
A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus galloprovincialis: An In Vitro Study


 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
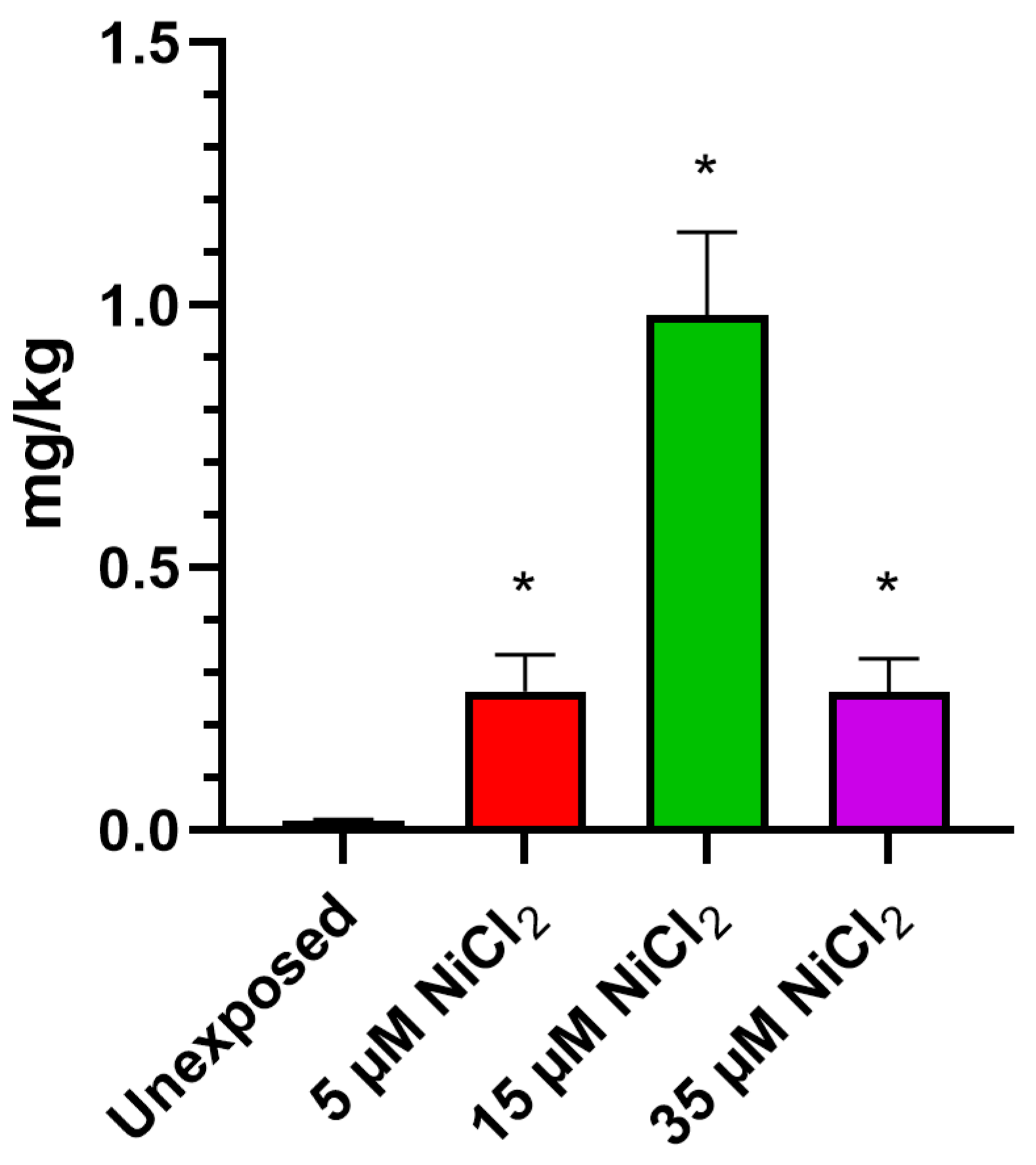
2.2. Bioaccumulation of Nickel
2.3. Exposure of Mussels to Nickel
2.4. Processing and Sampling of Spermatozoa
2.5. Extraction and Analysis of PL Proteins from M. galloprovincialis Spermatozoa
2.6. Electrophoretic Analysis
2.7. The Preparation and Analysis of Plasmid DNA
2.8. Evaluation of the Effect of PL Proteins from M. galloprovincialis on the Electrophoretic Mobility of DNA
2.9. Preparation of Sperm Nuclei and Salt-Induced M.galloprovincialis Sperm Nuclear Basic Protein Release
2.10. Micrococcal Nuclease Digestion
2.11. Homogenates’ Preparation from Mytilus galloprovincialis Male Gonads
2.12. Western Blotting of PARP
2.13. Statistical Analysis
3. Results
3.1. Bioaccumulation of Nickel
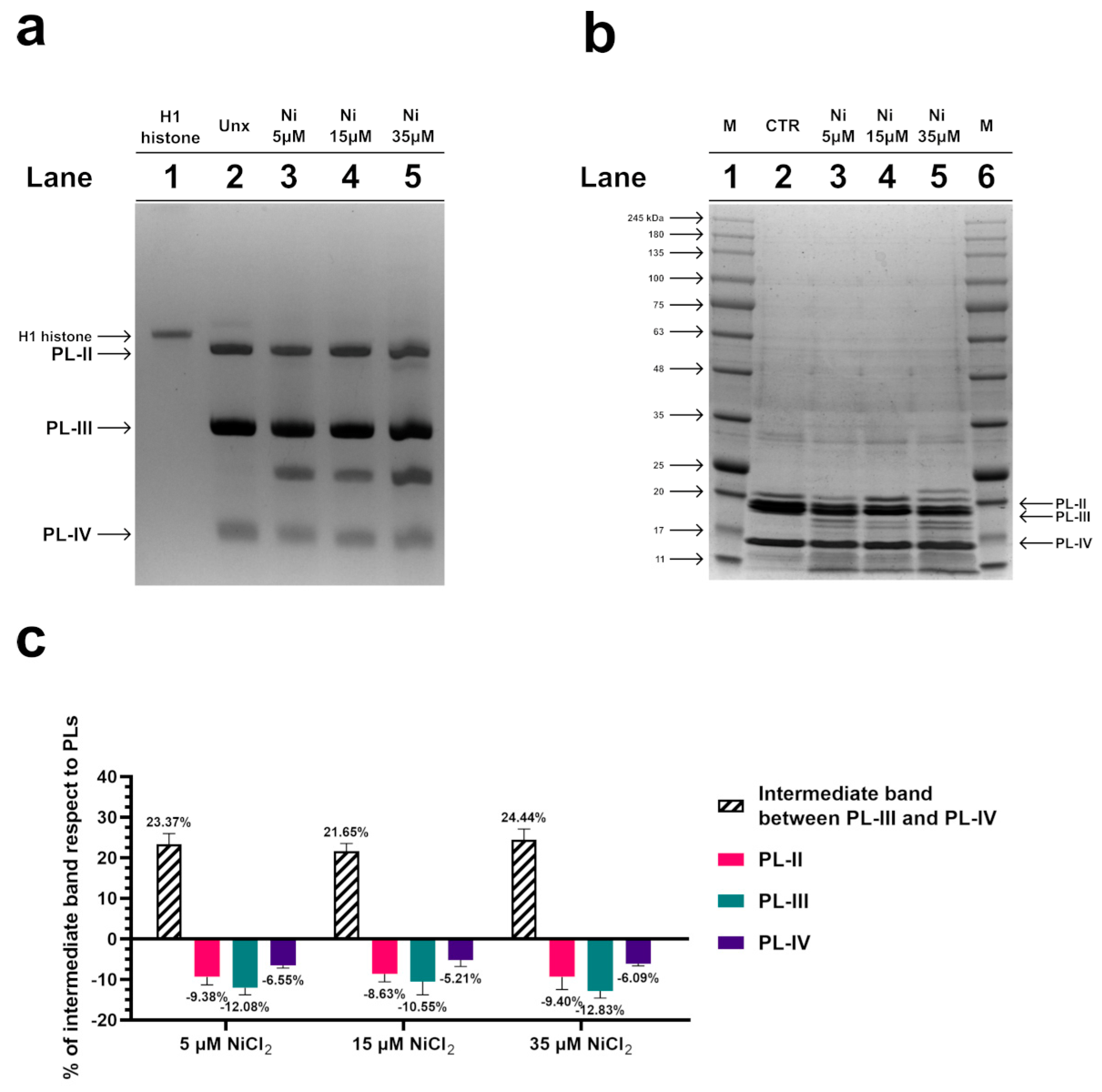
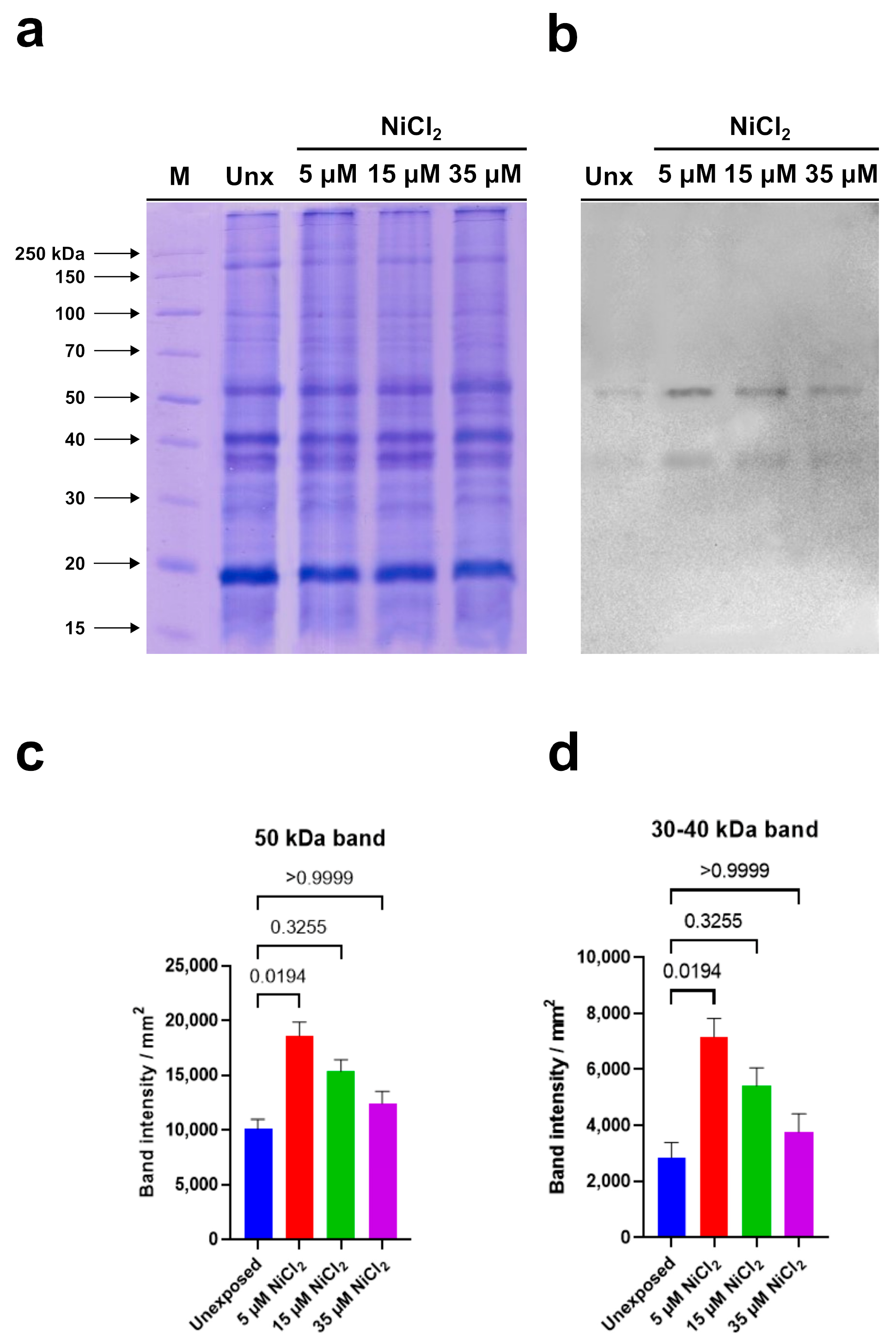
3.2. Electrophoretic Analyses of PL Proteins by AU-PAGE and SDS-PAGE
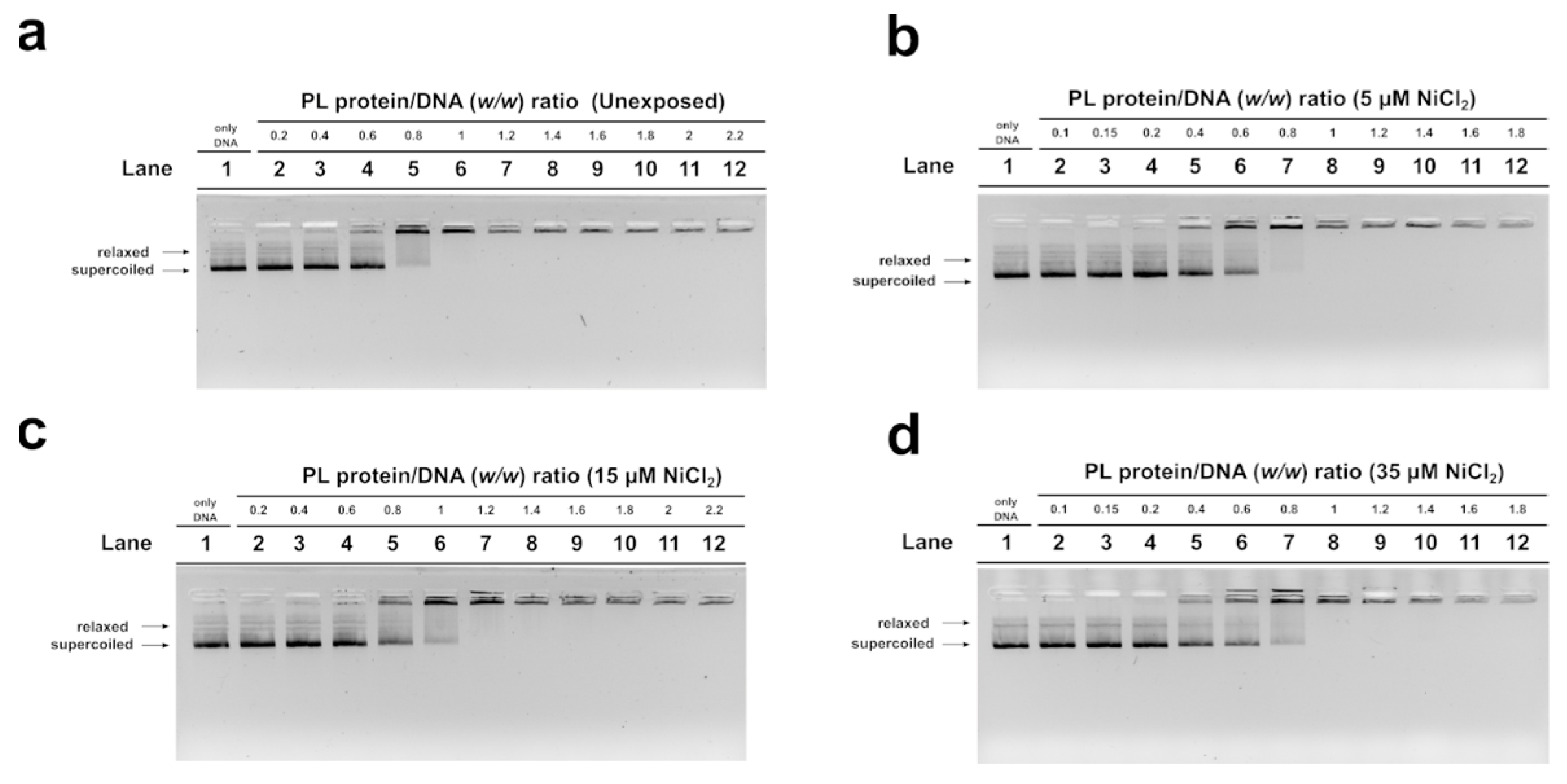
3.3. EMSA
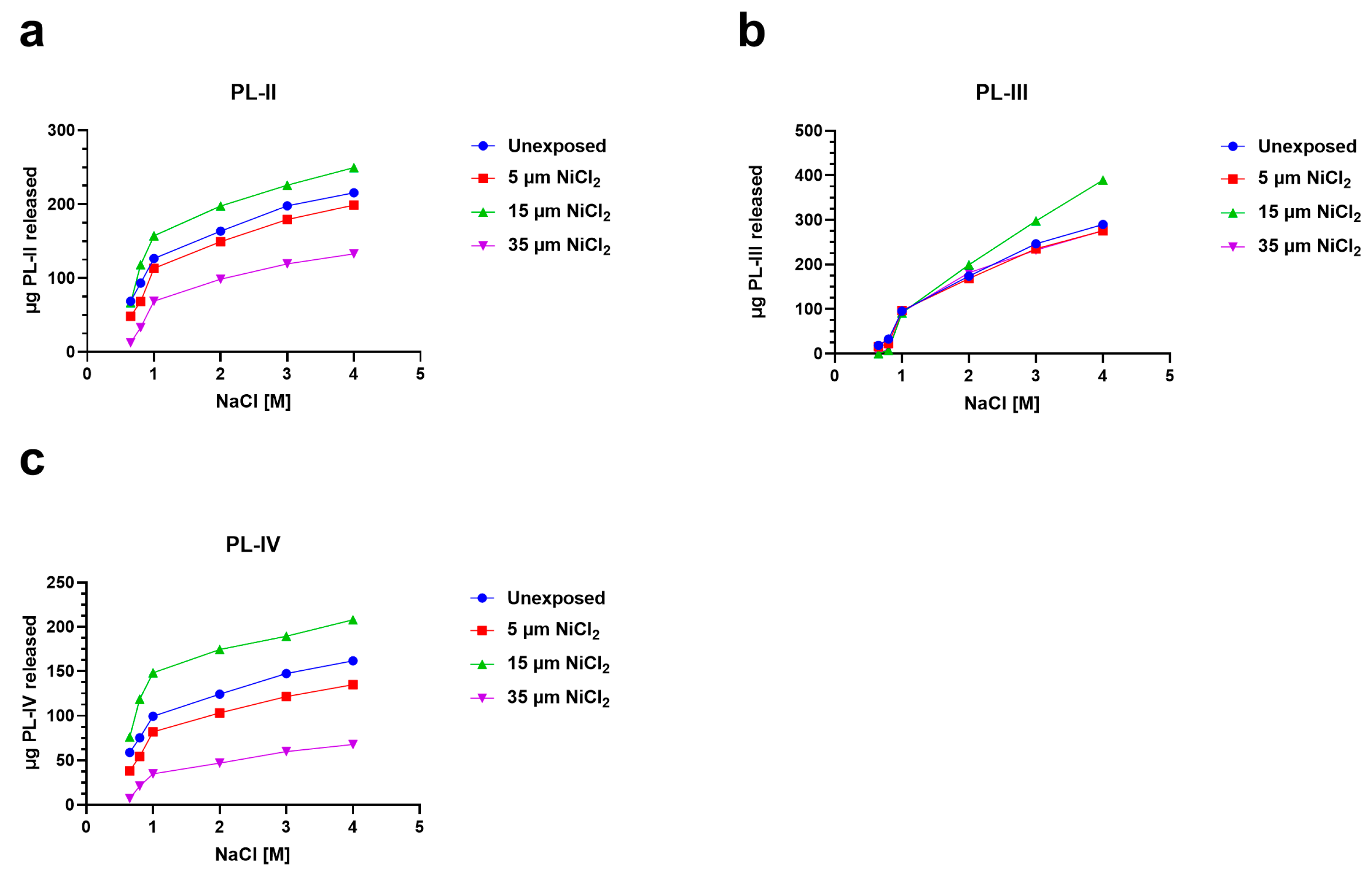
3.4. Release of PL Proteins from Sperm Nuclei
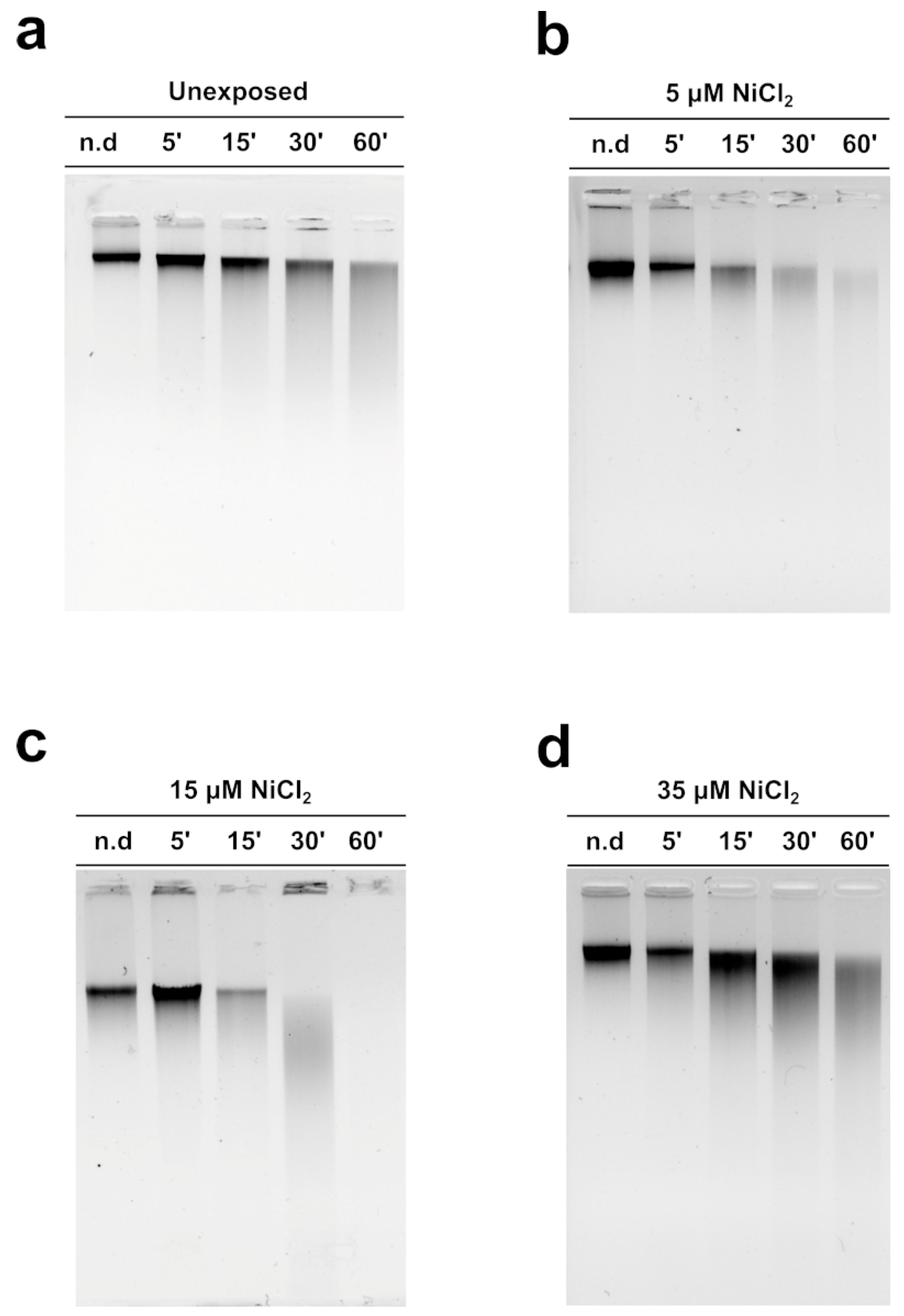
3.5. MNase Digestion Pattern of M. galloprovincialis Sperm Chromatin
3.6. PARP Expression
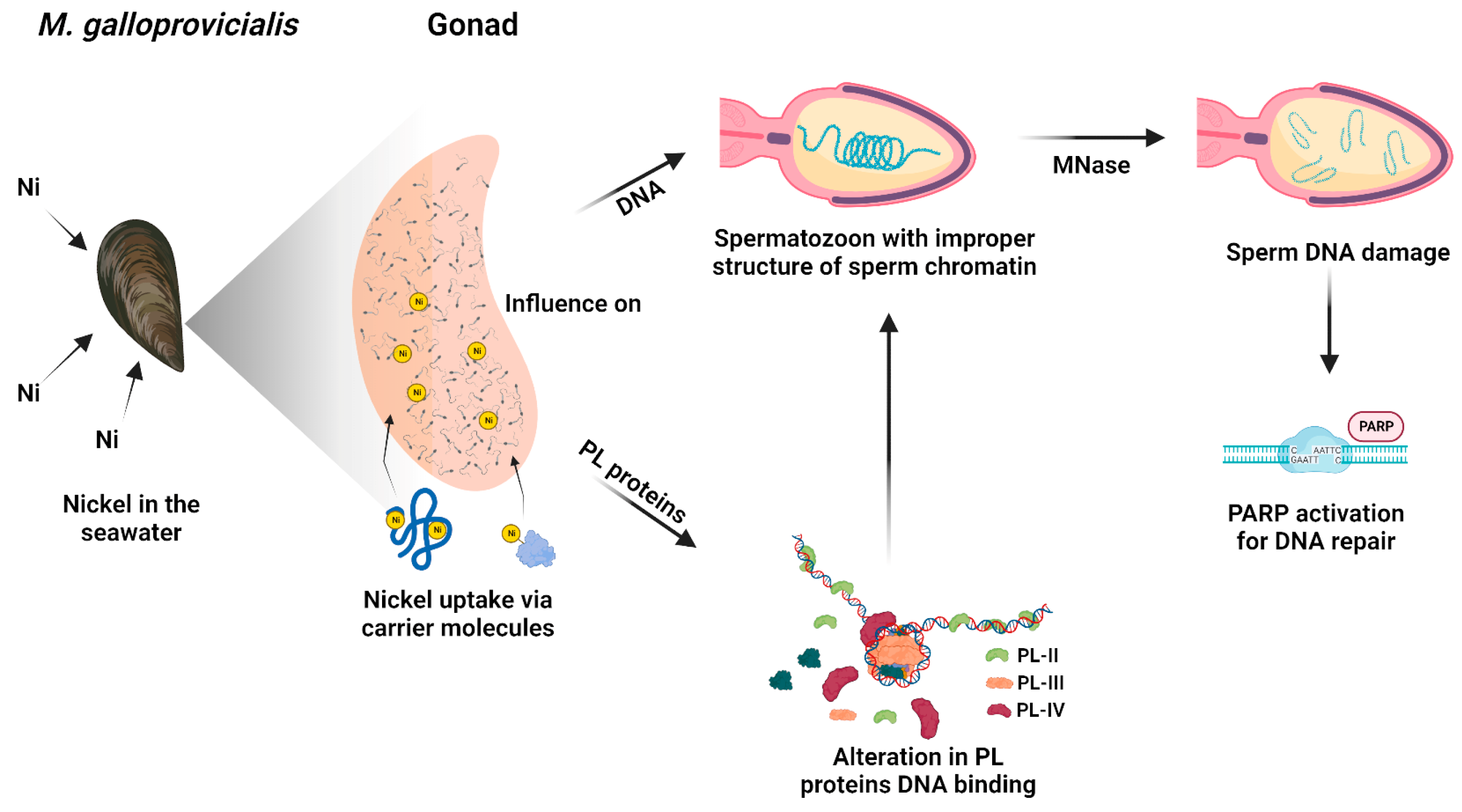
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gissi, F.; Stauber, J.L.; Binet, M.T.; Golding, L.A.; Adams, M.S.; Schlekat, C.E.; Garman, E.R.; Jolley, D.F. A Review of Nickel Toxicity to Marine and Estuarine Tropical Biota with Particular Reference to the South East Asian and Melanesian Region. Environ. Pollut. 2016, 218, 1308–1323. [Google Scholar] [CrossRef] [Green Version]
