
Osteopontin: A Bone-Derived Protein Involved in Rheumatoid Arthritis and Osteoarthritis Immunopathology
 , , , , ,
, , , , ,
Abstract
:1. Introduction
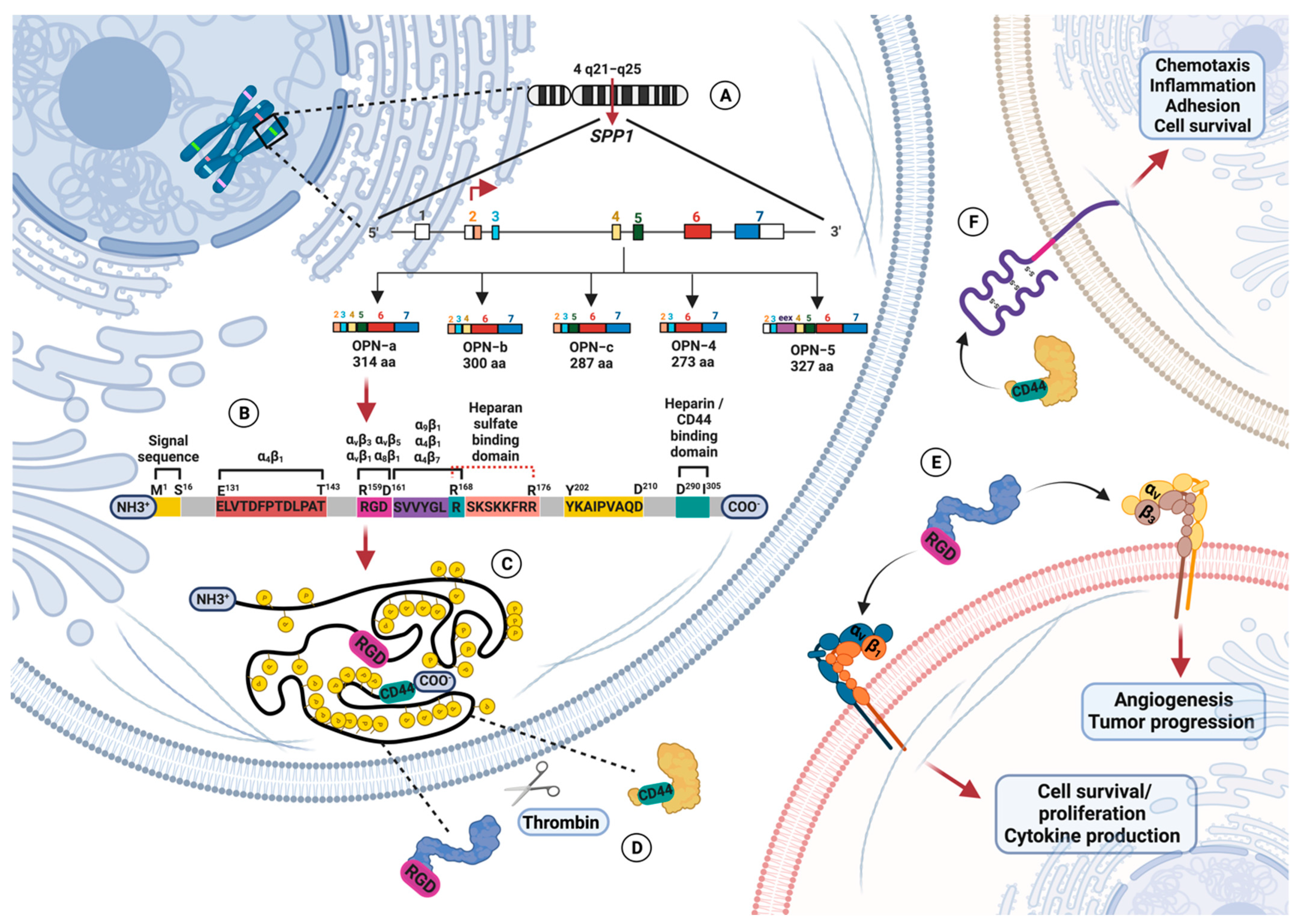
1.1. Osteopontin Gene and Protein Structure
1.2. Osteopontin Adhesion Motifs and Posttranslational Modifications
1.3. Osteopontin Mediates the Immune Response and Inflammation
1.4. Osteopontin Involvement in Inflammatory Diseases
2. A Brief Look at the Basic Immunopathology of Rheumatoid Arthritis and Osteoarthritis
3. Osteopontin: Could It Be Considered a Common Denominator in Rheumatoid Arthritis and Osteoarthritis Immunopathology?
3.1. Soluble Osteopontin Levels in Rheumatoid Arthritis and Osteoarthritis Patients
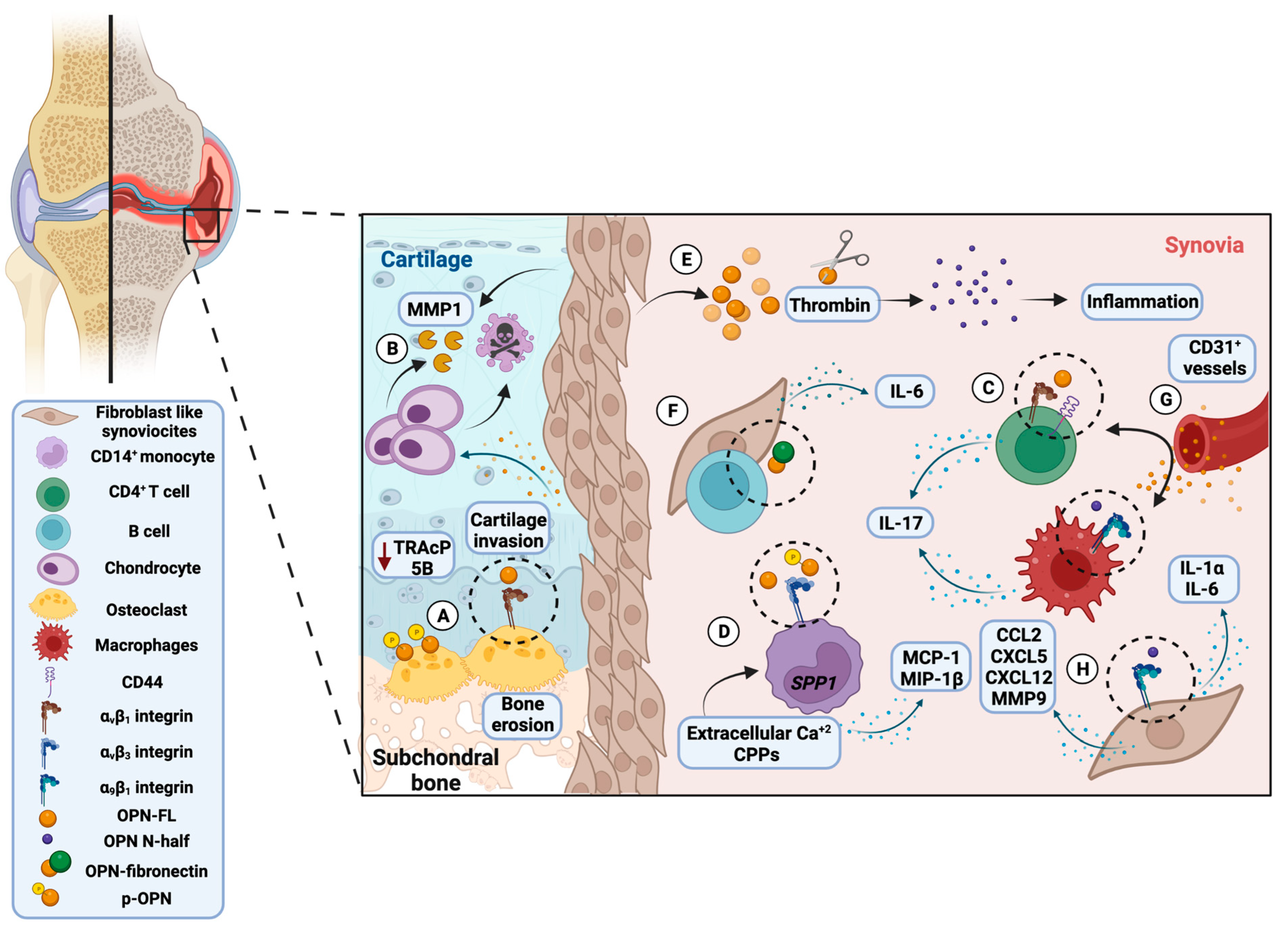
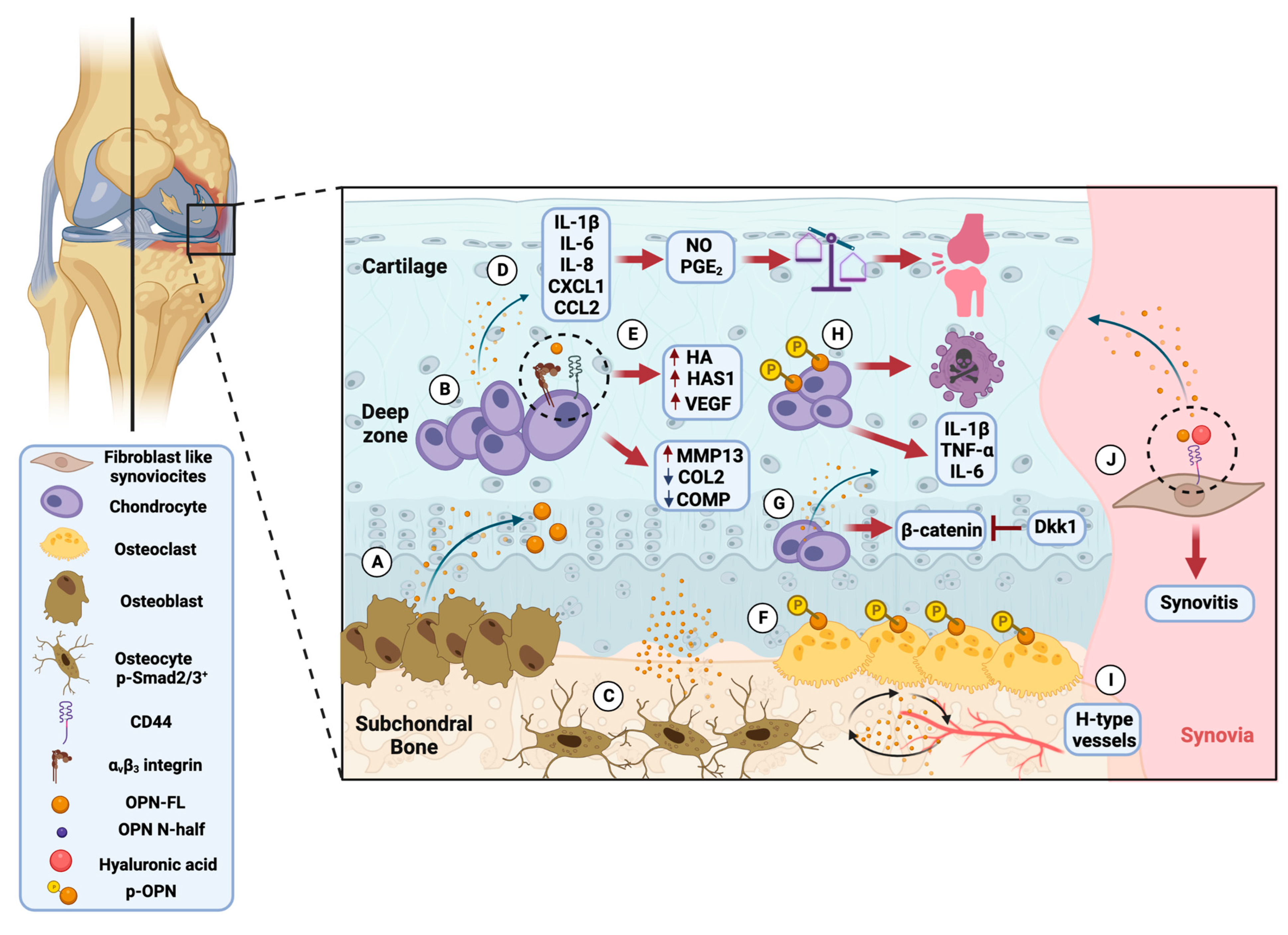
3.2. Proposed Molecular Mechanism of OPN in Rheumatoid Arthritis and Osteoarthritis
3.3. SPP1 Genetic Variations Associated with Rheumatoid Arthritis and Osteoarthritis
3.4. In Vitro Studies Reveal the Crucial Role of Osteopontin in Rheumatoid Arthritis and Osteoarthritis Pathology
3.5. Role of Osteopontin in Experimental Models of Rheumatoid Arthritis and Osteoarthritis
4. Osteopontin Regulation by MicroRNA in Rheumatoid Arthritis and Osteoarthritis
Are They Blocking Osteopontin Activity: An Alternative for Rheumatoid Arthritis and Osteoarthritis Treatment?
