


Full-Length Transcriptomes and Sex-Based Differentially Expressed Genes in the Brain and Ganglia of Giant River Prawn Macrobrachium rosenbergii

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Material
2.2. RNA Extraction and Compete Transcript Sequencing
2.3. Data Processing and Full-Length Transcript Functional Annotation
2.4. Identification of TF, CDS, and lncRNAs
2.5. Differential Expression Analysis via Illumina cDNA Library Sequencing
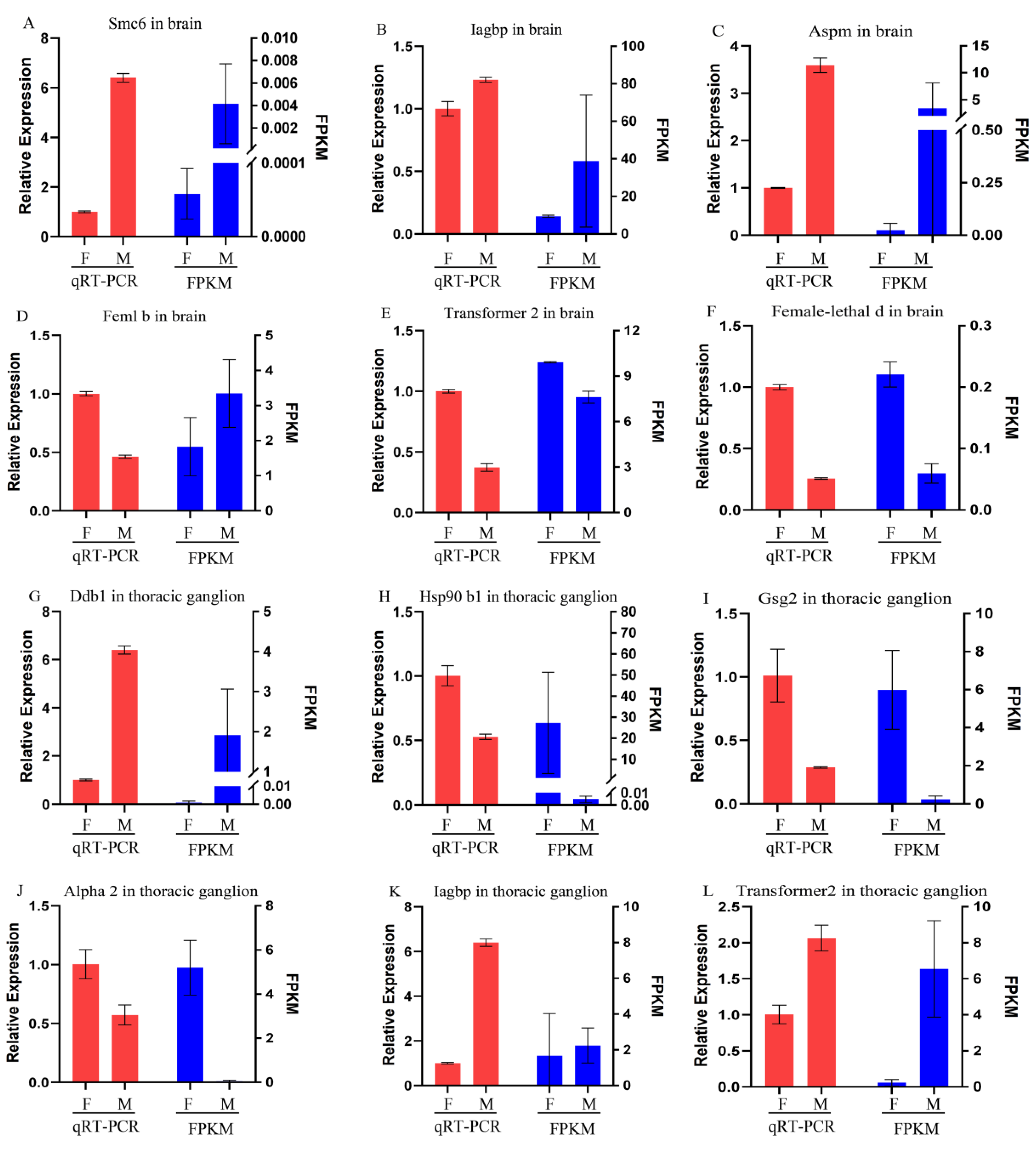
2.6. Differentially Expressed Gene Validation via qRT-PCR
2.7. Construction of LncRNA and Gene Co-Expression Networks
3. Results
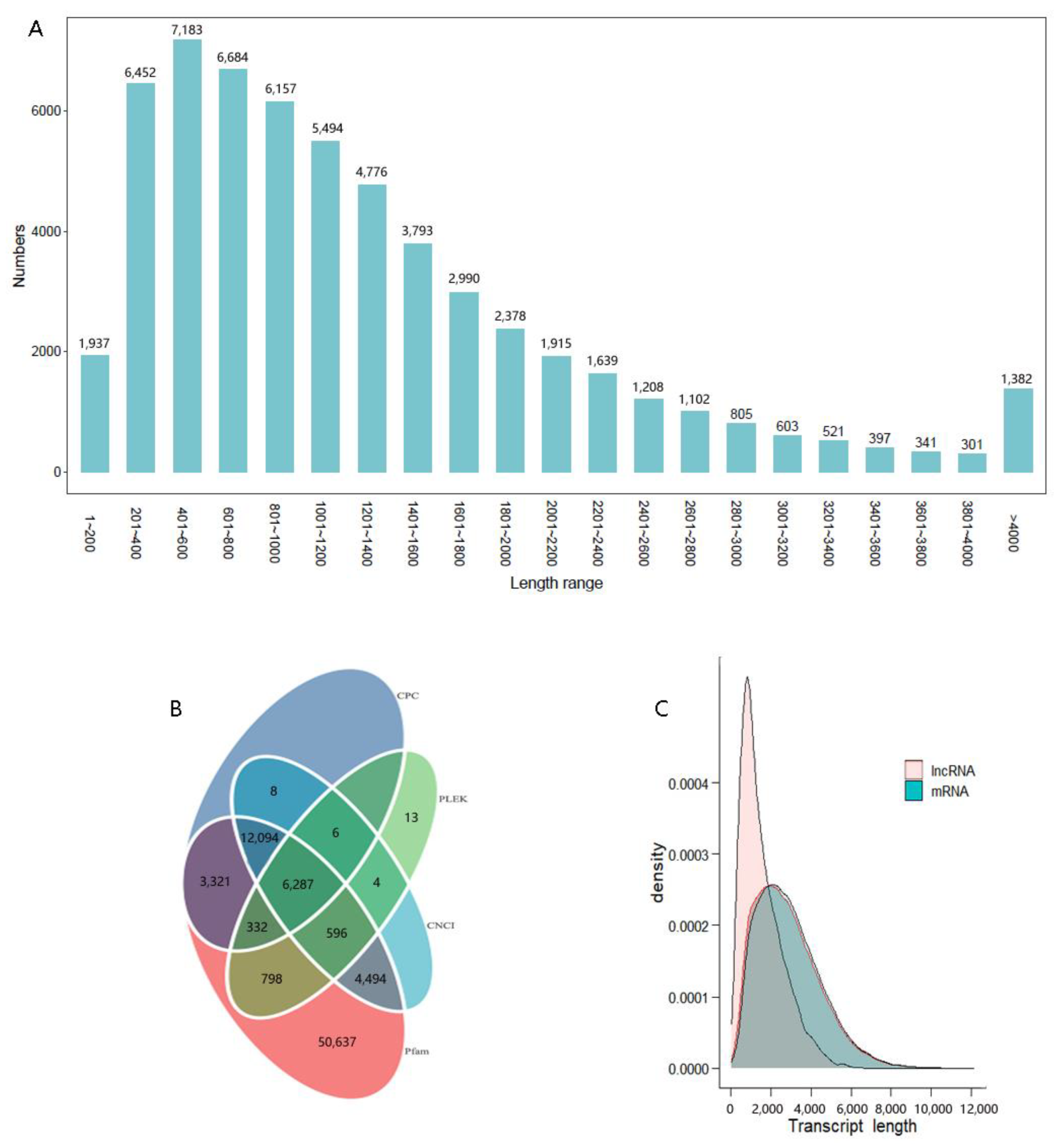
3.1. Full-Length Transcription Sequences
3.2. Assessment of Unigene Completeness
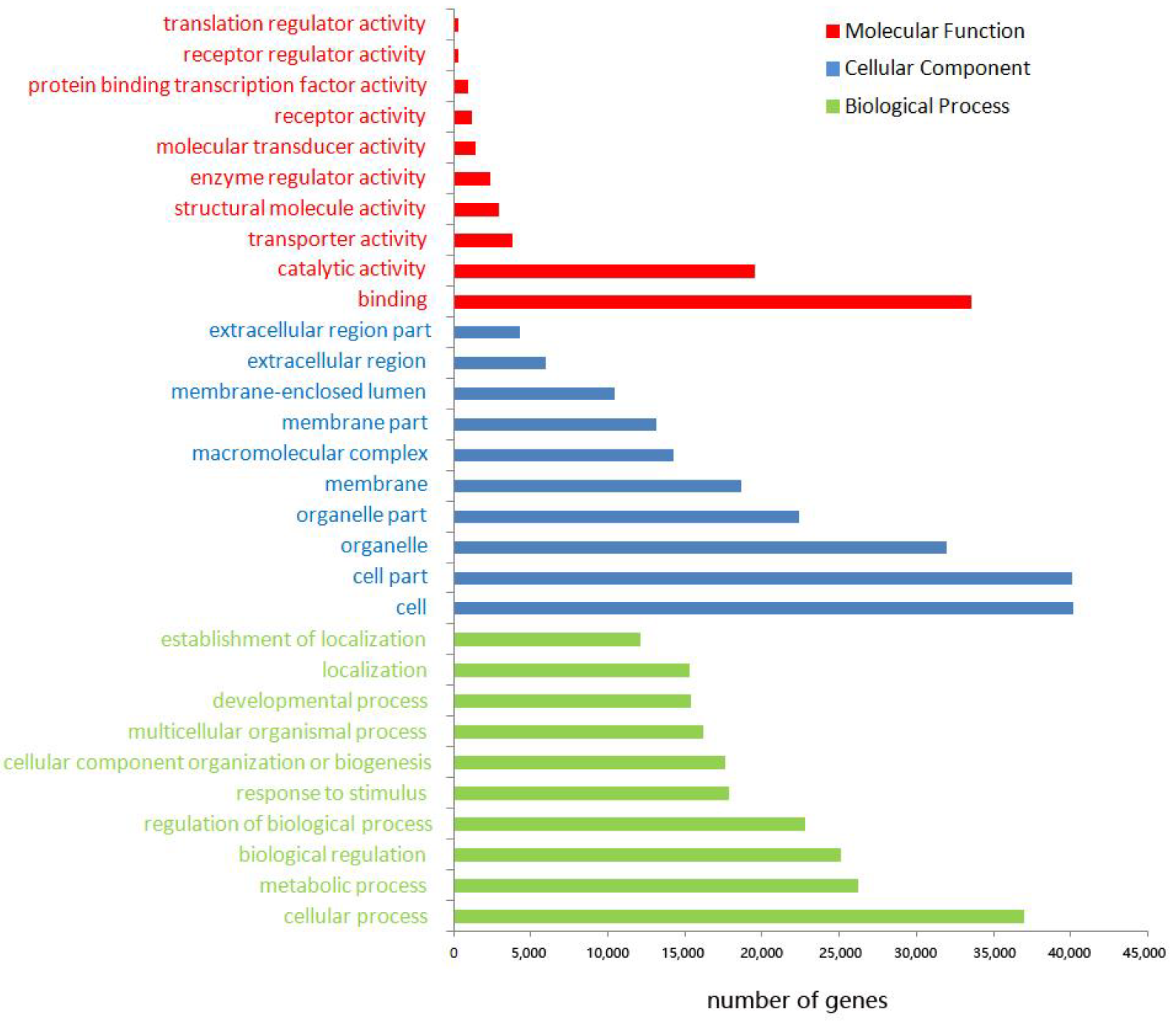
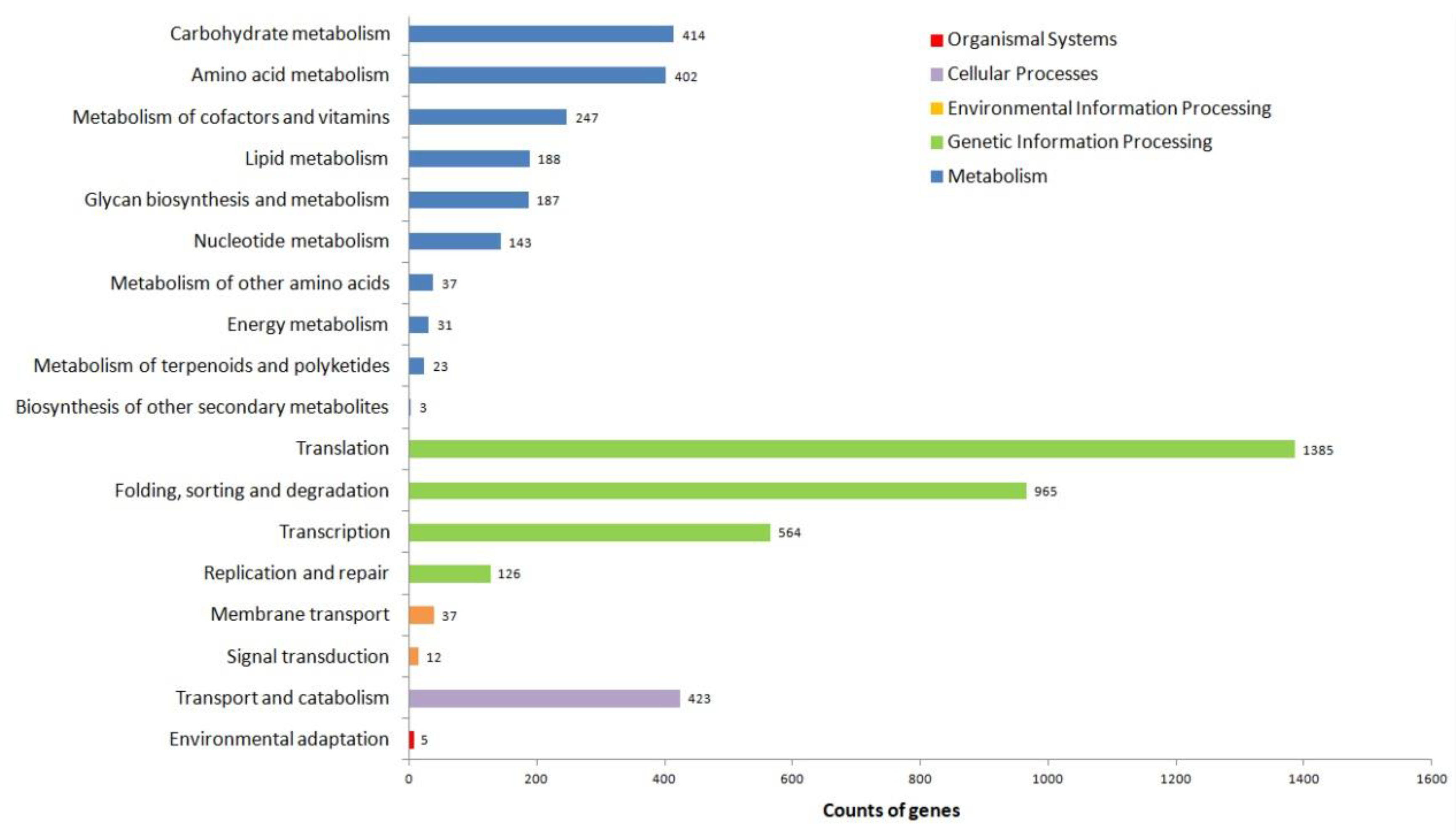
3.3. Functional Annotation of Unigenes
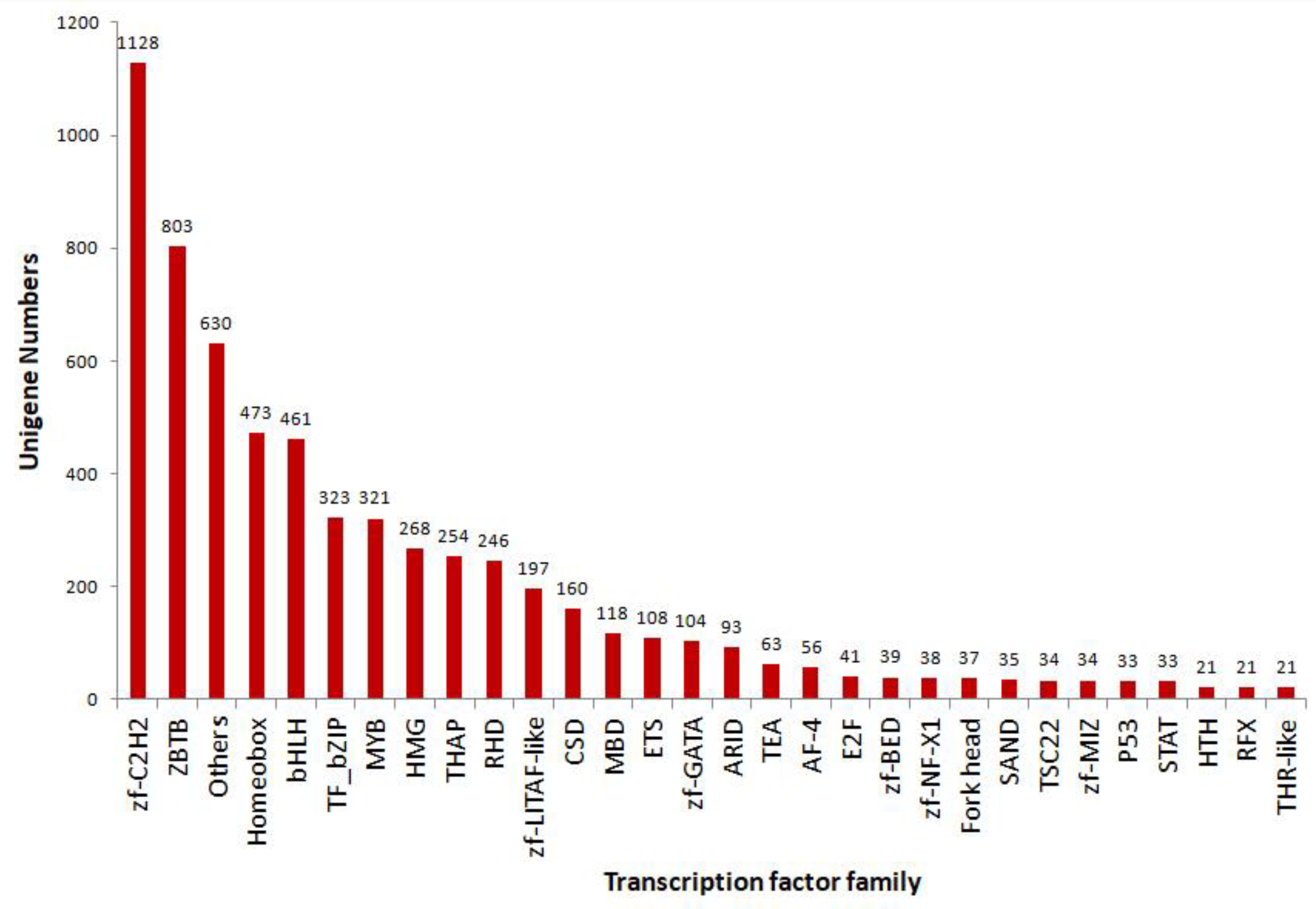
3.4. Results of TF, CDS, and LncRNAs
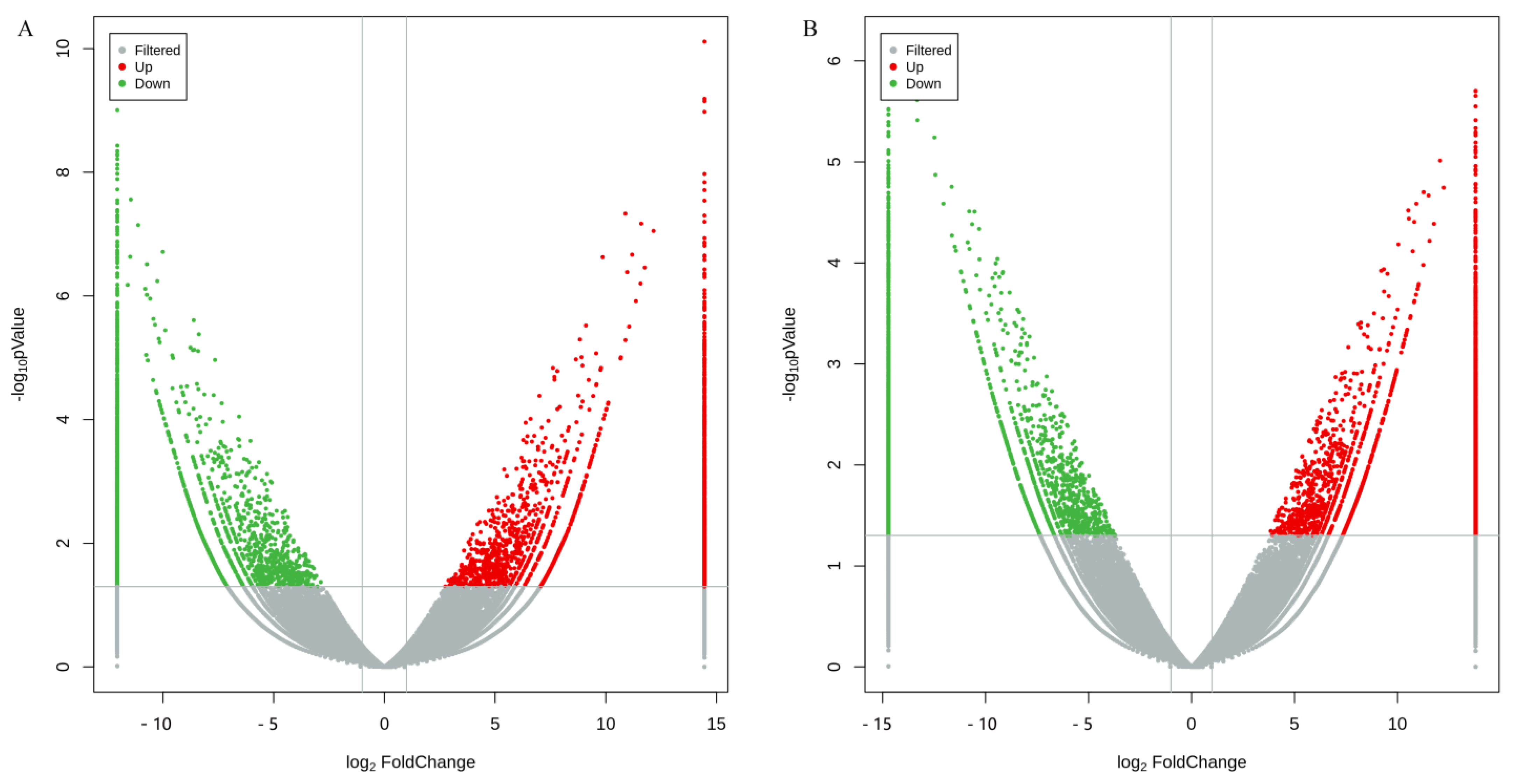
3.5. Differentially Expressed Genes in Brain and Thoracic Ganglion between Sexes Based on Illumina Sequencing Results
3.6. Validation of Differentially Expressed Genes
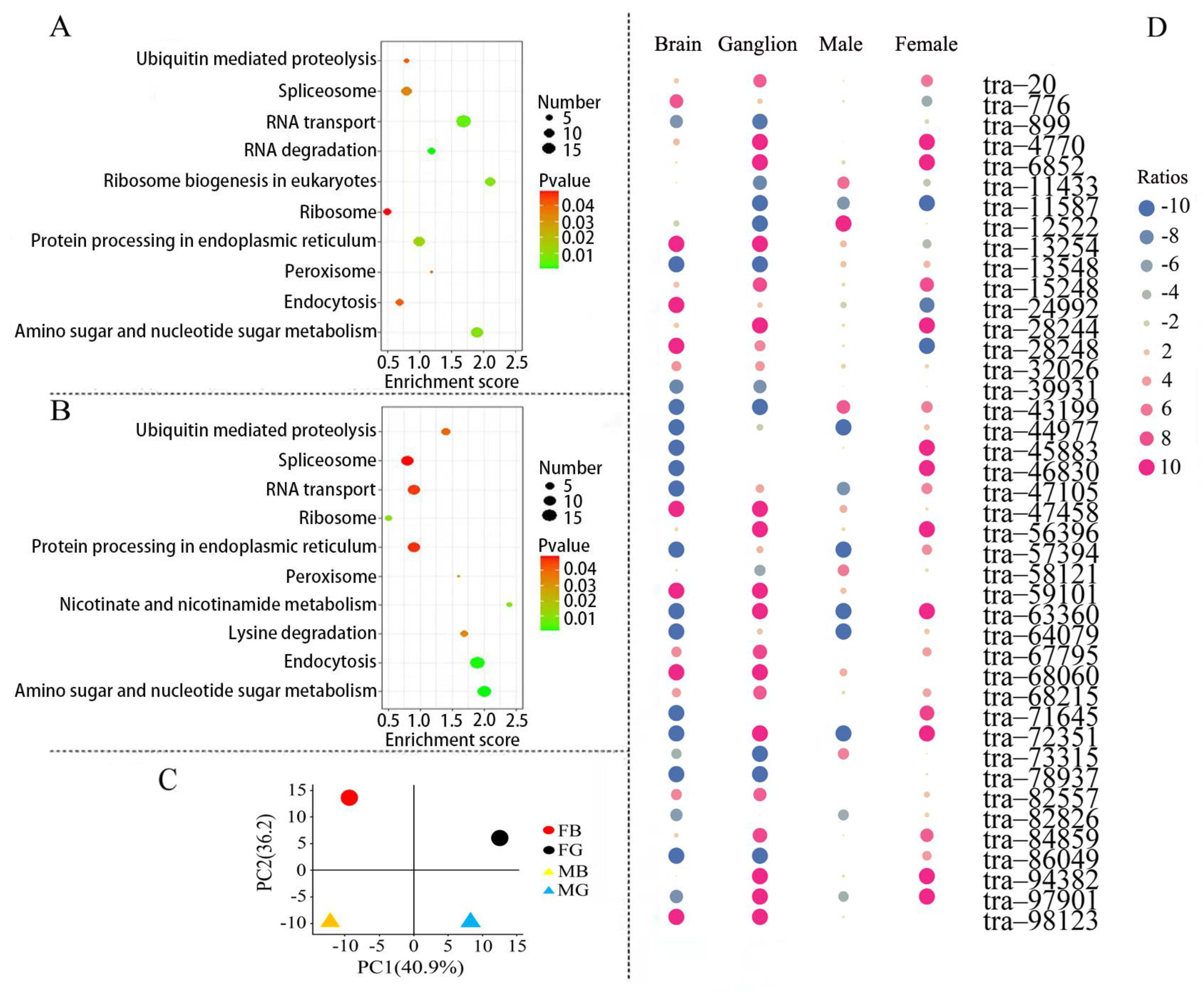
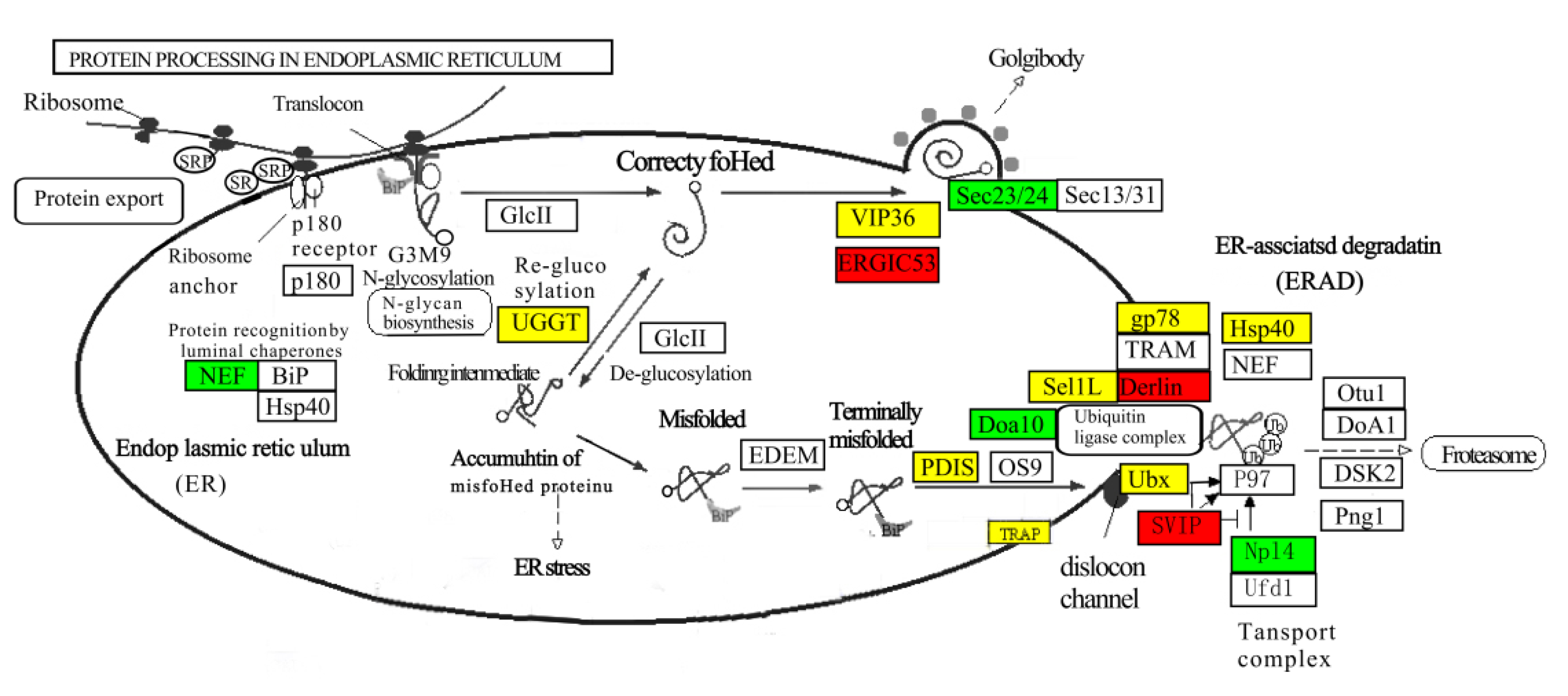
3.7. Gene Expression Analysis in Protein Processing Signaling Pathways
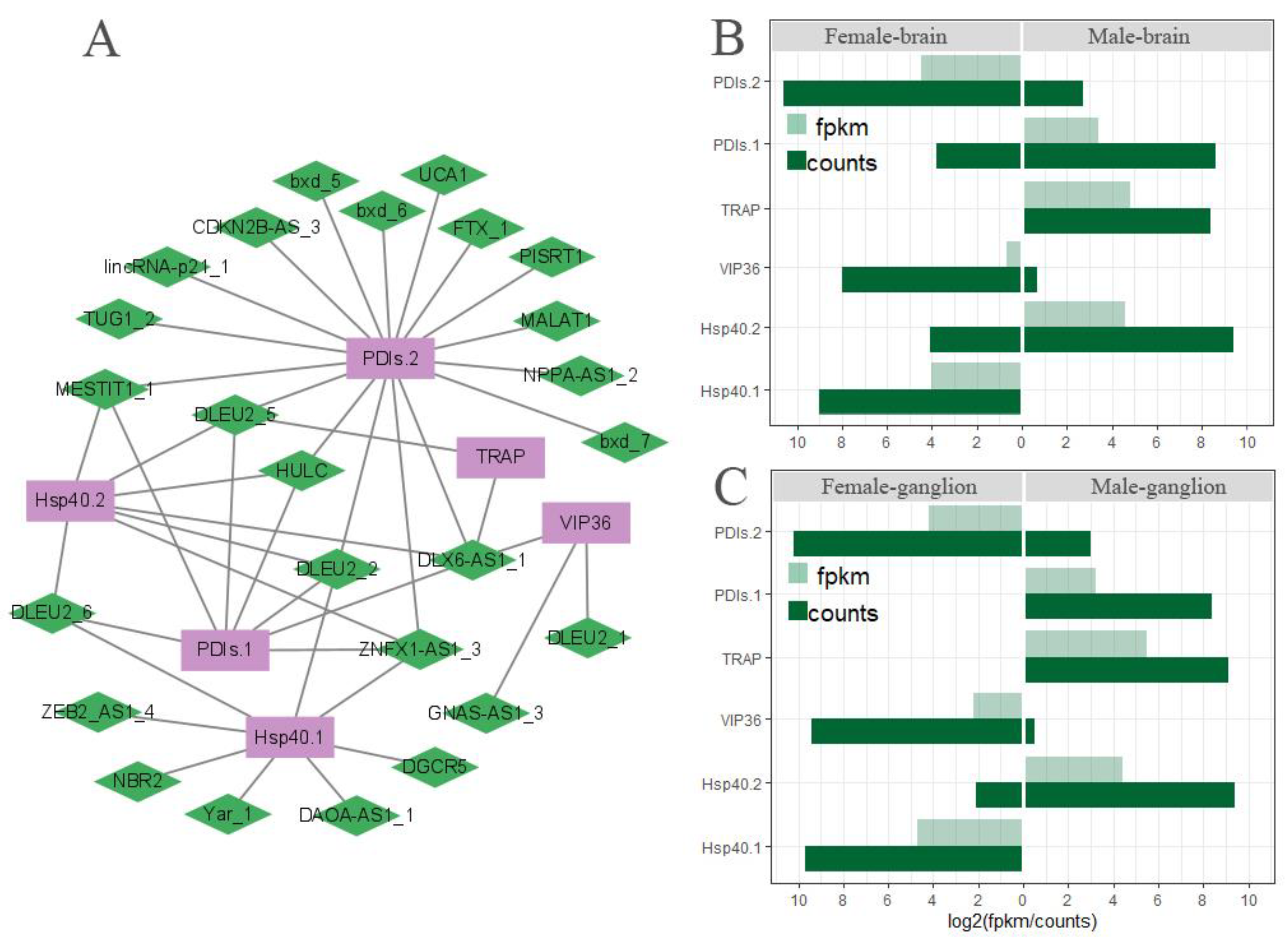
3.8. The Network of Differently Expressed lncRNA Gene Co-Expression in ERAD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.F.; Yang, G.L.; Kong, J.; Luan, S.; Zhang, Y.F.; Wang, J.Y.; Gao, Q.; Luo, K.; Gong, J.H.; Ye, S.Q. Effect of artificial culture and selective breeding on the genetic diversity of Macrobrachium rosenbergii. Acta Hydrobiol. Sin. 2012, 36, 866–873. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Affairs; National Fisheries Technology Extension Center; China Society of Fisheries. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2020. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022, Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Tan, K.; Jiang, H.G.; Jiang, D.; Wang, W. Sex reversal and the androgenic gland (AG) in Macrobrachium rosenbergii: A review. Aquac. Fish. 2020, 5, 283–288. [Google Scholar] [CrossRef]
- Levy, T.; Rosen, O.; Manor, R.; Dotan, S.; Azulay, D.; Abramov, A.; Sklarz, M.Y.; Chalifa-Caspi, V.; Baruch, K.; Shechter, A.; et al. Production of WW males lacking the masculine Z chromosome and mining the Macrobrachium rosenbergii genome for sex-chromosomes. Sci. Rep. 2019, 9, 12408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.L.; Xue, B.; Zhao, L.; Gao, H. Progress on the research of the growth-related genes in crustaceans. Trans. Oceanol. Limnol. 2016, 4, 97–104. [Google Scholar]
- Zhang, X.J.; Yuan, J.B.; Sun, Y.M.; Li, S.H.; Gao, Y.; Yu, Y.; Liu, C.Z.; Wang, Q.C.; Lv, X.J.; Zhang, X.X.; et al. Penaeid shrimp genome provides insights into benthic adaptation and frequent molting. Nat. Commun. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.A.; Andrade, S.C.S.; Teixeira, A.K.; Farias, F.; Guerrelhas, A.C.; Rocha, J.L.; Freitas, P.D. Transcriptome differential expression analysis reveals the activated genes in Litopenaeus vannamei shrimp families of superior growth performance. Aquaculture 2021, 531, 735871. [Google Scholar] [CrossRef]
- Bobkova, N.V.; Poltavtseva, R.A.; Leonov, S.V.; Sukhikh, G.T. Neuroregeneration: Regulation in Neurodegenerative Diseases and Aging. Biochemistry 2020, 85, 108–130. [Google Scholar] [CrossRef]
