
Upregulation of Apolipoprotein L6 Improves Tumor Immunotherapy by Inducing Immunogenic Cell Death

Abstract
:1. Introduction
2. Materials and Methods
2.1. Transcriptomic Profile of EBV-Associated Tumors and Tumor Immunotherapy Cohorts
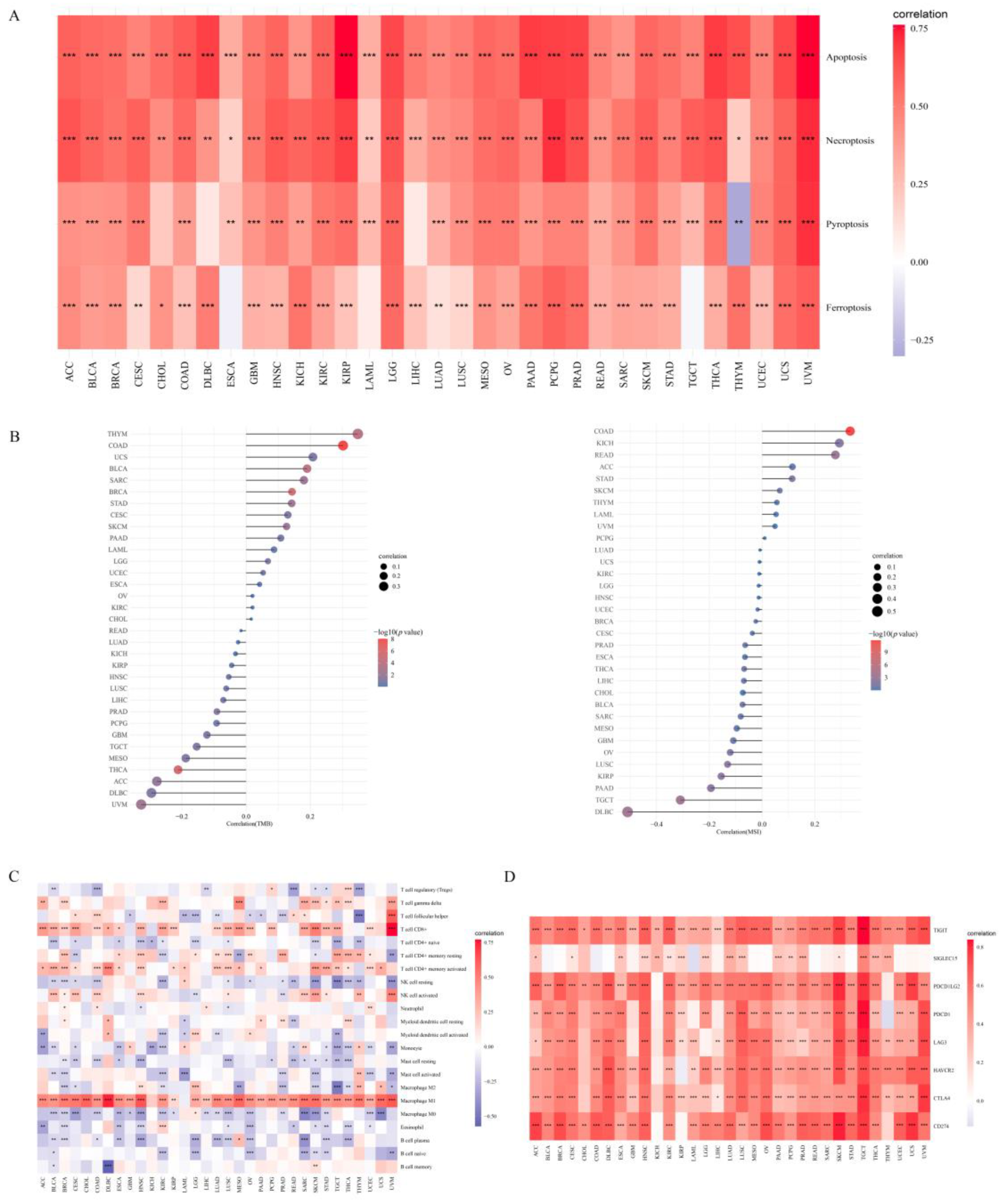
2.2. Association between APOL6 and the Tumor Microenvironment
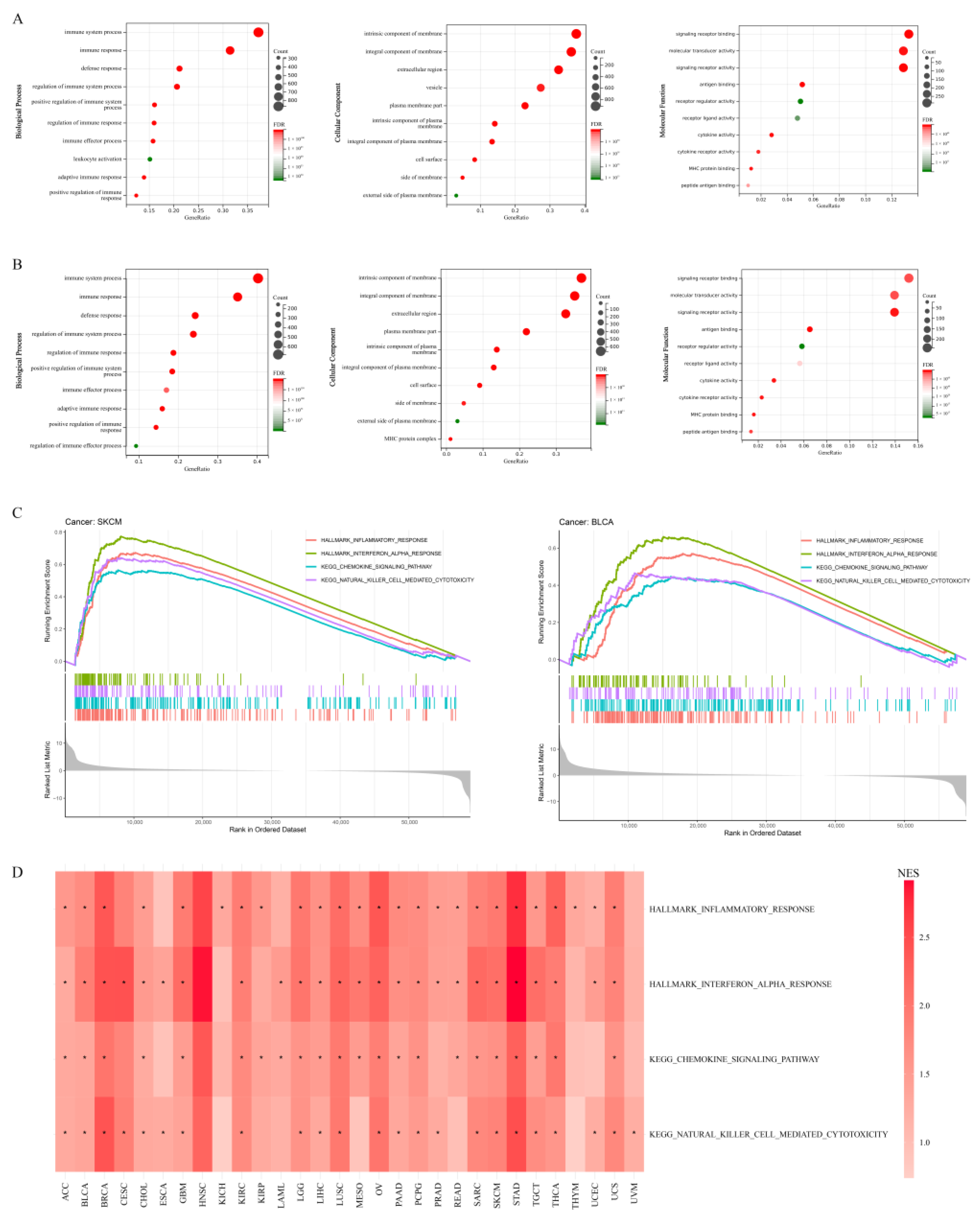
2.3. Gene Ontology (GO) and Gene Set Enrichment Analysis (GSEA)
2.4. Cell Counting Kit-8 (CCK8) Assay
2.5. Flow Cytometry
2.6. Western Blot Analysis
2.7. RNA Extraction, cDNA Synthesis, and qRT-PCR
2.8. Statistical Analysis
3. Results
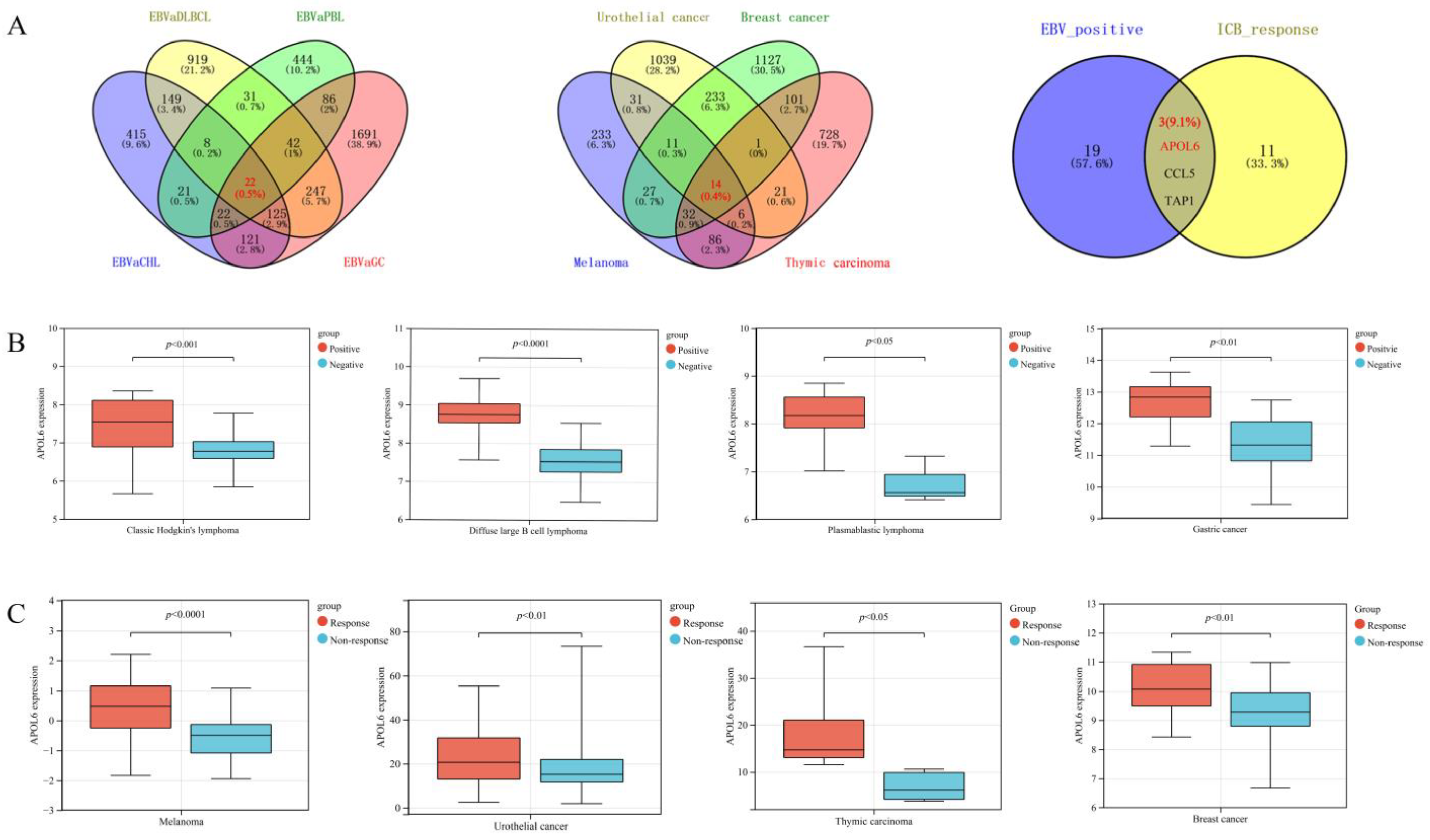
3.1. Exploring the Critical Genes Enhancing Antitumor Immunity across EBV-Positive Tumors
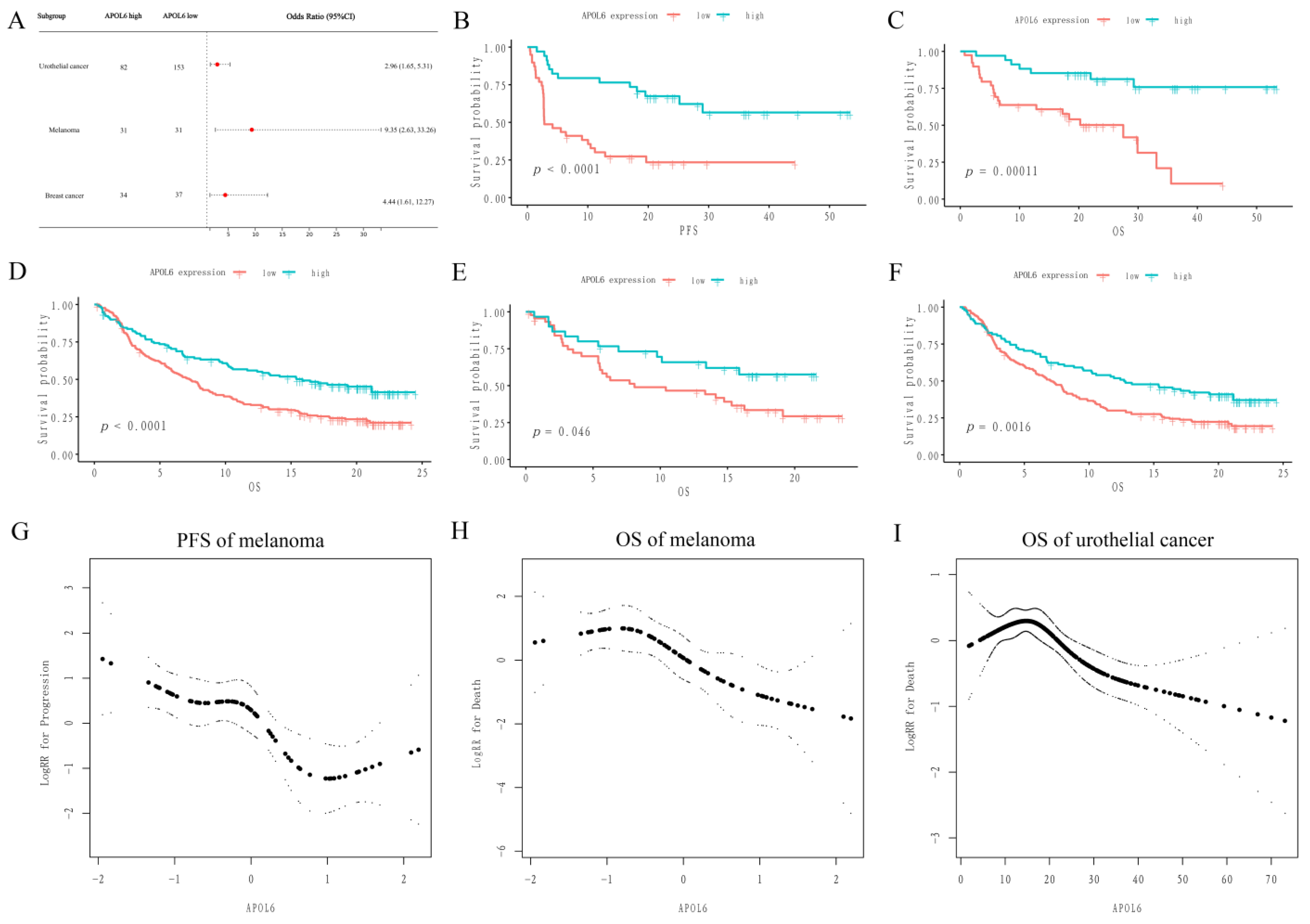
3.2. Upregulation of APOL6 Correlated with Better Immunotherapy Response and Prognosis
3.3. Role of APOL6 in Improving Cancer Immunotherapy Based on Bioinformatics Analyses
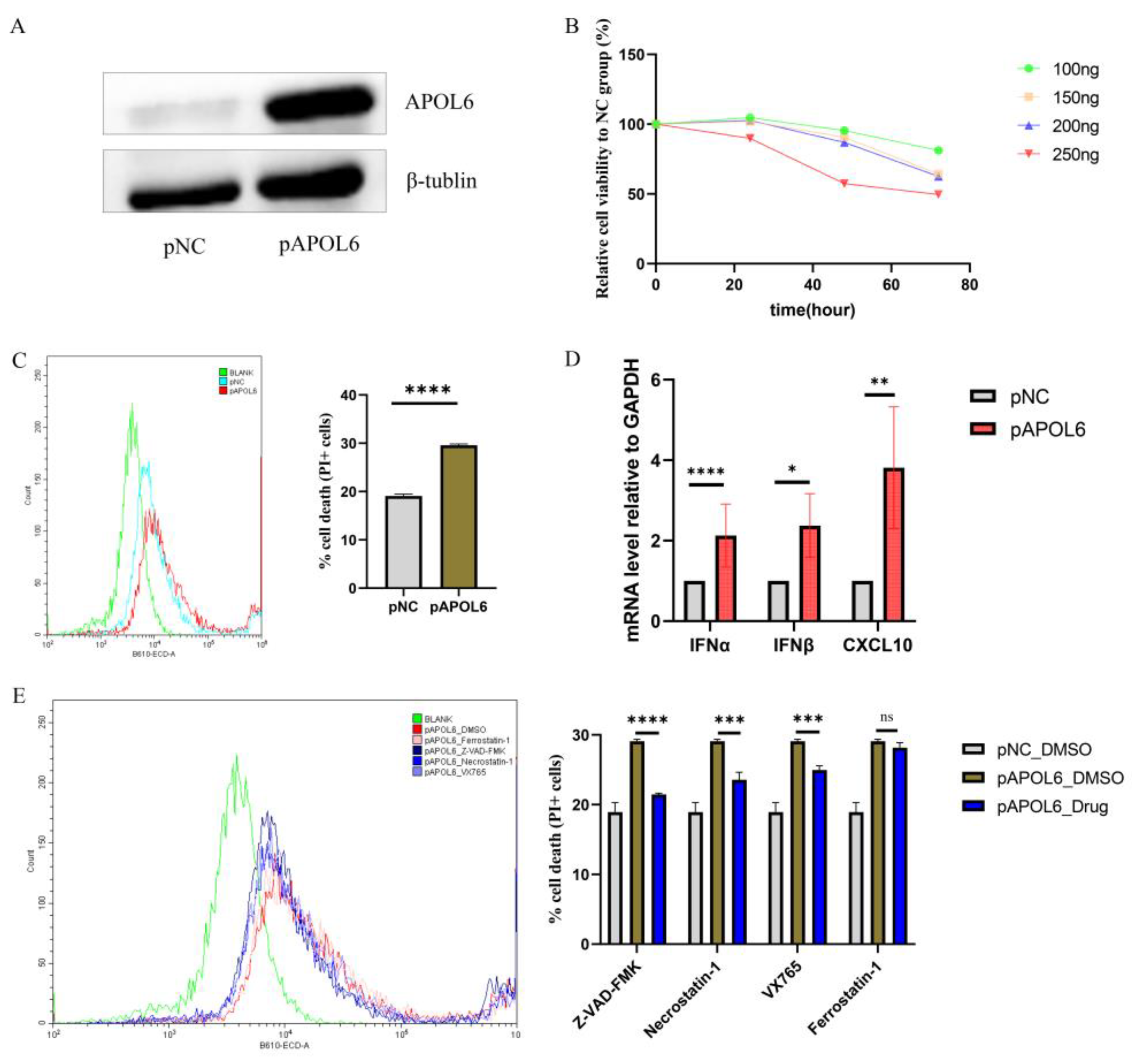
3.4. APOL6 Induced Immunogenic Tumor Cell Death In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Sanmamed, M.F.; Chen, L. A Paradigm Shift in Cancer Immunotherapy: From Enhancement to Normalization. Cell 2018, 175, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Litchfield, K.; Reading, J.L.; Puttick, C.; Thakkar, K.; Abbosh, C.; Bentham, R.; Watkins, T.B.K.; Rosenthal, R.; Biswas, D.; Rowan, A.; et