
Progress of Wnt Signaling Pathway in Osteoporosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Bone Growth Factors
2.1. Transforming Growth Factor β (TGF-β)
2.2. Bone Morphogenetic Protein (BMP)
2.3. Fibroblast Growth Factor (FGF)
3. Wnt Signaling Pathway
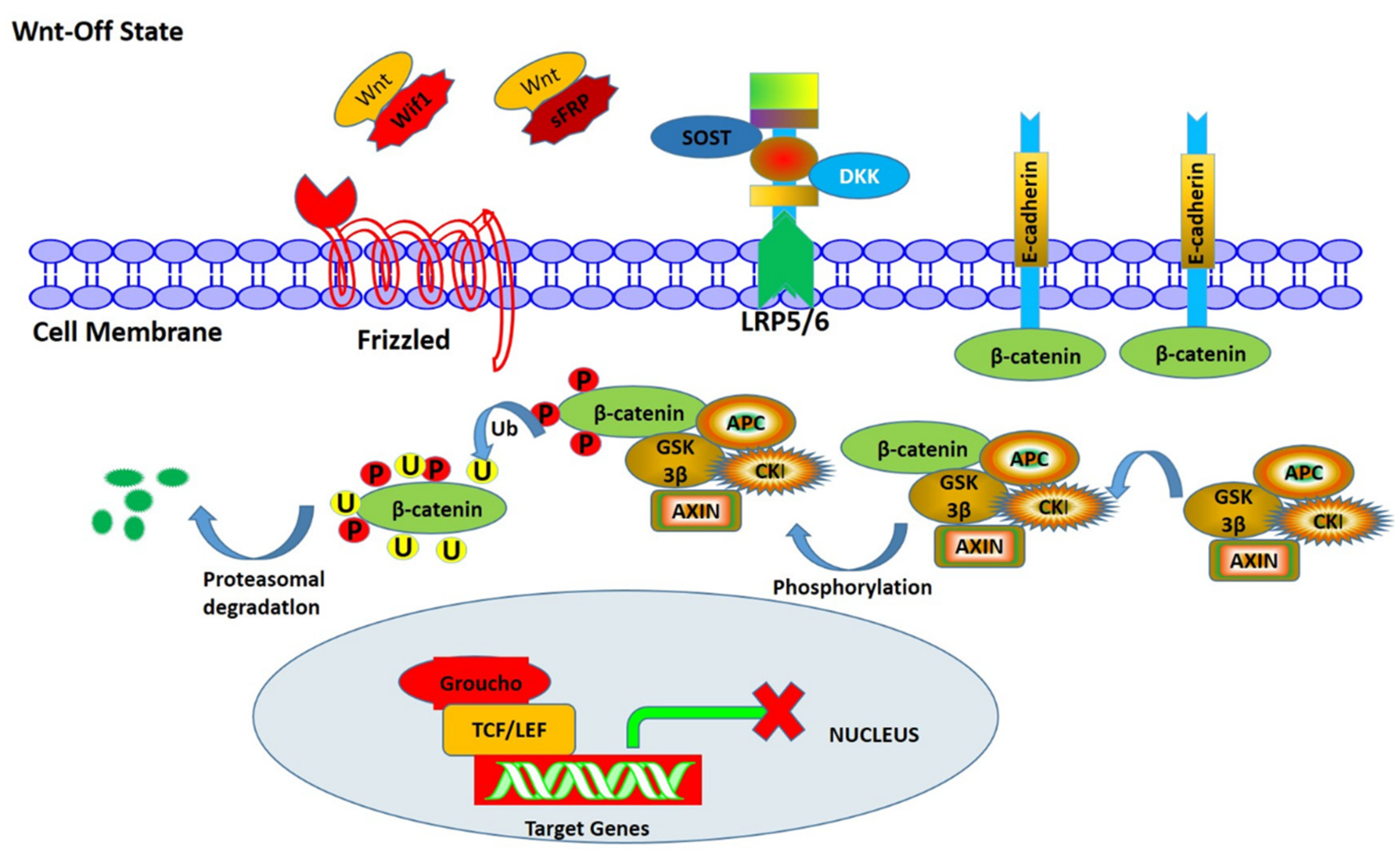
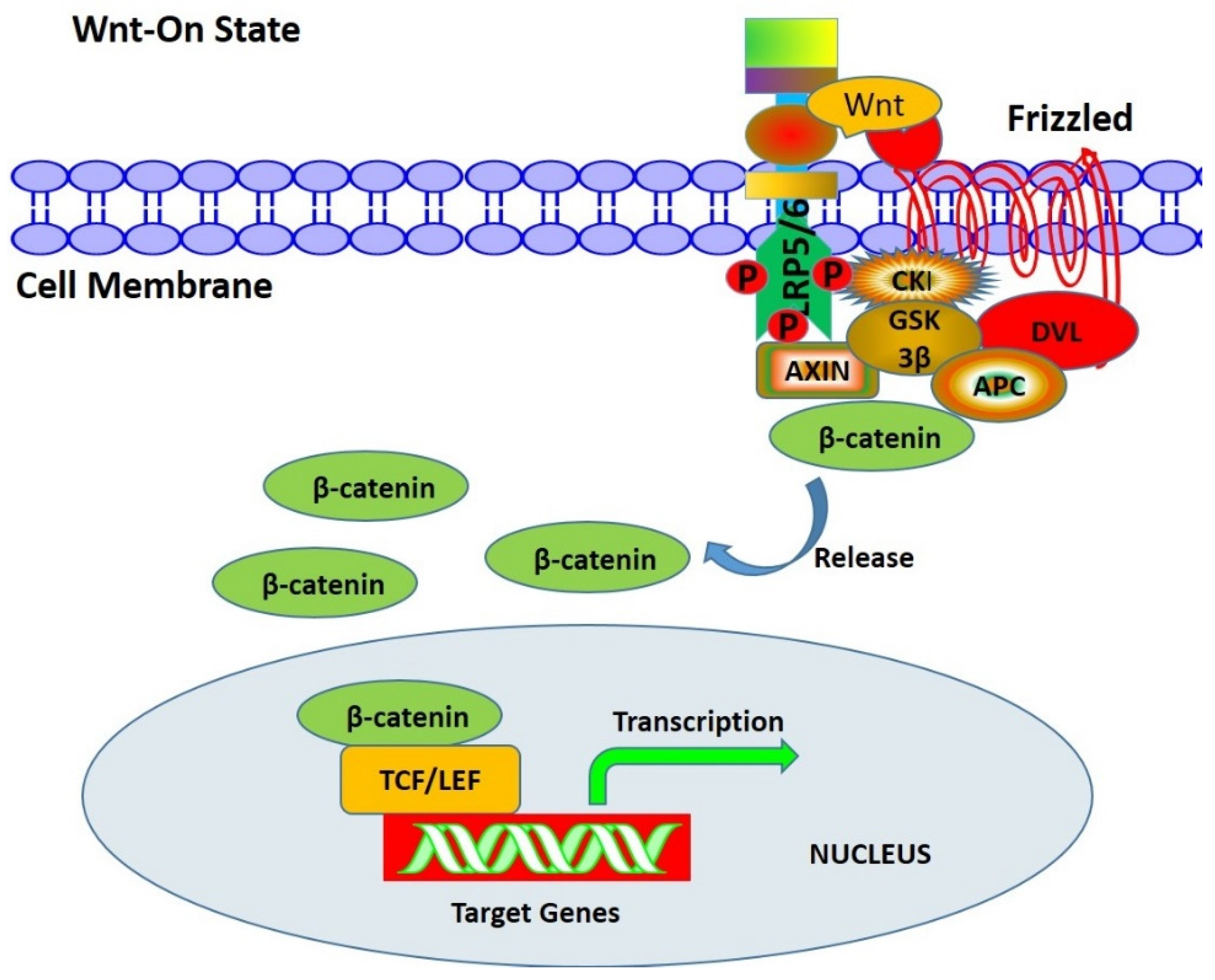
3.1. Wnt/β-Catenin Signaling Pathway
- (1)
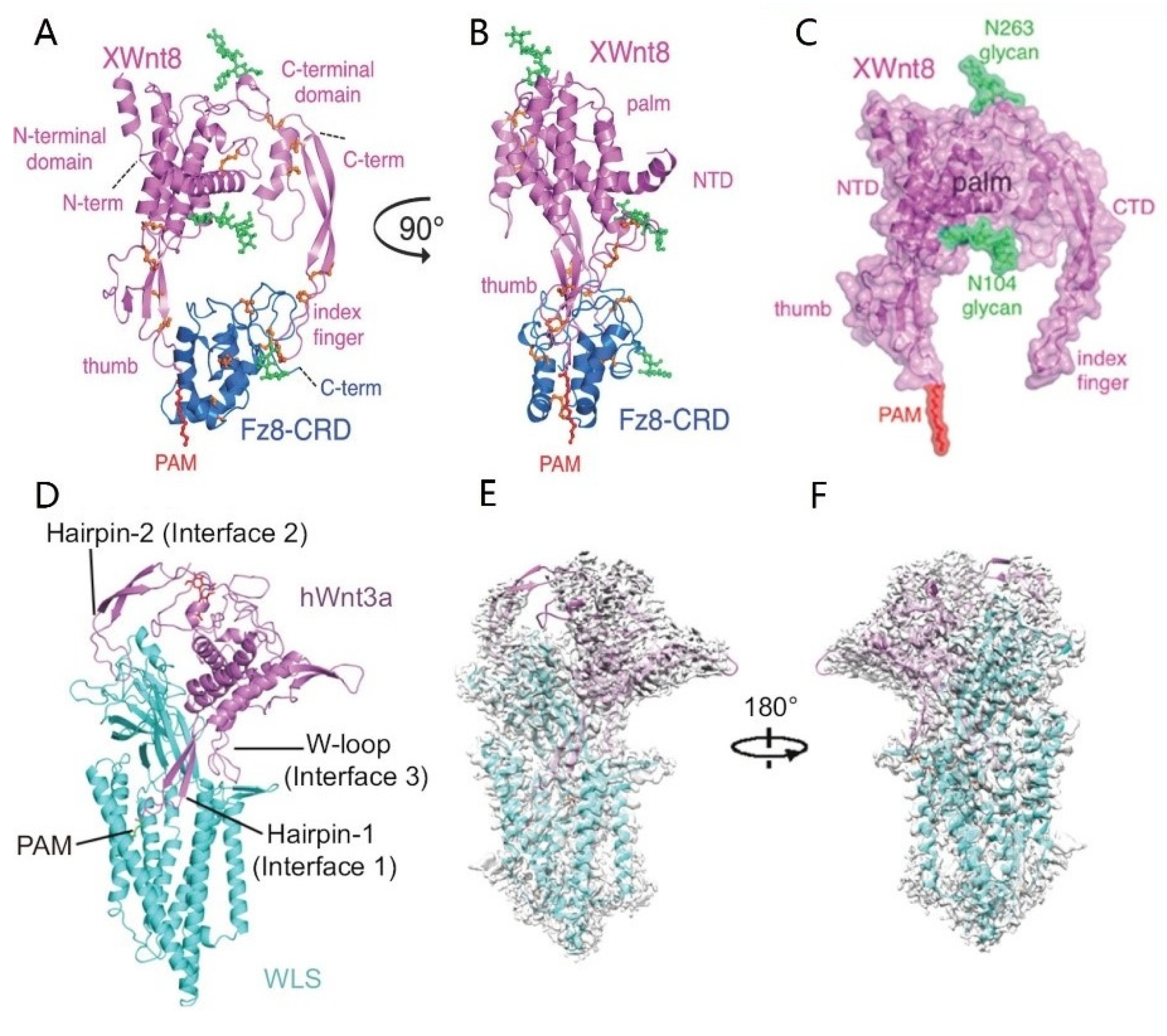
- Wnt proteins
- (2)
- β-catenin
- (3)
- Mechanisms of Wnt/β signal transduction
3.2. Wnt/Ca2+ Pathway
3.3. Wnt/Polarity Pathway (PCP)
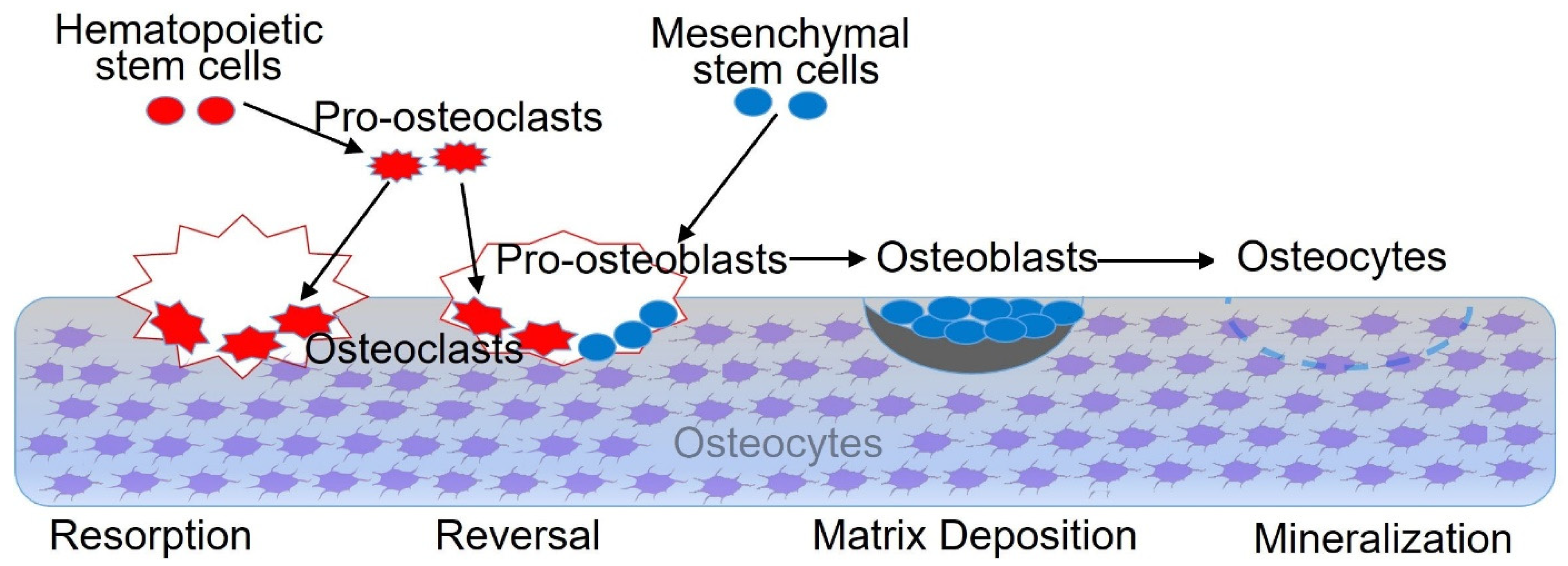
4. Wnt Signaling Pathway and Bone-Related Cells
4.1. Wnt Signaling Pathway and BMSCs
4.2. Wnt Signaling Pathway and Osteoblasts
4.3. Wnt Signaling Pathway and Osteoclasts
5. Osteoporosis and Wnt Signaling Pathway
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Financial Interest Statement
Abbreviations
References
- Gonzalez Rodriguez, E.; Debrach-Schneider, A.C.; Lamy, O. Osteoporosis. Rev. Med. Suisse. 2022, 18, 764–765. [Google Scholar]
- Anam, A.K.; Insogna, K. Update on Osteoporosis Screening and Management. Med. Clin. North. Am. 2021, 105, 1117–1134. [Google Scholar] [CrossRef]
- García-Gómez, M.C.; Vilahur, G. Osteoporosis and vascular calcification: A shared scenario. Clin. Investig. Arterioscler. 2020, 32, 33–42. [Google Scholar] [CrossRef]
- Watts, N.B.; Camacho, P.M.; Lewiecki, E.M.; Petak, S.M. American Association of Clinical Endocrinologists/American College of Endocrinology Clinical Practice Guidelines for the Diagnosis and Treatment of Postmenopausal Osteoporosis-2020 Update. Endocr. Pract. 2021, 27, 379–380. [Google Scholar] [CrossRef]
- Song, S.; Guo, Y.; Yang, Y.; Fu, D. Advances in pathogenesis and therapeutic strategies for osteoporosis. Pharmacol. Ther. 2022, 237, 108168. [Google Scholar] [CrossRef] [PubMed]
- Bolamperti, S.; Villa, I.; Rubinacci, A. Bone remodeling: An operational process ensuring survival and bone mechanical competence. Bone Res. 2022, 10, 48. [Google Scholar] [CrossRef]
- Marie, P.J. Bone cell-matrix protein interactions. Osteoporos Int. 2009, 20, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Wu, R.; Cao, L.; Gu, H.; Chai, F. Trelagliptin stimulates osteoblastic differentiation by increasing runt-related transcription factor 2 (RUNX2): A therapeutic implication in osteoporosis. Bioengineered 2021, 12, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Vescini, F.; Chiodini, I.; Falchetti, A.; Palermo, A.; Salcuni, A.S.; Bonadonna, S.; Geronimo, V.; Cesareo, R.; Giovanelli, L.; Brigo, M.; et al. Management of Osteoporosis in Men: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 13640. [Google Scholar] [CrossRef]
- Sirikul, W.; Siri-Angkul, N.; Chattipakorn, N.; Chattipakorn, S. Fibroblast Growth Factor 23 and Osteoporosis: Evidence from Bench to Bedside. Int. J. Mol. Sci. 2022, 23, 2500. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Sheng, M.; Lau, K.; Wilson, S.M.; Wongworawat, M.D.; Tang, X.; Ghahramanpouri, M.; Nehme, A.; Xu, Y.; Abdipour, A.; et al. Calcium released by osteoclastic resorption stimulates autocrine/paracrine activities in local osteogenic cells to promote coupled bone formation. Am. J. Physiol. Cell. Physiol. 2022, 322, C977–C990. [Google Scholar] [CrossRef]
- Ong, C.H.; Tham, C.L.; Harith, H.H.; Firdaus, N.; Israf, D.A. TGF-β-induced fibrosis: A review on the underlying mechanism and potential therapeutic strategies. Eur. J. Pharmacol. 2021, 911, 174510. [Google Scholar] [CrossRef]
- Li, S.N.; Wu, J.F. TGF-β/SMAD signaling regulation of mesenchymal stem cells in adipocyte commitment. Stem. Cell. Res. Ther. 2020, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Kassem, M.; Kveiborg, M.; Eriksen, E.F. Production and action of transforming growth factor-β in human osteoblast cultures: Dependence on cell differentiation and modulation by calcitriol. Eur. J. Clin. Investig. 2000, 30, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.F.; Li, Y.P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lieshout, E.M.; Alt, V. Bone graft substitutes and bone morphogenetic proteins for osteoporotic fractures: What is the evidence? Injury 2016, 47, S43–S46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumner, D.R.; Turner, T.M.; Cohen, M.; Losavio, P.; Urban, R.M.; Nichols, E.H.; McPherson, J.M. Aging Does Not Lessen the Effectiveness of TGFβ2-Enhanced Bone Regeneration. J. Bone Miner. Res. 2003, 18, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.A.; Liu, X.G.; Huo, J.Z.; Jiang, C.; Wen, X.J.; Chen, Z.R. Novel gene-modified-tissue engineering of cartilage using stable transforming growth factor-β1-transfected mesenchymal stem cells grown on chitosan scaffolds. J. Biosci. Bioeng. 2007, 103, 547–556. [Google Scholar] [CrossRef]
- Aslani, S.; Abhari, A.; Sakhinia, E.; Sanajou, D. Rajabi, H.; Rahimzadeh, S. Interplay between microRNAs and Wnt, transforming growth factor-β, and bone morphogenic protein signaling pathways promote osteoblastic differentiation of mesenchymal stem cells. J. Cell Physiol. 2019, 234, 8082–8093. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Park, H.; Lee, Y.L.; Oh, Y.; Park, J.; Kim, S.Y.; Weon, S.; Choi, S.H.; Yang, J.H.; Jo, S.; et al. Tgfβ Suppressed Matrix Mineralization of Osteoblasts Differentiation by Regulating Smurf-C Ebpβ-Dkk Axis. Int. J. Mol. Sci. 2020, 21, 9771. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, T.; Pagnotti, G.M.; Guise, T.A.; Mohammad, K.S. The Role of TGF-β in Bone Metastases. Biomolecules 2021, 11, 1643. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Xu, W.; Zhang, Z.; Jin, H.; Yang, Y.; Zhang, J.; Xu, H. Role of TGF-β1 in Fluoride-Treated Osteoblasts at Different Stages. Biol. Trace. Elem. Res. 2022, 200, 740–748. [Google Scholar] [CrossRef]
