
Potent GST Ketosteroid Isomerase Activity Relevant to Ecdysteroidogenesis in the Malaria Vector Anopheles gambiae


Abstract
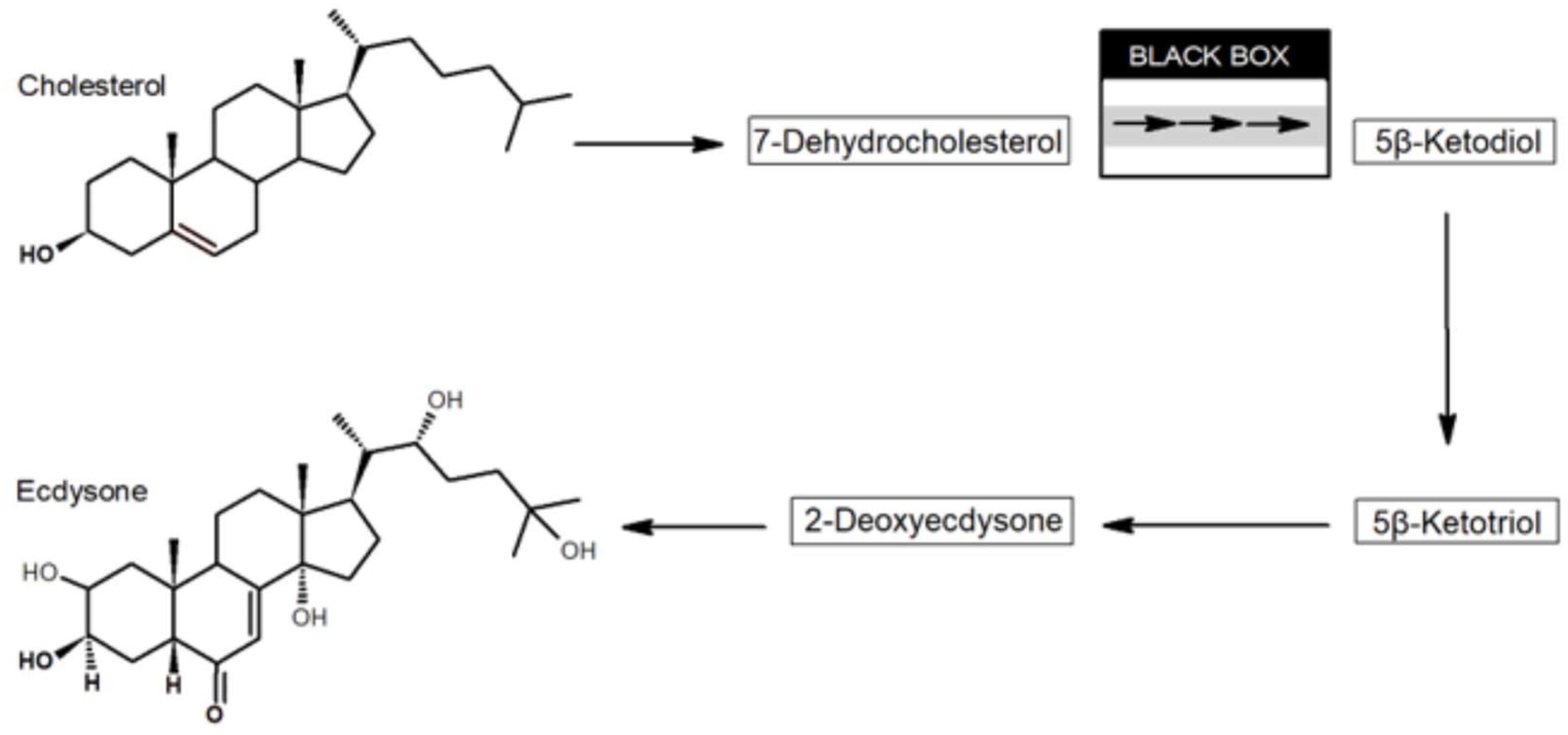
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Extracting Plasmid DNA from Filters