- Arendt, D.; Tosches, M.A.; Marlow, H. From nerve net to nerve ring, nerve cord and brain—Evolution of the nervous system. Nat. Rev. Neurosci. 2016, 17, 61–72. [Google Scholar] [CrossRef]
- Liao, J.Y. The Morphology and Structure of the Brain of the Prawn, Macrobrachium rosenbergii (Crustacea:Decapoda). Acta Sci. Nat. Univ. Sunyatseni 2001, 40, 85–88. [Google Scholar]
- Liao, J.Y.; Zhang, Y.F.; Sun, J.X.; Lin, Y.A. Partial isolation and bioassay of gonad-stimulating hormone from the brain of Macrobrachium rosenbergii. J. Fish. China 2001, 25, 5–10. [Google Scholar]
- Jayasankar, V.; Tomy, S.; Wilder, M.N. Insights on Molecular Mechanisms of Ovarian Development in Decapod Crustacea: Focus on Vitellogenesis-Stimulating Factors and Pathways. Front. Endocrinol. 2020, 11, 577925. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.S.; Diwan, A.D. Neuroendocrine regulation of ovarian maturation in the Indian white prawn Penaeus indicus H. Milne Edwards. Aquaculture 1991, 98, 381–393. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.K.; Luan, S.; Meng, X.H.; Cao, B.X.; Luo, K.; Kong, J. Comparative transcriptomic characterization of the eyestalk in Pacific white shrimp (Litopenaeus vannamei) during ovarian maturation. Gen. Comp. Endocr. 2019, 274, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Roach, N.P.; Sadowski, N.; Alessi, A.F.; Timp, W.; Taylor, J.; Kim, J.K. The full-length transcriptome of C. elegans using direct RNA sequencing. Genome Res. 2020, 30, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Liu, D.; Zhou, W.Z.; Yi, S.K.; Wang, X.H.; Li, B.L.; Jawad, M.; Xu, H.J.; Gui, L.; Li, M.Y. Comparative transcriptome analysis of brain and gonad reveals reproduction-related miRNAs in the giant prawn, Macrobrachium rosenbergii. Front. Genet. 2022, 13, 990677. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghany, S.E.; Hamilton, M.; Jacobi, J.L.; Ngam, P.; Devitt, N.; Schilkey, F.; Ben-Hur, A.; Reddy, A.S.N. A survey of the sorghum transcriptome using single-molecule long reads. Nat. Commun. 2016, 7, 11706. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.M.; Niu, B.F.; Zhu, Z.W.; Wu, S.T.; Li, W.Z. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopaedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Bairoch, A.; Bougueleret, L.; Altairac, S.; Amendolia, V.; Zhang, J. The Universal Protein Resource (UniProt) 2009. Nucleic Acids Res. 2010, 37, D169–D174. [Google Scholar]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.; Frank, M.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Miao, Y.R.; Jia, L.H.; Yu, Q.Y.; Zhang, Q.; Guo, A.Y. AnimalTFDB 3.0: A comprehensive resource for annotation and prediction of animal transcription factors. Nucleic Acids Res. 2019, 47, D33–D38. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.P.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Luo, H.T.; Bu, D.C.; Zhao, G.G.; Yu, K.T.; Zhang, C.H.; Liu, Y.N.; Chen, R.S.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Li, A.M.; Zhang, J.Y.; Zhou, Z.Y. PLEK: A tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 311. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2013, 10, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012; pp. 1–24. [Google Scholar]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Chen, W.; Li, J.; Huang, S.L.; Xu, X.L.; Zhang, X.; Xiang, S.L.; Liu, C.N. ZFLNC: A comprehensive and well-annotated database for zebrafish lncRNA. Database J. Biol. Databases Curation 2018, 2018, bay114. [Google Scholar] [CrossRef]
