
Pannexin-1 Modulates Inhibitory Transmission and Hippocampal Synaptic Plasticity
 ,
,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals and Reagents
2.3. Slice Preparation
2.4. Electrophysiology
2.5. Analysis
3. Results
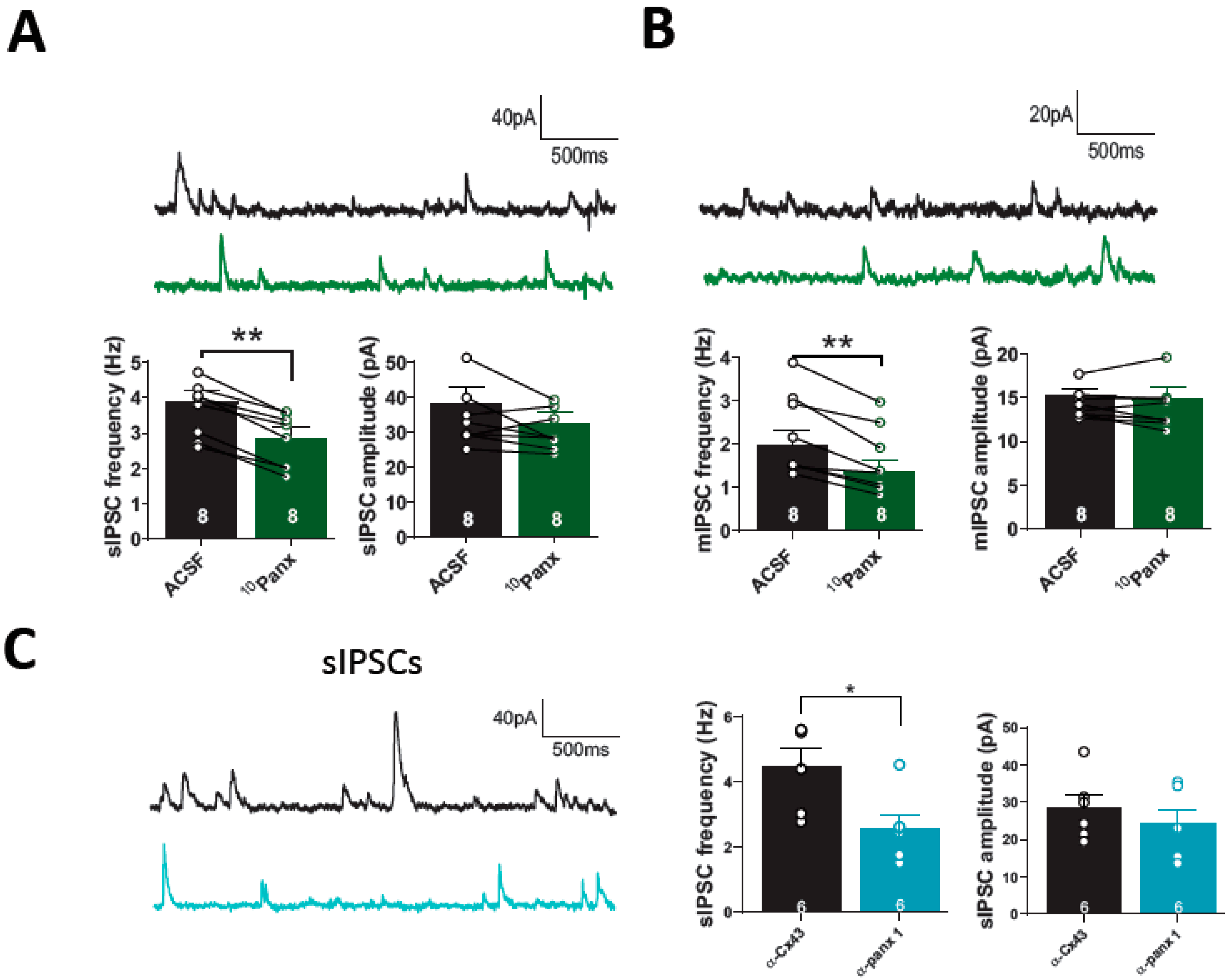
3.1. The Blockage of Panx1 Hemichannel Decrease Hippocampal GABAergic Efficacy
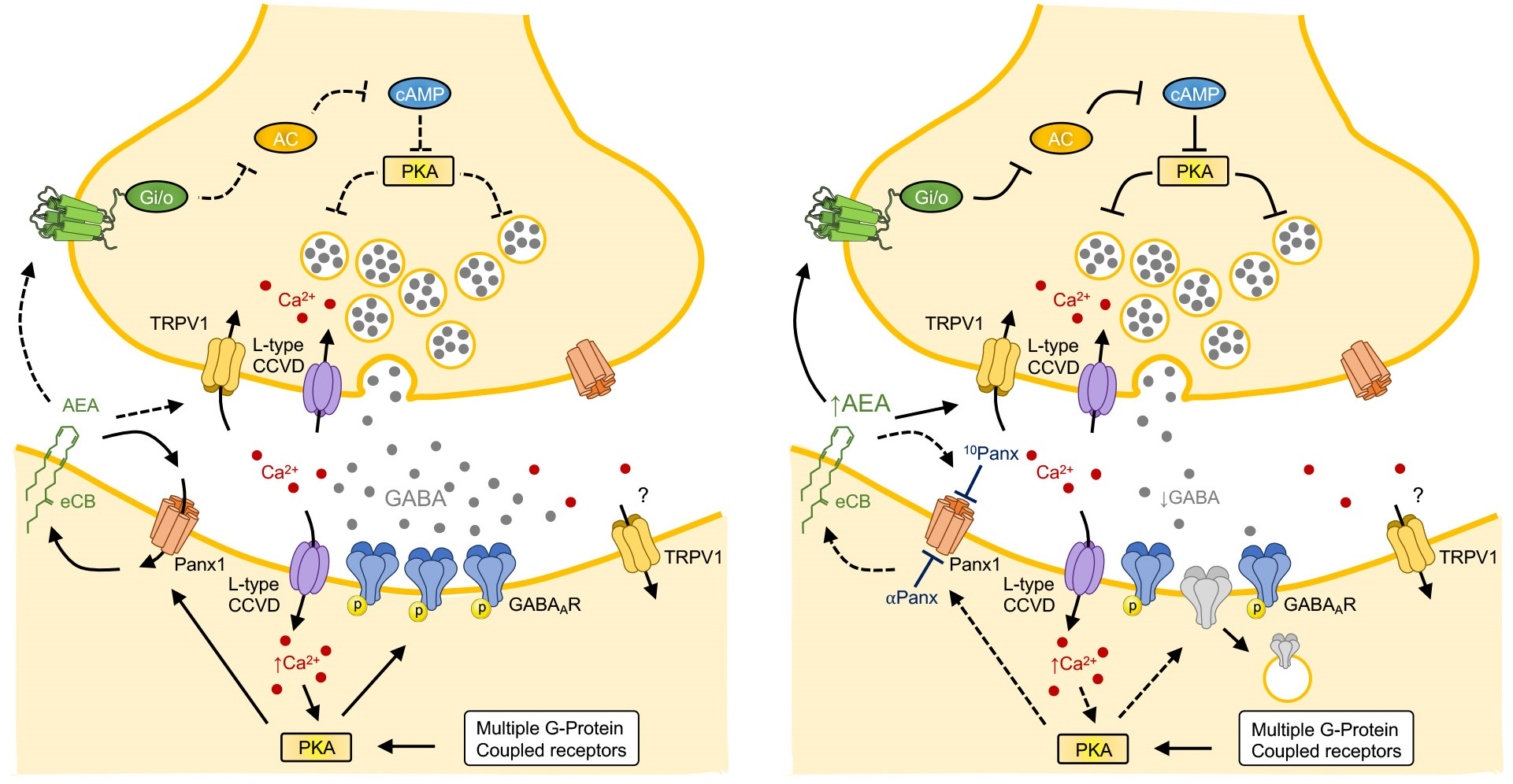
3.2. 10Panx1-Induced Depression of Inhibitory Transmission Requires CB1R and TRPV1 Activation and Presynaptic cAMP/PKA
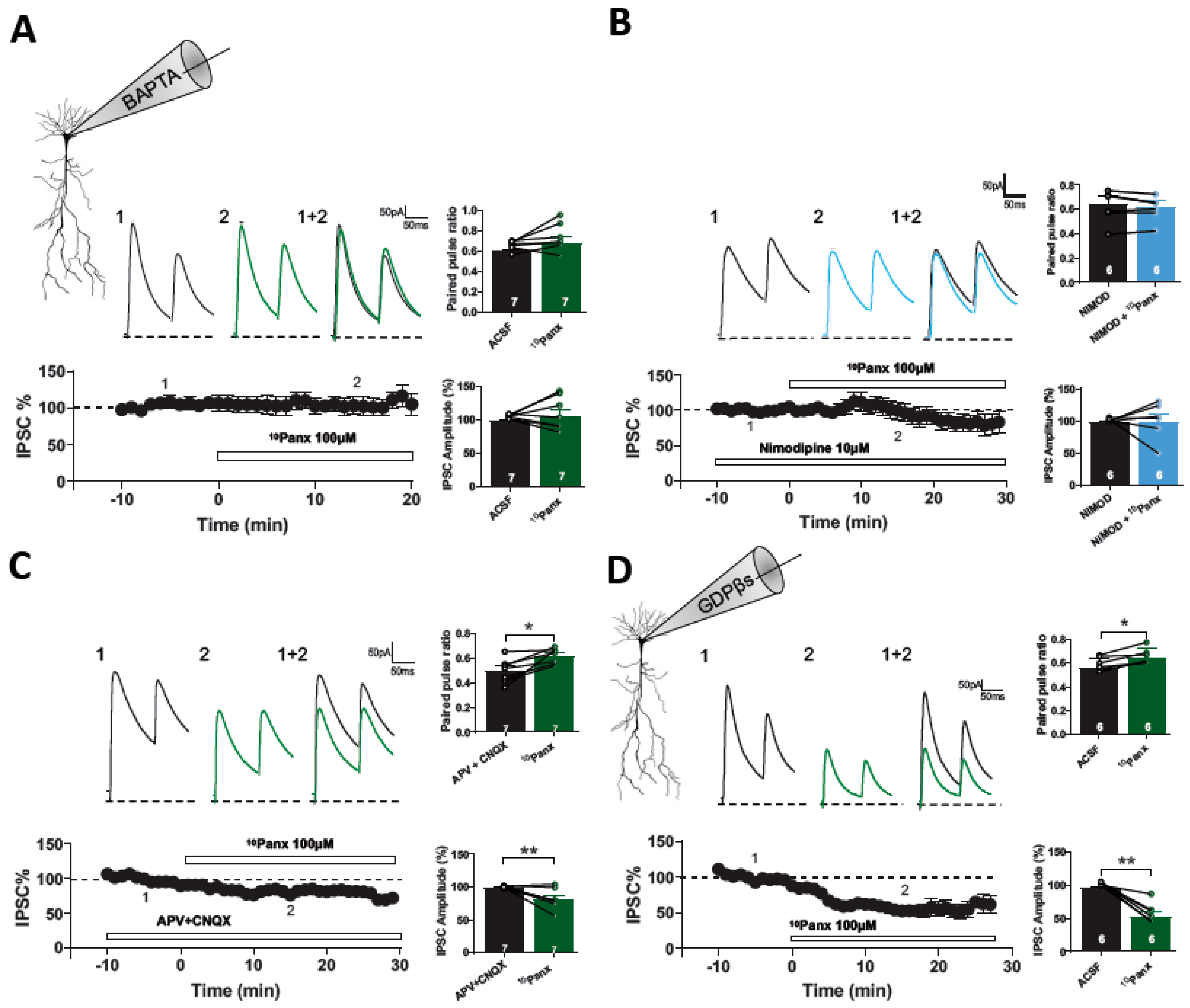
3.3. 10Panx1-Induced Inhibitory Depression Requires a Rise in Postsynaptic Intracellular Ca2+
3.4. Involvement of Panx1 Hemichannels in the Regulation of the E/I Ratio and the Threshold for Synaptic Plasticity in CA1 Pyramidal Neurons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, R.J.; Jackson, M.F.; Olah, M.E.; Rungta, R.L.; Hines, D.J.; Beasley, M.A.; MacDonald, J.F.; MacVicar, B.A. Activation of Pannexin-1 Hemichannels Augments Aberrant Bursting in the Hippocampus. Science 2008, 322, 1555–1559. [Google Scholar] [CrossRef] [PubMed]
- Bruzzone, R.; Hormuzdi, S.G.; Barbe, M.T.; Herb, A.; Monyer, H. Pannexins, a Family of Gap Junction Proteins Expressed in Brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13644–13649. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Prado, N.; Soto, P.A.; López, X.; Choi, E.J.; Marquez-Miranda, V.; Rojas, M.; Duarte, Y.; Lee, J.; González-Nilo, F.D.; Sáez, J.C. Endogenous Pannexin1 Channels Form Functional Intercellular Cell–Cell Channels with Characteristic Voltage-Dependent Properties. Proc. Natl. Acad. Sci. USA 2022, 119, e2202104119. [Google Scholar] [CrossRef] [PubMed]
- Ardiles, A.O.; Flores-Muñoz, C.; Toro-Ayala, G.; Cárdenas, A.M.; Palacios, A.G.; Muñoz, P.; Fuenzalida, M.; Saez, J.C.; Martinez, A.D. Pannexin 1 Regulates Bidirectional Hippocampal Synaptic Plasticity in Adult Mice. Front. Cell. Neurosci. 2014, 8, 326. [Google Scholar] [CrossRef]