2.2.2. Transformation of Bacteria
2.2.3. Expression and Purification of Recombinant GSTs
2.2.4. Kinetic Experiments
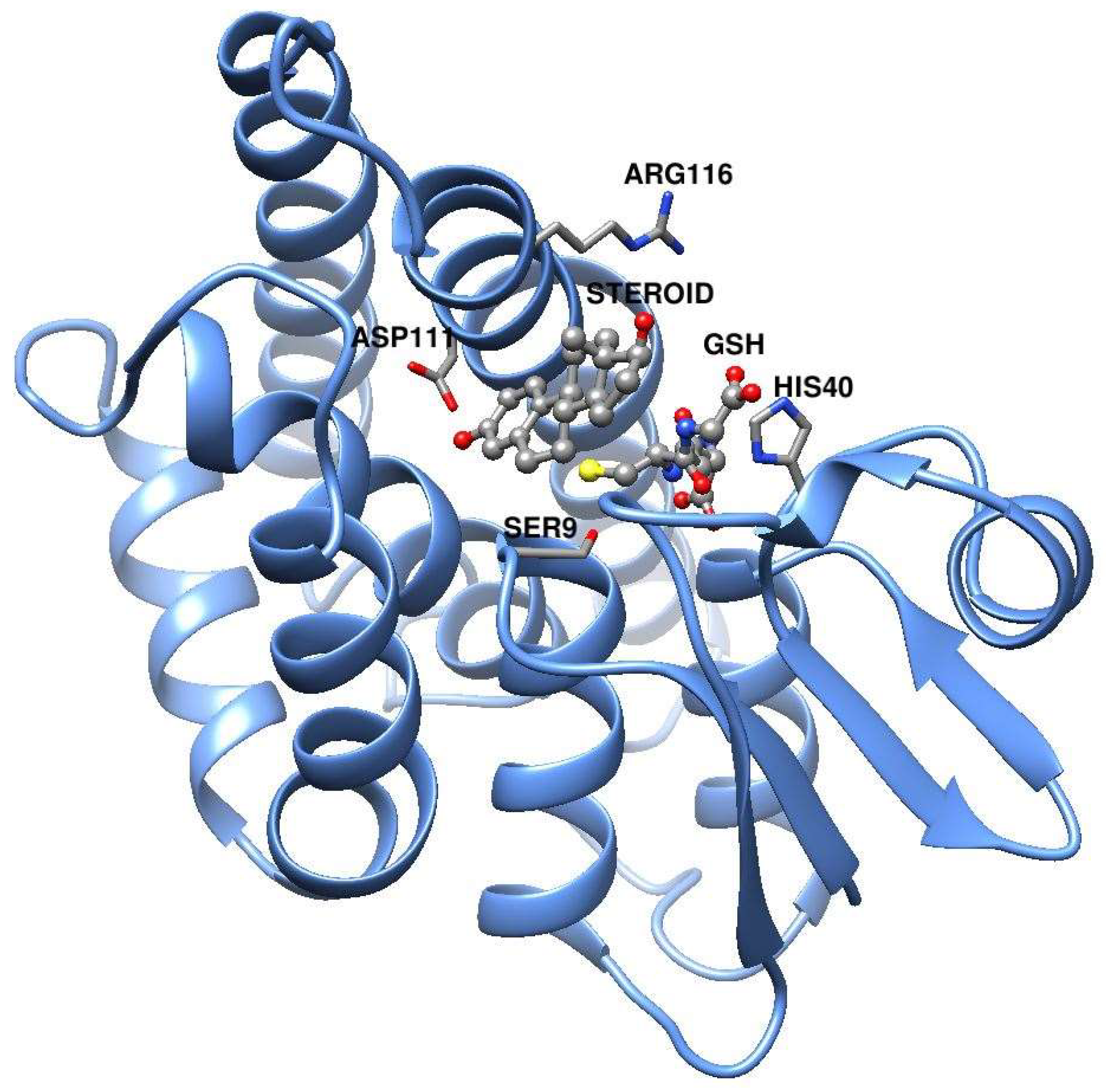
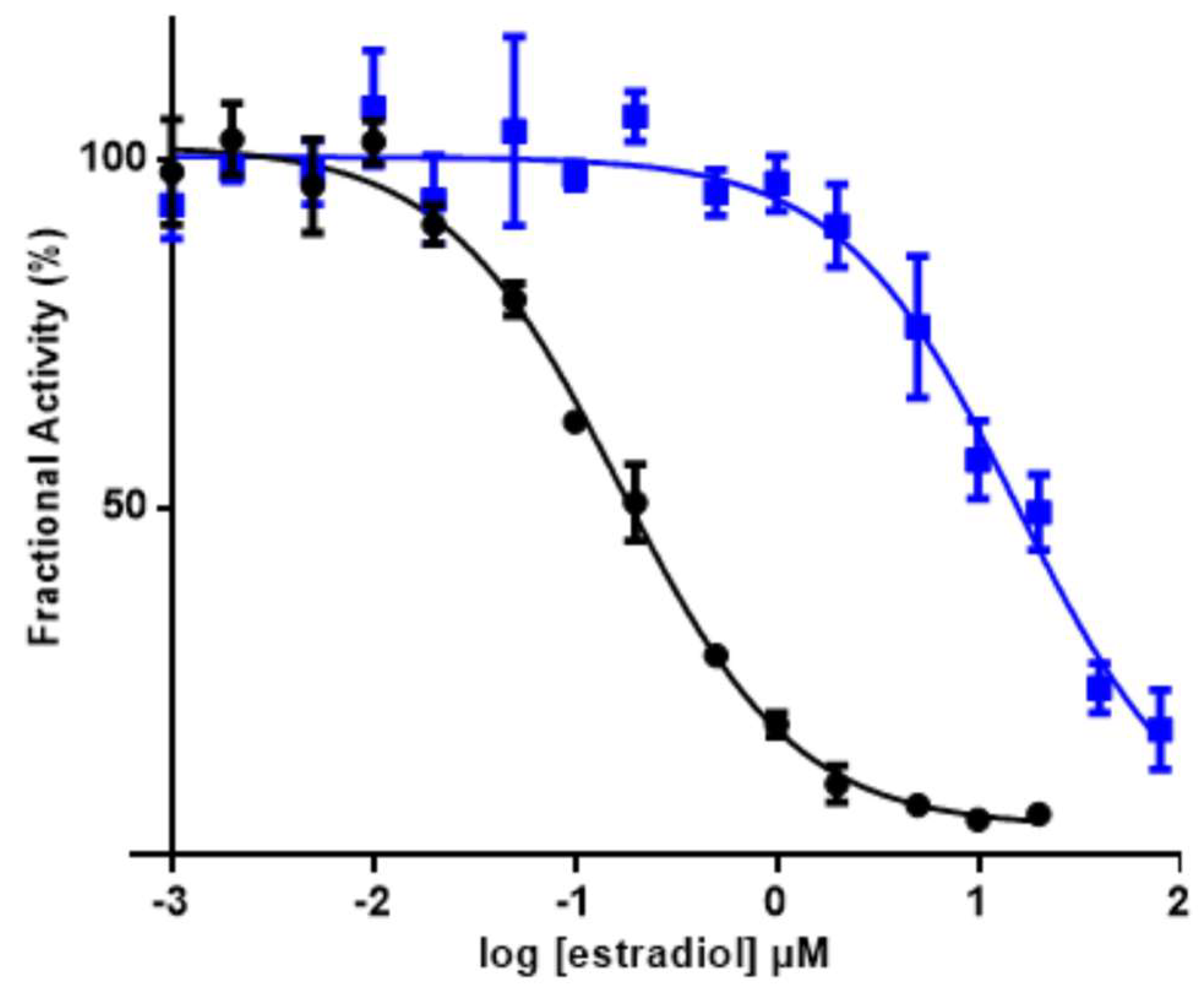
