
Piperitone (p-Menth-1-En-3-One): A New Repellent for Tea Shot Hole Borer (Coleoptera: Curculionidae) in Florida Avocado Groves
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lures, Repellents, and Traps
2.2. Field Evaluation of Piperitone
2.3. Field Comparison of Repellents
2.4. Electroantennography
2.5. Analysis of Repellent Contents
2.6. Volatile Collections and Analysis of Repellent Emissions
2.7. Statistical Analysis
3. Results
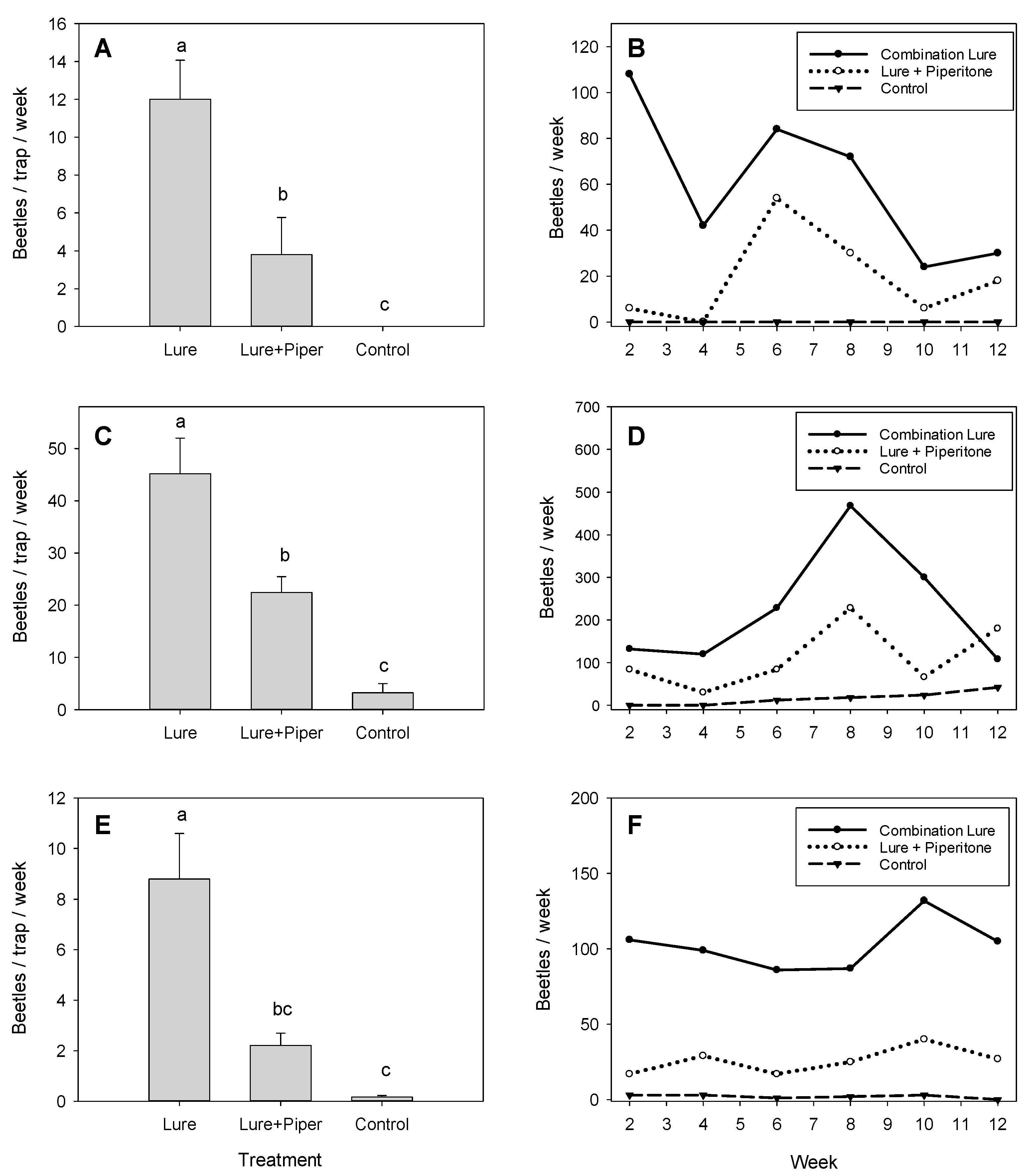
3.1. Field Evaluation of Piperitone
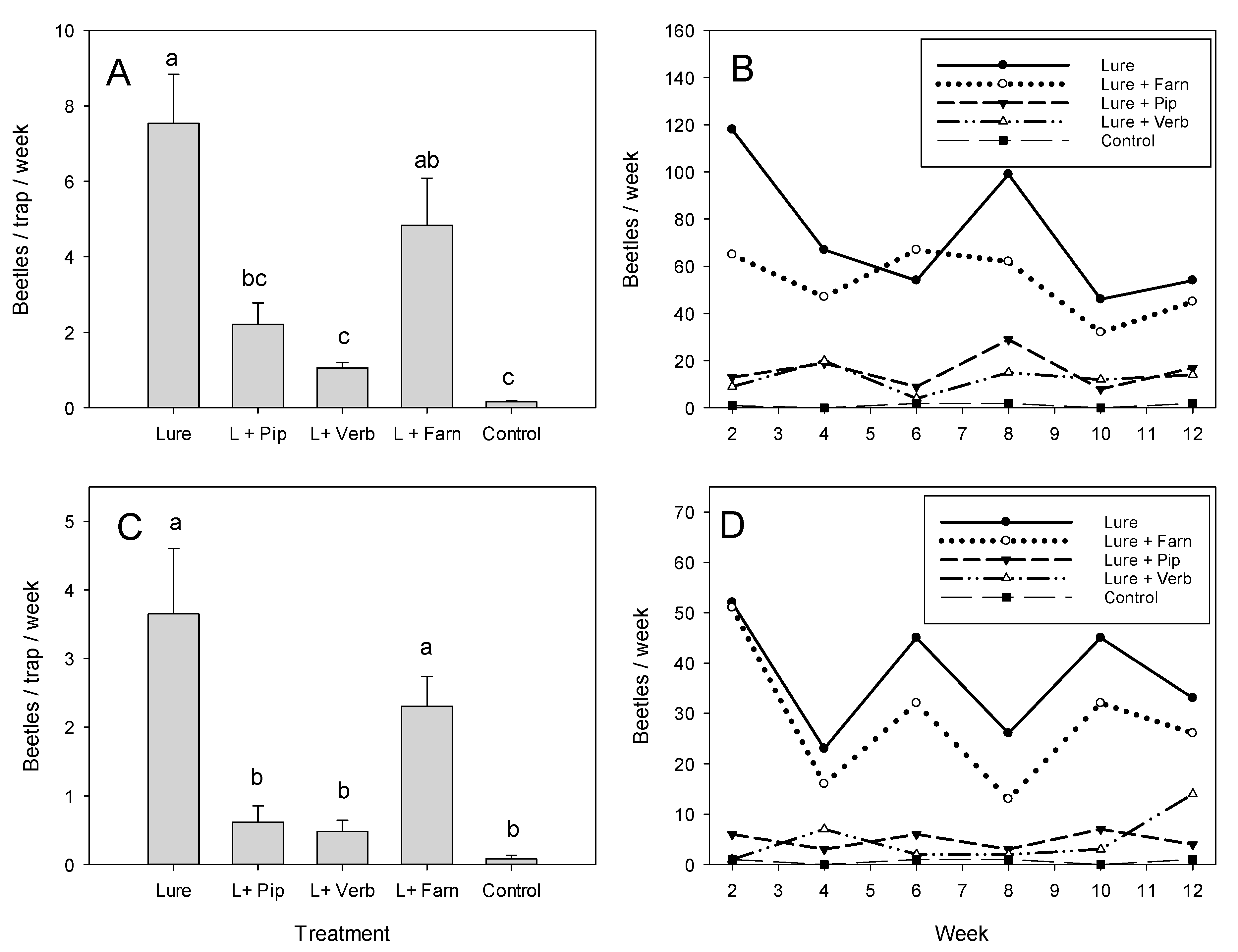
3.2. Field Comparison of Repellents
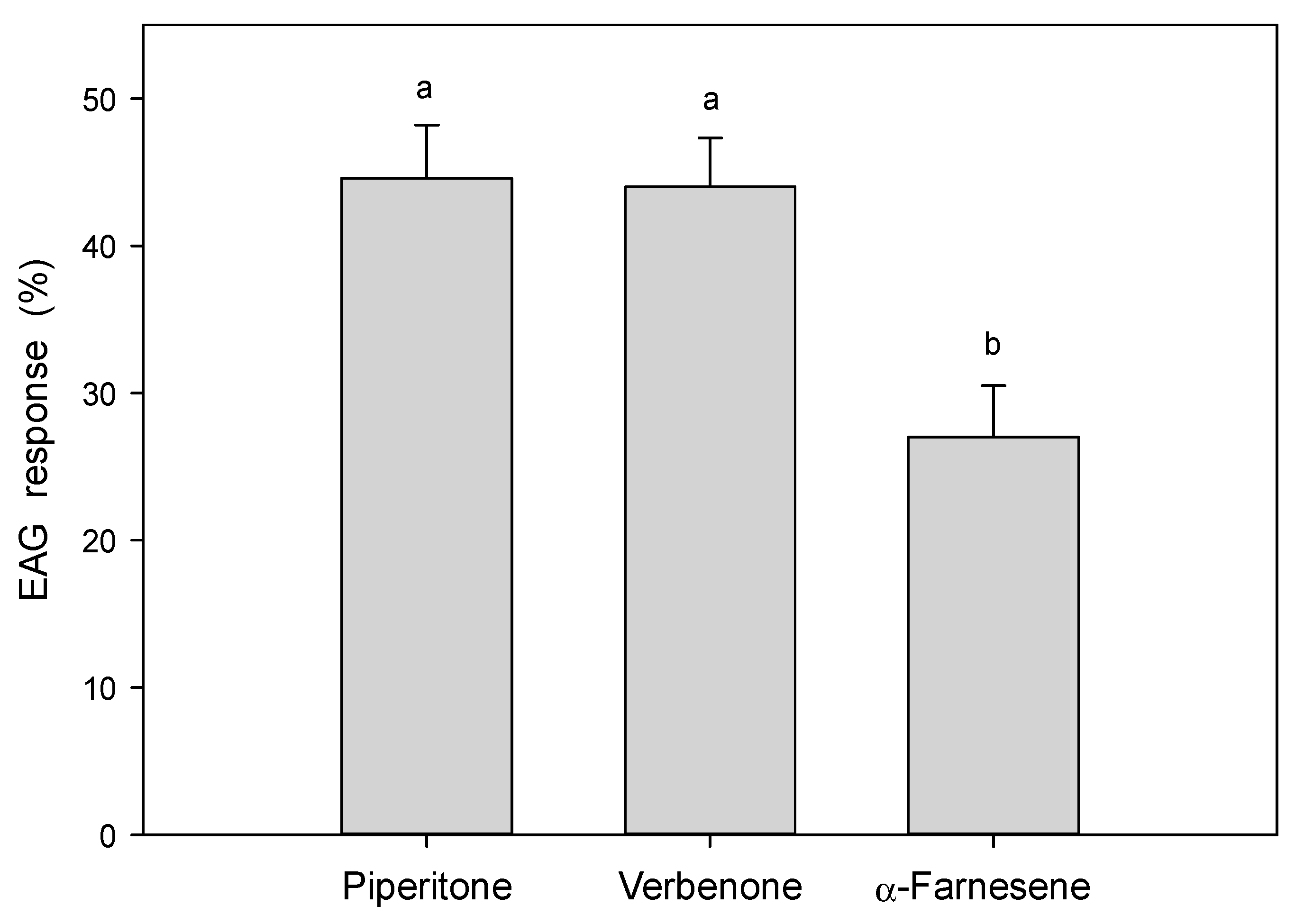
3.3. Electroantennography
3.4. Chemical Analysis of Repellent Contents
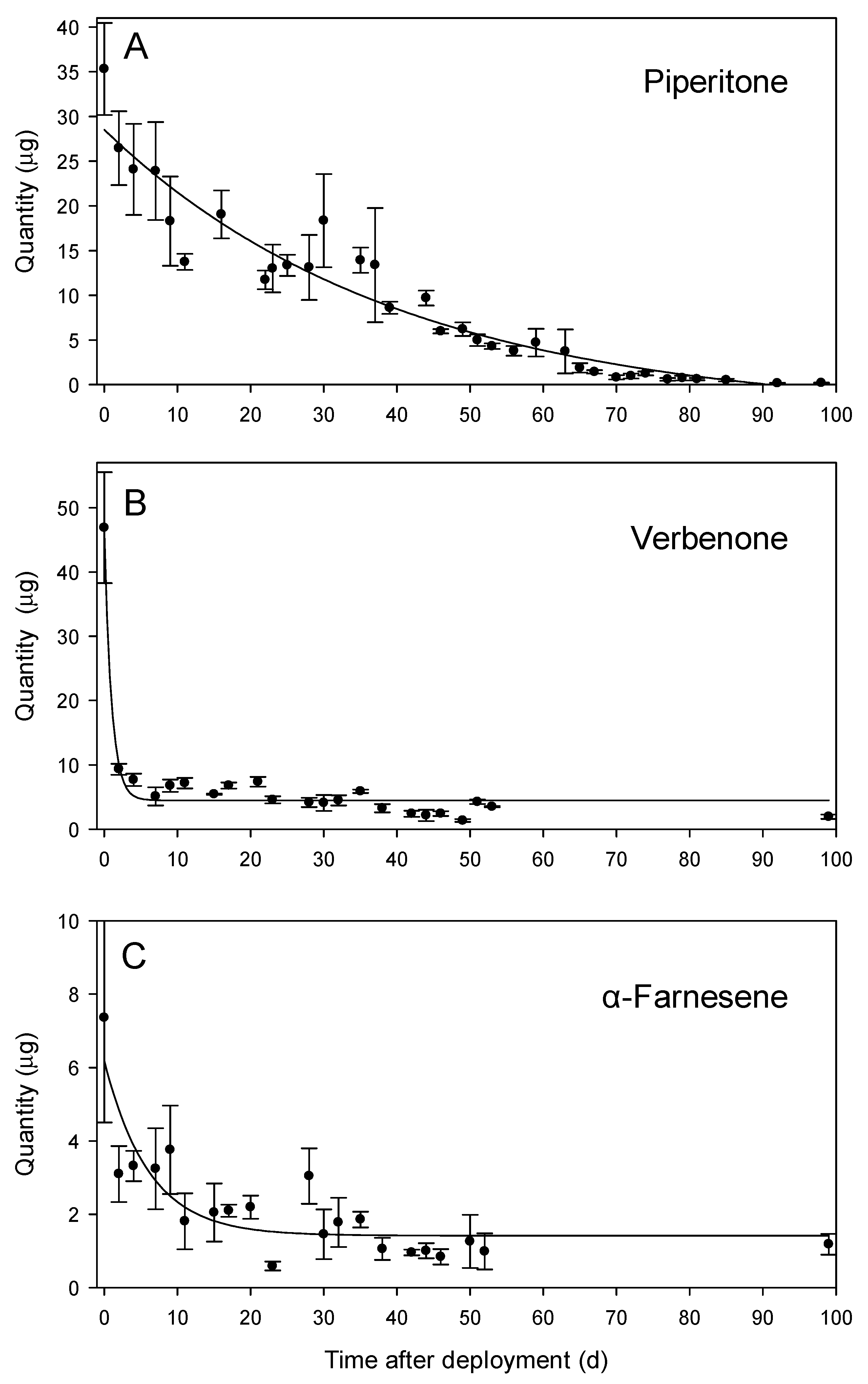
3.5. Temporal Analysis of Repellent Emissions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Danthanarayana, W. The distribution and Host Range of the Shot-Hole Borer (Xyleborus fornicatus Eichh.) of tea. Tea Q. 1968, 39, 61–69. Available online: http://dl.nsf.gov.lk/handle/1/12743 (accessed on 23 March 2023).
- Mendel, Z.; Protasov, A.; Sharon, M.; Zveibil, A.; Yehuda, S.B.; O’donnell, K.; Rabaglia, R.; Wysoki, M.; Freeman, S. An Asian ambrosia beetle Euwallacea fornicatus and its novel symbiotic fungus Fusarium sp. pose a serious threat to the Israeli avocado industry. Phytoparasitica 2012, 40, 235–238. [Google Scholar] [CrossRef]