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bandopadhyay, M.; Bulbule, A.; Butti, R.; Chakraborty, G.; Ghorpade, P.; Ghosh, P.; Gorain, M.; Kale, S.; Kumar, D.; Kumar, S.; et al. Osteopontin as a therapeutic target for cancer. Expert Opin. Ther. Targets 2014, 18, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Kahles, F.; Findeisen, H.M.; Bruemmer, D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol. Metab. 2014, 3, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Franzen, A.; Heinegård, D. Isolation and characterization of two sialoproteins present only in bone calcified matrix. Biochem. J. 1985, 232, 715–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldberg, A.; Franzén, A.; Heinegård, D. Cloning and sequence analysis of rat bone sialoprotein (osteopontin) cDNA reveals an Arg-Gly-Asp cell-binding sequence. Proc. Natl. Acad. Sci. USA 1986, 83, 8819–8823. [Google Scholar] [CrossRef] [Green Version]
- Reinholt, F.P.; Hultenby, K.; Oldberg, A.; Heinegård, D. Osteopontin—A possible anchor of osteoclasts to bone. Proc. Natl. Acad. Sci. USA 1990, 87, 4473–4475. [Google Scholar] [CrossRef] [Green Version]
- Karpinsky, G.; Fatyga, A.; Krawczyk, M.A.; Chamera, M.; Sande, N.; Szmyd, D.; Izycka-Swieszewska, E.; Bien, E. Osteopontin: Its potential role in cancer of children and young adults. Biomark. Med. 2017, 11, 389–402. [Google Scholar] [CrossRef]
- Xie, Y.; Sakatsume, M.; Nishi, S.; Narita, I.; Arakawa, M.; Gejyo, F. Expression, roles, receptors, and regulation of osteopontin in the kidney. Kidney Int. 2001, 60, 1645–1657. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Liu, Y.-J. Opn: Key regulator of pDC interferon production. Nat. Immunol. 2006, 7, 441–443. [Google Scholar] [CrossRef]
- Hasegawa, M.; Nakoshi, Y.; Iino, T.; Sudo, A.; Segawa, T.; Maeda, M.; Yoshida, T.; Uchida, A. Thrombin-Cleaved Osteopontin in Synovial Fluid of Subjects with Rheumatoid Arthritis. J. Rheumatol. 2009, 36, 240–245. [Google Scholar] [CrossRef]
- Anborgh, P.H.; Mutrie, J.C.; Tuck, A.B.; Chambers, A.F. Pre- and post-translational regulation of osteopontin in cancer. J. Cell Commun. Signal. 2011, 5, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Giacopelli, F.; Marciano, R.; Pistorio, A.; Catarsi, P.; Canini, S.; Karsenty, G.; Ravazzolo, R. Polymorphisms in the osteopontin promoter affect its transcriptional activity. Physiol. Genom. 2004, 20, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staines, K.A.; MacRae, V.E.; Farquharson, C. The importance of the SIBLING family of proteins on skeletal mineralisation and bone remodelling. J. Endocrinol. 2012, 214, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.-X.; Li, Z.J.; Jiang, X.O.; Lum, Y.L.; Khin, E.; Lee, N.P.; Wu, G.H.; Luk, J.M. Osteopontin as potential biomarker and therapeutic target in gastric and liver cancers. World J. Gastroenterol. 2012, 18, 3923–3930. [Google Scholar] [CrossRef] [Green Version]
- Castello, L.M.; Raineri, D.; Salmi, L.; Clemente, N.; Vaschetto, R.; Quaglia, M.; Garzaro, M.; Gentilli, S.; Navalesi, P.; Cantaluppi, V.; et al. Osteopontin at the Crossroads of Inflammation and Tumor Progression. Mediat. Inflamm. 2017, 2017, 4049098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eremo, A.G.; Lagergren, K.; Othman, L.; Montgomery, S.; Andersson, G.; Tina, E. Evaluation of SPP1/osteopontin expression as predictor of recurrence in tamoxifen treated breast cancer. Sci. Rep. 2020, 10, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimba, E.R.; Brum, M.C.; de Moraes, G.N. Full-length osteopontin and its splice variants as modulators of chemoresistance and radioresistance (Review). Int. J. Oncol. 2019, 54, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimba, E.; Tilli, T. Human osteopontin splicing isoforms: Known roles, potential clinical applications and activated signaling pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef]
- Weng, S.; Zhou, L.; Han, L.; Yuan, Y. Expression and purification of non-tagged recombinant mouse SPP1 in E. Coli and its biological significance. Bioengineered 2014, 5, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Kamalabadi-Farahani, M.; Atashi, A.; Jabbarpour, Z.; Aghayan, S.S. Expression of osteopontin-5 splice variant in the mouse primary and metastatic breast cancer cells. BMC Res. Notes 2022, 15, 286. [Google Scholar] [CrossRef]
- Mateos, B.; Holzinger, J.; Conrad-Billroth, C.; Platzer, G.; Żerko, S.; Sealey-Cardona, M.; Anrather, D.; Koźmiński, W.; Konrat, R. Hyperphosphorylation of Human Osteopontin and Its Impact on Structural Dynamics and Molecular Recognition. Biochemistry 2021, 60, 1347–1355. [Google Scholar] [CrossRef]
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal. 2009, 3, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Morser, J.; Leung, L.L. Thrombin Cleavage of Osteopontin Disrupts a Pro-chemotactic Sequence for Dendritic Cells, Which Is Compensated by the Release of Its Pro-chemotactic C-terminal Fragment. J. Biol. Chem. 2014, 289, 27146–27158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaw, L.; Lindner, V.; Schwartz, S.M.; Chambers, A.F.; Giachelli, C.M. Osteopontin and β 3 Integrin Are Coordinately Expressed in Regenerating Endothelium In Vivo and Stimulate Arg-Gly-Asp–Dependent Endothelial Migration In Vitro. Circ. Res. 1995, 77, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Schnapp, L.M.; Hatch, N.; Ramos, D.M.; Klimanskaya, I.V.; Sheppard, D.; Pytela, R. The Human Integrin α8β1 Functions as a Receptor for Tenascin, Fibronectin, and Vitronectin. J. Biol. Chem. 1995, 270, 23196–23202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.D.; Lin, E.C.; Kovach, N.L.; Hoyer, J.R.; Smith, J.W. A Biochemical Characterization of the Binding of Osteopontin to Integrins αvβ1 and αvβ5. J. Biol. Chem. 1995, 270, 26232–26238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faccio, R.; Grano, M.; Colucci, S.; Zallone, A.; Quaranta, V.; Pelletier, A. Activation of αvβ3Integrin on Human Osteoclast-like Cells Stimulates Adhesion and Migration in Response to Osteopontin. Biochem. Biophys. Res. Commun. 1998, 249, 522–525. [Google Scholar] [CrossRef]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of osteopontin signaling and function by post-translational phosphorylation and protein folding. J. Cell. Biochem. 2007, 102, 912–924. [Google Scholar] [CrossRef]
- Smith, L.L.; Cheung, H.K.; Ling, L.E.; Chen, J.; Sheppard, D.; Pytela, R.; Giachelli, C.M. Osteopontin N-terminal domain contains a cryptic adhesive sequence recognized by alpha9beta1 integrin. J. Biol. Chem. 1996, 271, 28485–28491. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L.; Giachelli, C.M. Structural Requirements for α9β1-Mediated Adhesion and Migration to Thrombin-Cleaved Osteopontin. Exp. Cell Res. 1998, 242, 351–360. [Google Scholar] [CrossRef]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The Integrin α9β1 Binds to a Novel Recognition Sequence (SVVYGLR) in the Thrombin-cleaved Amino-terminal Fragment of Osteopontin. J. Biol. Chem. 1999, 274, 36328–36334. [Google Scholar] [CrossRef] [Green Version]
- Barry, S.T.; Ludbrook, S.B.; Murrison, E.; Horgan, C.M. Analysis of the α4β1 Integrin–Osteopontin Interaction. Exp. Cell Res. 2000, 258, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Bayless, K.; Meininger, G.; Scholtz, J.; Davis, G. Osteopontin is a ligand for the alpha4beta1 integrin. J. Cell Sci. 1998, 111 Pt 9, 1165–1174. [Google Scholar] [CrossRef]
- Bayless, K.J.; Davis, G.E. Identification of Dual α4β1 Integrin Binding Sites within a 38 Amino Acid Domain in the N-terminal Thrombin Fragment of Human Osteopontin. J. Biol. Chem. 2001, 276, 13483–13489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, P.M.; Ludbrook, S.B.; Miller, D.D.; Horgan, C.M.; Barry, S.T. Structural elements of the osteopontin SVVYGLR motif important for the interaction with α4 integrins. FEBS Lett. 2001, 503, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchinaka, A.; Hamada, Y.; Mori, S.; Miyagawa, S.; Saito, A.; Sawa, Y.; Matsuura, N.; Yamamoto, H.; Kawaguchi, N. SVVYGLR motif of the thrombin-cleaved N-terminal osteopontin fragment enhances the synthesis of collagen type III in myocardial fibrosis. Mol. Cell. Biochem. 2015, 408, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Shao, Z.; Sharif, S.; Du, X.-Y.; Myles, T.; Merchant, M.; Harsh, G.; Glantz, M.; Recht, L.; Morser, J.; et al. Thrombin-cleaved Fragments of Osteopontin Are Overexpressed in Malignant Glial Tumors and Provide a Molecular Niche with Survival Advantage. J. Biol. Chem. 2013, 288, 3097–3111. [Google Scholar] [CrossRef] [Green Version]