3. Results
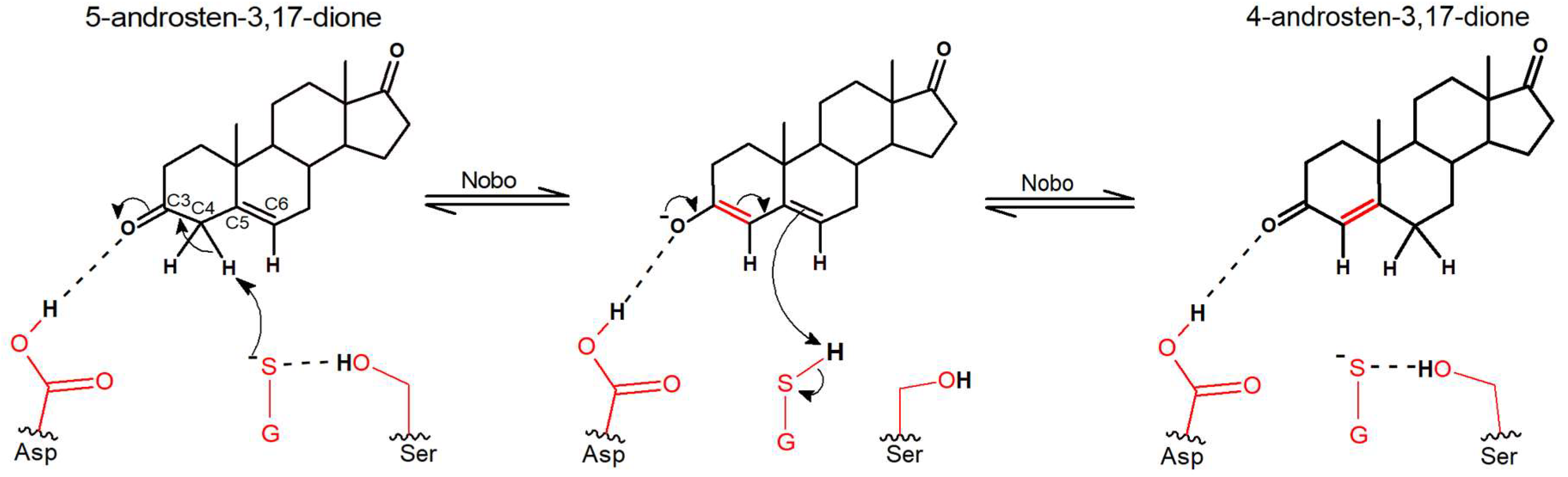
Nobo Is an Efficient Ketosteroid Isomerase
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Guidelines for Malaria; World Health Organization (WHO): Geneva, Switzerland, 2021.