- Eskalen, A.; Gonzalez, A.; Wang, D.H.; Twizeyimana, M.; Mayorquin, J.S.; Lynch, S.C. First Report of a Fusarium sp. and Its Vector Tea Shot Hole Borer (Euwallacea fornicatus) Causing Fusarium Dieback on Avocado in California. Plant Disease 2012, 96, 1070. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, D.; Cruz, L.F.; Kendra, P.E.; Narvaez, T.I.; Montgomery, W.S.; Monterroso, A.; De Grave, C.; Cooperband, M.F. Distribution, pest status and fungal associates of Euwallacea nr. fornicatus in Florida avocado groves. Insects 2016, 7, 55. [Google Scholar] [CrossRef]
- Carrillo, D.; Duncan, R.E.; Peña, J.E. Ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) that breed in avocado wood in Florida. Fla. Entomol. 2012, 95, 573–579. [Google Scholar] [CrossRef]
- García-Avila, C.D.J.; Trujillo-Arriaga, F.J.; López-Buenfil, J.A.; González-Gómez, R.; Carrillo, D.; Cruz, L.F.; Ruiz-Galván, I.; Quezada-Salinas, A.; Acevedo-Reyes, N. First Report of Euwallacea nr. fornicatus (Coleoptera: Curculionidae) in Mexico. Fla. Entomol. 2016, 99, 555–556. [Google Scholar] [CrossRef] [Green Version]
- Paap, T.; de Beer, Z.W.; Migliorini, D.; Nel, W.J.; Wingfield, M.J. The polyphagous shot hole borer (PSHB) and its fungal symbiont Fusarium euwallaceae: A new invasion in South Africa. Australas. Plant Pathol. 2018, 47, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Stouthamer, R.; Rugman-Jones, P.; Thu, P.Q.; Eskalen, A.; Thibault, T.; Hulcr, J.; Wang, L.; Jordal, B.H.; Chen, C.; Cooperband, M. Tracing the origin of a cryptic invader: Phylogeography of the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) species complex. Agric. For. Entomol. 