- Liu, X.; Ma, K.Y.; Liu, Z.Q.; Feng, J.B.; Ye, B.Q.; Qiu, G.F. Transcriptome analysis of the brain of the Chinese mitten crab, Eriocheir sinensis, for neuropeptide abundance profiles during ovarian development. Anim. Reprod. Sci. 2019, 201, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Qian, T.M.; Qiao, P.P.; Lu, Y.N.; Wang, H.K. Transcription factor SS18L1 regulates the proliferation, migration and differentiation of Schwann cells in peripheral nerve injury. Front. Vet. Sci. 2022, 9, 936620. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Guan, N.; Xu, E.S.; Miao, Y.; Li, C.G. Transcription Factors Leading to High Expression of Neuropeptide L1CAM in Brain Metastases from Lung Adenocarcinoma and Clinical Prognostic Analysis. Dis. Mrk. 2021, 2021, 8585633. [Google Scholar] [CrossRef] [PubMed]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grutzner, F.; Kaessmann, H. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 2014, 505, 635. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.L.; Jiang, S.W.; Sun, D.; Wu, Z.C.; Wei, C.; Dai, C.X.; Jiang, L.H.; Peng, S.H. Transcriptome profiling analysis of sex-based differentially expressed mRNAs and lncRNAs in the brains of mature zebrafish (Danio rerio). BMC Genom. 2019, 20, 830. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.; Rinn, J. Transposable elements reveal a stem cell-specific class of long noncoding RNAs. Genome Biol. 2012, 13, R107. [Google Scholar] [CrossRef] [Green Version]
- Vedula, P.; Kashina, A. The makings of the ‘actin code’: Regulation of actin’s biological function at the amino acid and nucleotide level. J. Cell Sci. 2018, 131, jcs215509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlerson, A.; Radaelli, G.; Mascarello, F.; Veggetti, A. Regeneration of skeletal muscle in two teleost fish: Sparus aurata and Brachydanio rerio. Cell Tissue Res. 1997, 289, 311–322. [Google Scholar] [CrossRef]