- Garg, P.; Mazur, M.M.; Buck, A.C.; Wandtke, M.E.; Liu, J.; Ebraheim, N.A. Prospective Review of Mesenchymal Stem Cells Differentiation into Osteoblasts. Orthop. Surg. 2017, 9, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ge, L.; Zhao, Y.; Zhai, Y.; Rao, N.; Yuan, X.; Yang, J.; Li, J.; Yu, S. TGF-β2 and TGF-β1 differentially regulate the odontogenic and osteogenic differentiation of mesenchymal stem cells. Arch. Oral. Biol. 2022, 135, 105357. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Oh, Y.; Jo, S.; Kim, T.H.; Ji, J.D. A dual role of TGF-β in human osteoclast differentiation mediated by Smad1 versus Smad3 signaling. Immunol. Lett. 2019, 206, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.M.W.; Itoh, K.; Udagawa, N.; Häusler, K.; Yasuda, H.; Shima, N.; Mizuno, A.; Higashio, K.; Takahashi, N.; Suda, T.; et al. Transforming Growth Factor β Affects Osteoclast Differentiation via Direct and Indirect Actions. J. Bone. Miner. Res. 2001, 16, 1787–1794. [Google Scholar] [CrossRef]
- Dai, G.M.; Ren, L.; Chen, H.; Liu, W.; Chen, Y.; He, X.Q.; Liu, W.; Tu, X.L.; Huang, W. Down-regulation of osteocytic TGF-β/Smad4 inhibits the osteoblastic and osteoclastic differentiation in mouse BMSCs. Basic Clin. Med. 2017, 37, 786–791. [Google Scholar]
- Li, H.; Fan, J.; Fan, L.; Li, T.; Yang, Y.; Xu, H.; Deng, L.; Li, J.; Li, T.; Weng, X.; et al. MiRNA-10b Reciprocally Stimulates Osteogenesis and Inhibits Adipogenesis Partly through the TGF-β/SMAD2 Signaling Pathway. Aging Dis. 2018, 9, 1058–1073. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Chen, R.; Wang, G.; Zhang, Y.; Liu, F. Exosomes derived from mesenchymal stem cells reverse EMT via TGF-β1/Smad pathway and promote repair of damaged endometrium. Stem. Cell. Res. Ther. 2019, 10, 225. [Google Scholar] [CrossRef]
- Ye, W.; Yang, Z.; Cao, F.; Li, H.; Zhao, T.; Zhang, H.; Zhang, Z.; Yang, S.; Zhu, J.; Liu, Z.; et al. Articular cartilage reconstruction with TGF-β1-simulating self-assembling peptide hydrogel-based composite scaffold. Acta Biomater. 2022, 146, 94–106. [Google Scholar] [CrossRef]
- Panahipour, L.; Omerbasic, A.; Nasirzade, J.; Gruber, R. TGF-β Activity of a Demineralized Bone Matrix. Int. J. Mol. Sci. 2021, 22, 664. [Google Scholar] [CrossRef]
- Mazziotta, C.; Lanzillotti, C.; Iaquinta, M.; Taraballi, F.; Torreggiani, E.; Rotondo, J.; Otòn-Gonzalez, L.; Mazzoni, E.; Frontini, F.; Bononi, I.; et al. MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 2362. [Google Scholar] [CrossRef] [PubMed]
- Heubel, B.; Nohe, A. The Role of BMP Signaling in Osteoclast Regulation. J. Dev. Biol. 2021, 9, 24. [Google Scholar] [CrossRef]
- Urist, M.R. Bone: Formation by Autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.M.; Rosen, V.; Celeste, A.J.; Mitsock, L.M.; Whitters, M.J.; Kriz, R.W.; Hewick, R.M.; Wang, E.A. Novel Regulators of Bone Formation: Molecular Clones and Activities. Science 1988, 242, 1528–1834. [Google Scholar] [CrossRef]
- Sconocchia, T.; Sconocchia, G. Regulation of the Immune System in Health and Disease by Members of the Bone Morphogenetic Protein Family. Front. Immunol. 2021, 12, 802346. [Google Scholar] [CrossRef]
- Xie, X.D.; Zhao, L.; Wu, Y.F.; Wang, J. Role of bone morphogenetic protein 1/tolloid proteinase family in the development of teeth and bone. Yi Xue Za Zhi 2020, 38, 589–593. [Google Scholar]
- Tajer, B.; Dutko, J.; Little, S.; Mullins, M. BMP heterodimers signal via distinct type I receptor class functions. Proc. Natl. Acad. Sci. USA 2021, 118, e201795211. [Google Scholar] [CrossRef]
- Karim, M.S.; Madamanchi, A.; Dutko, J.A.; Mullins, M.C.; Umulis, D.M. Heterodimer-heterotetramer formation mediates enhanced sensor activity in a biophysical model for BMP signaling. PLoS Comput. Biol. 2021, 17, e1009422. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, M.; Jayasuriya, C.T.; Peng, H.; Zhang, L.; Guan, Y.; Froehlich, J.A.; Terek, R.M.; Chen, Q. Human osteoarthritis cartilage-derived stromal cells activate joint degeneration through TGF-beta lateral signaling. FASEB J. 2020, 34, 16552–16566. [Google Scholar] [CrossRef]
- Song, D.; He, G.; Shi, Y.; Ni, J.; Long, F. Functional interaction between Wnt and Bmp signaling in periosteal bone growth. Sci. Rep. 2021, 11, 10782. [Google Scholar] [CrossRef]
- Biswas, T.; Jaswal, A.F.; Yadav, U.F.; Bandyopadhyay, A. Simultaneous differentiation of articular and transient cartilage: WNT-BMP interplay and its therapeutic implication. Int. J. Dev. Biol. 2020, 64, 203–211. [Google Scholar] [CrossRef]
- Zhang, R.; Oyajobi, B.O.; Harris, S.E.; Chen, D.; Tsao, C.; Deng, H.; Zhao, M. Wnt/β-catenin signaling activates bone morphogenetic protein 2 expression in osteoblasts. Bone 2013, 52, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Langton, P.F.; Kakugawa, S.; Vincent, J. Making, Exporting, and Modulating Wnts. Trends Cell Biol. 2016, 26, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Q.; Wang, N.X.; Luo, Y.; Yu, C.Y.; Xiao, J.H. Ganoderal A effectively induces osteogenic differentiation of human amniotic mesenchymal stem cells via cross-talk between Wnt/β-catenin and BMP/SMAD signaling pathways. Biomed. Pharmacother. 2020, 123, 109807. [Google Scholar] [CrossRef]