2017, 19, 366–375. [Google Scholar] [CrossRef]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. nov. A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Konkol, J.L.; Carrillo, D.; Freeman, S.; Mendel, Z. An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on avocado and other plant hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef]
- Lynch, S.C.; Twizeyimana, M.; Mayorquin, J.S.; Wang, D.H.; Na, F.; Kayim, M.; Kasson, M.T.; Thu, P.Q.; Bateman, C.; Rugman-Jones, P. Identification, pathogenicity and abundance of Paracremonium pembeum sp. nov. and Graphium euwallaceae sp. nov. Two newly discovered mycangial associates of the polyphagous shot hole borer (Euwallacea sp.) in California. Mycologia 2016, 108, 313–329. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Sink, S.; Libeskind-Hadas, R.; Hulcr, J.; Kasson, M.T.; Ploetz, R.C.; Konkol, J.L.; Ploetz, J.N.; Carrillo, D.; Campbell, A. Discordant phylogenies suggest repeated host shifts in the Fusarium–Euwallacea ambrosia beetle mutualism. Fungal Genet. Biol. 2015, 82, 277–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Gomez, D.F.; Beaver, R.A.; Hulcr, J.; Cognato, A.I. Reassessment of the species in the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) complex after the rediscovery of the “lost” type specimen. Insects 2019, 10, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaver, R.; Wilding, N.; Collins, N.; Hammond, P.; Webber, J. Insect-fungus relationships in the bark and ambrosia beetles. Insect Fungus Interact. 1989, 121, 143. [Google Scholar] [CrossRef]