- Prochnow, N.; Abdulazim, A.; Kurtenbach, S.; Wildförster, V.; Dvoriantchikova, G.; Hanske, J.; Petrasch-Parwez, E.; Shestopalov, V.I.; Dermietzel, R.; Manahan-Vaughan, D.; et al. Pannexin1 Stabilizes Synaptic Plasticity and Is Needed for Learning. PLoS ONE 2012, 7, e51767. [Google Scholar] [CrossRef]
- Ray, A.; Zoidl, G.; Weickert, S.; Wahle, P.; Dermietzel, R. Site-Specific and Developmental Expression of Pannexin1 in the Mouse Nervous System. Eur. J. Neurosci. 2005, 21, 3277–3290. [Google Scholar] [CrossRef]
- Vogt, A.; Hormuzdi, S.G.; Monyer, H. Pannexin1 and Pannexin2 Expression in the Developing and Mature Rat Brain. Mol. Brain Res. 2005, 141, 113–120. [Google Scholar] [CrossRef]
- Zoidl, G.; Petrasch-Parwez, E.; Ray, A.; Meier, C.; Bunse, S.; Habbes, H.-W.; Dahl, G.; Dermietzel, R. Localization of the Pannexin1 Protein at Postsynaptic Sites in the Cerebral Cortex and Hippocampus. Neuroscience 2007, 146, 9–16. [Google Scholar] [CrossRef]
- Flores-Muñoz, C.; García-Rojas, F.; Pérez, M.A.; Santander, O.; Mery, E.; Ordenes, S.; Illanes-González, J.; López-Espíndola, D.; González-Jamett, A.M.; Fuenzalida, M.; et al. The Long-Term Pannexin 1 Ablation Produces Structural and Functional Modifications in Hippocampal Neurons. Cells 2022, 11, 3646. [Google Scholar] [CrossRef]
- Gajardo, I.; Salazar, C.S.; Lopez-Espíndola, D.; Estay, C.; Flores-Muñoz, C.; Elgueta, C.; Gonzalez-Jamett, A.M.; Martínez, A.D.; Muñoz, P.; Ardiles, Á.O. Lack of Pannexin 1 Alters Synaptic GluN2 Subunit Composition and Spatial Reversal Learning in Mice. Front. Mol. Neurosci. 2018, 11, 114. [Google Scholar] [CrossRef]