al. Meta-analysis of tumor- and T cell-intrinsic mechanisms of sensitization to checkpoint inhibition. Cell 2021, 184, 596–614.e14. [Google Scholar] [CrossRef]
- Kim, S.T.; Cristescu, R.; Bass, A.J.; Kim, K.M.; Odegaard, J.I.; Kim, K.; Liu, X.Q.; Sher, X.; Jung, H.; Lee, M.; et al. Comprehensive molecular characterization of clinical responses to PD-1 inhibition in metastatic gastric cancer. Nat. Med. 2018, 24, 1449–1458. [Google Scholar] [CrossRef]
- Wei, X.L.; Liu, Q.W.; Liu, F.R.; Yuan, S.S.; Li, X.F.; Li, J.N.; Yang, A.L.; Ling, Y.H. The clinicopathological significance and predictive value for immunotherapy of programmed death ligand-1 expression in Epstein-Barr virus-associated gastric cancer. Oncoimmunology 2021, 10, 1938381. [Google Scholar] [CrossRef]
- Gravelle, P.; Péricart, S.; Tosolini, M.; Fabiani, B.; Coppo, P.; Amara, N.; Traverse-Gléhen, A.; Van Acker, N.; Brousset, P.; Fournie, J.-J.; et al. EBV infection determines the immune hallmarks of plasmablastic lymphoma. Oncoimmunology 2018, 7, e1486950. [Google Scholar] [CrossRef] [Green Version]
- Cristino, A.S.; Nourse, J.; West, R.A.; Sabdia, M.B.; Law, S.C.; Gunawardana, J.; Vari, F.; Mujaj, S.; Thillaiyampalam, G.; Snell, C.; et al. EBV microRNA-BHRF1-2-5p targets the 3′UTR of immune checkpoint ligands PD-L1 and PD-L2. Blood 2019, 134, 2261–2270. [Google Scholar] [CrossRef]