- Farrell, B.D.; Sequeira, A.S.; O’Meara, B.C.; Normark, B.B.; Chung, J.H.; Jordal, B.H. The evolution of agriculture in beetles (Curculionidae: Scolytinae and Platypodinae). Evolution 2001, 55, 2011–2027. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C. Ecology and evolution of mycophagous bark beetles and their fungal partners. In Ecological and Evolutionary Advances in Insect-Fungal Associations; Vega, F.E., Blackwell, M.A., Eds.; Oxford University Press: Oxford, UK, 2005; pp. 257–291. [Google Scholar]
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [Green Version]
- Six, D.L. Bark beetle-fungus symbioses. Insect Symbiosis 2003, 1, 97–114. [Google Scholar]
- Rabaglia, R., XVII. Scolytinae Latreille 1807. In American Beetles, Vol. 2, Polyphaga: Scarabaeoidea through Curculionoidea; Arnett, R., Thomas, M., Skelley, P., Frank, J., Eds.; CRC Press: Boca Raton, FL, USA, 2002; Volume 2, pp. 792–805. [Google Scholar]
- Eskalen, A.; Stouthamer, R.; Lynch, S.C.; Rugman-Jones, P.F.; Twizeyimana, M.; Gonzalez, A.; Thibault, T. Host range of Fusarium dieback and its ambrosia beetle (Coleoptera: Scolytinae) vector in southern California. Plant Dis. 2013, 97, 938–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, S.; Protasov, A.; Sharon, M.; Mohotti, K.; Eliyahu, M.; Okon-Levy, N.; Maymon, M.; Mendel, Z. Obligate feed requirement of Fusarium sp. nov., an avocado wilting agent, by the ambrosia beetle Euwallacea aff. fornicata. Symbiosis 2012, 58, 245–251. [Google Scholar] [CrossRef]