- Perrone, P.; Lettieri, G.; Marinaro, C.; Longo, V.; Capone, S.; Forleo, A.; Pappalardo, S.; Montano, L.; Piscopo, M. Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study. Int. J. Environ. Res. Public Health 2022, 19, 11023. [Google Scholar] [CrossRef]
- Peters, E.C.; Gassman, N.J.; Firman, J.C.; Richmond, R.H.; Power, E.A. Ecotoxicology of Tropical Marine Ecosystems. Environ. Toxicol. Chem. 1997, 16, 12–40. [Google Scholar] [CrossRef]
- Nunzio, A.D.; Giarra, A.; Toscanesi, M.; Amoresano, A.; Piscopo, M.; Ceretti, E.; Zani, C.; Lorenzetti, S.; Trifuoggi, M.; Montano, L. Comparison between Macro and Trace Element Concentrations in Human Semen and Blood Serum in Highly Polluted Areas in Italy. Int. J. Environ. Res. Public Health 2022, 19, 11635. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int. J. Mol. Sci. 2020, 21, 4198. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus Galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.; Singh, V.P. The Relative Impact of Toxic Heavy Metals (THMs) (Arsenic (As), Cadmium (Cd), Chromium (Cr)(VI), Mercury (Hg), and Lead (Pb)) on the Total Environment: An Overview. Environ. Monit. Assess 2019, 191, 419. [Google Scholar] [CrossRef]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A Review of Toxicity and Mechanisms of Individual and Mixtures of Heavy Metals in the Environment. Environ. Sci. Pollut. Res. 2016, 23, 8244–8259. [Google Scholar] [CrossRef]