- Chetaille, B.; Bertucci, F.; Finetti, P.; Esterni, B.; Stamatoullas, A.; Picquenot, J.M.; Copin, M.C.; Morschhauser, F.; Casasnovas, O.; Petrella, T.; et al. Molecular profiling of classical Hodgkin lymphoma tissues uncovers variations in the tumor microenvi-ronment and correlations with EBV infection and outcome. Blood 2009, 113, 2765–3775. [Google Scholar] [CrossRef] [Green Version]
- Kamper, P.; Bendix, K.; Hamilton-Dutoit, S.; Honoré, B.; Nyengaard, J.R.; d’Amore, F. Tumor-infiltrating macrophages cor-relate with adverse prognosis and Epstein-Barr virus status in classical Hodgkin’s lymphoma. Haematologica 2011, 96, 269–276. [Google Scholar] [CrossRef]
- Morscio, J.; Dierickx, D.; Ferreiro, J.F.; Herreman, A.; Van Loo, P.; Bittoun, E.; Verhoef, G.; Matthys, P.; Cools, J.; Wlodarska, I.; et al. Gene expression profiling reveals clear differences between EBV-positive and EBV-negative posttransplant lymphoproliferative disorders. Am. J. Transplant. 2013, 13, 1305–1316. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, C.; Kim, H.J.; Park, J.; Hwang, J.; Kim, J.I.; Choi, M.G.; Kim, S.; Kim, K.M.; Kang, M.S. Deregulation of immune response genes in patients with Epstein-Barr virus-associated gastric cancer and outcomes. Gastroenterology 2015, 148, 137–147.e9. [Google Scholar] [CrossRef]