- Mendel, Z.; Protasov, A.; Maoz, Y.; Maymon, M.; Miller, G.; Elazar, M.; Freeman, S. The role of Euwallacea nr. fornicatus (Coleoptera: Scolytinae) in the wilt syndrome of avocado trees in Israel. Phytoparasita 2017, 45, 341–359. [Google Scholar] [CrossRef]
- Carrillo, D.; Narvaez, T.; Cossé, A.A.; Stouthamer, R.; Cooperband, M. Attraction of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae) to lures containing quercivorol. Fla. Entomol. 2015, 98, 780–782. [Google Scholar] [CrossRef]
- Kendra, P.E.; Owens, D.; Montgomery, W.S.; Narvaez, T.I.; Bauchan, G.R.; Schnell, E.Q.; Tabanca, N.; Carrillo, D. α-Copaene is an attractant, synergistic with quercivorol, for improved detection of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae). PLoS ONE 2017, 12, e0179416. [Google Scholar] [CrossRef]
- Owens, D.; Cruz, L.F.; Montgomery, W.S.; Narvaez, T.I.; Schnell, E.Q.; Tabanca, N.; Duncan, R.E.; Carrillo, D.; Kendra, P.E. Host range expansion and increasing damage potential of Euwallacea nr. fornicatus (Coleoptera: Curculionidae) in Florida. Fla. Entomol. 2018, 101, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Boland, J.M. The impact of an invasive ambrosia beetle on the riparian habitats of the Tijuana River Valley, California. Peer J. 2016, 4, e2141. [Google Scholar] [CrossRef]
- United States Department of Agriculture, N.A.S.S. Quick Stats. Available online: https://quickstats.nass.usda.gov/ (accessed on 1 March 2023).