- Scatena, M.; Liaw, L.; Giachelli, C.M. Osteopontin: A multifunctional molecule regulating chronic inflammation and vascular disease. Arter. Thromb. Vasc. Biol. 2007, 27, 2302–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, S.A.; Du, X.; Myles, T.; Song, J.J.; Price, E.; Lee, D.M.; Goodman, S.B.; Nagashima, M.; Morser, J.; Robinson, W.H.; et al. Thrombin-activatable carboxypeptidase B cleavage of osteopontin regulates neutrophil survival and synoviocyte binding in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 2902–2912. [Google Scholar] [CrossRef] [Green Version]
- LaFoya, B.; Munroe, J.A.; Miyamoto, A.; Detweiler, M.A.; Crow, J.J.; Gazdik, T.; Albig, A.R. Beyond the Matrix: The Many Non-ECM Ligands for Integrins. Int. J. Mol. Sci. 2018, 19, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, L.R.; Teixeira, J.A.; Schmitt, F.L.; Paulsson, M.; Lindmark-Mansson, H. The Role of Osteopontin in Tumor Progression and Metastasis in Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1087–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-Ligand Interaction Between CD44 and Osteopontin (Eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-H.; Huang, C.-J.; Chao, J.-R.; Chen, S.-T.; Lee, S.-F.; Yen, J.J.-Y.; Yang-Yen, H.-F. Coupling of Osteopontin and Its Cell Surface Receptor CD44 to the Cell Survival Response Elicited by Interleukin-3 or Granulocyte-Macrophage Colony-Stimulating Factor. Mol. Cell. Biol. 2000, 20, 2734–2742. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F.; Zawaideh, S.; Hikita, S.; Kumar, V.A.; Cantor, H.; Ashkar, S. Phosphorylation-dependent interaction of osteopontin with its receptors regulates macrophage migration and activation. J. Leukoc. Biol. 2002, 72, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Crispín, J.C.; Keenan, B.T.; Finnell, M.D.; Bermas, B.L.; Schur, P.; Massarotti, E.; Karlson, E.W.; Fitzgerald, L.M.; Ergin, S.; Kyttaris, V.C.; et al. Expression of CD44 variant isoforms CD44v3 and CD44v6 is increased on T cells from patients with systemic lupus erythematosus and is correlated with disease activity. Arthritis Rheum. 2010, 62, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.X.; Denhardt, D.T. Osteopontin: Role in immune regulation and stress responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Rangaswami, H.; Bulbule, A.; Kundu, G.C. Osteopontin: Role in cell signaling and cancer progression. Trends Cell Biol. 2006, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 356. [Google Scholar] [CrossRef]
- Chakraborty, G.; Jain, S.; Behera, R.; Ahmed, M.; Sharma, P.; Kumar, V.; Kundu, G.C. The multifaceted roles of osteopontin in cell signaling, tumor progression and angiogenesis. Curr. Mol. Med. 2006, 6, 819–830. [Google Scholar] [CrossRef]
- Shinohara, M.L.; Jansson, M.; Hwang, E.S.; Werneck, M.B.F.; Glimcher, L.H.; Cantor, H. T-bet-dependent expression of osteopontin contributes to T cell polarization. Proc. Natl. Acad. Sci. USA 2005, 102, 17101–17106. [Google Scholar] [CrossRef] [Green Version]
- Bruemmer, D.; Collins, A.R.; Noh, G.; Wang, W.; Territo, M.; Arias-Magallona, S.; Fishbein, M.C.; Blaschke, F.; Kintscher, U.; Graf, K.; et al. Angiotensin II–accelerated atherosclerosis and aneurysm formation is attenuated in osteopontin-deficient mice. J. Clin. Investig. 2003, 112, 1318–1331. [Google Scholar] [CrossRef]
- Ogawa, D.; Stone, J.F.; Takata, Y.; Blaschke, F.; Chu, V.H.; Towler, D.A.; Law, R.E.; Hsueh, W.A.; Bruemmer, D. Liver X Receptor Agonists Inhibit Cytokine-Induced Osteopontin Expression in Macrophages Through Interference With Activator Protein-1 Signaling Pathways. Circ. Res. 2005, 96, e59-67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyström, T.; Dunér, P.; Hultgårdh-Nilsson, A. A constitutive endogenous osteopontin production is important for macrophage function and differentiation. Exp. Cell Res. 2007, 313, 1149–1160. [Google Scholar] [CrossRef]
- Martín-Márquez, B.-T.; Sandoval-García, F.; Corona-Meraz, F.-I.; Petri, M.-H.; Gutiérrez-Mercado, Y.-K.; Mercado, M.V.-D. Osteopontin: Another piece in the systemic lupus erythematosus immunopathology puzzle. Clin. Exp. Rheumatol. 2022, 40, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nakai, T.; Tamura, N.; Okamoto, S.; Matsuoka, K.; Sakuraba, A.; Fukushima, T.; Uede, T.; Hibi, T. Osteopontin/Eta-1 upregulated in Crohn’s disease regulates the Th1 immune response. Gut 2005, 54, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnholt, J.; Kelsen, J.; Schack, L.; Hvas, C.L.; Dahlerup, J.F.; Sørensen, E.S. Osteopontin, a Protein with Cytokine-like Properties, is Associated with Inflammation in Crohn’s Disease. Scand. J. Immunol. 2007, 65, 453–460. [Google Scholar] [CrossRef]
- Mishima, R.; Takeshima, F.; Sawai, T.; Ohba, K.; Ohnita, K.; Isomoto, H.; Omagari, K.; Mizuta, Y.; Ozono, Y.; Kohno, S. High Plasma Osteopontin Levels in Patients With Inflammatory Bowel Disease. J. Clin. Gastroenterol. 2007, 41, 167–172. [Google Scholar] [CrossRef]
- Li, Y.; Xiu, W.; Xu, J.; Chen, X.; Wang, G.; Duan, J.; Sun, L.; Liu, B.; Xie, W.; Pu, G.; et al. Increased CHCHD2 expression promotes liver fibrosis in nonalcoholic steatohepatitis via Notch/osteopontin signaling. JCI Insight 2022, 7, e162402. [Google Scholar] [CrossRef]
- Remmerie, A.; Martens, L.; Thoné, T.; Castoldi, A.; Seurinck, R.; Pavie, B.; Roels, J.; Vanneste, B.; De Prijck, S.; Vanhockerhout, M.; et al. Osteopontin Expression Identifies a Subset of Recruited Macrophages Distinct from Kupffer Cells in the Fatty Liver. Immunity 2020, 53, 641–657.e14. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, Y.; Zhang, W.; Qin, C.; Zhu, Y.; Fang, Y.; Wang, Y.; Tang, C.; Cao, F. Osteopontin-Targeted and PPARδ-Agonist-Loaded Nanoparticles Efficiently Reduce Atherosclerosis in Apolipoprotein E–/– Mice. ACS Omega 2022, 7, 28767–28778. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, J.; Cui, G.; Wei, Y.; Lu, C.; Wang, L.; Diao, H. Pathophysiological role of osteopontin and angiotensin II in atherosclerosis. Biochem. Biophys. Res. Commun. 2016, 471, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Lamort, A.-S.; Giopanou, I.; Psallidas, I.; Stathopoulos, G.T. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells 2019, 8, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewski, T.; Zyromska, A.; Makarewicz, R.; Zekanowska, E. Osteopontin And Angiogenic Factors As New Biomarkers Of Prostate Cancer. Urol. J. 2019, 16, 134–140. [Google Scholar] [CrossRef]
- Di Bartolomeo, M.; Pietrantonio, F.; Pellegrinelli, A.; Martinetti, A.; Mariani, L.; Daidone, M.G.; Bajetta, E.; Pelosi, G.; de Braud, F.G.M.; Floriani, I.; et al. Osteopontin, E-cadherin, and β-catenin expression as prognostic biomarkers in patients with radically resected gastric cancer. Gastric Cancer 2016, 19, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Clemente, N.; Raineri, D.; Cappellano, G.; Boggio, E.; Favero, F.; Soluri, M.F.; Dianzani, C.; Comi, C.; Dianzani, U.; Chiocchetti, A. Osteopontin Bridging Innate and Adaptive Immunity in Autoimmune Diseases. J. Immunol. Res. 2016, 2016, 7675437. [Google Scholar] [CrossRef] [Green Version]
- Bassi, M.S.; Buttari, F.; Gilio, L.; Iezzi, E.; Galifi, G.; Carbone, F.; Micillo, T.; Dolcetti, E.; Azzolini, F.; Bruno, A.; et al. Osteopontin Is Associated with Multiple Sclerosis Relapses. Biomedicines 2023, 11, 178. [Google Scholar] [CrossRef]
- Bazzichi, L.; Ghiadoni, L.; Rossi, A.; Bernardini, M.; Lanza, M.; De Feo, F.; Giacomelli, C.; Mencaroni, I.; Raimo, K.; Rossi, M.; et al. Osteopontin Is Associated with Increased Arterial Stiffness in Rheumatoid Arthritis. Mol. Med. 2009, 15, 402–406. [Google Scholar] [CrossRef]
- Radu, A.-F.; Bungau, S.G. Management of Rheumatoid Arthritis: An Overview. Cells 2021, 10, 2857. [Google Scholar] [CrossRef]
- Khan, M.W.A.; Khan, W.A. Autoantibodies and Cytokines in Pathogenesis of Rheumatoid Arthritis. In Autoantibodies and Cytokines; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Yumoto, K.; Ishijima, M.; Rittling, S.R.; Tsuji, K.; Tsuchiya, Y.; Kon, S.; Nifuji, A.; Uede, T.; Denhardt, D.T.; Noda, M. Osteopontin deficiency protects joints against destruction in anti-type II collagen antibody-induced arthritis in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 4556–4561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-M.; Chiang, Y.-C.; Huang, C.-Y.; Hsu, C.-J.; Fong, Y.-C.; Tang, C.-H. Osteopontin Promotes Oncostatin M Production in Human Osteoblasts: Implication of Rheumatoid Arthritis Therapy. J. Immunol. 2015, 195, 3355–3364. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Wu, B.; Goodman, S.B.; Berry, G.J.; Goronzy, J.J.; Weyand, C.M. Metabolic Control of Autoimmunity and Tissue Inflammation in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 652771. [Google Scholar] [CrossRef]