- Pusztai, L.; Yau, C.; Wolf, D.M.; Han, H.S.; Du, L.; Wallace, A.M.; String-Reasor, E.; Boughey, J.C.; Chien, A.J.; Elias, A.D.; et al. Durvalumab with olaparib and paclitaxel for high-risk HER2-negative stage II/III breast cancer: Results from the adaptively randomized I-SPY2 trial. Cancer Cell 2021, 39, 989–998.e5. [Google Scholar] [CrossRef]
- He, Y.; Ramesh, A.; Gusev, Y.; Bhuvaneshwar, K.; Giaccone, G. Molecular predictors of response to pembrolizumab in thymic carcinoma. Cell Rep. Med. 2021, 2, 100392. [Google Scholar] [CrossRef]
- Mariathasan, S.; Turley, S.J.; Nickles, D.; Castiglioni, A.; Yuen, K.; Wang, Y.; Kadel, E.E.I.; Koeppen, H.; Astarita, J.L.; Cubas, R.; et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 2018, 554, 544–548. [Google Scholar] [CrossRef]
- Gide, T.N.; Quek, C.; Menzies, A.M.; Tasker, A.T.; Shang, P.; Holst, J.; Madore, J.; Lim, S.Y.; Velickovic, R.; Wongchenko, M.; et al. Distinct Immune Cell Populations Define Response to Anti-PD-1 Monotherapy and Anti-PD-1/Anti-CTLA-4 Combined Therapy. Cancer Cell 2019, 35, 238–255.e6. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Gu, S.; Pan, D.; Fu, J.; Sahu, A.; Hu, X.; Li, Z.; Traugh, N.; Bu, X.; Li, B.; et al. Signatures of T cell dysfunction and exclusion predict cancer immunotherapy response. Nat. Med. 2018, 24, 1550–1558. [Google Scholar] [CrossRef]