- Ortelli, F.; Rossiter, L.C.; Vontas, J.; Ranson, H.; Hemingway, J. Heterologous expression of four glutathione transferase genes genetically linked to a major insecticide-resistance locus from the malaria vector Anopheles gambiae. Biochem. J. 2003, 373, 957–963. [Google Scholar] [CrossRef]
- Chanut-Delalande, H.; Hashimoto, Y.; Pelissier-Monier, A.; Spokony, R.; Dib, A.; Kondo, T.; Bohere, J.; Niimi, K.; Latapie, Y.; Inagaki, S.; et al. Pri peptides are mediators of ecdysone for the temporal control of development. Nat. Cell. Biol. 2014, 16, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Enya, S.; Ameku, T.; Igarashi, F.; Iga, M.; Kataoka, H.; Shinoda, T.; Niwa, R. A halloween gene noppera-bo encodes a glutathione s-transferase essential for ecdysteroid biosynthesis via regulating the behaviour of cholesterol in Drosophila. Sci. Rep. 2014, 4, 6586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enya, S.; Daimon, T.; Igarashi, F.; Kataoka, H.; Uchibori, M.; Sezutsu, H.; Shinoda, T.; Niwa, R. The silkworm glutathione s-transferase gene noppera-bo is required for ecdysteroid biosynthesis and larval development. Insect. Biochem. Mol. Biol. 2015, 61, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaba, K.; Ebihara, K.; Senda, M.; Yoshino, R.; Sakuma, C.; Koiwai, K.; Takaya, D.; Watanabe, C.; Watanabe, A.; Kawashima, Y.; et al. Molecular action of larvicidal flavonoids on ecdysteroidogenic glutathione s-transferase noppera-bo in Aedes aegypti. BMC Biol. 2022, 20, 43. [Google Scholar] [CrossRef]
- Mannervik, B.; Danielson, U.H. Glutathione transferases—Structure and catalytic activity. CRC Crit. Rev. Biochem. 1988, 23, 283–337. [Google Scholar] [CrossRef]
- Lindström, H.; Peer, S.M.; Ing, N.H.; Mannervik, B. Characterization of equine GST A3-3 as a steroid isomerase. J. Steroid. Biochem. Mol. Biol. 2018, 178, 117–126. [Google Scholar] [CrossRef]
- Skerlova, J.; Lindström, H.; Gonis, E.; Sjödin, B.; Neiers, F.; Stenmark, P.; Mannervik, B. Structure and steroid isomerase activity of Drosophila glutathione transferase E14 essential for ecdysteroid biosynthesis. FEBS Lett. 2020, 594, 1187–1195. [Google Scholar] [CrossRef] [Green Version]
- Mannervik, B.; Jemth, P. Measurement of glutathione transferases. In Current Protocols in Toxicology; Chapter 6; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Pettersson, P.L.; Mannervik, B. The role of glutathione in the isomerization of Δ5-androstene-3,17-dione catalyzed by human glutathione transferase A1-1. J. Biol. Chem. 2001, 276, 11698–11704. [Google Scholar] [CrossRef] [Green Version]
- Johansson, A.S.; Mannervik, B. Human glutathione transferase A3-3, a highly efficient catalyst of double-bond isomerization in the biosynthetic pathway of steroid hormones. J. Biol. Chem. 2001, 276, 33061–33065. [Google Scholar] [CrossRef] [Green Version]
- Raffalli-Mathieu, F.; Orre, C.; Stridsberg, M.; Hansson Edalat, M.; Mannervik, B. Targeting human glutathione transferase A3-3 attenuates progesterone production in human steroidogenic cells. Biochem. J. 2008, 414, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Koiwai, K.; Inaba, K.; Morohashi, K.; Enya, S.; Arai, R.; Kojima, H.; Okabe, T.; Fujikawa, Y.; Inoue, H.; Yoshino, R.; et al. An integrated approach to unravel a crucial structural property required for the function of the insect steroidogenic Halloween protein noppera-bo. J. Biol. Chem. 2020, 295, 7154–7167. [Google Scholar] [CrossRef] [Green Version]
