
Phytochemical Interactions with Calmodulin and Critical Calmodulin Binding Proteins Involved in Amyloidogenesis in Alzheimer’s Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
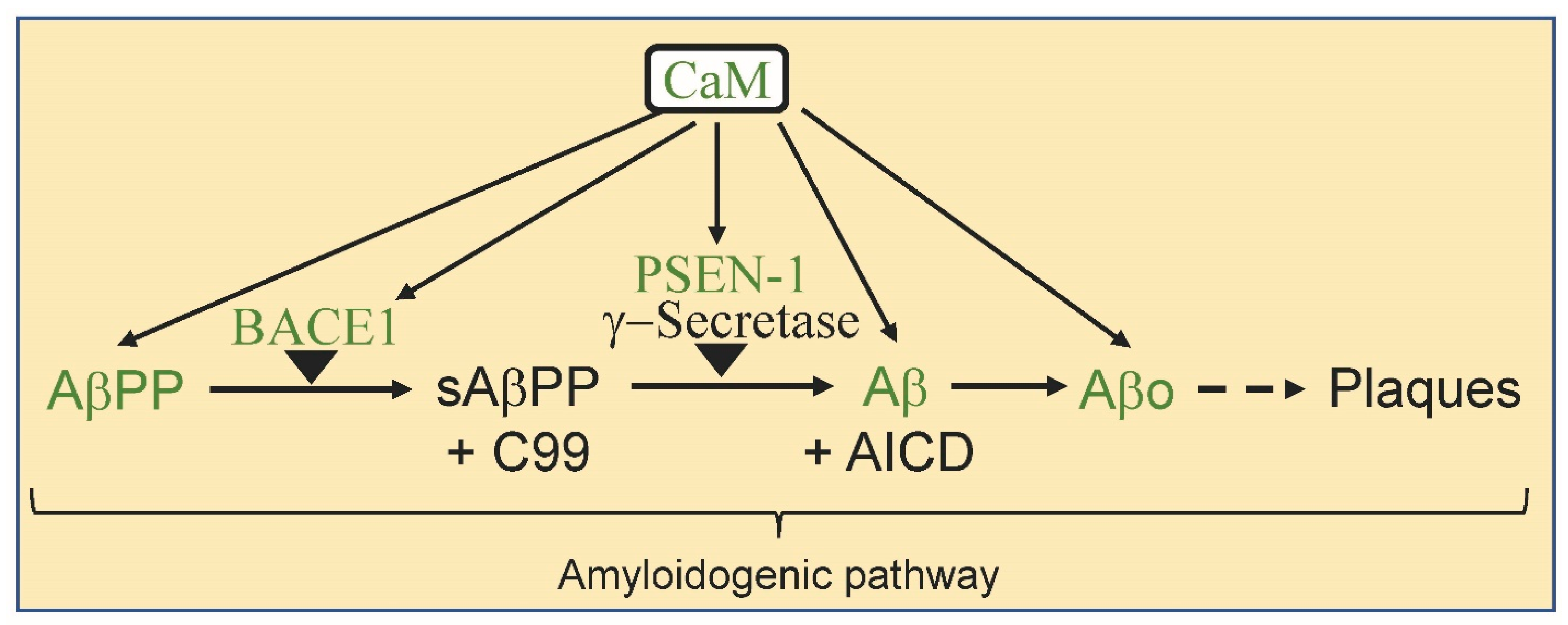
2. Calmodulin and Alzheimer’s Disease
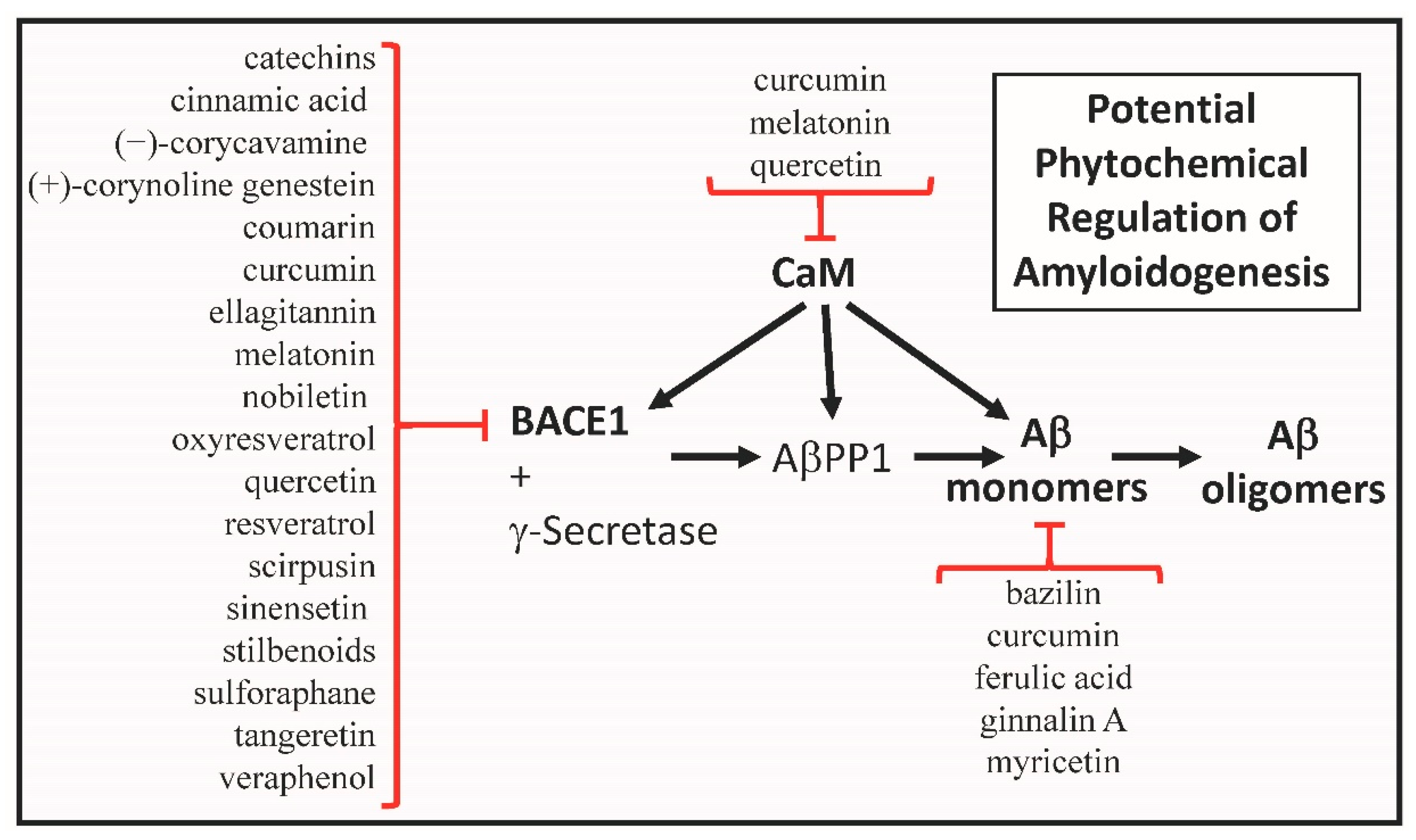
3. Calmodulin as a Phytochemical Target
4. Amyloid Beta Monomers as a Phytochemical Targets
5. Bace1 as a Phytochemical Target
6. Phytochemicals and Calmodulin Gene Expression
7. Discussion
8. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Aβ | amyloid beta |
| Aβo | amyloid beta oligomers |
| AβPP | amyloid-β precursor protein |
| AchR | acetylcholine receptor |
| AD | Alzheimer’s disease |
| AICD | APP intracellular domain |
| BACE1 | beta-secretase 1 |
| C99 | 99-residue C-terminal fragment |
| CaM | calmodulin |
| CaMBD | calmodulin binding domain |
| CaMBP | calmodulin binding protein |
| CaMKII | calcium/CaM-dependent kinase II |
| cdk5 | cyclin dependent kinase 5 |
| HBC | 4-[3,5-Bis-[2-(4-hydroxy-3-methoxy-phenyl)-ethyl]-4,5-dihydro-pyrazol-1-yl]-benzoic acid) |
| NMR | nuclear magnetic resonance |
| PMFs | polymehtoxyflavones |
| PP2B | calcineurin |
| pTau | phosphorylated Tau |
| W7 | N-6-aminohexyl-5-chloro-1-naphthalenesufonamide |
References
- Gauthier, S.; Webster, C.; Servaes, S.; Morais, J.A.; Rosa-Neto, P. World Alzheimer Report 2022: Life after Diagnosis: Navigating Treatment, Care and Support; Alzheimer’s Disease International: London, UK, 2022. [Google Scholar]
- Khachaturian, Z.S. Towards theories of brain aging. In Handbook of Studies on Psychiatry and Old Age; Kay, D.S., Burrows, G.W., Eds.; Elsevier Science Publishers, B.V.: Amsterdam, The Netherlands, 1984; pp. 7–30. [Google Scholar]
- Hampel, H.; Caraci, F.; Cuello, A.C.; Caruso, G.; Nisticò, R.; Corbo, M.; Baldacci1, F.; Toschi1, N.; Garaci, F.; Chiesa, P.A.; et al. A path toward precision medicine for neuroinflammatory mechanisms in Alzheimer’s disease. Front. Immunol. 2020, 11, 456. [Google Scholar] [CrossRef]
- Karen, E.; De Strooper, B. The amyloid hypothesis in Alzheimer disease: New insights from new therapeutics. Nature 2022, 21, 306–318. [Google Scholar] [CrossRef]
- O’Day, D.H.; Myre, M.A. Calmodulin-binding domains in Alzheimer’s disease proteins: Extending the calcium hypothesis. Biochem. Biophys. Res. Commun. 2004, 230, 1051–1054. [Google Scholar] [CrossRef]
- O’Day, D.H.; Eshak, K.; Myre, M.A. Calmodulin Binding Proteins and Alzheimer’s Disease: A Review. J. Alz. Dis. 2015, 46, 553–569. [Google Scholar] [CrossRef] [Green Version]
- O’Day, D.H. Calmodulin binding proteins and Alzheimer’s disease: Biomarkers, regulatory enzymes and receptors that are regulated by calmodulin. Int. J. Mol. Sci. 2020, 21, 7344. [Google Scholar] [CrossRef]