- Zhao, S.; Ye, Z.; Stanton, R. Misuse of RPKM or TPM normalization when comparing across samples and sequencing protocols. RNA 2020, 26, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2018, 48, 812–830.e14. [Google Scholar] [CrossRef] [Green Version]
- Bonneville, R.; Krook, M.A.; Kautto, E.A.; Miya, J.; Wing, M.R.; Chen, H.Z.; Reeser, J.W.; Yu, L.; Roychowdhury, S. Landscape of Microsatellite Instability Across 39 Cancer Types. JCO Precis. Oncol. 2017, 1, 1–15. [Google Scholar] [CrossRef]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, E.; Karasawa, T.; Watanabe, S.; Komada, T.; Kimura, H.; Kamata, R.; Ito, H.; Hishida, E.; Yamada, N.; Kasahara, T.; et al. GSDME-Dependent Incomplete Pyroptosis Permits Selective IL-1α Release under Caspase-1 Inhibition. iScience 2020, 23, 101070. [Google Scholar] [CrossRef]
- Liu, S.; Wu, W.; Chen, Q.; Zheng, Z.; Jiang, X.; Xue, Y.; Lin, D. TXNRD1: A Key Regulator Involved in the Ferroptosis of CML Cells Induced by Cysteine Depletion In Vitro. Oxidative Med. Cell. Longev. 2021, 2021, 7674565. [Google Scholar] [CrossRef]
- Perperoglou, A.; Sauerbrei, W.; Abrahamowicz, M.; Schmid, M. A review of spline function procedures in R. BMC Med. Res. Methodol. 2019, 19, 46. [Google Scholar] [CrossRef] [Green Version]