- Kuliopulos, A.; Mildvan, A.S.; Shortle, D.; Talalay, P. Kinetic and ultraviolet spectroscopic studies of active-site mutants of Δ5-3-ketosteroid isomerase. Biochemistry 1989, 28, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Pollack, R.M. Enzymatic mechanisms for catalysis of enolization: Ketosteroid isomerase. Bioorg. Chem. 2004, 32, 341–353. [Google Scholar] [CrossRef]
- Radzicka, A.; Wolfenden, R. A proficient enzyme. Science 1995, 267, 90–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Connacher, R.P.; O’Connor, M.B. Control of the insect metamorphic transition by ecdysteroid production and secretion. Curr. Opin. Insect Sci. 2021, 43, 11–20. [Google Scholar] [CrossRef]
- Enya, S.; Yamamoto, C.; Mizuno, H.; Esaki, T.; Lin, H.K.; Iga, M.; Morohashi, K.; Hirano, Y.; Kataoka, H.; Masujima, T.; et al. Dual roles of glutathione in ecdysone biosynthesis and antioxidant function during larval development in Drosophila. Genetics 2017, 207, 1519–1532. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, H.J.; Babbitt, P.C. Glutathione transferases are structural and functional outliers in the thioredoxin fold. Biochemistry 2009, 48, 11108–11116. [Google Scholar] [CrossRef] [Green Version]
- Dourado, D.F.; Fernandes, P.A.; Mannervik, B.; Ramos, M.J. Isomerization of Δ5-androstene-3,17-dione into Δ4-androstene-3,17-dione catalyzed by human glutathione transferase A3-3: A computational study identifies a dual role for glutathione. J. Phys. Chem. A 2014, 118, 5790–5800. [Google Scholar] [CrossRef]
- Yabukarski, F.; Doukov, T.; Pinney, M.M.; Biel, J.T.; Fraser, J.S.; Herschlag, D. Ensemble-function relationships to dissect mechanisms of enzyme catalysis. Sci. Adv. 2022, 8, eabn7738. [Google Scholar] [CrossRef]
- Mannervik, B.; Ismail, A.; Lindström, H.; Sjödin, B.; Ing, N.H. Glutathione transferases as efficient ketosteroid isomerases. Front. Mol. Biosci. 2021, 8, 765970. [Google Scholar] [CrossRef]
- Ranson, H.; Lissenden, N. Insecticide resistance in African Anopheles mosquitoes: A worsening situation that needs urgent action to maintain malaria control. Trends. Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Josephy, P.D.; Mannervik, B. Molecular Toxicology, 2nd ed.; Oxford University Press: New York, NY, USA, 2006; pp. 333–364. [Google Scholar]
- Koirala, B.K.S.; Moural, T.; Zhu, F. Functional and structural diversity of insect glutathione S-transferases in xenobiotic adaptation. Int. J. Biol. Sci. 2022, 18, 5713–5723. [Google Scholar] [CrossRef] [PubMed]
- Ketterman, A.J.; Saisawang, C.; Wongsantichon, J. Insect glutathione transferases. Drug Metab. Rev. 2011, 43, 253–265. [Google Scholar] [CrossRef]
- Adolfi, A.; Poulton, B.; Anthousi, A.; Macilwee, S.; Ranson, H.; Lycett, G.J. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2019, 116, 25764–25772. [Google Scholar] [CrossRef] [Green Version]