- French, J.A.; Koepp, M.; Naegelin, Y.; Vigevano, F.; Rho, J.M.; Rosenberg, E.; Devinsky, O.; Olofsson, P.S.; Dichter, M.A. Clinical studies and anti-inflammatory mechanisms of treatments. Epilepsia 2017, 58, 69–82. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.-G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An overview of oxidative stress, neuroinflammation, and neurodegenerative diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Sahlin, C.; Pettersson, F.E.; Nilsson, L.N.; Lannfelt, L.; Johansson, A.S. Docosahexaenoic acid stimulates non-amyloidogenic APP processing resulting in reduced Abeta levels in cellular models of Alzheimer’s disease. Eur. J. Neurosci. 2007, 26, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Canobbio, I.; Catricalà, S.; Balduini, C.; Torti, M. Calmodulin regulates the non-amyloidogenic metabolism of amyloid precursor protein in platelets. Biochim. Biophys. Acta Bioenergy 2011, 1813, 500–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Shi, J.; Zhang, P.; Zhang, Y.; Xu, J.; Zhao, L.; Zhang, R.; Wang, H.; Chen, H. Immunotherapy for Alzheimer’s disease: Targeting β-amyloid and beyond. Trans. Neurodegen. 2022, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Farías, G.A.; Guzmán-Martínez, L.; Delgado, C.; Maccioni, R.B. Nutraceuticals: A novel concept in the prevention and treatment of Alzheimer’s disease and related disorders. J. Alz. Dis. 2014, 42, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Jantan, I.; Ahmad, W.; Bukhari, S.N.A. Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Front. Plant Sci. 2015, 6, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlAli, M.; Alqubaisy, M.; Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Molouki, A.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. Nutraceuticals: Transformation of conventional foods into health promoters/disease preventers and safety considerations. Molecules 2021, 26, 2540. [Google Scholar] [CrossRef]
- Azlan, U.K.; Annuar, N.A.K.; Mediani, A.; Aizat, W.M.; Damanhuri, H.A.; Tong, X.; Yanagisawa, D.; Tooyama, I.; Ngah, W.Z.W.; Jantan, I.; et al. An insight into the neuroprotective and anti-neuroinflammatory effects and mechanisms of Moringa oleifera. Front. Pharmacol. 2022, 13, 1035220. [Google Scholar] [CrossRef]
- Maccioni, R.B.; Calfío, C.; González, A.; Lüttges, V. Novel Nutraceutical Compounds in Alzheimer Prevention. Biomolecules 2022, 12, 249. [Google Scholar] [CrossRef]
- Santini, A.; Novellino, E. To Nutraceuticals and back: Rethinking a concept. Foods 2017, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, A.R.; Friedberg, F. Sequence motifs for calmodulin recognition. FASEB J. 1997, 11, 331–340. [Google Scholar] [CrossRef]
- Tidow, H.; Nissen, P. Structural diversity of calmodulin binding to its target sites. FEBS J. 2013, 280, 5551–5565. [Google Scholar] [CrossRef]
- Grant, B.M.M.; Enomoto, M.; Ikura, M.; Marshall, C.B. A non-canonical calmodulin target motif comprising a polybasic region and lapidated terminal residue regulates localization. Int. J. Mol. Sci. 2020, 21, 2751. [Google Scholar] [CrossRef] [Green Version]