- Bialecki, J.; Werner, A.; Weilinger, N.L.; Tucker, C.M.; Vecchiarelli, H.A.; Egaña, J.; Mendizabal-Zubiaga, J.; Grandes, P.; Hill, M.N.; Thompson, R.J. Suppression of Presynaptic Glutamate Release by Postsynaptic Metabotropic NMDA Receptor Signalling to Pannexin-1. J. Neurosci. 2020, 40, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Sohal, V.S.; Rubenstein, J.L.R. Excitation-Inhibition Balance as a Framework for Investigating Mechanisms in Neuropsychiatric Disorders. Mol. Psychiatry 2019, 24, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Vogels, T.P.; Abbott, L.F. Gating Multiple Signals through Detailed Balance of Excitation and Inhibition in Spiking Networks. Nat. Neurosci. 2009, 12, 483–491. [Google Scholar] [CrossRef]
- Isaacson, J.S.; Scanziani, M. How Inhibition Shapes Cortical Activity. Neuron 2011, 72, 231–243. [Google Scholar] [CrossRef]
- Ferguson, B.R.; Gao, W.-J. Thalamic Control of Cognition and Social Behavior Via Regulation of Gamma-Aminobutyric Acidergic Signaling and Excitation/Inhibition Balance in the Medial Prefrontal Cortex. Biol. Psychiatry 2018, 83, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, D.M. Interneuron Networks in the Hippocampus. Curr. Opin. Neurobiol. 2011, 21, 709–716. [Google Scholar] [CrossRef]
- Mele, M.; Leal, G.; Duarte, C.B. Role of GABAAR Trafficking in the Plasticity of Inhibitory Synapses. J. Neurochem. 2016, 139, 997–1018. [Google Scholar] [CrossRef]
- Sprekeler, H. Functional Consequences of Inhibitory Plasticity: Homeostasis, the Excitation-Inhibition Balance and Beyond. Curr. Opin. Neurobiol. 2017, 43, 198–203. [Google Scholar] [CrossRef]
- Chiu, C.Q.; Barberis, A.; Higley, M.J. Preserving the Balance: Diverse Forms of Long-Term GABAergic Synaptic Plasticity. Nat. Rev. Neurosci. 2019, 20, 272–281. [Google Scholar] [CrossRef]
- Barberis, A. Postsynaptic Plasticity of GABAergic Synapses. Neuropharmacology 2020, 169, 107643. [Google Scholar] [CrossRef]