- Garzotto, M.; Beer, T.M.; Hudson, R.G.; Peters, L.; Hsieh, Y.C.; Barrera, E.; Klein, T.; Mori, M. Improved detection of prostate cancer using classification and regression tree analysis. J. Clin. Oncol. 2005, 23, 4322–4329. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Bao, X.; Zhang, R.; Ding, Y.; Zhang, M.; Li, B.; Zhang, H.; Li, X.; Tong, Z.; Liu, L.; et al. Depiction of the genomic and genetic landscape identifies CCL5 as a protective factor in colorectal neuroendocrine carcinoma. Br. J. Cancer 2021, 125, 994–1002. [Google Scholar] [CrossRef]
- Nesbeth, Y.C.; Martinez, D.G.; Toraya, S.; Scarlett, U.K.; Cubillos-Ruiz, J.R.; Rutkowski, M.R.; Conejo-Garcia, J.R. CD4+ T cells elicit host immune responses to MHC class II-negative ovarian cancer through CCL5 secretion and CD40-mediated licensing of dendritic cells. J. Immunol. 2010, 184, 5654–5662. [Google Scholar] [CrossRef] [Green Version]
- Doorduijn, E.M.; Sluijter, M.; Querido, B.J.; Oliveira, C.C.; Achour, A.; Ossendorp, F.; van der Burg, S.H.; van Hall, T. TAP-independent self-peptides enhance T cell recognition of immune-escaped tumors. J. Clin. Investig. 2016, 126, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Chen, L.; Li, Y.; Hu, Z.; He, F. Ferroptosis, necroptosis, and pyroptosis in the tumor microenvironment: Perspectives for immunotherapy of SCLC. Semin. Cancer Biol. 2022, 86, 273–285. [Google Scholar] [CrossRef]