- Léger, C.; Pitard, I.; Sadi, M.; Carvalho, N.; Brier, S.; Mechaly, A.; Raoux-Barbot, D.; Davi, M.; Hoos, S.; Weber, P.; et al. Patrice Vachette Dynamics and structural changes of calmodulin upon interaction with the antagonist calmidazolium. BMC Biol. 2022, 20, 176. [Google Scholar] [CrossRef]
- O’Day, D.H. Calmodulin and amyloid beta as coregulators of critical events during the onset and progression of Alzheimer’s disease. Int. J. Mol. Sci. 2023, 24, 1393. [Google Scholar] [CrossRef] [PubMed]
- Poejo, J.; Salazar, J.; Mata, A.M.; Gutierrez-Merino, C. The relevance of amyloid β-calmodulin complexation in neurons and brain degeneration in Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 4976. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Geise, K.P. Calcium/calmodulin-dependent kinase II and Alzheimer’s disease. Molec. Brain 2015, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Day, D.H. Calmodulin binding domains in critical risk proteins involved in neurodegeneration. Curr. Issues Mol. Biol. 2022, 44, 5802–5814. [Google Scholar] [CrossRef] [PubMed]
- Nishino, H.; Naito, E.; Iwashima, A.; Tanaka, K.I.; Matsuura, T.; Fujiki, H.; Sugimura, T. Interaction between quercetin and Ca2+-calmodulin complex: Possible mechanism for anti-tumor-promoting action of the flavonoid. GANN Jpn. J. Cancer Res. 1984, 75, 311–316. [Google Scholar]
- Benítez-King, G.; Huerto-Delgadillo, L.; Antón-Tay, F. Binding of 3H-melatonin to calmodulin. Life Sci. 1993, 53, 201–207. [Google Scholar] [CrossRef]
- Shin, J.S.; Lee, J.; Park, H.J.; Park, S.J.; Kwon, H.J. A new curcumin derivative, HBC, interferes with the cell cycle progression of colon cancer cells via antagonization of the Ca2+/calmodulin function. Chem Biol. 2004, 11, 1455–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.B.; Case, J.D. Melatonin. Fed. Proc. 1960, 19, 590–592. [Google Scholar]
- Roy, J.; Wong, K.Y.; Aquili, L.; Uddin, M.S.; Heng, B.C.; Tipoe, G.L.; Wong, K.H.; Fung, M.L.; Lim, L.W. Role of melatonin in Alzheimer’s disease: From preclinical studies to melatonin-based therapies. Front. Neuroendocrinol. 2022, 65, 100986. [Google Scholar] [CrossRef]
- Ouyang, H.; Vogel, H.J. Melatonin and serotonin interactions with calmodulin: NMR, spectroscopic and biochemical studies. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1383, 37–47. [Google Scholar] [CrossRef]
- Turjanski, A.G.; Estrin, D.A.; Rosenstein, R.E.; McCormick, J.E.; Martin, S.R.; Pastore, A.; Biekofsky, R.R.; Martorana, V. NMR and Molecular Dynamics Studies of the Interaction of Melatonin with Calmodulin. Protein Sci. 2004, 13, 2925–2938. [Google Scholar] [CrossRef] [PubMed]
- Villalobo, A. The multifunctional role of phosphor-calmodulin in pathophysiological processes. Biochem. J. 2018, 475, 4011–4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Vega, E.; Meza, I.; Ramirez-Rodriguez, G.; Benitez-King, G. Melatonin stimulates calmodulin phosphorylation by protein kinase C. J. Pineal Res. 2004, 37, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Flavonols and flavones as BACE-1 inhibitors: Structure–activity relationship in cell-free, cell-based and in silico studies reveal novel pharmacophore features. Biochim. Biophys. Acta 2008, 1780, 819–825. [Google Scholar] [CrossRef]