- Anselmi, F.; Hernandez, V.H.; Crispino, G.; Seydel, A.; Ortolano, S.; Roper, S.D.; Kessaris, N.; Richardson, W.; Rickheit, G.; Filippov, M.A.; et al. ATP Release through Connexin Hemichannels and Gap Junction Transfer of Second Messengers Propagate Ca2+ Signals across the Inner Ear. Proc. Natl. Acad. Sci. USA 2008, 105, 18770–18775. [Google Scholar] [CrossRef] [PubMed]
- Weilinger, N.L.; Tang, P.L.; Thompson, R.J. Anoxia-Induced NMDA Receptor Activation Opens Pannexin Channels via Src Family Kinases. J. Neurosci. 2012, 32, 12579–12588. [Google Scholar] [CrossRef]
- Weilinger, N.L.; Lohman, A.W.; Rakai, B.D.; Ma, E.M.M.; Bialecki, J.; Maslieieva, V.; Rilea, T.; Bandet, M.V.; Ikuta, N.T.; Scott, L.; et al. Metabotropic NMDA Receptor Signaling Couples Src Family Kinases to Pannexin-1 during Excitotoxicity. Nat. Neurosci. 2016, 19, 432–442. [Google Scholar] [CrossRef]
- Hsia, A.Y.; Malenka, R.C.; Nicoll, R.A. Development of Excitatory Circuitry in the Hippocampus. J. Neurophysiol. 1998, 79, 2013–2024. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, D.M.; Moreau, A.W.; Bakiri, Y.; Nicholson, E. Plasticity of Inhibition. Neuron 2012, 75, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Chevaleyre, V.; Takahashi, K.; Castillo, P. Endocannabinoid-Mediated Synaptic Plasticity in the CNS. Annu. Rev. Neurosci 2006, 29, 37–76. [Google Scholar] [CrossRef]
- Ahumada, J.; de Sevilla, D.F.; Couve, A.; Buño, W.; Fuenzalida, M. Long-Term Depression of Inhibitory Synaptic Transmission Induced by Spike-Timing Dependent Plasticity Requires Coactivation of Endocannabinoid and Muscarinic Receptors: CB1 and M AC H RECEPTORS REGULATE THE STDP- I LTD. Hippocampus 2013, 23, 1439–1452. [Google Scholar] [CrossRef]
- Chevaleyre, V.; Castillo, P.E. Heterosynaptic LTD of Hippocampal GABAergic Synapses: A Novel Role of Endocannabinoids in Regulating Excitability. Neuron 2003, 38, 461–472. [Google Scholar] [CrossRef]
- Shestopalov, V.I.; Slepak, V.Z. Molecular Pathways of Pannexin1-Mediated Neurotoxicity. Front. Physiol. 2014, 5, 23. [Google Scholar] [CrossRef]