- Yang, G.; Lu, Z.J.; Qin, Z.D.; Zhao, L.J.; Pan, G.; Shen, H.Y.; Zhang, M.L.; Liang, R.S.; Lin, L.; Zhang, K. Insight into the Regulatory Relationships between the Insulin-Like Androgenic Gland Hormone Gene and the Insulin-Like Androgenic Gland Hormone-binding Protein Gene in Giant Freshwater Prawns (Macrobrachium rosenbergii). Int. J. Mol. Sci. 2020, 21, 4207. [Google Scholar] [CrossRef]
- Wyganowska-Swiatkowska, M.; Urbaniak, P.; Nohawica, M.M.; Kotwicka, M.; Jankun, J. Enamel matrix proteins exhibit growth factor activity: A review of evidence at the cellular and molecular levels. Exp. Med. 2015, 9, 2025–2033. [Google Scholar] [CrossRef] [Green Version]
- Forouhan, M.; Lim, W.F.; Zanetti-Domingues, L.C.; Tynan, C.J.; Roberts, T.C.; Malik, B.; Manzano, R.; Speciale, A.A.; Ellerington, R.; Garcia-Guerra, A.; et al. AR cooperates with SMAD4 to maintain skeletal muscle homeostasis. Acta Neuropathol. 2022, 143, 713–731. [Google Scholar] [CrossRef] [PubMed]
- Khan, S. Endoplasmic Reticulum in Metaplasticity: From Information Processing to Synaptic Proteostasis. Mol. Neurobiol. 2022, 59, 5630–5655. [Google Scholar] [CrossRef] [PubMed]
- Nufer, O.; Kappeler, F.; Guldbrandsen, S.; Hauri, H.P. ER export of ERGIC-53 is controlled by cooperation of targeting determinants in all three of its domains. J. Cell Sci. 2003, 116, 4429–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yutaka, K.; Noriko, Y.; Yuichi, K.; Takeshi, O. Effect of co-overexpression of the cargo receptor ERGIC-53/MCFD2 on antibody production and intracellular IgG secretion in recombinant Chinese hamster ovary cells. J. Biosci. Bioeng. 2022, 22, 190–196. [Google Scholar] [CrossRef]