- Piccialli, I.; Tedeschi, V.; Caputo, L.; D’Errico, S.; Ciccone, R.; De Feo, V.; Secondo, A.; Pannaccione, A. Exploring the therapeutic potential of phytochemicals in Alzheimer’s. disease: Focus on polyphenols and monoterpenes. Front. Pharmacol. 2020, 13, 876614. [Google Scholar] [CrossRef]
- Pagano, K.; Tomaselli, S.; Molinari, H.; Ragona, L. Natural Compounds as Inhibitors of Ab Peptide Aggregation: Chemical Requirements and Molecular Mechanisms. Front. Neurosci. 2020, 14, 619667. [Google Scholar] [CrossRef]
- Fan, Q.; Liu, Y.; Wang, X.; Zhang, Z.; Fu, Y.; Liu, L.; Wang, P.; Ma, H.; Ma, H.; Seeram, N.P.; et al. Ginnalin A Inhibits Aggregation, Reverses Fibrillogenesis, and Alleviates Cytotoxicity of Amyloid β(1-42). ACS Chem. Neurosci. 2020, 11, 638–647. [Google Scholar] [CrossRef]
- Ahmed, M.; Davis, J.; Aucoin, D.; Sato, T.; Ahuja, S.; Aimoto, S.; Elliott, J.I.; Van Nostrand, W.E.; Smith, S.O. Structural conversion of neurotoxic amyloid-beta (1-42) oligomers to fibrils. Nat. Struct. Mol. Biol. 2010, 17, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Du, W.J.; Guo, J.J.; Gao, M.T.; Hu, S.Q.; Dong, X.Y.; Han, Y.F.; Liu, F.F.; Jiang, S.; Sun, Y. Brazilin inhibits amyloid beta-protein fibrillogenesis, remodels amyloid fibrils and reduces amyloid cytotoxicity. Sci. Rep. 2015, 5, 7992. [Google Scholar] [CrossRef] [Green Version]
- Ono, K.; Li, L.; Takamura, Y.; Yoshiike, Y.; Zhu, L.; Han, F.; Mao, X.; Ikeda, T.; Takasaki, J.-I.; Nishijo, H.; et al. Phenolic compounds prevent amyloid beta-protein oligomerization and synaptic dysfunction by site-specific binding. J. Biol. Chem. 2012, 287, 14631–14643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, R.; VanSchouwen, B.; Jafari, N.; Ni, X.; Ortega, J.; Melacini, G. Molecular mechanism for the (-)-epigallocatechin gallate-induced toxic to nontoxic remodeling of abeta oligomers. J. Am. Chem. Soc. 2017, 139, 13720–13734. [Google Scholar] [CrossRef]
- Panyatip, P.; Tadtong, S.; Sousa, E.; Puthongking, P. BACE1 inhibitor, neuroprotective, and neuritogenic activities of melatonin derivatives. Sci. Pharm. 2020, 88, 58. [Google Scholar] [CrossRef]
- Yusof, M.N.I.S.; Abdullah, Z.L.; Othman, N.; Fauzi, F.M. Structure–activity relationship analysis of flavonoids and its inhibitory activity against BACE1 enzyme toward a better therapy for Alzheimer’s disease. Front. Chem. 2022, 10, 874615. [Google Scholar] [CrossRef]
- Hemming, M.L.; Elias, J.E.; Gygi, S.P.; Selkoe, D.J. Identification of beta-secretase (BACE1) substrates using quantitative proteomics. PLoS ONE 2009, 4, e8477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, R. Physiological functions of the b-site amyloid precursor protein cleaving enzyme 1 and 2. Front. Mol. Neurosci. 2017, 10, 97–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConlogue, L.; Buttini, M.; Anderson, J.P.; Brigham, E.F.; Chen, K.S.; Freedman, S.B.; Games, D.; Johnson-Wood, K.; Lee, M.; Zeller, M.; et al. Partial reduction of BACE1 has dramatic effects on Alzheimer plaque and synaptic pathology in APP Transgenic Mice. J. Biol. Chem. 2007, 282, 26326–26334. [Google Scholar] [CrossRef] [Green Version]