- Ross, R.A. Anandamide and Vanilloid TRPV1 Receptors. Br. J. Pharmacol. 2003, 140, 790–801. [Google Scholar] [CrossRef]
- Chevaleyre, V.; Heifets, B.D.; Kaeser, P.S.; Südhof, T.C.; Castillo, P.E. Endocannabinoid-Mediated Long-Term Plasticity Requires CAMP/PKA Signaling and RIM1α. Neuron 2007, 54, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Lonart, G.; Schoch, S.; Kaeser, P.S.; Larkin, C.J.; Südhof, T.C.; Linden, D.J. Phosphorylation of RIM1alpha by PKA Triggers Presynaptic Long-Term Potentiation at Cerebellar Parallel Fiber Synapses. Cell 2003, 115, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Shosaku, T.; Maejima, T.; Kano, M. Endogenous Cannabinoids Mediate Retrograde Signals from Depolarized Postsynaptic Neurons to Presynaptic Terminals. Neuron 2001, 29, 729–738. [Google Scholar] [CrossRef]
- Wilson, R.I.; Nicoll, R.A. Endogenous Cannabinoids Mediate Retrograde Signalling at Hippocampal Synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Thibault, O.; Landfield, P.W. Increase in Single L-Type Calcium Channels in Hippocampal Neurons during Aging. Science 1996, 272, 1017–1020. [Google Scholar] [CrossRef]
- Kapur, A.; Yeckel, M.F.; Gray, R.; Johnston, D. L-Type Calcium Channels Are Required for One Form of Hippocampal Mossy Fiber LTP. J. Neurophysiol. 1998, 79, 2181–2190. [Google Scholar] [CrossRef] [PubMed]
- Froemke, R.C. Plasticity of Cortical Excitatory-Inhibitory Balance. Annu. Rev. Neurosci. 2015, 38, 195–219. [Google Scholar] [CrossRef]
- Rubin, R.; Abbott, L.F.; Sompolinsky, H. Balanced Excitation and Inhibition Are Required for High-Capacity, Noise-Robust Neuronal Selectivity. Proc. Natl. Acad. Sci. USA 2017, 114, E9366–E9375. [Google Scholar] [CrossRef]
- Nanou, E.; Lee, A.; Catterall, W.A. Control of Excitation/Inhibition Balance in a Hippocampal Circuit by Calcium Sensor Protein Regulation of Presynaptic Calcium Channels. J. Neurosci. 2018, 38, 4430–4440. [Google Scholar] [CrossRef]