- Tang, R.; Xu, J.; Zhang, B.; Liu, J.; Liang, C.; Hua, J.; Meng, Q.; Yu, X.; Shi, S. Ferroptosis, necroptosis, and pyroptosis in anticancer immunity. J. Hematol. Oncol. 2020, 13, 110. [Google Scholar] [CrossRef]
- Sistigu, A.; Yamazaki, T.; Vacchelli, E.; Chaba, K.; Enot, D.P.; Adam, J.; Vitale, I.; Goubar, A.; Baracco, E.E.; Remédios, C.; et al. Cancer cell–autonomous contribution of type I interferon signaling to the efficacy of chemotherapy. Nat. Med. 2014, 20, 1301–1309. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Warren, S.; Adjemian, S.; Agostinis, P.; Martinez, A.B.; Chan, T.A.; Coukos, G.; Demaria, S.; Deutsch, E.; et al. Consensus guidelines for the definition, detection and interpretation of immunogenic cell death. J. Immunother. Cancer 2020, 8, e000337. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Lu, H.; Jiang, Z.; Pastuszyn, A.; Hu, C.-A.A. Apolipoprotein L6, a Novel Proapoptotic Bcl-2 Homology 3–Only Protein, Induces Mitochondria-Mediated Apoptosis in Cancer Cells. Mol. Cancer Res. 2005, 3, 21–31. [Google Scholar] [CrossRef]
- Fujikura, D.; Miyazaki, T. Programmed Cell Death in the Pathogenesis of Influenza. Int. J. Mol. Sci. 2018, 19, 2065. [Google Scholar] [CrossRef] [Green Version]