- Morro, B.; Broughton, R.; Balseiro, P.; Handeland, S.O.; Mackenzie, S.; Doherty, M.K.; Whitfield, P.D.; Shimizu, M.; Gorissen, M.; Sveier, H.; et al. Endoplasmic reticulum stress as a key mechanism in stunted growth of seawater rainbow trout (Oncorhynchus mykiss). BMC Genom. 2021, 22, 824. [Google Scholar] [CrossRef]
- van de Weijer, M.L.; Bassik, M.C.; Luteijn, R.D.; Voorburg, C.M.; Lohuis, M.A.M.; Kremmer, E.; Hoeben, R.C.; LeProust, E.M.; Chen, S.Y.; Hoelen, H.; et al. A high-coverage shRNA screen identifies TMEM129 as an E3 ligase involved in ER-associated protein degradation. Nat. Commun. 2014, 5, 3832. [Google Scholar] [CrossRef] [Green Version]
- Ballar, P.; Zhong, Y.; Nagahama, M.; Tagaya, M.; Shen, Y.; Fang, S. Identification of SVIP as an endogenous inhibitor of endoplasmic reticulum-associated degradation. J. Biol. Chem. 2007, 282, 33908–33914. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Najafi, S.; Hussen, B.M.; Ganjo, A.R.; Taheri, M.; Samadian, M. DLX6-AS1: A Long Non-coding RNA With Oncogenic Features. Front. Cell Dev. Biol. 2022, 10, 746443. [Google Scholar] [CrossRef]
- Xian, H.P.; Zhuo, Z.L.; Sun, Y.J.; Liang, B.; Zhao, X.T. Circulating long non-coding RNAs HULC and ZNFX1-AS1 are potential biomarkers in patients with gastric cancer. Oncol. Lett. 2018, 16, 4689–4698. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, J. LncRNAs: Macromolecules with big roles in neurobiology and neurological diseases. Metab. Brain Dis. 2017, 32, 281–291. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unigene ID | Gene Name | Primer Sequence (5- > 3) | |
|---|---|---|---|
| Forward | Reverse | ||
| transcript_40005 | SMCP6 | TGCAGTGGTTGTTGCACTTG | ACTGCACAGTAGCTGTTTGC |