- Jiang, Y.; Yao, M.; Liu, Q.; Zhou, C. OPN gene polymorphisms influence the risk of knee OA and OPN levels in synovial fluid in a Chinese population. Arthritis Res. Ther. 2013, 15, R3. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Guo, X.-H.; Yue, S.-T.; Shi, L.; Tang, C.; Qiang, B. Effects of Artesunate on the Expressions of Insulin-Like Growth Factor-1, Osteopontin and C-Telopeptides of Type II Collagen in a Rat Model of Osteoarthritis. Pharmacology 2018, 101, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-G.; Zeng, C.; Song, Y.; Tian, J.; Cheng, C.; Yang, T.; Li, H.; Zhang, F.-J.; Lei, G.-H. Effect of osteopontin on the mRNA expression of ADAMTS4 and ADAMTS5 in chondrocytes from patients with knee osteoarthritis. Exp. Ther. Med. 2015, 9, 1979–1983. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Zhang, F.-J.; Tian, J.; Tu, M.; Xiong, Y.-L.; Luo, W.; Li, Y.-S.; Song, B.-B.; Gao, S.-G.; Lei, G.-H. Osteopontin inhibits HIF-2α mRNA expression in osteoarthritic chondrocytes. Exp. Ther. Med. 2015, 9, 2415–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.-G.; Yu, Y.; Zeng, C.; Lu, S.-T.; Tian, J.; Cheng, C.; Li, L.-J.; Lei, G.-H. Phosphorylation of osteopontin has proapoptotic and proinflammatory effects on human knee osteoarthritis chondrocytes. Exp. Ther. Med. 2016, 12, 3488–3494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Cheng, C.; Kuang, S.-D.; Su, C.; Zhao, X.; Xiong, Y.-L.; Li, Y.-S.; Gao, S.-G. OPN Deficiency Increases the Severity of Osteoarthritis Associated with Aberrant Chondrocyte Senescence and Apoptosis and Upregulates the Expression of Osteoarthritis-Associated Genes. Pain Res. Manag. 2020, 2020, 3428587. [Google Scholar] [CrossRef]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 10922. [Google Scholar] [CrossRef]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef]
- Pandolfi, F.; Franza, L.; Carusi, V.; Altamura, S.; Andriollo, G.; Nucera, E. Interleukin-6 in Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 5238. [Google Scholar] [CrossRef]
- Laavola, M.; Leppänen, T.; Hämäläinen, M.; Vuolteenaho, K.; Moilanen, T.; Nieminen, R.; Moilanen, E. IL-6 in Osteoarthritis: Effects of Pine Stilbenoids. Molecules 2018, 24, 109. [Google Scholar] [CrossRef] [Green Version]
- Levescot, A.; Chang, M.H.; Schnell, J.; Nelson-Maney, N.; Yan, J.; Martínez-Bonet, M.; Grieshaber-Bouyer, R.; Lee, P.Y.; Wei, K.; Blaustein, R.B.; et al. IL-1β–driven osteoclastogenic Tregs accelerate bone erosion in arthritis. J. Clin. Investig. 2021, 131, e141008. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, J.; Liang, X.; Zhao, H.; Lu, J.; Ma, Q.; Jing, B.; Tian, F. IL-1β increases the expression of inflammatory factors in synovial fluid-derived fibroblast-like synoviocytes via activation of the NF-κB-mediated ERK-STAT1 signaling pathway. Mol. Med. Rep. 2019, 20, 4993–5001. [Google Scholar] [CrossRef] [Green Version]
- Dayer, J.-M.; Oliviero, F.; Punzi, L. A Brief History of IL-1 and IL-1 Ra in Rheumatology. Front. Pharmacol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenei-Lanzl, Z.; Meurer, A.; Zaucke, F. Interleukin-1β signaling in osteoarthritis—Chondrocytes in focus. Cell. Signal. 2019, 53, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Dayer, J.-M.; Manger, B. Interleukin-1 function and role in rheumatic disease. Nat. Rev. Rheumatol. 2016, 12, 14–24. [Google Scholar] [CrossRef]
- Kurowska, W.; Przygodzka, M.; Jakubaszek, M.; Kwiatkowska, B.; Maslinski, W. Interleukin-15 as a Biomarker Candidate of Rheumatoid Arthritis Development. J. Clin. Med. 2020, 9, 1555. [Google Scholar] [CrossRef]
- Koper-Lenkiewicz, O.M.; Sutkowska, K.; Wawrusiewicz-Kurylonek, N.; Kowalewska, E.; Matowicka-Karna, J. Proinflammatory Cytokines (IL-1, -6, -8, -15, -17, -18, -23, TNF-α) Single Nucleotide Polymorphisms in Rheumatoid Arthritis—A Literature Review. Int. J. Mol. Sci. 2022, 23, 2106. [Google Scholar] [CrossRef]
- Warner, S.C.; Nair, A.; Marpadga, R.; Chubinskaya, S.; Doherty, M.; Valdes, A.M.; Scanzello, C.R. IL-15 and IL15RA in Osteoarthritis: Association With Symptoms and Protease Production, but Not Structural Severity. Front. Immunol. 2020, 11, 1385. [Google Scholar] [CrossRef]
- Wang, J.; He, L.; Li, W.; Lv, S. A Role of IL-17 in Rheumatoid Arthritis Patients Complicated With Atherosclerosis. Front. Pharmacol. 2022, 13, 828933. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Liu, Y.-Z.; Zhou, X.-D.; Huang, Y.; Xu, N.-W. Role of IL-17 gene polymorphisms in osteoarthritis: A meta-analysis based on observational studies. World J. Clin. Cases 2020, 8, 2280–2293. [Google Scholar] [CrossRef]
- Dinesh, P.; Rasool, M. Multifaceted role of IL-21 in rheumatoid arthritis: Current understanding and future perspectives. J. Cell. Physiol. 2018, 233, 3918–3928. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Qi, C.; Liu, Y.; Gao, H.; Zhao, D.; Jiang, Y. Increased frequency of peripheral blood follicular helper T cells and elevated serum IL-21 levels in patients with knee osteoarthritis. Mol. Med. Rep. 2017, 15, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Aarts, J.; van Caam, A.; Chen, X.; Marijnissen, R.J.; Helsen, M.M.; Walgreen, B.; Vitters, E.L.; van de Loo, F.A.; van Lent, P.L.; van der Kraan, P.M.; et al. Local inhibition of TGF-β1 signaling improves Th17/Treg balance but not joint pathology during experimental arthritis. Sci. Rep. 2022, 12, 3182. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Xiao, H.; Liao, J.; Zhou, N.; Zhao, C.; Xu, W.; Xu, W.; Liang, X.; Huang, W. Osteocyte TGFβ1-Smad2/3 is positively associated with bone turnover parameters in subchondral bone of advanced osteoarthritis. Int. J. Mol. Med. 2020, 46, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, Y.; Mimura, T. Matrix Metalloproteinase Gene Activation Resulting from Disordred Epigenetic Mechanisms in Rheumatoid Arthritis. Int. J. Mol. Sci. 2017, 18, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, B.J.; Kooyman, D.L. A Tale of Two Joints: The Role of Matrix Metalloproteases in Cartilage Biology. Dis. Markers 2016, 2016, 4895050. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zhang, G.; Li, X.; Qiu, X.; Ouyang, J.; Dai, J.; Min, S. Matrix Metalloproteinase 3: A Promoting and Destabilizing Factor in the Pathogenesis of Disease and Cell Differentiation. Front. Physiol. 2021, 12, 663978. [Google Scholar] [CrossRef]
- Xue, M.; McKelvey, K.; Shen, K.; Minhas, N.; March, L.; Park, S.-Y.; Jackson, C.J. Endogenous MMP-9 and not MMP-2 promotes rheumatoid synovial fibroblast survival, inflammation and cartilage degradation. Rheumatology 2014, 53, 2270–2279. [Google Scholar] [CrossRef] [Green Version]
- Stojanovic, S.K.; Stamenkovic, B.N.; Cvetkovic, J.M.; Zivkovic, V.G.; Apostolovic, M.R.A. Matrix Metalloproteinase-9 Level in Synovial Fluid—Association with Joint Destruction in Early Rheumatoid Arthritis. Medicina 2023, 59, 167. [Google Scholar] [CrossRef]
- Luo, S.; Li, W.; Wu, W.; Shi, Q. Elevated expression of MMP8 and MMP9 contributes to diabetic osteoarthritis progression in a rat model. J. Orthop. Surg. Res. 2021, 16, 64. [Google Scholar] [CrossRef]
- Itoh, Y. Metalloproteinases in Rheumatoid Arthritis: Potential Therapeutic Targets to Improve Current Therapies. Prog. Mol. Biol. Transl. Sci. 2017, 148, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-L.; Duan, H.-X.; Liang, Q.; Huang, Y.-L.; Wang, L.-Y.; Zhang, Q.; Wu, C.-J.; Liu, S.-Q.; Peng, W. Targeting matrix metalloproteases: A promising strategy for herbal medicines to treat rheumatoid arthritis. Front. Immunol. 2022, 13, 1046810. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E. MMP-12, An Old Enzyme Plays a New Role in the Pathogenesis of Rheumatoid Arthritis? Am. J. Pathol. 2004, 165, 1069–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Rajasekaran, N.; Hartenstein, B.; Szabowski, S.; Gajda, M.; Angel, P.; Bräuer, R.; Illges, H. Collagenase-3 (MMP-13) deficiency protects C57BL/6 mice from antibody-induced arthritis. Arthritis Res. Ther. 2013, 15, R222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehana, E.-S.E.; Khafaga, A.F.; El-Blehi, S.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Park, J.M.; Lee, S.Y.M. A review of osteoarthritis signaling intervention using small-molecule inhibitors. Medicine 2022, 101, e29501. [Google Scholar] [CrossRef]
- Elemam, N.M.; Hannawi, S.; Maghazachi, A.A. Role of Chemokines and Chemokine Receptors in Rheumatoid Arthritis. Immunotargets Ther. 2020, 9, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Moadab, F.; Khorramdelazad, H.; Abbasifard, M. Role of CCL2/CCR2 axis in the immunopathogenesis of rheumatoid arthritis: Latest evidence and therapeutic approaches. Life Sci. 2021, 269, 119034. [Google Scholar] [CrossRef]