- Rehman, K.; Fatima, F.; Waheed, I.; Akash, M.S.H. Prevalence of Exposure of Heavy Metals and Their Impact on Health Consequences. J. Cell. Biochem. 2018, 119, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Xi, S. The Effects of Heavy Metals on Human Metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Iamiceli, A.; Ubaldi, A.; Lucchetti, D.; Brambilla, G.; Abate, V.; De Felip, E.; De Filippis, S.P.; Dellatte, E.; De Luca, S.; Ferri, F.; et al. Metals in Mediterranean Aquatic Species. Mar. Pollut. Bull. 2015, 94, 278–283. [Google Scholar] [CrossRef]
- Ahlström, M.G.; Thyssen, J.P.; Wennervaldt, M.; Menné, T.; Johansen, J.D. Nickel Allergy and Allergic Contact Dermatitis: A Clinical Review of Immunology, Epidemiology, Exposure, and Treatment. Contact Dermat. 2019, 81, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Montano, L.; Pironti, C.; Pinto, G.; Ricciardi, M.; Buono, A.; Brogna, C.; Venier, M.; Piscopo, M.; Amoresano, A.; Motta, O. Polychlorinated Biphenyls (PCBs) in the Environment: Occupational and Exposure Events, Effects on Human Health and Fertility. Toxics 2022, 10, 365. [Google Scholar] [CrossRef]
- Maresca, V.; Lettieri, G.; Sorbo, S.; Piscopo, M.; Basile, A. Biological Responses to Cadmium Stress in Liverwort Conocephalum Conicum (Marchantiales). Int. J. Mol. Sci. 2020, 21, 6485. [Google Scholar] [CrossRef] [PubMed]
- Ahlström, M.G.; Thyssen, J.P.; Menné, T.; Johansen, J.D. Prevalence of Nickel Allergy in Europe Following the EU Nickel Directive—A Review. Contact Dermat. 2017, 77, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Buxton, S.; Garman, E.; Heim, K.E.; Lyons-Darden, T.; Schlekat, C.E.; Taylor, M.D.; Oller, A.R. Concise Review of Nickel Human Health Toxicology and Ecotoxicology. Inorganics 2019, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Pane, E.F.; Haque, A.; Wood, C.M. Mechanistic Analysis of Acute, Ni-Induced Respiratory Toxicity in the Rainbow Trout (Oncorhynchus Mykiss): An Exclusively Branchial Phenomenon. Aquat. Toxicol. 