- Meredith, R.M.; Floyer-Lea, A.M.; Paulsen, O. Maturation of Long-Term Potentiation Induction Rules in Rodent Hippocampus: Role of GABAergic Inhibition. J. Neurosci. 2003, 23, 11142–11146. [Google Scholar] [CrossRef]
- Paulsen, O.; Moser, E. A Model of Hippocampal Memory Encoding and Retrieval: GABAergic Control of Synaptic Plasticity. Trends Neurosci. 1998, 21, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Arias, J.C.; Liu, M.; Choi, C.S.W.; Ebert, S.N.; Brown, C.E.; Swayne, L.A. Pannexin 1 Regulates Network Ensembles and Dendritic Spine Development in Cortical Neurons. eNeuro 2019, 6, ENEURO.0503-18.2019. [Google Scholar] [CrossRef] [PubMed]
- Vasile, F.; Dossi, E.; Moulard, J.; Ezan, P.; Lecoin, L.; Cohen-Salmon, M.; Mailly, P.; Le Bert, M.; Couillin, I.; Bemelmans, A.; et al. Pannexin 1 Activity in Astroglia Sets Hippocampal Neuronal Network Patterns. PLoS Biol. 2022, 20, e3001891. [Google Scholar] [CrossRef] [PubMed]
- Pouille, F.; Scanziani, M. Enforcement of Temporal Fidelity in Pyramidal Cells by Somatic Feed-Forward Inhibition. Science 2001, 293, 1159–1163. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, R.; Lee, S.; Rudy, B. GABAergic Interneurons in the Neocortex: From Cellular Properties to Circuits. Neuron 2016, 91, 260–292. [Google Scholar] [CrossRef]
- Devane, W.; Hanus, L.; Breuer, A.; Pertwee, R.; Stevenson, L.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Stella, N.; Schweitzer, P.; Piomelli, D. A Second Endogenous Cannabinoid That Modulates Long-Term Potentiation. Nature 1997, 388, 773–778. [Google Scholar] [CrossRef]
- Irving, A.J.; Coutts, A.A.; Harvey, J.; Rae, M.G.; Mackie, K.; Bewick, G.S.; Pertwee, R.G. Functional Expression of Cell Surface Cannabinoid CB1 Receptors on Presynaptic Inhibitory Terminals in Cultured Rat Hippocampal Neurons. Neuroscience 2000, 98, 253–262. [Google Scholar] [CrossRef]