- Tang, C.; Wu, M.; Zhao, D.; Edwards, D.; McVicar, A.; Luo, Y.; Zhu, G.; Wang, Y.; Zhou, H.D.; Chen, W.; et al. Runx1 is a central regulator of osteogenesis for bone homeostasis by orchestrating BMP and WNT signaling pathways. PLoS Genet. 2021, 17, e1009233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Li, H.N.; Chen, F.; Chen, Y.P.; Chai, Y.; Liao, J.Z.; Gan, B.; Chen, D.P.; Li, S.; Liu, Y.Q. Icariin regulates miR-23a-3p-mediated osteogenic differentiation of BMSCs via BMP-2/Smad5/Runx2 and WNT/β-catenin pathways in osteonecrosis of the femoral head. Saudi Pharm. J. 2021, 29, 1405–1415. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Wang, J.H. Bone morphogenetic protein-2, -4, -6, -7 and -9 differentially mediated osteogenic differentiation of immortalized odontoblasts. Shanghai Kou Qiang 2020, 29, 242–249. [Google Scholar]
- Tang, N.; Song, W.; Luo, J.; Luo, X.; Chen, J.; Sharff, K.A.; Bi, Y.; He, B.; Huang, J.; Zhu, G.; et al. BMP-9-induced osteogenic differentiation of mesenchymal progenitors requires functional canonical Wnt/β-catenin signalling. J. Cell. Mol. Med. 2009, 13, 2448–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Liu, Y.; Wang, A.; Zhu, Z.; Li, Y.; Zhu, C.; Che, Z.; Liu, T.; Liu, H.; Huang, L. Application of BMP in Bone Tissue Engineering. Front. Bioeng. Biotechnol. 2022, 10, 810880. [Google Scholar] [CrossRef]
- Pérez-Lozano, M.; Sudre, L.; van Eegher, S.; Citadelle, D.; Pigenet, A.; Lafage-Proust, M.H.; Pastoureau, P.; Ceuninck, F.; Berenbaum, F.; Houard, X. Gremlin-1 and BMP-4 Overexpressed in Osteoarthritis Drive an Osteochondral-Remodeling Program in Osteoblasts and Hypertrophic Chondrocytes. Int. J. Mol. Sci. 2022, 23, 2084. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, G.; Gong, Y.; Li, B.; Li, W.; Liu, D.; Yang, X. BMP-2 promotes osteogenic differentiation of mesenchymal stem cells by enhancing mitochondrial activity. J. Musculoskelet. Neuronal Interact. 2022, 22, 123–131. [Google Scholar] [PubMed]
- Kuwahara, M.; Akasaki, Y.; Goto, N.; Kurakazu, I.; Sueishi, T.; Toya, M.; Uchida, T.; Tsutsui, T.; Hirose, R.; Tsushima, H.; et al. Fluvastatin promotes chondrogenic differentiation of adipose-derived mesenchymal stem cells by inducing bone morphogenetic protein. BMC Pharmacol. Toxicol. 2022, 23, 61. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell. Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef] [Green Version]
- Claus, S.; Aubert-Foucher, E.; Demoor, M.; Camuzeaux, B.; Paumier, A.; Piperno, M.; Damour, O.; Duterque-Coquillaud, M.; Galéra, P.; Mallein-Gerin, F. Chronic exposure of bone morphogenetic protein-2 favors chondrogenic expression in human articular chondrocytes amplified in monolayer cultures. J. Cell. Biochem. 2010, 111, 1642–1651. [Google Scholar] [CrossRef]
- Zolbin, M.; Ersoy, G.S.; Aliakbari, F.; Amidi, F.; Daghigh, F.; Abbasi, M.; Johnson, J. Basal characterization and in vitro differentiation of putative stem cells derived from the adult mouse ovary. In Vitro Cell. Dev. Biol. Anim. 2020, 56, 59–66. [Google Scholar] [CrossRef]
- Steinert, A.F.; Proffen, B.; Kunz, M.; Hendrich, C.; Ghivizzani, S.C.; Nöth, U.; Rethwilm, A.; Eulert, J.; Evans, C.H. Hypertrophy is induced during the in vitrochondrogenic differentiation of human mesenchymal stem cells by bone morphogenetic protein-2 and bone morphogenetic protein-4 gene transfer. Arthritis Res. Ther. 2009, 11, R148. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wong, S.W.; Han, D.; Cai, T. Genetic analysis: Wnt and other pathways in nonsyndromic tooth agenesis. Oral Dis. 2019, 25, 646–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ran, S.; Liu, B.; Gu, S. Monitoring of canonical BMP and Wnt activities during postnatal stages of mouse first molar root formation. J. Appl. Oral. Sci. 2021, 13, e20210281. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Pan, Y.; Ji, J.J.; Xu, Y.X.; Zhang, Q.Y.; Qin, L.P. Roles and action mechanisms of WNT4 in cell differentiation and human diseases: A review. Cell Death Discov. 2021, 7, 287. [Google Scholar] [CrossRef]
- Halloran, D.; Durbano, H.W.; Nohe, A. Bone Morphogenetic Protein-2 in Development and Bone Homeostasis. J. Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef]
- Chhabra, S.; Warmflash, A. BMP-treated human embryonic stem cells transcriptionally resemble amnion cells in the monkey embryo. Biol. Open 2021, 10, 58617. [Google Scholar] [CrossRef]
- Tóth, F.; Tőzsér, J.; Hegedűs, C. Effect of Inducible BMP-7 Expression on the Osteogenic Differentiation of Human Dental Pulp Stem Cells. Int. J. Mol. Sci. 2021, 22, 6182. [Google Scholar] [CrossRef]
- Si, W.; Kang, Q.; Luu, H.H.; Park, J.K.; Luo, Q.; Song, W.X.; Jiang, W.; Luo, X.; Li, X.; Yin, H. CCN1/Cyr61 Is Regulated by the Canonical Wnt Signal and Plays an Important Role in Wnt3A-Induced Osteoblast Differentiation of Mesenchymal Stem Cells. Mol. Cell. Biol. 2006, 26, 2955–2964. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, Z.; Cheng, Y.; Kardami, E.; Loh, Y.P. Low and High Molecular Weight FGF-2 Have Differential Effects on Astrocyte Proliferation, but Are Both Protective Against Aβ-Induced Cytotoxicity. Front. Mol. Neurosci. 2019, 12, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, P.; Saikia, B.B.; Sonnaila, S.; Agrawal, S.; Alraawi, Z.; Kumar, T.K.S.; Iyer, S. The Saga of Endocrine FGFs. Cells 2021, 10, 2418. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Sato, R. Endocrine Fibroblast Growth Factors in Relation to Stress Signaling. Cells 2022, 11, 505. [Google Scholar] [CrossRef]