- Kimura, R.; Devi, L.; Ohno, M. Partial reduction of BACE1 improves synaptic plasticity, recent and remote memories in Alzheimer’s disease transgenic mice. J. Neurochem. 2010, 113, 248–261. [Google Scholar] [CrossRef] [Green Version]
- Barão, S.; Moechars, D.; Lichtenthaler, S.F.; De Strooper, B. BACE1 Physiological Functions May Limit Its Use as Therapeutic Target for Alzheimer’s Disease. Trends Neurosci. 2016, 39, 158–169. [Google Scholar] [CrossRef]
- Ali, M.Y.; Seong, S.H.; Jung, H.A.; Jannat, S.; Choi, J.S. Kinetics and molecular docking of dihydroxanthyletin-type coumarins from Angelica decursiva that inhibit cholinesterases and BACE1. Arch. Pharm. Res. 2018, 401, 753–764. [Google Scholar] [CrossRef]
- Janat, S.; Balupuri, A.; Ali, M.Y.; Hong, S.S.; Choi, C.W.; Choi, Y.-H.; Ku, J.-M.; Kim, W.J.; Leem, J.Y.; Kim, J.E.; et al. Inhibition of β-site amyloid precursor protein cleaving enzyme 1 and cholinesterases by pterosins via a specific structure-activity relationship with a strong BBB permeability. Exp. Mol. Med. 2019, 51, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, M.; Iraji, A.; Vahedi-Mazdabadi, Y.; Alizadeh, A.; Edraki, N.; Firuzi, O.; Eftekhari, M.; Akbarzadeh, T. Cinnamomum verum J. Presl. bark essential oil: In vitro investigation of anti-cholinesterase, anti-BACE1, and neuroprotective activity. BMC Complement. Med. Therap. 2022, 22, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Rapposelli, S.; Sestito, S.; Herrera-Bravo, J.; Arancibia-Diaz, A.; Salazar, L.A.; Yeskaliyeva, B.; Beyatli, A.; Leyva-Gómez, G.; González-Contreras, C.; et al. Multi-target mechanisms of phytochemicals in Alzheimer’s disease: Effects on oxidative stress, neuroinflammation and protein aggregation. J. Pers. Med. 2022, 12, 1515. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.Y.; Bae, K.; Seong, Y.H.; Song, K.S. Green tea catechins as a BACE1 (beta-secretase) inhibitor. Bioorg. Med. Chem. Lett. 2003, 13, 3905–3908. [Google Scholar] [CrossRef] [PubMed]
- Zhumanova, K.; Lee, G.; Baiseitova, A.; Shah, A.B.; Kim, J.H.; Kim, J.Y.; Lee, K.W.; Park, K.H. Inhibitory mechanism of O-methylated quercetins, highly potent β-secretase inhibitors isolated from Caragana balchaschensis (Kom.) Pojark. J. Ethnopharmacol. 2021, 272, 113935. [Google Scholar] [CrossRef]
- Youn, K.; Yu, Y.; Lee, J.; Jeong, W.-S.; Ho, C.-T.; Jun, M. Polymethoxyflavones: Novel β-secretase (BACE1) inhibitors from citrus peels. Nutrients 2017, 9, 973. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Rodríguez, M.; Correa-Basurto, J.; Gutiérrez, A.; Vitorica, J.; Rosales-Hernández, M.C. Asp32 and Asp228 Determine the Selective Inhibition of BACE1 as Shown by Docking and Molecular Dynamics Simulations. Eur. J. Med. Chem. 2016, 124, 1142–1154. [Google Scholar] [CrossRef]