2004, 69, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Oller, A.R.; Kirkpatrick, D.T.; Radovsky, A.; Bates, H.K. Inhalation Carcinogenicity Study with Nickel Metal Powder in Wistar Rats. Toxicol. Appl. Pharmacol. 2008, 233, 262–275. [Google Scholar] [CrossRef]
- Khlifi, R.; Olmedo, P.; Gil, F.; Hammami, B.; Hamza-Chaffai, A. Cadmium and Nickel in Blood of Tunisian Population and Risk of Nasosinusal Polyposis Disease. Environ. Sci. Pollut. Res. 2015, 22, 3586–3593. [Google Scholar] [CrossRef] [PubMed]
- Bolek, E.; Erden, A.; Kulekci, C.; Kalyoncu, U.; Karadag, O. Rare Occupational Cause of Nasal Septum Perforation: Nickel Exposure. Int. J. Occup. Med. Environ. Health 2017, 30, 963–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewett, T.A.; Leonard, E.M. Mechanisms of Nickel Toxicity to Fish and Invertebrates in Marine and Estuarine Waters. Environ. Pollut. 2017, 223, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.C.; Pereira, M.L.; Sousa, J.P. Evaluation of Nickel Toxicity on Liver, Spleen, and Kidney of Mice after Administration of High-Dose Metal Ion. J. Biomed. Mater. Res. 1998, 40, 40–47. [Google Scholar] [CrossRef]
- Macomber, L.; Hausinger, R.P. Mechanisms of Nickel Toxicity in Microorganisms†. Metallomics 2011, 3, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Gong, C.; Ren, J.; Zhang, X.; Wang, G.; Liu, Y.; Ren, Y.; Zhao, Y.; Yu, Q.; Wang, Y.; et al. Toxicity of Nickel and Cobalt in Japanese Flounder. Environ. Pollut. 2020, 263, 114516. [Google Scholar] [CrossRef] [PubMed]