- Chen, H.; Cui, Y.; Zhang, D.; Xie, J.; Zhou, X. The role of fibroblast growth factor 8 in cartilage development and disease. J. Cell. Mol. Med. 2022, 26, 990–999. [Google Scholar] [CrossRef]
- Song, H.; Du, H.; Li, J.; Wang, M.; Wang, J.; Ju, X.; Mu, W. Effect of fibroblast growth factor 2 on degenerative endplate chondrocyte: From anabolism to catabolism. Exp. Mol. Pathol. 2021, 118, 104590. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Kaji, Y.; Nakamura, O.; Tobiume, S.; Nomura, Y.; Oka, K.; Yamamoto, T. Bone Union Enhancement by bFGF-Containing HAp/Col in Prefabricated Vascularized Allo-Bone Grafts. J. Reconstr. Microsurg. 2021, 37, 346–352. [Google Scholar] [CrossRef]
- Antunes, B.P.; Vainieri, M.L.; Alini, M.; Monsonego-Ornan, E.; Grad, S.; Yayon, A. Enhanced chondrogenic phenotype of primary bovine articular chondrocytes in Fibrin-Hyaluronan hydrogel by multi-axial mechanical loading and FGF. Acta Biomaterialia 2020, 105, 170–179. [Google Scholar] [CrossRef]
- Maija, P.V.; Teuvo, H.; Qiang, Q.; Eeva, M.V.; Anna, H.; Jani, A.; Seppänen, H.; Kalervo, V.; Pirkko, L.H. Regulation of osteoblast differentiation: A novel function for fibroblast growth factor 8. Endocrinology 2006, 147, 2171–2182. [Google Scholar]
- Hayes, A.J.; Whitelock, J.; Melrose, J. Regulation of FGF-2, FGF-18 and Transcription Factor Activity by Perlecan in the Maturational Development of Transitional Rudiment and Growth Plate Cartilages and in the Maintenance of Permanent Cartilage Homeostasis. Int. J. Mol. Sci. 2022, 23, 1934. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.; Emani, C.; Kochan, A.; Ghebrehawariat, K.; Tyburski, J.; Johnston, M.; Rabago, D. Prolotherapy agent P2G is associated with upregulation of fibroblast growth factor-2 genetic expression in vitro. J. Exp. Orthop. 2020, 7, 97. [Google Scholar] [CrossRef]
- Fei, Y.; Hurley, M.M. Role of fibroblast growth factor 2 and wnt signaling in anabolic effects of parathyroid hormone on bone formation. J. Cell. Physiol. 2012, 227, 3539–3545. [Google Scholar] [CrossRef] [Green Version]
- Charoenlarp, P.; Rajendran, A.K.; Iseki, S. Role of fibroblast growth factors in bone regeneration. Inflamm. Regen. 2017, 37, 10. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Tensho, K.; Nakaya, H.; Nawata, M.; Okabe, T.; Wakitani, S. Low dose fibroblast growth factor-2 (FGF-2) enhances bone morphogenetic protein-2 (BMP-2)-induced ectopic bone formation in mice. Bone 2005, 36, 399–407. [Google Scholar] [CrossRef]
- Kodama, N.; Nagata, M.; Tabata, Y.; Ozeki, M.; Ninomiya, T.; Takagi, R. A local bone anabolic effect of rhFGF2-impregnated gelatin hydrogel by promoting cell proliferation and coordinating osteoblastic differentiation. Bone 2009, 44, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Sakano, S.; Hasegawa, Y.; Murata, Y.; Ito, T.; Genda, E.; Iwata, H.; Ishiguro, N.; Seo, H. Inhibitory effect of bFGF on endochondral heterotopic ossification. Biochem. Biophys. Res. Commun. 2002, 293, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Homer-Bouthiette, C.; Doetschman, T.; Xiao, L.; Hurley, M.M. Knockout of Nuclear High Molecular Weight FGF2 Isoforms in Mice Modulates Bone and Phosphate Homeostasis. J. Biol. Chem. 2014, 289, 36303–36314. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Yang, P.; Liu, P.; Wang, H.; Ke, E.; Li, K.; Yan, H. Targeting Filamin A alleviates ovariectomy-induced bone loss in mice via the WNT/β-catenin signaling pathway. Cell Signal 2022, 90, 110191. [Google Scholar] [CrossRef]
- Balmukhanova, A.; Kabulbayev, K.; Alpay, H.; Kanatbayeva, A.; Balmukhanova, A. FGF-23 and Phosphate in Children with Chronic Kidney Disease: A Cross-Sectional Study in Kazakhstan. Medicina 2021, 57, 15. [Google Scholar] [CrossRef]
- Miyakawa, H.; Hsu, H.H.; Ogawa, M.; Akabane, R.; Miyagawa, Y.; Takemura, N. Association between serum fibroblast growth factor-23 concentration and development of hyperphosphatemia in normophosphatemic dogs with chronic kidney disease. J. Vet. Intern. Med. 2021, 35, 2296–2305. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Sánchez, S.; Poveda, J.; Navarro-García, J.A.; González-Lafuente, L.; Rodríguez-Sánchez, E.; Ruilope, L.M.; Ruiz-Hurtado, G. An Overview of FGF-23 as a Novel Candidate Biomarker of Cardiovascular Risk. Front. Physiol. 2021, 12, 632260. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, H.; Sun, W.; Bai, X.; Karaplis, A.C.; Goltzman, D.; Miao, D. Fibroblast growth factor 23 overexpression impacts negatively on dentin mineralization and dentinogenesis in mice. Clin. Exp. Pharmacol. Physiol. 2011, 38, 395–402. [Google Scholar] [CrossRef]
- Lv, Z.T. The Role and Mechanism of FGF23 on Mineralization of Chicken Osteoblasts. Master’s Thesis, Shandong Agricultural University, Taian, China, 2020. [Google Scholar]
- Kawai, M.; Kinoshita, S.; Kimoto, A.; Hasegawa, Y.; Miyagawa, K. FGF23 suppresses chondrocyte proliferation in the presence of soluble α-klotho both in vitro and in vivo. J. Biol. Chem. 2013, 288, 2414–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Sato, T.; Densmore, M.; Saito, H.; Schüler, C.; Erben, R.G.; Lanske, B. FGF-23/Klotho signaling is not essential for the phosphaturic and anabolic functions of PTH. J. Bone. Miner. Res. 2011, 26, 2026–2035. [Google Scholar] [CrossRef] [Green Version]