- Katona, I.; Sperlágh, B.; Sík, A.; Käfalvi, A.; Vizi, E.S.; Mackie, K.; Freund, T.F. Presynaptically Located CB1 Cannabinoid Receptors Regulate GABA Release from Axon Terminals of Specific Hippocampal Interneurons. J. Neurosci. 1999, 19, 4544–4558. [Google Scholar] [CrossRef]
- Fykse, E.; Li, C.; Sudhof, T. Phosphorylation of Rabphilin-3A by Ca2+/Calmodulin- and CAMP-Dependent Protein Kinases in Vitro. J. Neurosci. 1995, 15, 2385–2395. [Google Scholar] [CrossRef]
- Hirling, H.; Scheller, R. Phosphorylation of Synaptic Vesicle Proteins: Modulation of the ASNAP Interaction with the Core Complex. Proc. Natl. Acad. Sci. USA 1996, 93, 11945–11949. [Google Scholar] [CrossRef]
- Nguyen, P.; Woo, N. Regulation of Hippocampal Synaptic Plasticity by Cyclic AMP-Dependent Protein Kinases. Prog. Neurobiol. 2003, 71, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-C.; Mackie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef]
- Shonesy, B.; Stephenson, J.; Marks, C.; Colbran, R. Cyclic AMP-dependent protein kinase and D1 dopamine receptors regulate diacylglycerol lipase-α and synaptic 2-arachidonoyl glycerol signaling. J. Neurochem. 2020, 153, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.E. Presynaptic LTP and LTD of Excitatory and Inhibitory Synapses. Cold Spring Harb. Perspect. Biol. 2012, 4, a005728. [Google Scholar] [CrossRef]
- Kittler, J.T.; Moss, S.J. Modulation of GABAA Receptor Activity by Phosphorylation and Receptor Trafficking: Implications for the Efficacy of Synaptic Inhibition. Curr. Opin. Neurobiol. 2003, 13, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Nusser, Z.; Hájos, N.; Somogyi, P.; Mody, I. Increased Number of Synaptic GABAA Receptors Underlies Potentiation at Hippocampal Inhibitory Synapses. Nature 1998, 395, 172–177. [Google Scholar] [CrossRef]