- David, M.; Qurat-Ul-Ain; Afzal, M.; Shoaib, M.; Aman, F.; Cloete, K.J.; Turi, N.; Jahan, S. Study of Occupational Exposure to Brick Kiln Emissions on Heavy Metal Burden, Biochemical Profile, Cortisol Level and Reproductive Health Risks among Female Workers at Rawat, Pakistan. Environ. Sci. Pollut. Res. 2020, 27, 44073–44088. [Google Scholar] [CrossRef]
- Lyu, K.; Wang, X.; Wang, L.; Wang, G. Rare-Earth Element Yttrium Enhances the Tolerance of Curly-Leaf Pondweed (Potamogeton Crispus) to Acute Nickel Toxicity. Environ. Pollut. 2019, 248, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, K.S.; Bal, W.; Karaczyn, A.A. The Role of Chromatin Damage in Nickel-Induced Carcinogenesis. A Review of Recent Developments. J. Environ. Monitor. 2003, 5, 183–187. [Google Scholar] [CrossRef]
- Shainkin-Kestenbaum, R.; Caruso, C.; Berlyne, G.M. Effect of Nickel on Oxygen Free Radical Metabolism. Inhibition of Superoxide Dismutase and Enhancement of Hydroxydopamine Autoxidation. Biol. Trace Elem. Res. 1991, 28, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B.; Uversky, V.N.; Ciurli, S. Nickel Impact on Human Health: An Intrinsic Disorder Perspective. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2016, 1864, 1714–1731. [Google Scholar] [CrossRef]
- Yang, J.; Ma, Z. Research Progress on the Effects of Nickel on Hormone Secretion in the Endocrine Axis and on Target Organs. Ecotoxicol. Environ. Saf. 2021, 213, 112034. [Google Scholar] [CrossRef]
- Djebbi, E.; Bonnet, D.; Pringault, O.; Tlili, K.; Yahia, M.N.D. Effects of Nickel Oxide Nanoparticles on Survival, Reproduction, and Oxidative Stress Biomarkers in the Marine Calanoid Copepod Centropages Ponticus under Short-Term Exposure. Environ. Sci. Pollut. Res. 2021, 28, 21978–21990. [Google Scholar] [CrossRef]