- Knowles, H.J. Hypoxia-Induced Fibroblast Growth Factor 11 Stimulates Osteoclast-Mediated Resorption of Bone. Calcif. Tissue Int. 2017, 100, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Kettunen, P.; Furmanek, T.; Chaulagain, R.; Kvinnsland, I.; Luukko, K. Developmentally regulated expression of intracellular Fgf11-13, hormone-like Fgf15 and canonical Fgf16, -17 and -20 mRNAs in the developing mouse molar tooth. Acta. Odontol. Scand. 2011, 69, 360–366. [Google Scholar] [CrossRef]
- Sieber, S.; Gigout, A. Sprifermin (recombinant human FGF18) is internalized through clathrin- and dynamin-independent pathways and degraded in primary chondrocytes. Exp. Cell Res. 2020, 395, 112236. [Google Scholar] [CrossRef]
- Xie, Y.; Zinkle, A.; Chen, L.; Mohammadi, M. Fibroblast growth factor signalling in osteoarthritis and cartilage repair. Nat. Rev. Rheumatol. 2020, 16, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.; Bossaer, J.B. Fibroblast growth factor receptor (FGFR) inhibitors: A review of a novel therapeutic class. J. Oncol. Pharm. Pract. 2021, 27, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Liyis, B.G.; Nolan, J.; Maharjana, M.A. Fibroblast growth factor receptor 1-bound extracellular vesicle as novel therapy for osteoarthritis. Biomedicine 2022, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Shin, H.R.; Kim, H.J.; Yoon, H.; Cho, Y.D.; Choi, K.Y.; Choi, J.Y.; Kim, W.J.; Ryoo, H.M. Septal chondrocyte hypertrophy contributes to midface deformity in a mouse model of Apert syndrome. Sci. Rep. 2021, 11, 7979. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.H.; Zhang, M.Z.; Lai, C.Z.; Li, C.C.; Du, L.; Song, G.D.; Zong, X.L.; Jin, X.L. Dura cells in the etiopathogenesis of Crouzon syndrome: The effects of FGFR2 mutations in the dura cells on the proliferation of osteoblasts through the hippo/YAP mediated transcriptional regulation pathway. Am. J. Transl. Res. 2021, 13, 11255–11270. [Google Scholar] [PubMed]
- Wang, X.; He, J.; Wang, H.; Zhao, D.; Geng, B.; Wang, S.; An, J.; Wang, C.; Han, H.; Xia, Y. Fluid shear stress regulates osteoblast proliferation and apoptosis via the lncRNA TUG1/miR-34a/FGFR1 axis. J. Cell. Mol. Med. 2021, 25, 8734–8747. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Tao, D.Y.; Zhang, J.J.; Niu, H.H.; Luo, J.F.; Cheng, S.Q. Clinical features and FGFR3 mutations of children with achondroplasia. Zhongguo Dang Dai Er Ke Za Zhi 2022, 24, 405–410. [Google Scholar]
- Savarirayan, R.; De Bergua, J.M.; Arundel, P.; McDevitt, H.; Cormier-Daire, V.; Saraff, V.; Skae, M.; Delgado, B.; Leiva-Gea, A.; Santos-Simarro, F.; et al. Infigratinib in children with achondroplasia: The PROPEL and PROPEL 2 studies. Ther. Adv. Musculoskelet. Dis. 2022, 14, 1759720X221084848. [Google Scholar] [CrossRef]
- Ea, C.; Hennocq, Q.; Picard, A.; Polak, M.; Collet, C.; Legeai-Mallet, L.; Arnaud, É.; Paternoster, G.; Khonsari, R.H. Growth charts in FGFR2- and FGFR3-related faciocraniosynostoses. Bone Rep. 2022, 16, 101524. [Google Scholar] [CrossRef]
- Scheers, E.; Borgmans, C.; Keung, C.; Bohets, H.; Wynant, I.; Poggesi, I.; Cuyckens, F.; Leclercq, L.; Mamidi, R. Metabolism and disposition in rats, dogs, and humans of erdafitinib, an orally administered potent pan-fibroblast growth factor receptor (FGFR) tyrosine kinase inhibitor. Xenobiotica 2021, 51, 177–193. [Google Scholar] [CrossRef]
- Collin-Osdoby, P.; Rothe, L.; Bekker, S.; Anderson, F.; Huang, Y.; Osdoby, P. Basic Fibroblast Growth Factor Stimulates Osteoclast Recruitment, Development, and Bone Pit Resorption in Association With Angiogenesis In Vivo on the Chick Chorioallantoic Membrane and Activates Isolated Avian Osteoclast Resorption In Vitro. J. Bone. Miner. Res. 2002, 17, 1859–1871. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Su, N.; Yang, J.; Huang, W.; Li, C.; Zhao, L.; He, Q.; Du, X.; Shen, Y.; Chen, B.; et al. Fibroblast growth factor receptor 1 regulates the differentiation and activation of osteoclasts through Erk1/2 pathway. Biochem. Biophys. Res. Commun. 2009, 390, 494–499. [Google Scholar] [CrossRef]
- Okada, Y.; Montero, A.; Zhang, X.; Sobue, T.; Lorenzo, J.; Doetschman, T.; Coffin, J.D.; Hurley, M.M. Impaired Osteoclast Formation in Bone Marrow Cultures of Fgf2 Null Mice in Response to Parathyroid Hormone *. J. Biol. Chem. 2003, 278, 21258–21266. [Google Scholar] [CrossRef] [Green Version]
- Su, N.; Li, X.; Tang, Y.; Yang, J.; Wen, X.; Guo, J.; Tang, J.; Du, X.; Chen, L. Deletion of FGFR3 in Osteoclast Lineage Cells Results in Increased Bone Mass in Mice by Inhibiting Osteoclastic Bone Resorption. J. Bone Miner. Res. 2016, 31, 1676–1687. [Google Scholar] [CrossRef] [Green Version]
- Bayle, A.; Martin-Romano, P.; Loriot, Y. FIGHT against FGF/FGFR alterations: What are the next steps? Ann. Oncol. 2022, 33, 460–462. [Google Scholar] [CrossRef]
- Vuong, L.T.; Mlodzik, M. Different strategies by distinct Wnt-signaling pathways in activating a nuclear transcriptional response. Curr. Top. Dev. Biol. 2022, 149, 59–89. [Google Scholar] [PubMed]
- Yang, R.B.; Lin, F.F.; Yang, J.; Chen, B.; Zhang, M.H.; Lu, Q.P.; Xiao, B.; Liu, Y.; Zheng, K.; Qiu, Y.R. Retraction Note: Overexpression of CAV3 facilitates bone formation via the Wnt signaling pathway in osteoporotic rats. Endocrine 2022, 76, 751. [Google Scholar] [CrossRef]