- Tarozzi, A.; Angeloni, C.; Malaguti, M.; Morroni, F.; Hrelia, S.; Hrelia, P. Sulforaphane as a potential protective phytochemical against neurodegenerative diseases. Oxid. Med. Cell. Longev. 2013, 2013, 415078. [Google Scholar] [CrossRef] [Green Version]
- Brandenburg, L.O.; Kipp, M.; Lucius, R.; Pufe, T.; Wruck, C.J. Sulforaphane suppresses LPS-induced inflammation in primary rat microglia. Inflamm. Res. 2010, 59, 443–450. [Google Scholar] [CrossRef]
- Zhang, R.; Miao, Q.W.; Zhu, C.X.; Zhao, Y.; Liu, L.; Yang, J.; Li, A. Sulforaphane ameliorates neurobehavioral deficits and protects the brain from amyloid beta deposits and peroxidation in mice with Alzheimer-like lesions. Am. J. Alzheimer’s Dis. Other Dement. 2015, 30, 183–191. [Google Scholar] [CrossRef]
- Youn, K.; Yoon, J.-H.; Lee, N.; Lim, G.; Lee, J.; Sang, S.; Ho, C.-T.; Jun, M. Discovery of sulforaphane as a potent BACE1 inhibitor based on kinetics and computational studies. Nutrients 2020, 12, 3026. [Google Scholar] [CrossRef]
- Williams, P.; Sorribas, A.; Howes, M.J. Natural products as a source of Alzheimer’s drug leads. Nat. Prod. Rep. 2010, 28, 48–77. [Google Scholar] [CrossRef]
- Fang, W.-S.; Sun, D.-Y.; Yang, S.; Cheng, C.; Moschke, K.; Li, T.; Sun, S.; Lichtenthaler, S.F.; Huang, J.; Wang, Y. Discovery of a series of selective and cell permeable beta-secretase (BACE1) inhibitors by fragment linking with the assistance of STD-NMR. Bioorg. Chem. 2019, 92, 103253. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.Y.; Kwon, S.H.; Seong, Y.H.; Bae, K.; Hur, J.M.; Lee, Y.-Y.; Suh, D.-Y.; Song, K.-S. b-Secretase (BACE1)-inhibiting stilbenoids from Smilax Rhizoma. Phytomedicine 2007, 14, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Youn, K.; Park, J.; Lee, S.; Lee, S.; Lee, J.; Yun, E.; Jeong, W.-S.; Jun, M. BACE1 Inhibition by genistein: Biological evaluation, kinetic analysis, and molecular docking simulation. J. Med. Food 2018, 21, 416–420. [Google Scholar] [CrossRef]
- Ali, M.Y.; Jannat, S.; Jung, H.A.; Choi, R.J.; Roy, A.; Choi, J.S. Anti-Alzheimer’s disease potential of coumarins from Angelica decursiva and Artemisia capillaris and structure-activity analysis. Asian Pac. J. Trop. Med. 2016, 9, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chlebek, J.; De Simone, A.; Hošťálková, A.; Opletal, L.; Pérez, C.; Pérez, D.I.; Havlíková, L.; Cahlíková, L.; Andrisano, V. Application of BACE1 immobilized enzyme reactor for the characterization of multifunctional alkaloids from Corydalis cava (Fumariaceae) as Alzheimer’s disease targets. Fitoterapia 2016, 109, 241–247. [Google Scholar] [CrossRef]
- Gruenwald, J.; Freder, J.; Armbruester, N. Cinnamon and health. Crit. Rev. Food Sci. Nutr. 2010, 50, 822–834. [Google Scholar] [CrossRef]
- Qian, D.; Wang, Q.; Lin, S.; Li, Y.; Gu, X.; Xia, C.; Xu, Y.; Zhang, T.; Yang, L.; Wu, Q.; et al. Identification of potential targets of cinnamon for treatment against Alzheimer’s disease-related GABA-ergic synaptic dysfunction using network pharmacology. Sci. Rep. 2022, 12, 19959. [Google Scholar] [CrossRef]