- Mohammed, E.H.; Wang, G.; Jiang, J. The Effects of Nickel on the Reproductive Ability of Three Different Marine Copepods. Ecotoxicology 2010, 19, 911–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Boni, R.; Buttino, I.; Tosti, E. Spermiotoxicity of Nickel Nanoparticles in the Marine Invertebrate Ciona Intestinalis (Ascidians). Nanotoxicology 2016, 10, 1096–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettieri, G.; Maione, M.; Ranauda, M.A.; Mele, E.; Piscopo, M. Molecular Effects on Spermatozoa of Mytilus Galloprovincialis Exposed to Hyposaline Conditions. Mol. Reprod. Dev. 2019, 86, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus Galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Notariale, R.; Carusone, N.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. New Insights into Alterations in PL Proteins Affecting Their Binding to DNA after Exposure of Mytilus Galloprovincialis to Mercury-A Possible Risk to Sperm Chromatin Structure? Int. J. Mol. Sci. 2021, 22, 5893. [Google Scholar] [CrossRef] [PubMed]
- Śmietanka, B.; Burzyński, A.; Hummel, H.; Wenne, R. Glacial History of the European Marine Mussels Mytilus, Inferred from Distribution of Mitochondrial DNA Lineages. Heredity 2014, 113, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Lassoued, J.; Padín, X.A.; Comeau, L.A.; Bejaoui, N.; Pérez, F.F.; Babarro, J.M.F. The Mediterranean Mussel Mytilus Galloprovincialis: Responses to Climate Change Scenarios as a Function of the Original Habitat. Conserv. Physiol. 2021, 9, coaa114. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2014; FAO Publisher: Rome, Italy, 2014; p. 223. ISBN 978-92-5-108276-8. Available online: www.fao.org/publications (accessed on 1 November 2022).