- Chagay, N.B.; Khayt, G.Y.; Vdovina, T.M.; Shaforost, A.A. Cystic fibrosis being a polyendocrine disease. Probl. Endokrinol. 2021, 67, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, J.; Fu, J.; Li, S.; Chai, Z.; Sun, Y. Associations between WNT signaling pathway-related gene polymorphisms and risks of osteoporosis development in Chinese postmenopausal women: A case-control study. Climacteric 2022, 25, 257–263. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Gavin, B.J.; McMahon, J.A.; McMahon, A.P. Expression of multiple novel Wnt-1/int-1-related genes during fetal and adult mouse development. Genes Dev. 1990, 4, 2319–2332. [Google Scholar] [CrossRef]
- Janda, C.Y.; Waghray, D.; Levin, A.; Thomas, C.; Garcia, K.C. Structural basis of Wnt recognition by Frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Q.; Zhao, Y.; Ye, F.; Xiao, Z.; Huang, G.; Xu, M.; Zhang, Y.; Zhan, X.; Sun, K.; Wang, Z.; et al. Cryo-EM structure of human Wntless in complex with Wnt3a. Nat. Commun. 2021, 12, 4541. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Manzoor, M.; Hussain, A. Wnt signaling pathway: A comprehensive review. Cell Biol. Int. 2022, 46, 863–877. [Google Scholar] [CrossRef]
- Bänziger, C.; Soldini, D.; Schütt, C.; Zipperlen, P.; Hausmann, G.; Basler, K. Wntless, a Conserved Membrane Protein Dedicated to the Secretion of Wnt Proteins from Signaling Cells. Cell 2006, 125, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Amjadi-Moheb, F.; Akhavan-Niaki, H. Wnt signaling pathway in osteoporosis: Epigenetic regulation, interaction with other signaling pathways, and therapeutic promises. J. Cell. Physiol. 2019, 234, 14641–14650. [Google Scholar] [CrossRef] [PubMed]
- Teufel, S.; Hartmann, C. Wnt-signaling in skeletal development. Curr. Top. Dev. Biol. 2019, 133, 235–279. [Google Scholar]
- Visweswaran, M.; Pohl, S.; Arfuso, F.; Newsholme, P.; Dilley, R.; Pervaiz, S.; Dharmarajan, A. Multi-lineage differentiation of mesenchymal stem cells—To Wnt, or not Wnt. Int. J. Biochem. Cell. Biol. 2015, 68, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.F.; Shao, J.; Shi, C.J.; Li, Z.P.; Fu, W.M.; Zhang, J.F. Apigenin promotes osteogenic differentiation of mesenchymal stem cells and accelerates bone fracture healing via activating Wnt/β-catenin signaling. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E760–E771. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, X.; Zhou, R.; Song, D.; Jiang, Y.; Xue, W. Chaetocin Promotes Osteogenic Differentiation via Modulating Wnt/Beta-Catenin Signaling in Mesenchymal Stem Cells. Stem. Cells Int. 2021, 2021, 8888416. [Google Scholar] [CrossRef]
- Lademann, F.; Tsourdi, E.; Hofbauer, L.C.; Rauner, M. Thyroid Hormone Actions and Bone Remodeling-The Role of the Wnt Signaling Pathway. Exp. Clin. Endocrinol. Diabetes 2020, 128, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.; Deng, F.; Huang, E.; Yan, Z.; Wang, Z.; Deng, Y.; Zhang, Q.; Zhang, Z.; Ye, J. Canonical Wnt signaling acts synergistically on BMP9-induced osteo/odontoblastic differentiation of stem cells of dental apical papilla (SCAPs). Biomaterials 2015, 39, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Liang, K.; Du, Y.; Chen, L.; Wang, L.; Li, R.; Yan, Z.; Liu, Y. Contrary Roles of Wnt/β-Catenin Signaling in BMP9-Induced Osteogenic and Adipogenic Differentiation of 3T3-L1 Preadipocytes. Cell. Biochem. Biophys. 2020, 78, 347–356. [Google Scholar] [CrossRef]
- Bilkovski, R.; Schulte, D.M.; Oberhauser, F.; Gomolka, M.; Udelhoven, M.; Hettich, M.M.; Roth, B.; Heidenreich, A.; Gutschow, C.; Krone, W.; et al. Role of Wnt-5a in the Determination of Human Mesenchymal Stem Cells into Preadipocytes. J. Biol. Chem. 2010, 285, 6170–6178. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.; Fromigué, O.; Dieudonné, F.X.; Marty, C.; Chen, J.; Dahan, J.; Wei, Y.; Marie, P.J. The LIM-only protein FHL2 controls mesenchymal cell osteogenic differentiation and bone formation through Wnt5a and Wnt10b. Bone 2013, 53, 6–12. [Google Scholar] [CrossRef]
- He, X.; Wang, H.; Jin, T.; Xu, Y.; Mei, L.; Yang, J. TLR4 Activation Promotes Bone Marrow MSC Proliferation and Osteogenic Differentiation via Wnt3a and Wnt5a Signaling. PloS ONE 2016, 11, e0149876. [Google Scholar] [CrossRef]
- Okamoto, M.; Udagawa, N.; Uehara, S.; Maeda, K.; Yamashita, T.; Nakamichi, Y.; Kato, H.; Saito, N.; Minami, Y.; Takahashi, N.; et al. Noncanonical Wnt5a enhances Wnt/β-catenin signaling during osteoblastogenesis. Sci. Rep. 2014, 4, 4493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, V.; Nohno, T.; Linker, C.; Marcelle, C.; Francis, P. Wnt regulation of chondrocyte differentiation. J. Cell. Sci. 2002, 115, 4809–4818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padwal, M.; Liu, L.; Margetts, P.J. The role of WNT5A and Ror2 in peritoneal membrane injury. J. Cell. Mol. Med. 2020, 24, 3481–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Chan, R.W.S.; Lee, C.L.; Chiu, P.C.N.; Li, R.H.W.; Ng, E.H.Y.; Yeung, W.S.B. WNT5A Interacts With FZD5 and LRP5 to Regulate Proliferation and Self-Renewal of Endometrial Mesenchymal Stem-Like Cells. Front. Cell. Dev. Biol. 2022, 10, 837827. [Google Scholar] [CrossRef]
- Cawthorn, W.P.; Bree, A.J.; Yao, Y.; Du, B.; Hemati, N.; Martinez-Santibañez, G.; MacDougald, O.A. Wnt6, Wnt10a and Wnt10b inhibit adipogenesis and stimulate osteoblastogenesis through a β-catenin-dependent mechanism. Bone 2012, 50, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Kries, J.P.; Winbeck, G.; Asbrand, C.; Schwarz-Romond, T.; Sochnikova, N.; Dell’Oro, A.; Behrens, J.; Birchmeier, W. Hot spots in β-catenin for interactions with LEF-1, conductin and APC. Nat. Struct. Biol. 2000, 7, 800–807. [Google Scholar] [PubMed]