- Willcockson, H.; Ozkan, H.; Chubinskaya, S.; Loeser, R.F.; Longobardi, L. CCL2 induces articular chondrocyte MMP expression through ERK and p38 signaling pathways. Osteoarthr. Cartil. Open 2021, 3, 100136. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, M.; Deng, J.; Lv, X.; Liu, J.; Xiao, Y.; Yang, W.; Zhang, Y.; Li, C. Chemokine Signaling Pathway Involved in CCL2 Expression in Patients with Rheumatoid Arthritis. Yonsei Med. J. 2015, 56, 1134–1142. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yang, Z.; Zhang, Z.; Kang, Y.; Huang, G.; Wang, S.; Huang, H.; Liao, W. CCL3 serves as a potential plasma biomarker in knee degeneration (osteoarthritis). Osteoarthr. Cartil. 2015, 23, 1405–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowin, T.; Straub, R.H. Integrins and their ligands in rheumatoid arthritis. Arthritis Res. Ther. 2011, 13, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshitomi, H. Regulation of Immune Responses and Chronic Inflammation by Fibroblast-Like Synoviocytes. Front. Immunol. 2019, 10, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Jiang, S.; Wang, R.; Zhang, Y.; Dong, J.; Li, Y. Mechanistic Insight Into the Roles of Integrins in Osteoarthritis. Front. Cell Dev. Biol. 2021, 9, 693484. [Google Scholar] [CrossRef]
- Morshed, A.; Abbas, A.B.; Hu, J.; Xu, H. Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules 2019, 24, 1537. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, S.; Yamaguchi, N.; Nishioka, K.; Mima, T.; Ishii, T.; Umeshita-Sasai, M.; Kobayashi, H.; Shimizu, M.; Katada, Y.; Wakitani, S.; et al. Enhanced local production of osteopontin in rheumatoid joints. J. Rheumatol. 2002, 29, 2061–2067. [Google Scholar]
- Gattorno, M.; Gregorio, A.; Ferlito, F.; Gerloni, V.; Parafioriti, A.; Felici, E.; Sala, E.; Gambini, C.; Picco, P.; Martini, A. Synovial expression of osteopontin correlates with angiogenesis in juvenile idiopathic arthritis. Rheumatology 2004, 43, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Iwadate, H.; Kobayashi, H.; Kanno, T.; Asano, T.; Saito, R.; Sato, S.; Suzuki, E.; Watanabe, H.; Ohira, H. Plasma osteopontin is correlated with bone resorption markers in rheumatoid arthritis patients. Int. J. Rheum. Dis. 2014, 17, 50–56. [Google Scholar] [CrossRef]
- Shio, K.; Kobayashi, H.; Asano, T.; Saito, R.; Iwadate, H.; Watanabe, H.; Sakuma, H.; Segawa, T.; Maeda, M.; Ohira, H. Thrombin-cleaved Osteopontin Is Increased in Urine of Patients with Rheumatoid Arthritis. J. Rheumatol. 2010, 37, 704–710. [Google Scholar] [CrossRef]
- Honsawek, S.; Tanavalee, A.; Sakdinakiattikoon, M.; Chayanupatkul, M.; Yuktanandana, P. Correlation of plasma and synovial fluid osteopontin with disease severity in knee osteoarthritis. Clin. Biochem. 2009, 42, 808–812. [Google Scholar] [CrossRef]
- El Deeb, S.; Abdelnaby, R.; Khachab, A.; Bläsius, K.; Tingart, M.; Rath, B. Osteopontin as a Biochemical Marker and Severity Indicator for Idiopathic Hip Osteoarthritis. HIP Int. 2016, 26, 397–403. [Google Scholar] [CrossRef]
- Abdelnaby, R.; El Deeb, S.; Khachab, A.; Bläsius, K.; Tingart, M.; Rath, B. Plasma level of Osteopontin does not respond to total replacement Surgery in patients with severe Primary knee/Hip Osteoarthritis. J. Orthop. 2017, 14, 354–357. [Google Scholar] [CrossRef]
- Slovacek, H.; Khanna, R.; Poredos, P.; Jezovnik, M.; Hoppensteadt, D.; Fareed, J.; Hopkinson, W. Interrelationship of Osteopontin, MMP-9 and ADAMTS4 in Patients With Osteoarthritis Undergoing Total Joint Arthroplasty. Clin. Appl. Thromb. 2020, 26, 1076029620964864. [Google Scholar] [CrossRef]
- Dong, X.; Zheng, Y.; Liu, H.-Y. The clinical significance of serum and joint fluid osteopontin, and thrombin-cleaved osteopontin levels in osteoarthritis. Zhonghua Nei Ke Za Zhi 2013, 52, 1023–1027. [Google Scholar]
- Ji, H.-I.; Lee, S.-H.; Song, R.; Yang, H.-I.; Lee, Y.-A.; Hong, S.-J.; Kim, S.; Park, Y.-B.; Lee, S.-K.; Yoo, M.C.; et al. Serum level of osteopontin as an inflammatory marker does not indicate disease activity or responsiveness to therapeutic treatments in patients with rheumatoid arthritis. Clin. Rheumatol. 2014, 33, 397–402. [Google Scholar] [CrossRef]
- Min, S.; Shi, T.; Han, X.; Chen, D.; Xu, Z.; Shi, D.; Teng, H.; Jiang, Q. Serum levels of leptin, osteopontin, and sclerostin in patients with and without knee osteoarthritis. Clin. Rheumatol. 2021, 40, 287–294. [Google Scholar] [CrossRef]
- Gao, S.; Li, K.; Zeng, K.; Tu, M.; Xu, M.; Lei, G. Elevated osteopontin level of synovial fluid and articular cartilage is associated with disease severity in knee osteoarthritis patients. Osteoarthr. Cartil. 2010, 18, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.-F.; Wang, W.-C.; Fang, H.; Mao, X.-Z.; Huang, G.-L.; Chen, Y.; Zhou, H.-D.; Shen, Y.; Peng, D. Expression of NF-κB and osteopontin of synovial fluid of patients with knee osteoarthritis. Asian Pac. J. Trop. Med. 2013, 6, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Nourkami-Tutdibi, N.; Graf, N.; Beier, R.; Zemlin, M.; Tutdibi, E. Plasma levels of osteopontin from birth to adulthood. Pediatr. Blood Cancer 2020, 67, e28272. [Google Scholar] [CrossRef] [Green Version]
- Petrow, P.K.; Hummel, K.M.; Schedel, J.; Franz, J.K.; Klein, C.L.; Muller-Ladner, U.; Kriegsmann, J.; Chang, P.L.; Prince, C.W.; Gay, R.E.; et al. Expression of osteopontin messenger RNA and protein in rheumatoid arthritis: Effects of osteopontin on the release of collagenase 1 from articular chondrocytes and synovial fibroblasts. Arthritis Rheumatol. 2000, 43, 1597–1605. [Google Scholar] [CrossRef]
- Suzuki, K.; Zhu, B.; Rittling, S.R.; Denhardt, D.T.; Goldberg, H.A.; McCulloch, C.A.G.; Sodek, J. Colocalization of Intracellular Osteopontin With CD44 Is Associated With Migration, Cell Fusion, and Resorption in Osteoclasts. J. Bone Miner. Res. 2002, 17, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.-J.; Yu, W.-B.; Luo, W.; Gao, S.-G.; Li, Y.-S.; Lei, G.-H. Effect of osteopontin on TIMP-1 and TIMP-2 mRNA in chondrocytes of human knee osteoarthritis in vitro. Exp. Ther. Med. 2014, 8, 391–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Nie, H.; Li, N.; Zheng, W.; Zhang, D.; Feng, G.; Ni, L.; Xu, R.; Hong, J.; Zhang, J.Z. Role of osteopontin in amplification and perpetuation of rheumatoid synovitis. J. Clin. Investig. 2005, 115, 1060–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Li, R.; Pan, H.; He, D.; Xu, R.; Guo, T.B.; Guo, Y.; Zhang, J.Z. Role of osteopontin in induction of monocyte chemoattractant protein 1 and macrophage inflammatory protein 1β through the NF-κB and MAPK pathways in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1957–1965. [Google Scholar] [CrossRef]
- Varisco, P.A.; Péclat, V.; Van Ness, K.; Bischof-Delaloye, A.; So, A.; Busso, N. Effect of thrombin inhibition on synovial inflammation in antigen induced arthritis. Ann. Rheum. Dis. 2000, 59, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, S.; Kobayashi, H.; Yamaguchi, N.; Nishioka, K.; Umeshita-Sasai, M.; Mima, T.; Nomura, S.; Kon, S.; Inobe, M.; Uede, T.; et al. Expression of osteopontin at sites of bone erosion in a murine experimental arthritis model of collagen-induced arthritis: Possible involvement of osteopontin in bone destruction in arthritis. Arthritis Rheum. 2002, 46, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Zhang, X.; Li, R.; Fang, L.; Niu, X.; Zheng, Y.; He, D.; Xu, R.; Zhang, J.Z. Role of osteopontin in synovial Th17 differentiation in rheumatoid arthritis. Arthritis Rheum. 2010, 62, 2900–2908. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Liu, S.-C.; Wang, Y.-H.; Su, C.-M.; Huang, C.-C.; Hsu, C.-J.; Tang, C.-H. Osteopontin inhibition of miR-129-3p enhances IL-17 expression and monocyte migration in rheumatoid arthritis. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 15–22. [Google Scholar] [CrossRef]
- Luukkonen, J.; Pascual, L.M.; Patlaka, C.; Lång, P.; Turunen, S.; Halleen, J.; Nousiainen, T.; Valkealahti, M.; Tuukkanen, J.; Andersson, G.; et al. Increased amount of phosphorylated proinflammatory osteopontin in rheumatoid arthritis synovia is associated to decreased tartrate-resistant acid phosphatase 5B/5A ratio. PLoS ONE 2017, 12, e0182904. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.-F.; Wang, J.; Bai, H.-H.; He, J.-J.; Jia, R.-H.; Wang, X.; Zhang, W.-Q.; Zhao, X.-C.; Zhang, X.-C.; Liu, G.-Y.; et al. A Decreased Absolute Number of Treg Cells in Patients with Active Rheumatoid Arthritis is Associated with Elevated Serum Osteopontin Levels with Disease Progression. Adv. Ther. 2022, 39, 3280–3291. [Google Scholar] [CrossRef]
- Izumi, K.; Kaneko, Y.; Hashizume, M.; Yoshimoto, K.; Takeuchi, T. Baseline Serum Osteopontin Levels Predict the Clinical Effectiveness of Tocilizumab but Not Infliximab in Biologic-Naïve Patients with Rheumatoid Arthritis: A Single-Center Prospective Study at 1 Year (the Keio First-Bio Cohort Study). PLoS ONE 2015, 10, e0145468. [Google Scholar] [CrossRef] [PubMed]