- Fraedrich, S. California laurel is susceptible to laurel wilt caused by Raffaelea lauricola. Plant Dis. 2008, 92, 1469. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.; Fraedrich, S.; Aghayeva, D. Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Hughes, M.A.; Riggins, J.J.; Koch, F.H.; Cognato, A.I.; Anderson, C.; Formby, J.P.; Dreaden, T.J.; Ploetz, R.C.; Smith, J.A. No rest for the laurels: Symbiotic invaders cause unprecedented damage to southern USA forests. Biol. Invasions 2017, 19, 2143–2157. [Google Scholar] [CrossRef]
- Carrillo, D.; Duncan, R.; Ploetz, J.; Campbell, A.; Ploetz, R.; Peña, J. Lateral transfer of a phytopathogenic symbiont among native and exotic ambrosia beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Konkol, J.L.; Narvaez, T.; Duncan, R.E.; Saucedo, R.J.; Campbell, A.; Mantilla, J.; Carrillo, D.; Kendra, P.E. Presence and prevalence of Raffaelea lauricola, cause of laurel wilt, in different species of ambrosia beetle in Florida, USA. J. Econ. Entomol. 2017, 110, 347–354. [Google Scholar] [CrossRef]
- Chen, Y.; Coleman, T.W.; Poloni, A.L.; Nelson, L.; Seybold, S.J. Reproduction and control of the invasive polyphagous shot hole borer, Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae), in three species of hardwoods: Effective sanitation through felling and chipping. Environ. Entomol. 2020, 49, 1155–1163. [Google Scholar] [CrossRef]
- Jones, M.E.; Kabashima, J.; Eskalen, A.; Dimson, M.; Mayorquin, J.S.; Carrillo, J.D.; Hanlon, C.C.; Paine, T.D. Evaluations of insecticides and fungicides for reducing attack rates of a new invasive ambrosia beetle (Euwallacea sp., Coleoptera: Curculionidae: Scolytinae) in infested landscape trees in California. J. Econ. Entomol. 2017, 110, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Kendra, P.E.; Montgomery, W.S.; Narvaez, T.I.; Carrillo, D. Comparison of trap designs for detection of Euwallacea nr. fornicatus and other Scolytinae (Coleoptera: Curculionidae) that vector fungal pathogens of avocado trees in Florida. J. Econ. Entomol. 2020, 113, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.; Kendra, P.E.; Tabanca, N.; Narvaez, T.I.; Montgomery, W.S.; Schnell, E.Q.; Carrillo, D. Quantitative analysis of contents and volatile emissions from α-copaene and quercivorol lures, and longevity for attraction of Euwallacea nr. fornicatus in Florida. J. Pest Sci. 2019, 92, 237–252. [Google Scholar] [CrossRef]
- Dodge, C.; Coolidge, J.; Cooperband, M.; Cossé, A.; Carrillo, D.; Stouthamer, R. Quercivorol as a lure for the polyphagous and Kuroshio shot hole borers, Euwallacea spp. nr. fornicatus (Coleoptera: Scolytinae), vectors of Fusarium dieback. Peer J. 2017, 5, e3656. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-L.; Rugman-Jones, P.; Liao, Y.-C.; Fernandez, V.; Chien, I.; Dodge, C.; Cooperband, M.F.; Tuan, S.-J.; Stouthamer, R. The attractiveness of α-copaene to members of the Euwallacea fornicatus (Coleoptera: Curculionidae) species complex in California and Taiwan. J. Econ. Entomol. 2021, 115, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.; Seo, M.; Montgomery, W.S.; Rivera, M.J.; Stelinski, L.L.; Kendra, P.E. Dispersal behaviour of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae) in avocado groves and estimation of lure sampling range. Agric. For. Entomol. 2019, 21, 199–208. [Google Scholar] [CrossRef]
- Byers, J.A.; Maoz, Y.; Fefer, D.; Levi-Zada, A. Semiochemicals affecting attraction of ambrosia beetle Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) to quercivorol: Developing push-pull control. J. Econ. Entomol. 2020, 113, 2120–2127. [Google Scholar] [CrossRef]
- Byers, J.A.; Maoz, Y.; Wakarchuk, D.; Fefer, D. Inhibitory effects of semiochemicals on the attraction of an ambrosia beetle Euwallacea nr. fornicatus to quercivorol. J. Chem. Ecol. 2018, 44, 565–575. [Google Scholar] [CrossRef]
- Vega, F.E.; Simpkins, A.; Miranda, J.; Harnly, J.M.; Infante, F.; Castillo, A.; Wakarchuk, D.; Cossé, A. A potential repellent against the coffee berry borer (Coleoptera: Curculionidae: Scolytinae). J. Insect Sci. 2017, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, S.B.; Miller, D.R. Effect of verbenone on five species of bark beetles (Coleoptera: Scolytidae) in lodgepole pine forests. Environ. Entomol. 2002, 31, 759–765. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Deyrup, M.A.; Guillén, L.; Epsky, N.D. Xyleborus glabratus, X. affinis, and X. ferrugineus (Coleoptera: Curculionidae: Scolytinae): Electroantennogram responses to host-based attractants and temporal patterns in host-seeking flight. Environ. Entomol. 2012, 41, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Kendra, P.E.; Epsky, N.D.; Montgomery, W.S.; Heath, R.R. Response of Anastrepha suspensa (Diptera: Tephritidae) to Terminal Diamines in a Food-based Synthetic Attractant. Environ. Entomol. 2008, 37, 1119–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corp: Carol Stream, IL, USA, 2007; pp. 1–804. [Google Scholar]
- van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef] [PubMed]
- MassFinder. MassFinder Software; Version 3; Hochmuth Scientific Consulting: Hamburg, Germany, 2004. [Google Scholar]
- FFNSC3. Flavors and fragrances of natural and synthetic compounds. 2015.