- Barth, A.I.M.; Näthke, I.S.; Nelson, W.J. Cadherins, catenins and APC protein: Interplay between cytoskeletal complexes and signaling pathways. Curr. Opin. Cell. Biol. 1997, 9, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.R.; Fagotto, F. The ins and outs of APC and β-catenin nuclear transport. EMBO Rep. 2002, 3, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Westendorf, J.J.; Kahler, R.A.; Schroeder, T.M. Wnt signaling in osteoblasts and bone diseases. Gene 2004, 341, 19–39. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, Y.; Jia, B.; Wang, Y.; Li, T. Icariin stimulates osteogenesis and suppresses adipogenesis of human bone mesenchymal stem cells via miR-23a-mediated activation of the Wnt/β-catenin signaling pathway. Phytomedicine 2021, 85, 153485. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, J.; Zhao, J.; Kuai, F.; Yang, H. Shikonin promotes osteogenesis and suppresses osteoclastogenesis in vitro. Am. J. Transl. Res. 2020, 12, 8099–8110. [Google Scholar]
- Dell’Accio, F.; De Bari, C.; Eltawil, N.M.; Vanhummelen, P.; Pitzalis, C. Identification of the molecular response of articular cartilage to injury, by microarray screening: Wnt-16 expression and signaling after injury and in osteoarthritis. Arthritis. Rheum. 2008, 58, 1410–1421. [Google Scholar] [CrossRef]
- Hui, T.; Zhou, Y.; Wang, T.; Li, J.; Zhang, S.; Liao, L.; Gu, J.; Ye, L.; Zhao, L.; Chen, D. Activation of β-catenin signaling in aggrecan-expressing cells in temporomandibular joint causes osteoarthritis-like defects. Int. J. Oral. Sci. 2018, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Tamamura, Y.; Otani, T.; Kanatani, N.; Koyama, E.; Kitagaki, J.; Komori, T.; Yamada, Y.; Costantini, F.; Wakisaka, S.; Pacifici, M.; et al. Developmental Regulation of Wnt/β-Catenin Signals Is Required for Growth Plate Assembly, Cartilage Integrity, and Endochondral Ossification. J. Biol. Chem. 2005, 280, 19185–19195. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Lyons, J.P.; Mori-Akiyama, Y.; Yang, X.; Zhang, R.; Zhang, Z.; Deng, J.M.; Taketo, M.M.; Nakamura, T.; Behringer, R.R.; et al. Interactions between Sox9 and beta-catenin control chondrocyte differentiation. Genes Dev. 2004, 18, 1072–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Mak, K.K.; Taketo, M.M.; Yang, Y. The Wnt/beta-catenin pathway interacts differentially with PTHrP signaling to control chondrocyte hypertrophy and final maturation. PLoS ONE 2009, 4, e6067. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Zhang, Z.; Yu, Y.; Chu, H.Y.; Yu, S.; Yao, S.; Zhang, G.; Zhang, B.T. Drug Discovery of DKK1 Inhibitors. Front. Pharmacol. 2022, 13, 847387. [Google Scholar] [CrossRef]
- Kawazoe, M.; Kaneko, K.; Nanki, T. Glucocorticoid therapy suppresses Wnt signaling by reducing the ratio of serum Wnt3a to Wnt inhibitors, sFRP-1 and Wif-1. Clin. Rheumatol. 2021, 40, 2947–2954. [Google Scholar] [CrossRef]
- Shen, Z.; Zhu, F.P.; Huang, Y.; Fan, Y.M.; Zhou, F.Q.; Pan, C.X. Research progress of Wnt /Ca2+ signaling pathway in mechanism of limb ischemia/reperfusion injury. Zhongguo Yaolixue Tongbao 2018, 34, 4. [Google Scholar]
- Wang, H.; He, X.Q.; Jin, T.; Li, Y.; Fan, X.Y.; Wang, Y.; Xu, Y.Q. Wnt11 plays an important role in the osteogenesis of human mesenchymal stem cells in a PHA/FN/ALG composite scaffold: Possible treatment for infected bone defect. Stem. Cell. Res. Ther. 2016, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Y.; Xing, L.; Tian, F.M. Roles of noncanonical Wnt signaling in bone homeostasis. Chin. J. Osteoporos. Bone Miner. Res. 2020, 13, 86–94. [Google Scholar]
- Rogers, S.; Scholpp, S. Vertebrate Wnt5a-At the crossroads of cellular signalling. Semin. Cell. Dev. Biol. 2022, 125, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Tian, H.; Zhang, K.; Chen, D.; Chen, D.; Wang, X.; Zhao, J. Wnt5a/FZD4 Mediates the Mechanical Stretch-Induced Osteogenic Differentiation of Bone Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2018, 48, 215–226. [Google Scholar] [CrossRef]
- Maeda, K.; Takahashi, N.; Kobayashi, Y. Roles of Wnt signals in bone resorption during physiological and pathological states. J. Mol. Med. 2013, 91, 15–23. [Google Scholar] [CrossRef]
- Hang, K.; Ye, C.; Xu, J.; Chen, E.; Wang, C.; Zhang, W.; Ni, L.; Kuang, Z.; Ying, L.; Xue, D.; et al. Apelin enhances the osteogenic differentiation of human bone marrow mesenchymal stem cells partly through Wnt/β-catenin signaling pathway. Stem. Cell. Res. Ther. 2019, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, M.; Hong, S.; Kim, E.Y.; Lee, H.; Jung, H.S.; Sohn, Y. Albiflorin Promotes Osteoblast Differentiation and Healing of Rat Femoral Fractures Through Enhancing BMP-2/Smad and Wnt/β-Catenin Signaling. Front. Pharmacol. 2021, 12, 69011. [Google Scholar] [CrossRef]
- Eijken, M.; Meijer, I.M.J.; Westbroek, I.; Koedam, M.; Chiba, H.; Uitterlinden, A.G.; Pols, H.A.P.; Van Leeuwen, J.P.T.M. Wnt signaling acts and is regulated in a human osteoblast differentiation dependent manner. J. Cell. Biochem. 2008, 104, 568–579. [Google Scholar] [CrossRef]