- Sennels, H.P.; Sørensen, S.; Østergaard, M.; Knudsen, L.S.; Hansen, M.S.; Skjødt, H.; Peters, N.D.; Colic, A.; Grau, K.; Jacobsen, S. Circulating levels of osteopontin, osteoprotegerin, total soluble receptor activator of nuclear factor-kappa B ligand, and high sensitivity C-reactive protein in patients with active rheumatoid arthritis randomized to etanercept alone or in combination with methotrexate. Scand. J. Rheumatol. 2008, 37, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Pullig, O.; Weseloh, G.; Gauer, S.; Swoboda, B. Osteopontin is expressed by adult human osteoarthritic chondrocytes: Protein and mRNA analysis of normal and osteoarthritic cartilage. Matrix Biol. 2000, 19, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Yagi, R.; McBurney, D.; Laverty, D.; Weiner, S.; Horton, W.E., Jr. Intrajoint comparisons of gene expression patterns in human osteoarthritis suggest a change in chondrocyte phenotype. J. Orthop. Res. 2005, 23, 1128–1138. [Google Scholar] [CrossRef]
- Sanchez, C.; Deberg, M.A.; Bellahcène, A.; Castronovo, V.; Msika, P.; Delcour, J.P.; Crielaard, J.M.; Henrotin, Y.E. Phenotypic characterization of osteoblasts from the sclerotic zones of osteoarthritic subchondral bone. Arthritis Rheum. 2008, 58, 442–455. [Google Scholar] [CrossRef]
- Cheng, C.; Gao, S.; Lei, G. Association of osteopontin with osteoarthritis. Rheumatol. Int. 2014, 34, 1627–1631. [Google Scholar] [CrossRef]
- Kulkarni, P.; Srivastava, V.; Tootsi, K.; Electricwala, A.; Kharat, A.; Bhonde, R.; Koks, S.; Martson, A.; Harsulkar, A. Synovial Fluid in Knee Osteoarthritis Extends Proinflammatory Niche for Macrophage Polarization. Cells 2022, 11, 4115. [Google Scholar] [CrossRef]
- Tanaka, S.; Narusawa, K.; Onishi, H.; Miura, M.; Hijioka, A.; Kanazawa, Y.; Nishida, S.; Ikeda, S.; Nakamura, T. Lower osteocalcin and osteopontin contents of the femoral head in hip fracture patients than osteoarthritis patients. Osteoporos. Int. 2011, 22, 587–597. [Google Scholar] [CrossRef]
- Yamaga, M.; Tsuji, K.; Miyatake, K.; Yamada, J.; Abula, K.; Ju, Y.-J.; Sekiya, I.; Muneta, T. Osteopontin Level in Synovial Fluid Is Associated with the Severity of Joint Pain and Cartilage Degradation after Anterior Cruciate Ligament Rupture. PLoS ONE 2012, 7, e49014. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zhang, L.; Zhao, L.; Gao, S.; Han, R.; Su, D.; Lei, G. Phosphorylation of osteopontin in osteoarthritis degenerative cartilage and its effect on matrix metalloprotease 13. Rheumatol. Int. 2013, 33, 1313–1319. [Google Scholar] [CrossRef]
- Urcelay, E.; Martínez, A.; Mas-Fontao, A.; Peris-Pertusa, A.; Pascual-Salcedo, D.; Balsa, A.; Fernández-Arquero, M.; de la Concha, E. Osteopontin gene polymorphisms in Spanish patients with rheumatoid arthritis. J. Rheumatol. 2005, 32, 405–409. [Google Scholar]
- Xu, G.; Sun, W.; He, D.; Wang, L.; Zheng, W.; Nie, H.; Ni, L.; Zhang, D.; Li, N.; Zhang, J. Overexpression of osteopontin in rheumatoid synovial mononuclear cells is associated with joint inflammation, not with genetic polymorphism. J. Rheumatol. 2005, 32, 410–416. [Google Scholar]
- Marciano, R.; Giacopelli, F.; Divizia, M.T.; Gattorno, M.; Felici, E.; Pistorio, A.; Martini, A.; Ravazzolo, R.; Picco, P. A polymorphic variant inside the osteopontin gene shows association with disease course in oligoarticular juvenile idiopathic arthritis. Ann. Rheum. Dis. 2006, 65, 662–665. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, F.; D’Alfonso, S.; Perricone, C.; Carlomagno, Y.; Alessandri, C.; Croia, C.; Barizzone, N.; Montecucco, C.; Galeazzi, M.; Sebastiani, G.D.; et al. The role of eight polymorphisms in three candidate genes in determining the susceptibility, phenotype, and response to anti-TNF therapy in patients with rheumatoid arthritis. Clin. Exp. Rheumatol. 2012, 30, 939–942. [Google Scholar]
- Juge, P.-A.; van Steenbergen, H.W.; Constantin, A.; Tobon, G.J.; Schaeverbeke, T.; Gazal, S.; Combe, B.; Devauchelle-Pensec, V.; Nigon, D.; van der Helm-van Mil, A.H.; et al. SPP1rs9138 variant contributes to the severity of radiological damage in anti-citrullinated protein autoantibody-negative rheumatoid arthritis. Ann. Rheum. Dis. 2014, 73, 1840–1843. [Google Scholar] [CrossRef] [Green Version]
- Gazal, S.; Sacre, K.; Allanore, Y.; Teruel, M.; Goodall, A.H.; Tohma, S.; Alfredsson, L.; Okada, Y.; Xie, G.; Constantin, A.; et al. Identification of secreted phosphoprotein 1 gene as a new rheumatoid arthritis susceptibility gene. Ann. Rheum. Dis. 2015, 74, e19. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Li, Y.; Xu, J.; Cao, H.; Li, X.; Ma, B.; Ning, P.; Wang, Y.; Wang, M.; Zhu, S.; et al. Association of SPP1 promoter variants with hip osteoarthritis susceptibility in Chinese population. Gene 2015, 564, 9–13. [Google Scholar] [CrossRef]
- Borgonio-Cuadra, V.M.; González-Huerta, N.C.; Rojas-Toledo, E.X.; Morales-Hernández, E.; Pérez-Hernández, N.; Rodríguez-Pérez, J.M.; Tovilla-Zárate, C.A.; González-Castro, T.B.; Hernández-Díaz, Y.; López-Narváez, M.L.; et al. Genetic association analysis of Osteopontin and Matrix Gla Protein genes polymorphisms with primary knee osteoarthritis in Mexican population. Clin. Rheumatol. 2019, 38, 223–228. [Google Scholar] [CrossRef]
- Shang, H.; Hao, Y.; Hu, W.; Hu, X.; Jin, Q. OPN gene locus is associated with the risk of knee osteoarthritis: A case–control study. Biosci. Rep. 2019, 39, BSR20182023. [Google Scholar] [CrossRef] [Green Version]
- Take, Y.; Nakata, K.; Hashimoto, J.; Tsuboi, H.; Nishimoto, N.; Ochi, T.; Yoshikawa, H. Specifically modified osteopontin in rheumatoid arthritis fibroblast-like synoviocytes supports interaction with B cells and enhances production of interleukin-6. Arthritis Rheum. 2009, 60, 3591–3601. [Google Scholar] [CrossRef]
- Mehta, B.B.; Sharma, S.; Vasishta, R.K.; Sen, R.K.; Sharma, A.; Luthra-Guptasarma, M. Blocking osteopontin-fibronectin interactions reduce extracellular fibronectin deployment and arthritic immunopathology. Int. Immunopharmacol. 2018, 55, 297–305. [Google Scholar] [CrossRef]
- Asano, T.; Iwasaki, N.; Kon, S.; Kanayama, M.; Morimoto, J.; Minami, A.; Uede, T. 9 1 integrin acts as a critical intrinsic regulator of human rheumatoid arthritis. Rheumatology 2014, 53, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Oh, I.; Morimoto, J.; Uede, T.; Morimoto, A. Osteopontin Has a Crucial Role in Osteoclast-Like Multinucleated Giant Cell Formation. J. Cell. Biochem. 2014, 115, 585–595. [Google Scholar] [CrossRef]
- Shi, L.; Shi, L.; Wang, X.; He, J. Regulatory Roles of Osteopontin in Production of Monocyte-Origin MCP-1. Cell Transplant. 2018, 27, 1185–1194. [Google Scholar] [CrossRef] [Green Version]
- Murthy, S.; Karkossa, I.; Schmidt, C.; Hoffmann, A.; Hagemann, T.; Rothe, K.; Seifert, O.; Anderegg, U.; von Bergen, M.; Schubert, K.; et al. Danger signal extracellular calcium initiates differentiation of monocytes into SPP1/osteopontin-producing macrophages. Cell Death Dis. 2022, 13, 53. [Google Scholar] [CrossRef]
- Gao, C.; Guo, H.; Mi, Z.; Wai, P.Y.; Kuo, P.C. Transcriptional Regulatory Functions of Heterogeneous Nuclear Ribonucleoprotein-U and -A/B in Endotoxin-Mediated Macrophage Expression of Osteopontin. J. Immunol. 2005, 175, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-J.; Gao, S.-G.; Cheng, L.; Tian, J.; Xu, W.-S.; Luo, W.; Song, Y.; Yang, Y.; Lei, G.-H. The effect of hyaluronic acid on osteopontin and CD44 mRNA of fibroblast-like synoviocytes in patients with osteoarthritis of the knee. Rheumatol. Int. 2012, 33, 79–83. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Zhang, F.; Luo, W. Effect of ligand-receptor interaction of osteopontin-CD44 on the expression of hyaluronic acid in human knee osteoarthritic chondrocytes in vitro. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2020, 45, 1015–1023. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Wu, Z.; Luo, W. Mechanistic Insight on the Interaction between OPN and Integrin ανβ3 in Osteoarthritis. Biomed. Res. Int. 2020, 2020, 2905634. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, T.; Cao, Y. Osteopontin can decrease the expression of Col-II and COMP in cartilage cells in vitro. Int. J. Clin. Exp. Med. 2015, 8, 2254–2260. [Google Scholar]
- Xu, J.; Yi, Y.; Li, L.; Zhang, W.; Wang, J. Osteopontin induces vascular endothelial growth factor expression in articular cartilage through PI3K/AKT and ERK1/2 signaling. Mol. Med. Rep. 2015, 12, 4708–4712. [Google Scholar] [CrossRef] [Green Version]
- Katayama, Y.; House, C.M.; Udagawa, N.; Kazama, J.J.; McFarland, R.J.; Martin, T.J.; Findlay, D.M. Casein kinase 2 phosphorylation of recombinant rat osteopontin enhances adhesion of osteoclasts but not osteoblasts. J. Cell. Physiol. 1998, 176, 179–187. [Google Scholar] [CrossRef]
- Liu, Q.; Zeng, H.; Yuan, Y.; Wang, Z.; Wu, Z.; Luo, W. Osteopontin inhibits osteoarthritis progression via the OPN/CD44/PI3K signal axis. Genes Dis. 2022, 9, 128–139. [Google Scholar] [CrossRef]