- Sun, D.-Y.; Cheng, C.; Moschke, K.; Huang, J.; Fang, W.-S. Extensive-structure modification on luteolin-cinnamic acid conjugates leading to BACE1 inhibitors with optimal pharmacological properties. Molecules 2020, 25, 102. [Google Scholar]
- Khan, A.H. The Greatest Elixir (Exir Azam); Tehran Research Institute for Islamic and Complementary Medicine, Tehran University of Medical Sciences: Tehran, Iran, 2009; pp. 267–280. [Google Scholar]
- Benny, A.; Thomas, J. Essential oils as treatment strategy for Alzheimer’s disease: Current and future perspectives. Planta Med. 2019, 85, 239–248. [Google Scholar] [PubMed] [Green Version]
- Videira, R.; Castanheira, P.; Grãos, M.; Salgueiro, L.; Faro, C.; Cavaleiro, C. A necrodane monoterpenoid from Lavandula luisieri essential oil as a cell-permeable inhibitor of BACE-1, the β-secretase in Alzheimer’s disease. Flavour Fragr. J. 2013, 28, 380–388. [Google Scholar] [CrossRef]
- Marambaud, P.; Zhao, H.; Davies, P. Resveratrol promotes clearance of Alzheimer’s disease amyloid-b peptides. J. Biol. Chem. 2005, 280, 37377–37382. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.W.; Choi, Y.H.; Cha, M.-R.; Kim, Y.S.; Yon, G.H.; Hong, K.S.; Park, W.-K.; Kim, Y.H.; Ryu, S.Y. In vitro BACE-1 inhibitory activity of resveratrol oligomers from the seed extract of Paeonia lactiflora. Planta Med. 2011, 77, 374–376. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yoo, M.Y.; Choi, C.W.; Cha, M.R.; Yon, G.H.; Kwon, D.Y.; Kim, Y.S.; Park, W.-K.; Ryu, S.Y. A new specific BACE-1 inhibitor from the stem bark extract of Vitis vinifera. Planta Med. 2009, 75, 537–540. [Google Scholar] [CrossRef]
- Koukoulitsa, C.; Villalonga-Barber, C.; Csonka, R.; Alexi, X.; Leonis, G.; Dellis, D.; Hamelink, E.; Belda, O.; Barry, R.; Micha-Screttas, M.; et al. Biological and computational evaluation of resveratrol inhibitors against Alzheimer’s disease. J. Enzyme Inhib. Med. Chem. 2016, 31, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Thangthaeng, M. Walnut extract modulates activation of microglia through alteration in intracellular calcium concentration. Nutrition Res. 2018, 49, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3 kinase signalling. J. Inflamm. 2012, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Carey, A.N.; Fisher, D.R.; Joseph, J.A.; Shukitt-Hale, B. The ability of walnut extract and fatty acids to protect against the deleterious effects of oxidative stress and inflammation in hippocampal cells. Nutr. Neurosci. 2013, 16, 13–20. [Google Scholar] [CrossRef]
- Casal, C.; Tusell, J.M.; Serratosa, J. Role of calmodulin in the differentiation/activation of microglial cells. Brain Res. 2001, 902, 101–107. [Google Scholar] [CrossRef]