- Wiley Science Solution. Wiley Registry 12th Edition/NIST 2020 Mass Spectral Library; Scientific Instrument Services Inc.: Ringoes, NJ, USA, 2020. [Google Scholar]
- Kendra, P.E.; Tabanca, N.; Cruz, L.F.; Menocal, O.; Schnell, E.Q.; Carrillo, D. Volatile emissions and relative attraction of the fungal symbionts of tea shot hole borer (Coleoptera: Curculionidae). Biomolecules 2022, 12, 97. [Google Scholar] [CrossRef] [PubMed]
- Lazarević, J.; Radulović, N.; Palić, R.; Zlatković, B. Chemical Composition of the Essential Oil of Doronicum austriacum Jacq. subsp. giganteum (Griseb.) Stoj. et Stef. (Compositae) from Serbia. J. Essent. Oil Res. 2009, 21, 507–510. [Google Scholar] [CrossRef]
- Pino, J.A.; Marbot, R.; Payo, A.; Chao, D.; Herrera, P.; Martí, M.P. Leaf Oils of Two Cuban Asteraceae Species: Pluchea carolinensis Jacq. and Ambrosia hispida Pursh. J. Essent. Oil Res. 2005, 17, 318–320. [Google Scholar] [CrossRef]
- Upadhyaya, K.; Dixit, V.K.; Padalia, R.C.; Mathela, C.S. Chemical Composition of the Essential Oil of Caryopteris grata Benth. J. Essent. Oil Res. 2009, 21, 69–70. [Google Scholar] [CrossRef]
- Remoroza, C.A.; Liang, Y.; Mak, T.D.; Mirokhin, Y.; Sheetlin, S.L.; Yang, X.; San Andres, J.V.; Power, M.L.; Stein, S.E. Increasing the coverage of a mass spectral library of milk oligosaccharides using a hybrid-search-based bootstrapping method and milks from a wide variety of mammals. Anal. Chem. 2020, 92, 10316–10326. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The Use of Push-Pull Strategies in Integrated Pest Management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.R.; Pickett, J.A. The ‘Push-Pull’Strategy for Stemborer management: A Case Study in Exploiting Biodiversity and Chemical Ecology; CSIRO: Canberra, Australia, 2004; pp. 155–164.
- Byers, J.A.; Wood, D.L.; Craig, J.; Hendry, L.B. Attractive and inhibitory pheromones produced in the bark beetle, Dendroctonus brevicomis, during host colonization: Regulation of inter- and intraspecific competition. J. Chem. Ecol. 1984, 10, 861–877. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Kivimäenpää, M.; Nizkorodov, S.A. Plant-derived secondary organic material in the air and ecosystems. Trends Plant Sci. 2017, 22, 744–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindgren, B.S.; Nordlander, G.; Birgersson, G. Feeding deterrence of verbenone to the pine weevil, Hylobius abietis (L.) (Col., Curculionidae). J. Appl. Entomol. 1996, 120, 397–403. [Google Scholar] [CrossRef]
- Salom, S.M.; Carlson, J.A.; Ang, B.N.; Grosman, D.M.; Day, E.R. Laboratory evaluation of biologically-based compounds as antifeedants for the pales weevil, Hylobius pales (Herbst) (Coleoptera: Curculionidae). J. Entomol. Sci. 1994, 29, 407–419. [Google Scholar] [CrossRef]
- Cloonan, K.R.; Montgomery, W.S.; Narvaez, T.I.; Carrillo, D.; Kendra, P.E. Community of bark and ambrosia beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in agricultural and forest ecosystems with laurel wilt. Insects 2022, 13, 971. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Pruett, G.E.; Mayfield, A.E., III; MacKenzie, M.; Deyrup, M.A.; Bauchan, G.R.; Ploetz, R.C.; Epsky, N.D. North American Lauraceae: Terpenoid emissions, relative attraction and boring preferences of redbay ambrosia beetle, Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae). PLoS ONE 2014, 9, e102086. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.F.; Willhalm, B.; Bowie, J.H. Mass spectra and organic analysis. Part VIII. The mass spectra of piperitone, the piperitols, and related products. J. Chem. Soc. B. Phys. Org. 1967, 392–400. [Google Scholar] [CrossRef]
- Synergy Semiochemicals Corporation. Verbenone. Available online: https://semiochemical.com/verbenone/ (accessed on 23 March 2023).