- Lewis, J.D.; Ausió, J. Protamine-like Proteins: Evidence for a Novel Chromatin Structure. Biochem. Cell Biol. 2002, 80, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Ausió, J. Histone H1 and Evolution of Sperm Nuclear Basic Proteins. J. Biol. Chem. 1999, 274, 31115–31118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piscopo, M.; Conte, M.; Di Paola, F.; Conforti, S.; Rana, G.; De Petrocellis, L.; Fucci, L.; Geraci, G. Relevance of Arginines in the Mode of Binding of H1 Histones to DNA. DNA Cell Biol. 2010, 29, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Fioretti, F.M.; Febbraio, F.; Carbone, A.; Branno, M.; Carratore, V.; Fucci, L.; Ausió, J.; Piscopo, M. A Sperm Nuclear Basic Protein from the Sperm of the Marine Worm Chaetopterus Variopedatus with Sequence Similarity to the Arginine-Rich C-Termini of Chordate Protamine-Likes. DNA Cell Biol. 2012, 31, 1392–1402. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus Galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, Q.A.; Caccavale, F.; Avagnano, S.; Murolo, A.; Guerriero, G.; Fucci, L.; Ausió, J.; Piscopo, M. New Insights into Protamine-like Component Organization in Mytilus Galloprovincialis’ Sperm Chromatin. DNA Cell Biol. 2015, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Middag, R.; Rolison, J.M.; George, E.; Gerringa, L.J.A.; Rijkenberg, M.J.A.; Stirling, C.H. Basin Scale Distributions of Dissolved Manganese, Nickel, Zinc and Cadmium in the Mediterranean Sea. Mar. Chem. 2022, 238, 104063. [Google Scholar] [CrossRef]
- Piscopo, M.; Ricciardiello, M.; Palumbo, G.; Troisi, J. Selectivity of Metal Bioaccumulation and Its Relationship with Glutathione S-Transferase Levels in Gonadal and Gill Tissues of Mytilus Galloprovincialis Exposed to Ni (II), Cu (II) and Cd (II). Rend. Fis. Acc. Lincei 2016, 27, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Notariale, R.; Basile, A.; Montana, E.; Romano, N.C.; Cacciapuoti, M.G.; Aliberti, F.; Gesuele, R.; Ruberto, F.D.; Sorbo, S.; Tenore, G.C.; et al. Protamine-like Proteins Have Bactericidal Activity. The First Evidence in Mytilus Galloprovincialis. Acta Biochim. Pol. 2018, 65, 585–594. [Google Scholar] [CrossRef]
- Piscopo, M.; Trifuoggi, M.; Notariale, R.; Labar, S.; Troisi, J.; Giarra, A.; Rabbito, D.; Puoti, R.; Brundo, M.V.; Basile, A.; et al. Protamine-like Proteins Analyses as Emerging Biotechnique for Cadmium Impact Assessment on Male Mollusk Mytilus Galloprovincialis (Lamarck 1819). Acta Biochim. Pol. 2018, 65, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Fioretti, F.M.; Fucci, L.; Ausió, J.; Piscopo, M. High Efficiency Method to Obtain Supercoiled DNA with a Commercial Plasmid Purification Kit. Acta Biochim. Pol. 2012, 59, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Olivares, C.; Ruiz, S. Nucleosomal Organization of Chromatin in Sperm Nuclei of the Bivalve Mollusc Aulacomya Ater. Mol. Cell Biochem. 1991, 101, 93–99. [Google Scholar] [CrossRef]
- De Guglielmo, V.; Puoti, R.; Notariale, R.; Maresca, V.; Ausió, J.; Troisi, J.; Verrillo, M.; Basile, A.; Febbraio, F.; Piscopo, M. Alterations in the Properties of Sperm Protamine-like II Protein after Exposure of Mytilus Galloprovincialis (Lamarck 1819) to Sub-Toxic Doses of Cadmium. Ecotoxicol. Environ. Saf. 2019, 169, 600–606. [Google Scholar] [CrossRef]
- Capriello, T.; Di Meglio, G.; De Maio, A.; Scudiero, R.; Bianchi, A.R.; Trifuoggi, M.; Toscanesi, M.; Giarra, A.; Ferrandino, I. Aluminium Exposure Leads to Neurodegeneration and Alters the Expression of Marker Genes Involved to Parkinsonism in Zebrafish Brain. Chemosphere 2022, 307, 135752. [Google Scholar] [CrossRef]
- Vlahogianni, T.; Dassenakis, M.; Scoullos, M.J.; Valavanidis, A. Integrated Use of Biomarkers (Superoxide Dismutase, Catalase and Lipid Peroxidation) in Mussels Mytilus Galloprovincialis for Assessing Heavy Metals’ Pollution in Coastal Areas from the Saronikos Gulf of Greece. Mar. Pollut. Bull. 2007, 54, 1361–1371. [Google Scholar] [CrossRef]
- De Maio, A.; Porzio, E.; Rotondo, S.; Bianchi, A.R.; Faraone-Mennella, M.R. In Sulfolobus Solfataricus, the Poly(ADP-Ribose) Polymerase-Like Thermoprotein Is a Multifunctional Enzyme. Microorganisms 2020, 8, 1523. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Brundo, M.V.; Labar, S.; Bianchi, A.R.; Trocchia, S.; Rabbito, D.; Palumbo, G.; Abdel-Gawad, F.K.; De Maio, A. Frog (Pelophylax Bergeri, Günther 1986) Endocrine Disruption Assessment: Characterization and Role of Skin Poly(ADP-Ribose) Polymerases. Environ. Sci. Pollut. Res. 2018, 25, 18303–18313. [Google Scholar] [CrossRef]
- Longo, V.; Forleo, A.; Radogna, A.V.; Siciliano, P.; Notari, T.; Pappalardo, S.; Piscopo, M.; Montano, L.; Capone, S. A Novel Human Biomonitoring Study by Semiconductor Gas Sensors in Exposomics: Investigation of Health Risk in Contaminated Sites. Environ. Pollut. 2022, 304, 119119. [Google Scholar] [CrossRef]