| transcript_88064 | IAGBP | AACATGGCTGACGGTTCTTC | TCCGGACGTTGATGTTCATG |
| transcript_16449 | ASMP | ACAGATCAGTGCCACACATG | ACGGGAACAAACCATCAAGC |
| transcript_29898 | Fem1b | AATTGCATGTGGGGCTGATG | TGTCCATGTGTGCTCCATTC |
| transcript_72021 | transformer 2 | ACATCATGGAAGCAGAGAGGAC | TGGCATCCAGAACAACTTGC |
| transcript_45783 | female lethal d-like protein | GGTGAATTGGCCCTTCAAAGG | TTCGCCTGCCTTAATTGCTG |
| transcript_47685 | DNA damage- binding protein 1 | TTCGAGTGGACGAACGAAAG | ACGTCTTGTACTGCAGCAAG |
| transcript_10906 | alpha-2-macroglobulin | TCAGTGAAGCAGCCTTTTGC | TGATGTCTCTGTGGCCAAAC |
| transcript_57773 | heat shock protein 90 | TCCGCAAGAACTTGGTCAAG | AGCCAACTTCTTGCGGTTAG |
| transcript_524 | ganglioside GM2 activator | TATCGGTTTCATGTGGGTGGAG | ATGGATCAGGGCAAGGTTCG |
| transcript_60749 | Actin | AATCGTGCGTGACATCAAGG | TCTCGTTACCGATGGTGATGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Hong, Z.; Gui, L.; Zhao, L.; Wang, Y.; Sun, S.; Li, M. Full-Length Transcriptomes and Sex-Based Differentially Expressed Genes in the Brain and Ganglia of Giant River Prawn Macrobrachium rosenbergii. Biomolecules 2023, 13, 460. https://doi.org/10.3390/biom13030460
Liu D, Hong Z, Gui L, Zhao L, Wang Y, Sun S, Li M. Full-Length Transcriptomes and Sex-Based Differentially Expressed Genes in the Brain and Ganglia of Giant River Prawn Macrobrachium rosenbergii. Biomolecules. 2023; 13(3):460. https://doi.org/10.3390/biom13030460
Chicago/Turabian StyleLiu, Dong, Zhenzhen Hong, Lang Gui, Li Zhao, Yude Wang, Shengming Sun, and Mingyou Li. 2023. "Full-Length Transcriptomes and Sex-Based Differentially Expressed Genes in the Brain and Ganglia of Giant River Prawn Macrobrachium rosenbergii" Biomolecules 13, no. 3: 460. https://doi.org/10.3390/biom13030460