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| *LKIexp | **LKIlit | Compounds | % | ***Sim. | ****IM |
|---|---|---|---|---|---|
| 1440 | 1440 a | aromadendrene | 0.01 ± 0.00 | 922 | RI, MS, Std |
| 1457 | 1456 a | (E)-β-farnesene | 39.52 ± 0.47 | 908 | RI, MS, Std |
| 1480 | 1480 a | ar-curcumene | 0.92 ± 0.03 | 927 | RI, MS, Std |
| 1482 | 1482 a | γ-curcumene | 0.43 ± 0.02 | 921 | RI, MS, |
| 1491 | 1495 a | cis-cadina-1,4-diene | 1.39 ± 0.03 | 890 | RI, MS, |
| 1494 | 1499 b | (Z,E)-α-farnesene | 1.30 ± 0.00 | 875 | RI, MS, |
| 1502 | 1505 c | (Z)-α-bisabolene | 5.74 ± 0.06 | 922 | RI, MS, |
| 1506 | 1505 a | (E,E)-α-farnesene | 42.74 ± 0.31 | 921 | RI, MS, Std |
| 1511 | 1512 d | β-curcumene | 0.20 ± 0.02 | 866 | RI, MS, |
| 1513 | 1513 a | γ-cadinene | 1.05 ± 0.02 | 867 | RI, MS, |
| 1515 | 1515 a | (Z)-γ-bisabolene | 3.35 ± 0.05 | 894 | RI, MS, |
| 1521 | 1522 a | trans-calamenene | 0.12 ± 0.01 | 849 | RI, MS, |
| 1522 | 1523 a | δ-cadinene | 1.10 ± 0.02 | 888 | RI, MS, |
| 1530 | 1531 a | (E)-γ-bisabolene | 1.29 ± 0.02 | 874 | RI, MS, |
| 1545 | 1547 a | (E)-α-bisabolene | 0.85 ± 0.02 | 913 | RI, MS, |
| Total | 100 ± 0.00 |
| Samples | (R)-(-)- Piperitone | (S)-(+)- Piperitone | (R)-(+)- Verbenone | (S)-(-)- Verbenone |
|---|---|---|---|---|
| Piperitone dispenser | 82.93 ± 0.21 | 17.07 ± 0.21 | ||
| Piperitone standard | 87.66 ± 0.10 | 12.34 ± 0.10 | ||
| Verbenone dispenser | 16.18 ± 0.01 | 83.82 ± 0.01 | ||
| Verbenone standard | 22.68 ± 0.12 | 77.32 ± 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kendra, P.E.; Montgomery, W.S.; Tabanca, N.; Schnell, E.Q.; Vázquez, A.; Menocal, O.; Carrillo, D.; Cloonan, K.R. Piperitone (p-Menth-1-En-3-One): A New Repellent for Tea Shot Hole Borer (Coleoptera: Curculionidae) in Florida Avocado Groves. Biomolecules 2023, 13, 656. https://doi.org/10.3390/biom13040656
Kendra PE, Montgomery WS, Tabanca N, Schnell EQ, Vázquez A, Menocal O, Carrillo D, Cloonan KR. Piperitone (p-Menth-1-En-3-One): A New Repellent for Tea Shot Hole Borer (Coleoptera: Curculionidae) in Florida Avocado Groves. Biomolecules. 2023; 13(4):656. https://doi.org/10.3390/biom13040656
Chicago/Turabian StyleKendra, Paul E., Wayne S. Montgomery, Nurhayat Tabanca, Elena Q. Schnell, Aimé Vázquez, Octavio Menocal, Daniel Carrillo, and Kevin R. Cloonan. 2023. "Piperitone (p-Menth-1-En-3-One): A New Repellent for Tea Shot Hole Borer (Coleoptera: Curculionidae) in Florida Avocado Groves" Biomolecules 13, no. 4: 656. https://doi.org/10.3390/biom13040656