- Liu, D.; Qin, H.; Yang, J.; Yang, L.; He, S.; Chen, S.; Bao, Q.; Zhao, Y.; Zong, Z. Different effects of Wnt/β-catenin activation and PTH activation in adult and aged male mice metaphyseal fracture healing. BMC Musculoskelet. Disord. 2020, 21, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Jin, H.; Zhou, H.; Yu, L.; Wan, H.; He, Y. Guhong Injection promotes fracture healing by activating Wnt/beta-catenin signaling pathway in vivo and in vitro. Biomed. Pharmacother. 2019, 120, 109436. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, D.; He, S.; Yang, L.; Bao, Q.; Qin, H.; Liu, H.; Zhao, Y.; Zong, Z. Differential effects of type 1 diabetes mellitus and subsequent osteoblastic β-catenin activation on trabecular and cortical bone in a mouse model. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M. Research on Wnt/b-catenin in Signaling Acting Synergistically on bmp9-Induced Os-teo/odontoblastic Differentiation of Stem Cells from the Apical Papilla (SCAPs). Ph.D. Thesis, ChongQing Medical University, ChongQing, China, 2015. [Google Scholar]
- Maeda, K.; Kobayashi, Y.; Koide, M.; Uehara, S.; Okamoto, M.; Ishihara, A.; Kayama, T.; Saito, M.; Marumo, K. The Regulation of Bone Metabolism and Disorders by Wnt Signaling. Int. J. Mol. Sci. 2019, 20, 5525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, K.C.; Ding, H.; Tieu, R.; Sparks, N.R.; Ehnes, D.D.; Zur Nieden, N.I. Wnt5a Supports Osteogenic Lineage Decisions in Embryonic Stem Cells. Stem. Cells Dev. 2016, 25, 1020–1032. [Google Scholar] [CrossRef]
- Bullock, W.A.; Hoggatt, A.M.; Horan, D.J.; Elmendorf, A.J.; Sato, A.Y.; Bellido, T.; Loots, G.G.; Pavalko, F.M.; Robling, A.G. Lrp4 Mediates Bone Homeostasis and Mechanotransduction through Interaction with Sclerostin In Vivo. iScience 2019, 20, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Fei, Y.; Hurley, M.M. FGF2 crosstalk with Wnt signaling in mediating the anabolic action of PTH on bone formation. Bone Rep. 2018, 9, 136–144. [Google Scholar] [CrossRef]
- Hsueh, Y.C.; Hodgkinson, C.P.; Gomez, J. The role of Sfrp and DKK proteins in cardiomyocyte development. Physiol. Rep. 2021, 9, e14678. [Google Scholar] [CrossRef]
- Qiang, Y.W.; Barlogie, B.; Rudikoff, S.; Shaughnessy, J.D. Dkk1-induced inhibition of Wnt signaling in osteoblast differentiation is an underlying mechanism of bone loss in multiple myeloma. Bone 2008, 42, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Glantschnig, H.; Hampton, R.; Wei, N. Fully human anti-DKK1 antibodies increase bone formation and resolve osteopenia in mouse models of estrogen-deficiency induced bone loss. J. Bone Miner. Res. 2008, 23, S60–S61. [Google Scholar]
- Sasaki, F.; Hayashi, M.; Ono, T.; Nakashima, T. The regulation of RANKL by mechanical force. J. Bone Miner. Metab. 2021, 39, 34–44. [Google Scholar] [CrossRef]
- Soen, S. Control of inflammatory bone destruction by targeting the Wnt signaling pathway. ]. Clin. Calcium. 2019, 29, 337–341. [Google Scholar]
- Yang, T.; Zhang, J.; Cao, Y.; Zhang, M.; Jing, L.; Jiao, K.; Yu, S.; Chang, W.; Chen, D.; Wang, M. Wnt5a/Ror2 Mediates Temporomandibular Joint Subchondral Bone Remodeling. J. Dent. Res. 2015, 94, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wei, W.; Zeve, D.; Suh, J.M.; Wang, X.; Du, Y.; Zerwekh, J.E.; Dechow, P.C.; Graff, J.M. Biphasic and dosage-dependent regulation of osteoclastogenesis by beta-catenin. Bone 2011, 48, S160. [Google Scholar] [CrossRef]
- Huang, P.; Yan, R.; Zhang, X.; Wang, L.; Ke, X.; Qu, Y. Activating Wnt/β-catenin signaling pathway for disease therapy: Challenges and opportunities. Pharmacol. Ther. 2019, 196, 79–90. [Google Scholar] [CrossRef]
- Heiland, G.R.; Zwerina, K.; Baum, W.; Kireva, T.; Distler, J.H.; Grisanti, M.; Asuncion, F.; Li, X.; Ominsky, M.; Richards, W.; et al. Neutralisation of Dkk-1 protects from systemic bone loss during inflammation and reduces sclerostin expression. Ann. Rheum. Dis. 2010, 69, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Piters, E.; Balemans, W.; Nielsen, T.L.; Andersen, M.; Boudin, E.; Brixen, K.; Van Hul, W. Common Genetic Variation in the DKK1 Gene is Associated with Hip Axis Length but not with Bone Mineral Density and Bone Turnover Markers in Young Adult Men: Results from the Odense Androgen Study. Calcif. Tissue Int. 2010, 86, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhu, X.; He, X.; Yi, X.; Jin, A. The Wnt pathway regulator expression levels and their relationship to bone metabolism in thoracolumbar osteoporotic vertebral compression fracture patients. Am. J. Transl. Res. 2021, 13, 4812–4818. [Google Scholar] [PubMed]
- Glantschnig, H.; Scott, K.; Hampton, R.; Wei, N.; McCracken, P.; Nantermet, P.; Zhao, J.Z.; Vitelli, S.; Huang, L.; Haytko, P.; et al. A rate-limiting role for Dickkopf-1 in bone formation and the remediation of bone loss in mouse and primate models of postmenopausal osteoporosis by an experimental therapeutic antibody. J. Pharmacol. Exp. Ther. 2011, 338, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Voskaridou, E.; Christoulas, D.; Xirakia, C.; Varvagiannis, K.; Boutsikas, G.; Bilalis, A.; Kastritis, E.; Papatheodorou, A.; Terpos, E. Serum Dickkopf-1 is increased and correlates with reduced bone mineral density in patients with thalassemia-induced osteoporosis. Reduction post-zoledronic acid administration. Haematologica 2009, 94, 1182. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Chen, N.; Fu, Z.; Zhang, Q. Progress of Wnt Signaling Pathway in Osteoporosis. Biomolecules 2023, 13, 483. https://doi.org/10.3390/biom13030483
Gao Y, Chen N, Fu Z, Zhang Q. Progress of Wnt Signaling Pathway in Osteoporosis. Biomolecules. 2023; 13(3):483. https://doi.org/10.3390/biom13030483
Chicago/Turabian StyleGao, Yongguang, Na Chen, Zhanda Fu, and Qing Zhang. 2023. "Progress of Wnt Signaling Pathway in Osteoporosis" Biomolecules 13, no. 3: 483. https://doi.org/10.3390/biom13030483