- Ranson, H.; Hemingway, J. Mosquito glutathione transferases. Methods Enzymol. 2005, 401, 226–241. [Google Scholar]
- Bresell, A.; Weinander, R.; Lundqvist, G.; Raza, H.; Shimoji, M.; Sun, T.H.; Balk, L.; Wiklund, R.; Eriksson, J.; Jansson, C.; et al. Bioinformatic and enzymatic characterization of the MAPEG superfamily. FEBS J. 2005, 272, 1688–1703. [Google Scholar] [CrossRef]
- Warren, J.T.; O’Connor, M.B.; Gilbert, L.I. Studies on the black box: Incorporation of 3-oxo-7-dehydrocholesterol into ecdysteroids by Drosophila melanogaster and Manduca sexta. Insect Biochem. Mol. Biol. 2009, 39, 677–687. [Google Scholar] [CrossRef]
- Kamiyama, T.; Niwa, R. Transcriptional regulators of ecdysteroid biosynthetic enzymes and their roles in insect development. Front. Physiol. 2022, 13, 823418. [Google Scholar] [CrossRef]
- Swevers, L. An update on ecdysone signaling during insect oogenesis. Curr. Opin. Insect. Sci. 2019, 31, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef]
- Poulton, B.C.; Colman, F.; Anthousi, A.; Grigoraki, L.; Adolfi, A.; Lynd, A.; Lycett, G.J. Using the GAL4-UAS system for functional genetics in Anopheles gambiae. J. Vis. Exp. 2021, 170, e62131. [Google Scholar]
- Grisales, N.; Lees, R.S.; Maas, J.; Morgan, J.C.; Wangrawa, D.W.; Guelbeogo, W.M.; N’Fale, S.; Lindsay, S.W.; McCall, P.J.; Ranson, H. Pyriproxyfen-treated bed nets reduce reproductive fitness and longevity of pyrethroid-resistant Anopheles gambiae under laboratory and field conditions. Malar. J. 2021, 20, 273. [Google Scholar] [CrossRef] [PubMed]
- Baldini, F.; Gabrieli, P.; South, A.; Valim, C.; Mancini, F.; Catteruccia, F. The interaction between a sexually transferred steroid hormone and a female protein regulates oogenesis in the malaria mosquito Anopheles gambiae. PLoS Biol. 2013, 11, e1001695. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | WT | D111N | S9A | H40N | R116A |
|---|---|---|---|---|---|
| CDNB | 24.7 ± 3.4 | 26.2 ± 4.8 | 1.8 ± 0.2 | 14.0 ± 0.5 | 3.3 ± 0.1 |
| DCNB | 16.4 ± 1.9 | 14.7 ± 0.5 | 0.49 ± 0.03 | 5.0 ± 0.1 | 2.34 ± 0.02 |
| CuOOH | 5.0 ± 0.5 | 3.8 ± 0.4 | 0.05 ± 0.01 | 0.9 ± 0.1 | 0.16 ± 0.02 |
| Allyl-ITC | 3.3 ± 0.4 | 3.4 ± 0.5 | 0.2 ± 0.1 | 2.7 ± 0.3 | 2.0 ± 0.2 |
| Phenethyl-ITC | 0.4 ± 0.1 | 1.2 ± 0.3 | 0.04 ± 0.01 | 0.34 ± 0.02 | 0.1 ± 0.04 |
| 5-AD | 245 ± 8 | 0.8 ± 0.1 | 16.0 ± 2.0 | 69.3 ± 3.7 | 48.1 ± 1.2 |
| 5-PD | 49.7 ± 6.1 | 0.6 ± 0.2 | 5.3 ± 0.5 | 23.7 ± 4.1 | 14.7 ± 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musdal, Y.; Ismail, A.; Sjödin, B.; Mannervik, B. Potent GST Ketosteroid Isomerase Activity Relevant to Ecdysteroidogenesis in the Malaria Vector Anopheles gambiae. Biomolecules 2023, 13, 976. https://doi.org/10.3390/biom13060976
Musdal Y, Ismail A, Sjödin B, Mannervik B. Potent GST Ketosteroid Isomerase Activity Relevant to Ecdysteroidogenesis in the Malaria Vector Anopheles gambiae. Biomolecules. 2023; 13(6):976. https://doi.org/10.3390/biom13060976
Chicago/Turabian StyleMusdal, Yaman, Aram Ismail, Birgitta Sjödin, and Bengt Mannervik. 2023. "Potent GST Ketosteroid Isomerase Activity Relevant to Ecdysteroidogenesis in the Malaria Vector Anopheles gambiae" Biomolecules 13, no. 6: 976. https://doi.org/10.3390/biom13060976