- Yoshitake, H.; Rittling, S.R.; Denhardt, D.T.; Noda, M. Osteopontin-deficient mice are resistant to ovariectomy-induced bone resorption. Proc. Natl. Acad. Sci. USA 1999, 96, 8156–8160. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Sakai, F.; Kon, S.; Morimoto, J.; Kimura, C.; Yamazaki, H.; Okazaki, I.; Seki, N.; Fujii, T.; Uede, T. Essential role of the cryptic epitope SLAYGLR within osteopontin in a murine model of rheumatoid arthritis. J. Clin. Investig. 2003, 112, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, M.; Kurotaki, D.; Morimoto, J.; Asano, T.; Matsui, Y.; Nakayama, Y.; Saito, Y.; Ito, K.; Kimura, C.; Iwasaki, N.; et al. α9 Integrin and Its Ligands Constitute Critical Joint Microenvironments for Development of Autoimmune Arthritis. J. Immunol. 2009, 182, 8015–8025. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, M.; Morimoto, J.; Matsui, Y.; Ikesue, M.; Danzaki, K.; Kurotaki, D.; Ito, K.; Yoshida, T.; Uede, T. α9β1 Integrin-Mediated Signaling Serves as an Intrinsic Regulator of Pathogenic Th17 Cell Generation. J. Immunol. 2011, 187, 5851–5864. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.P.; Pettit, A.R.; Shinohara, M.L.; Jansson, M.; Cantor, H.; Gravallese, E.M.; Mathis, D.; Benoist, C. Lack of requirement of osteopontin for inflammation, bone erosion, and cartilage damage in the K/BxN model of autoantibody-mediated arthritis. Arthritis Rheum. 2004, 50, 2685–2694. [Google Scholar] [CrossRef]
- Matsui, Y.; Iwasaki, N.; Kon, S.; Takahashi, D.; Morimoto, J.; Matsui, Y.; Denhardt, D.T.; Rittling, S.; Minami, A.; Uede, T. Accelerated development of aging-associated and instability-induced osteoarthritis in osteopontin-deficient mice. Arthritis Rheum. 2009, 60, 2362–2371. [Google Scholar] [CrossRef]
- Almonte-Becerril, M.; Costell, M.; Kouri, J.B. Changes in the integrins expression are related with the osteoarthritis severity in an experimental animal model in rats. J. Orthop. Res. 2014, 32, 1161–1166. [Google Scholar] [CrossRef]
- Gao, S.; Cheng, L.; Zeng, C.; Wei, L.; Zhang, F.; Tian, J.; Tu, M.; Luo, W.; Lei, G. Usefulness of specific OA biomarkers, thrombin-cleaved osteopontin, in the posterior cruciate ligament OA rabbit model. Osteoarthr. Cartil. 2013, 21, 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Calleja, A.; Velasquillo, C.; Vega-López, M.; Arellano-Jiménez, J.; Tsutsumi-Fujiyoshi, V.; Mondragón-Flores, R.; Kouri-Flores, J. Osteopontin expression and localization of Ca2+ deposits in early stages of osteoarthritis in a rat model. Histol. Histopathol. 2014, 29, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.F.; Kong, W.K.; Liu, L.; Liu, Y.; Liu, F.M.; Liu, W.J.; Yu, H.; Yang, W.L.; Li, G.Q.; Sun, Q.R. Osteopontin accelerates chondrocyte proliferation in osteoarthritis rats through the NF-kappab signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2836–2842. [Google Scholar] [CrossRef]
- Lin, C.; Chen, Z.; Guo, D.; Zhou, L.; Lin, S.; Li, C.; Li, S.; Wang, X.; Lin, B.; Ding, Y. Increased expression of osteopontin in subchondral bone promotes bone turnover and remodeling, and accelerates the progression of OA in a mouse model. Aging 2022, 14, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Moran-Moguel, M.C.; Petarra-Del Rio, S.; Mayorquin-Galvan, E.E.; Zavala-Cerna, M.G. Rheumatoid Arthritis and miRNAs: A Critical Review through a Functional View. J. Immunol. Res. 2018, 2018, 2474529. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qian, K.; Li, C.; Ma, Y.; Chen, X. Roles of microRNA-539 and osteopontin in rheumatoid arthritis. Exp. Ther. Med. 2018, 15, 2681–2687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Xu, L.; Sun, X.; Wang, Y.; Xuan, W.; Zhang, Q.; Zhao, P.; Wu, Q.; Liu, R.; Che, N.; et al. Adiponectin aggravates bone erosion by promoting osteopontin production in synovial tissue of rheumatoid arthritis. Arthritis Res. Ther. 2018, 20, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.; Li, Y.; Zeng, C.; Deng, Z.; Gao, S.; Xiao, W.; Luo, W.; Jiang, W.; Li, L.; Lei, G. MicroRNA-127-5p regulates osteopontin expression and osteopontin-mediated proliferation of human chondrocytes. Sci. Rep. 2016, 6, 25032. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Tian, X.; Huang, X.; He, L.; Xu, F. microRNA-186 inhibition of PI3K–AKT pathway via SPP1 inhibits chondrocyte apoptosis in mice with osteoarthritis. J. Cell. Physiol. 2019, 234, 6042–6053. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.; Liu, J.; Yang, B.; Wang, T.; Zhang, Z.; Jiang, X.; Guo, Y.; Zhang, Y. Roles of long noncoding RNA in osteoarthritis (Review). Int. J. Mol. Med. 2021, 48, 1–8. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, W.; Zhang, F.; Deng, Y.; Long, Z. NEAT1/miR-181c Regulates Osteopontin (OPN)-Mediated Synoviocyte Proliferation in Osteoarthritis. J. Cell. Biochem. 2017, 118, 3775–3784. [Google Scholar] [CrossRef]
- Liang, J.; Xu, L.; Zhou, F.; Liu, A.; Ge, H.; Chen, Y.; Tu, M. MALAT1/miR-127-5p Regulates Osteopontin (OPN)-Mediated Proliferation of Human Chondrocytes Through PI3K/Akt Pathway. J. Cell. Biochem. 2018, 119, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lv, G.; Wang, B.; Kuang, L. XIST/miR-376c-5p/OPN axis modulates the influence of proinflammatory M1 macrophages on osteoarthritis chondrocyte apoptosis. J. Cell. Physiol. 2020, 235, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Asila, A.; Deng, Y.; Liao, J.; Liu, Z.; Fang, R. Osteopontin-induced lncRNA HOTAIR expression is involved in osteoarthritis by regulating cell proliferation. BMC Geriatr. 2021, 21, 57. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Tu, M. The lncRNA MIAT/miR-181a-5p axis regulates osteopontin (OPN)-mediated proliferation and apoptosis of human chondrocytes in osteoarthritis. J. Mol. Histol. 2022, 53, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Nakashima, T.; Torikai, M.; Naruse, T.; Morimoto, J.; Kon, S.; Sakai, F.; Uede, T. Successful treatment of collagen-induced arthritis in non-human primates by chimeric anti-osteopontin antibody. Int. Immunopharmacol. 2007, 7, 1460–1470. [Google Scholar] [CrossRef]
- Fan, K.; Dai, J.; Wang, H.; Wei, H.; Cao, Z.; Hou, S.; Qian, W.; Wang, H.; Li, B.; Zhao, J.; et al. Treatment of collagen-induced arthritis with an anti-osteopontin monoclonal antibody through promotion of apoptosis of both murine and human activated T cells. Arthritis Rheum. 2008, 58, 2041–2052. [Google Scholar] [CrossRef]
- Boumans, M.J.H.; Houbiers, J.G.A.; Verschueren, P.; Ishikura, H.; Westhovens, R.; Brouwer, E.; Rojkovich, B.; Kelly, S.; Adel, M.D.; Isaacs, J.; et al. Safety, tolerability, pharmacokinetics, pharmacodynamics and efficacy of the monoclonal antibody ASK8007 blocking osteopontin in patients with rheumatoid arthritis: A randomised, placebo controlled, proof-of-concept study. Ann. Rheum. Dis. 2012, 71, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Farrokhi, V.; Chabot, J.R.; Neubert, H.; Yang, Z. Assessing the Feasibility of Neutralizing Osteopontin with Various Therapeutic Antibody Modalities. Sci. Rep. 2018, 8, 7781. [Google Scholar] [CrossRef]
- Takanashi, M.; Oikawa, K.; Sudo, K.; Tanaka, M.; Fujita, K.; Ishikawa, A.; Nakae, S.; Kaspar, R.L.; Matsuzaki, M.; Kudo, M.; et al. Therapeutic silencing of an endogenous gene by siRNA cream in an arthritis model mouse. Gene Ther. 2009, 16, 982–989. [Google Scholar] [CrossRef]
- Tsai, S.-W.; Lin, C.-C.; Lin, S.-C.; Wang, S.-P.; Yang, D.-H. Isorhamnetin ameliorates inflammatory responses and articular cartilage damage in the rats of monosodium iodoacetate-induced osteoarthritis. Immunopharmacol. Immunotoxicol. 2019, 41, 504–512. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ref. | Disease | Immunopathogenic Function | |
|---|---|---|---|
| Cytokine | |||
| Immunopathogenesis key regulator. Activates | |||
| [78] | RA | endothelial cells and recruits proinflammatory | |
| TNF-α | cytokines such as IL-6 and IL-1β. | ||
| [79] | OA | Leads joint and cartilage degeneration by | |
| collagen disruption and proteoglycan degradation. | |||
| Crucial role in inflammation process in joints, in | |||
| [80] | RA | OC-mediated bone resorption and pannus | |
| IL-6 | development. | ||
| [81] | OA | Suppresses proteoglycan production, induces | |
| oxidative stress, and increases ROS production. | |||
| Mediates bone erosion through induction | |||
| [82,83] | RA | of osteoclastogenic Treg, Promotes the upregulation | |
| IL-1β | of TNF-α and IL-17. | ||
| Drives MMP-9, ADAMTS-4, ADAMTS-5, | |||
| [84,85,86] | OA | and RANKL production associated with | |
| cartilage catabolism and matrix degradation. | |||
| Promotes TNF-α and IL-17 production and osteoclastogenesis. | |||
| [87,88] | RA | Stimulates and activates T and NK cells, neutrophils, and | |
| IL-15 | macrophages. | ||
| [79,89] | OA | Enhances MMP-9 production and induces MMP-1 | |
| and MMP-3 secretion from cartilage chondrocytes in vitro. | |||
| Related to an OC and osteoblast imbalance, | |||