- Fuloria, S.; Mehta, J.; Chandel, A.; Sekar, M.; Rani, N.N.I.M.; Begum, M.Y.; Subramaniyan, V.; Chidambaram, K.; Thangavelu, L.; Nordin, R.; et al. A comprehensive review on the therapeutic potential of Curcuma longa Linn. in relation to its major active constituent curcumin. Front. Pharmacol. 2022, 13, 820806. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Guo, F.; Luo, J.; Zhang, Y.; Liu, J.; Zhang, Y.; Zheng, X.; Wan, F.; Ding, W. Functional analysis of an up regulated calmodulin gene related to the agaricidal activity of curcumin against Tetranychus cinnabarinus (Boisduval). Pest Management Sci. 2021, 77, 719–730. [Google Scholar] [CrossRef]
- McLachlan, D.R.C.; Wong, L.; Bergeron, C.; Baimbridge, K.G. Calmodulin and Calbindin D28K in Alzheimer Disease. Alzheimer Dis. Assoc. Disord. 1987, 1, 171–179. [Google Scholar] [CrossRef]
- Caldwell, G.W.; Yan, Z.; Lang, W.; Masucci, J.A. The IC(50) concept revisited. Curr. Top. Med. Chem. 2012, 12, 1282–1290. [Google Scholar] [CrossRef]
- Cummings, J.; Lee, G.; Nahed, P.; Kambar, M.E.Z.N.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2022. Alzheimer’s Dement. 2022, 8, e12295. [Google Scholar] [CrossRef] [PubMed]
- Alaqeel, N.K.; AlSheikh, M.H.; Al-Hariri, M.T. Quercetin nanoemulsion ameliorates neuronal dysfunction in experimental Alzheimer’s disease model. Antioxidants 2022, 11, 1986. [Google Scholar] [CrossRef]
- Campisi, A.; Sposito, G.; Pellitteri, R.; Santonocito, D.; Bisicchia, J.; Raciti, G.; Russo, C.; Nardiello, P.; Pignatello, R.; Casamenti, F.; et al. Effect of unloaded and curcumin-loaded solid lipid nanoparticles on tissue transglutaminase isoforms expression levels in an experimental model of Alzheimer’s disease. Antioxidants 2022, 11, 1863. [Google Scholar] [CrossRef]
- Gregory, J.; Vengalasetti, Y.V.; Bredesen, D.E.; Rao, R.V. Neuroprotective Herbs for the Management of Alzheimer’s Disease. Biomolecules 2021, 11, 543. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective effects of quercetin in Alzheimer’s disease. Biomolecules 2020, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Kola, P.; Kaur, D.; Singla, G.; Mishra, V.; Panesar, P.S.; Mallikarjunan, K.; Krishania, M. Therapeutic potential of nutraceuticals and dietary supplements in the prevention of viral diseases: A review. Front. Nutr. 2021, 8, 679312. [Google Scholar] [CrossRef]
- Zhou, X.; Venigalla, M.; Raju, R.; Münch, G. Pharmacological considerations for treating neuroinflammation with curcumin in Alzheimer’s disease. J. Neural Trans. 2022, 129, 755–771. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O'Day, D.H. Phytochemical Interactions with Calmodulin and Critical Calmodulin Binding Proteins Involved in Amyloidogenesis in Alzheimer’s Disease. Biomolecules 2023, 13, 678. https://doi.org/10.3390/biom13040678
O'Day DH. Phytochemical Interactions with Calmodulin and Critical Calmodulin Binding Proteins Involved in Amyloidogenesis in Alzheimer’s Disease. Biomolecules. 2023; 13(4):678. https://doi.org/10.3390/biom13040678
Chicago/Turabian StyleO'Day, Danton H. 2023. "Phytochemical Interactions with Calmodulin and Critical Calmodulin Binding Proteins Involved in Amyloidogenesis in Alzheimer’s Disease" Biomolecules 13, no. 4: 678. https://doi.org/10.3390/biom13040678