- McDonald, B.J.; Amato, A.; Connolly, C.N.; Benke, D.; Moss, S.J.; Smart, T.G. Adjacent Phosphorylation Sites on GABAA Receptor β Subunits Determine Regulation by CAMP-Dependent Protein Kinase. Nat. Neurosci. 1998, 1, 23–28. [Google Scholar] [CrossRef]
- López, X.; Escamilla, R.; Fernández, P.; Duarte, Y.; González-Nilo, F.; Palacios-Prado, N.; Martinez, A.D.; Sáez, J.C. Stretch-Induced Activation of Pannexin 1 Channels Can Be Prevented by PKA-Dependent Phosphorylation. Int. J. Mol. Sci. 2020, 21, 9180. [Google Scholar] [CrossRef]
- Ma, W.; Compan, V.; Zheng, W.; Martin, E.; North, R.A.; Verkhratsky, A.; Surprenant, A. Pannexin 1 Forms an Anion-Selective Channel. Pflüg. Arch.-Eur. J. Physiol. 2012, 463, 585–592. [Google Scholar] [CrossRef]
- Kalsi, K.; González-Alonso, J. Temperature-dependent release of ATP from human erythrocytes: Mechanism for the control of local tissue perfusion. Exp. Physiol. 2012, 97, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, L. Xenopus connexin38 forms hemi-gap-junctional channels in the nonjunctional plasma membrane of Xenopus oocytes. Biophys J. 1996, 71, 742–748. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rojas, F.; Flores-Muñoz, C.; Santander, O.; Solis, P.; Martínez, A.D.; Ardiles, Á.O.; Fuenzalida, M. Pannexin-1 Modulates Inhibitory Transmission and Hippocampal Synaptic Plasticity. Biomolecules 2023, 13, 887. https://doi.org/10.3390/biom13060887
García-Rojas F, Flores-Muñoz C, Santander O, Solis P, Martínez AD, Ardiles ÁO, Fuenzalida M. Pannexin-1 Modulates Inhibitory Transmission and Hippocampal Synaptic Plasticity. Biomolecules. 2023; 13(6):887. https://doi.org/10.3390/biom13060887
Chicago/Turabian StyleGarcía-Rojas, Francisca, Carolina Flores-Muñoz, Odra Santander, Pamela Solis, Agustín D. Martínez, Álvaro O. Ardiles, and Marco Fuenzalida. 2023. "Pannexin-1 Modulates Inhibitory Transmission and Hippocampal Synaptic Plasticity" Biomolecules 13, no. 6: 887. https://doi.org/10.3390/biom13060887