- Maresca, V.; Fusaro, L.; Sorbo, S.; Siciliano, A.; Loppi, S.; Paoli, L.; Monaci, F.; Karam, E.A.; Piscopo, M.; Guida, M.; et al. Functional and Structural Biomarkers to Monitor Heavy Metal Pollution of One of the Most Contaminated Freshwater Sites in Southern Europe. Ecotoxicol. Environ. Saf. 2018, 163, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Basile, A.; Loppi, S.; Piscopo, M.; Paoli, L.; Vannini, A.; Monaci, F.; Sorbo, S.; Lentini, M.; Esposito, S. The Biological Response Chain to Pollution: A Case Study from the “Italian Triangle of Death” Assessed with the Liverwort Lunularia Cruciata. Environ. Sci. Pollut. Res. 2017, 24, 26185–26193. [Google Scholar] [CrossRef]
- Cameron, K.S.; Buchner, V.; Tchounwou, P.B. Exploring the Molecular Mechanisms of Nickel-Induced Genotoxicity and Carcinogenicity: A Literature Review. Rev. Environ. Health 2011, 26, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Forgacs, Z.; Massányi, P.; Lukac, N.; Somosy, Z. Reproductive Toxicology of Nickel—Review. J. Environ. Sci. Health 2012, 47, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Kopera, E.; Krȩżel, A.; Protas, A.M.; Belczyk, A.; Bonna, A.; Wysłouch-Cieszyńska, A.; Poznański, J.; Bal, W. Sequence-Specific Ni(II)-Dependent Peptide Bond Hydrolysis for Protein Engineering: Reaction Conditions and Molecular Mechanism. Inorg. Chem. 2010, 49, 6636–6645. [Google Scholar] [CrossRef] [PubMed]
- Tesei, G.; Vazdar, M.; Jensen, M.R.; Cragnell, C.; Mason, P.E.; Heyda, J.; Skepö, M.; Jungwirth, P.; Lund, M. Self-Association of a Highly Charged Arginine-Rich Cell-Penetrating Peptide. Proc. Natl. Acad. Sci. USA 2017, 114, 11428–11433. [Google Scholar] [CrossRef] [Green Version]
- Argudo, P.G.; Giner-Casares, J.J. Folding and Self-Assembly of Short Intrinsically Disordered Peptides and Protein Regions. Nanoscale Adv. 2021, 3, 1789–1812. [Google Scholar] [CrossRef] [PubMed]
- Salvati, D.; Conforti, S.; Conte, M.; Matassa, D.S.; Fucci, L.; Piscopo, M. Self-Association of Chaetopterus Variopedatus Sperm Histone H1-like. Relevance of Arginine Content and Possible Physiological Role. Acta Biochim. Pol. 2008, 55, 701–706. [Google Scholar] [CrossRef]
- Moriello, C.; Costabile, M.; Spinelli, M.; Amoresano, A.; Palumbo, G.; Febbraio, F.; Piscopo, M. Altered Expression of Protamine-like and Their DNA Binding Induced by Cr(VI): A Possible Risk to Spermatogenesis? Biomolecules 2022, 12, 700. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Kraus, W.L. The PARP Side of the Nucleus: Molecular Actions, Physiological Outcomes, and Clinical Targets. Mol. Cell 2010, 39, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.P.; Pereira Bastos de Siqueira, M.E.; Sérgio da Silva, C. Urinary Nickel as Bioindicator of Workers’ Ni Exposure in a Galvanizing Plant in Brazil. Int. Arch. Occup. Environ. Health 2000, 73, 65–68. [Google Scholar] [CrossRef]
- Murawska-Ciałowicz, E.; Bal, W.; Januszewska, L.; Zawadzki, M.; Rychel, J.; Zuwała-Jagiełło, J. Oxidative Stress Level in the Testes of Mice and Rats during Nickel Intoxication. Sci. World J. 2012, 2012, 395741. [Google Scholar] [CrossRef] [Green Version]
- Menezo, Y.; Russo, G.; Tosti, E.; El Mouatassim, S.; Benkhalifa, M. Expression Profile of Genes Coding for DNA Repair in Human Oocytes Using Pangenomic Microarrays, with a Special Focus on ROS Linked Decays. J. Assist. Reprod. Genet. 2007, 24, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Ménézo, Y.; Dale, B.; Cohen, M. DNA Damage and Repair in Human Oocytes and Embryos: A Review. Zygote 2010, 18, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. The Amoroso Lecture. The Human Spermatozoon—A Cell in Crisis? J. Reprod. Fertil. 1999, 115, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dunn’s Multiple Comparisons Test | Significant? | Summary | Adjusted p Value |
|---|---|---|---|
| Unexposed vs. 5 µM NiCl2 | Yes | * | 0.0194 |
| Unexposed vs. 15 µM NiCl2 | No | ns | 0.3255 |
| Unexposed vs. 35 µM NiCl2 | No | ns | >0.9999 |
| 5 µM NiCl2 vs. 15 µM NiCl2 | No | ns | >0.9999 |
| 5 µM NiCl2 vs. 35 µM NiCl2 | No | ns | 0.1887 |
| 15 µM NiCl2 vs. 35 µM NiCl2 | No | ns | >0.9999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbone, G.; Lettieri, G.; Marinaro, C.; Costabile, M.; Notariale, R.; Bianchi, A.R.; De Maio, A.; Piscopo, M. A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus galloprovincialis: An In Vitro Study. Biomolecules 2023, 13, 520. https://doi.org/10.3390/biom13030520
Carbone G, Lettieri G, Marinaro C, Costabile M, Notariale R, Bianchi AR, De Maio A, Piscopo M. A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus galloprovincialis: An In Vitro Study. Biomolecules. 2023; 13(3):520. https://doi.org/10.3390/biom13030520
Chicago/Turabian StyleCarbone, Gelsomina, Gennaro Lettieri, Carmela Marinaro, Martina Costabile, Rosaria Notariale, Anna Rita Bianchi, Anna De Maio, and Marina Piscopo. 2023. "A Molecular Mechanism to Explain the Nickel-Induced Changes in Protamine-like Proteins and Their DNA Binding Affecting Sperm Chromatin in Mytilus galloprovincialis: An In Vitro Study" Biomolecules 13, no. 3: 520. https://doi.org/10.3390/biom13030520