| [90] | RA | it promotes MMPs release in synoviocytes, causing | |
| IL-17 | joint destruction. | ||
| [91] | OA | Induces the release of chemokines by chondrocytes | |
| and FLS, contributing to cartilage collapse. | |||
| Stimulates OC formation by upregulating RANKL | |||
| [78] | RA | production from T cells in RA synovitis. Role in | |
| IL-18 | onset and maintenance of inflammatory response. | ||
| [79] | OA | Induces the synthesis of MMP-1, MMP-3, and MMP-13, | |
| enhancing cartilage degradation. | |||
| Activates T and B cells, monocytes/macrophages, and | |||
| [78,92] | RA | FLS through JAK/STAT, MAPK, and PI3K/Akt signaling | |
| IL-21 | pathways. | ||
| [93] | OA | Produced by TFH cells, contributes to inflammation and | |
| progression of OA stages. | |||
| Promotes TH17 differentiation and regulates | |||
| [94] | RA | Treg function. TGF-β1 induces IL-6 production | |
| TGF-β | in arthritic synovium. | ||
| Alters the osteogenic activity of MSC. Generates | |||
| [95] | OA | early non-coupled osteogenesis/osteoclastogenesis, | |
| accelerating local bone island formation. | |||
| MMP | |||
| MMP-1/MMP-3 | [96,97] | AR | Peripheral levels can predict progression |
| of joint destruction. | |||
| Promotes vascular invasion, degrades cartilage matrix, | |||
| MMP-3 | [98] | OA | induces OC differentiation, and facilitates inflammatory |
| cell accumulation in articular cartilage. | |||
| Contributes to FLS survival, proliferation, migration, and | |||
| [99,100] | AR | Invasion, promoting joint destruction. | |
| Stimulates TNF-α, IL-6, and IL-8 production, and its | |||
| MMP-9 | SF levels can predict radiographic progression. | ||
| [101] | OA | Downregulates COL2A1 and COL1A1 cartilage | |
| expression and promotes knee pathological changes. | |||
| Promotes macrophage infiltration in the pannus, | |||
| MMP-12 | [102,103,104] | AR | enhancing inflammation. Increases synovial |
| thickening and cartilage destruction. | |||
| Promotes inflammation and joint destruction in arthritis | |||
| [105] | AR | models. Participates in cartilage destruction in modulating | |
| the inflammatory response. | |||
| MMP-13 | Plays a crucial role in COL2, ADAMTS-4, | ||
| [106,107] | OA | ADAMTS-5, and aggrecan degradation. Causes synovial | |
| hyperplasia and synovitis with diffuse mononuclear | |||
| cell infiltration. | |||
| Chemokine | |||
| [108,109] | AR | Promotes the locomotion and recruitment of monocytes | |
| CCL2 (MCP-1) | and macrophages to the inflamed joint. | ||
| [110] | OA | Induces articular chondrocyte MMP-1, MMP-3, and | |
| MMP-13 upregulation. | |||
| [111] | AR | Acts as a major chemoattractant for CCR5+ leucocytes | |
| CCL4 (MIP-1β) | from peripheral blood to articular tissue. | ||
| [112] | OA | Upregulated in response to IL-1β and resistin. | |
| Biomarker of K-OA severity. |
| Sample | Ref. | n | sOPN Levels |
|---|---|---|---|
| Plasma | [118] | 45 JIA | 0.25 ng/mL (0–1.160 ng/mL) |
| [66] | 41 RA | 9.93 ng/mL (4.36–47.80 ng/mL) | |
| 28 SSc | 4.3 ng/mL (2.1–19.7 ng/mL) | ||
| 18 Co | 5.2 ng/mL (4.1–9.4 ng/mL) | ||
| [119] | 37 RA | RA-R = 34.8 ng/mL (39.8–1150.9 ng/mL) | |
| RA-NR = 114 ng/mL (38.4–854.9 ng/mL) | |||
| [120] | 70 RA | 499.4 ng/mL (58.8–1492.8 ng/mL) | |
| 20 OA | 413.9 ng/mL (195.6–1047.8 ng/mL) | ||
| 46 Co | 277.9 ng/mL (114.0–655.1 ng/mL) | ||
| [121] | 32 OA | K-OA = 168.8 ± 15.6 ng/mL | |
| 15 Co | 67.2 ± 7.7 ng/mL | ||
| 309.78 ng/mL (212.2–521.5 ng/mL) | |||
| [122] | 21 OA | H-OA KL-3 = 283.41 ng/mL (212.2–400.1 ng/mL) | |
| H-OA KL-4 = 362. 51 ng/mL (256.3–521.5 ng/mL) | |||
| Pre-operative | |||
| K-OA = 305.85 ± 55.31 ng/mL | |||
| [123] | 30 OA | H-OA = 303.06 ± ng/mL | |
| Post-operative | |||
| K-OA = 434.2 ± 144.85 ng/mL | |||
| H-OA = 376.14 ± 90.69 ng/mL | |||
| Pre-operative | |||
| 155 ± 12.8 ng/mL | |||
| Day 1 post-operative | |||
| [124] | 63 OA | 152 ± 11.6 ng/mL | |
| Day 5–7 post-operative | |||
| 140 ± 13.8 ng/mL | |||
| 150 ± 7.20 ng/mL | |||
| 50 Co | 66.3 ± 2.49 ng/mL | ||
| Serum | FL = 1.7 ± 1.1 ng/mL | ||
| [125] | 22 OA | N-half = 21.1 ± 11.7 ng/mL | |
| F-OA N-half = 23.2 ± 11.8 ng/mL | |||
| M-OA N-half = 11.1 ± 4.9 ng/mL | |||
| 12 Co | FL = 1.8 ± 0.5 ng/mL | ||
| N-half = 14.7 ± 3.6 ng/mL | |||
| 21.804 ± 14.932 ng/mL | |||
| RA-M = 16.943 ± 8.055 ng/mL | |||
| [126] | 40 RA | RA-S = 25.397 ± 17.785 ng/mL | |
| RA-R baseline = 22.223 ± 20.106 ng/mL | |||
| RA-R after DMARD = 11.248 ± 9.583 ng/mL | |||
| 11 Co | 6.900 ± 4.593 ng/mL | ||
| K-OA = 4.908 ± 0.769 ng/mL | |||
| [127] | 148 OA | KL-2 = 5.684 ± 0.727 ng/mL | |
| KL-3 = 4.494 ± 0.592 ng/mL | |||
| KL-4 = 5.136 ± 0.673 ng/mL | |||
| 101 Co | 2.182 ± 0.217 ng/mL | ||
| SF | [118] | 45 JIA | 2.55 ng/mL |
| [121] | 32 OA | K-OA = 272.1 ± 15.0 ng/mL | |
| K-OA = 4.519 ± 1.830 ng/mL | |||
| [128] | 50 OA | KL-2 = 3.543 ± 0.811 ng/mL | |
| KL-3 = 4.013 ± 0.676 ng/mL | |||
| KL-4 = 6.170 ± 0.773 ng/mL | |||
| 10 Co | 1.179. ± 0.303 ng/mL | ||
| [125] | 22 OA | FL = 7.75 ± 2.33 ng/mL | |
| F-OA N-half = 0.91 ± 0.49 ng/mL | |||
| M-OA N-half = 0.12 ± 0.08 ng/mL | |||
| KL-2 N-half = 0.669 ± 0.476 ng/mL | |||
| [72] | 750 OA | KL-3 N-half = 5.203 ± 0.385 ng/mL | |
| KL-4 N-half = 5.642 ± 0.350 ng/mL | |||
| [129] | 42 OA | 3.378 ± 4.195 ng/mL | |
| 40 Co | 0.892 ± 0.323 ng/mL | ||
| Urine | 70 RA | FL = 2044.6 ng/mmol Cr | |
| N-half = 143.5 pmol/mmol Cr | |||
| [120] | 20 OA | FL = 945.7 ng/mmol Cr | |
| N half = 69.8 pmol/mmol Cr | |||
| 46 Co | 67.9 pmol/mmol Cr |
| Population | Ref. | n | SPP1 SNP | Main Findings |
|---|---|---|---|---|
| 327 T/C | ||||
| [152] | 263 RA | 795 C/T | Strong linkage disequilibrium | |
| 478 Co | 1128 A/G | in SPP1 SNP. | ||
| 1284 A/C | ||||
| +245 first intron | Association of variant 2 with | |||
| [154] | 119 JIA | biallelic ins/del | persistent oligoarticular JIA vs. | |
| 200 Co | (variant 1 = TG allele; | extended form. | ||
| variant 2 = TGTG allele) | ||||
| Association of rs11439060 with | ||||
| European | [155] | 377 RA | rs11439060 (−156/GG) | RA susceptibility (p = 0.023, |
| 391 Co | rs9138 (+1239 A/C) | OR 1.47). | ||
| Association of rs9138 with | ||||
| joint destruction in 358 ACPA- | ||||
| negative patients (p = 0.034). | ||||
| Association of rs4754 in complete | ||||
| [156] | 652 RA | rs9138 | linkage disequilibrium with | |
| (ESPOIR) | rs11439060 | rs9138 with joint damage | ||
| 273 RA | rs4754 | progression in ACPA-negative | ||
| (EAC) | patients at 2 (p = 0.019) and 7 | |||
| year follow-ups (p = 0.050). | ||||
| −631 G/T | ||||
| −458 T/C | SPP1 genotype and allele | |||
| 192 RA | rs4754 | frequencies do not differ between | ||
| 288 Co | rs1126616 | RA patients and Co. | ||
| rs1126772 | ||||
| rs9138 | ||||
| Association of −443 C/T and −66 | ||||
| [72] | 750 OA | −156 GG/G | T/G with OA risk and | |
| Han Chinese | 794 Co | −443 C/T | radiographic severity, and | |
| −66 T/G | affect SF sOPN N-half levels. | |||
| 389 OA | rs17524488 | rs17524488 delG > insG increase | ||
| [158] | 315 Co | rs11730582 | H-OA risk (OR 1.48, 95% CI | |
| 1.18–1.85, p < 0.01). | ||||
| rs11730582 C allele of C | ||||
| 403 OA | genotype related to decreased | |||
| [160] | 536 Co | rs11730582 | risk of H-OA. Association | |
| among females and BMI | ||||
| <25 kg/m2 groups. | ||||
| Mexican | [159] | 296 OA | rs11730582 | No association between |
| 354 Co | rs11730582 and primary K-OA. | |||
| Nine | rs11439060 | Association of rs11439060 and | ||
| independent | [157] | 11,715 RA | (−156/GG) | rs9138 risk allele with ACPA |
| Cohorts * | 26,493 C | rs9138 | negative (p = 1.29 × 10−5). | |
| (+1239 A/C) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Márquez, B.T.; Sandoval-García, F.; Corona-Meraz, F.I.; Martínez-García, E.A.; Sánchez-Hernández, P.E.; Salazar-Páramo, M.; Fletes-Rayas, A.L.; González-Inostroz, D.; Vazquez-Del Mercado, M. Osteopontin: A Bone-Derived Protein Involved in Rheumatoid Arthritis and Osteoarthritis Immunopathology. Biomolecules 2023, 13, 502. https://doi.org/10.3390/biom13030502
Martín-Márquez BT, Sandoval-García F, Corona-Meraz FI, Martínez-García EA, Sánchez-Hernández PE, Salazar-Páramo M, Fletes-Rayas AL, González-Inostroz D, Vazquez-Del Mercado M. Osteopontin: A Bone-Derived Protein Involved in Rheumatoid Arthritis and Osteoarthritis Immunopathology. Biomolecules. 2023; 13(3):502. https://doi.org/10.3390/biom13030502
Chicago/Turabian StyleMartín-Márquez, Beatriz Teresita, Flavio Sandoval-García, Fernanda Isadora Corona-Meraz, Erika Aurora Martínez-García, Pedro Ernesto Sánchez-Hernández, Mario Salazar-Páramo, Ana Lilia Fletes-Rayas, Daniel González-Inostroz, and Monica Vazquez-Del Mercado. 2023. "Osteopontin: A Bone-Derived Protein Involved in Rheumatoid Arthritis and Osteoarthritis Immunopathology" Biomolecules 13, no. 3: 502. https://doi.org/10.3390/biom13030502