- Sauter, B.; Albert, M.L.; Francisco, L.; Larsson, M.; Somersan, S.; Bhardwaj, N. Consequences of cell death: Exposure to necrotic tumor cells, but not primary tissue cells or apoptotic cells, induces the maturation of immunostimulatory dendritic cells. J. Exp. Med. 2000, 191, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Christofi, T.; Baritaki, S.; Falzone, L.; Libra, M.; Zaravinos, A. Current Perspectives in Cancer Immunotherapy. Cancers 2019, 11, 1472. [Google Scholar] [CrossRef] [Green Version]
- Brahmer, J.R.; Tykodi, S.S.; Chow, L.Q.; Hwu, W.J.; Topalian, S.L.; Hwu, P.; Drake, C.G.; Camacho, L.H.; Kauh, J.; Odunsi, K.; et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N. Engl. J. Med. 2012, 366, 2455–2465. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.S.; Badgwell, B.D. Current treatment and recent progress in gastric cancer. CA A Cancer J. Clin. 2021, 71, 264–279. [Google Scholar] [CrossRef]
- Kim, H.; Heo, Y.J.; Cho, Y.A.; Kang, S.Y.; Ahn, S.; Kim, K.M. Tumor immune microenvironment is influenced by frameshift mutations and tumor mutational burden in gastric cancer. Clin. Transl. Oncol. 2021, 24, 556–567. [Google Scholar] [CrossRef]
- Tan, Y.; Gan, M.; Fan, Y.; Li, L.; Zhong, Z.; Li, X.; Bai, L.; Zhao, Y.; Niu, L.; Shang, Y.; et al. miR-10b-5p regulates 3T3-L1 cells differentiation by targeting Apol6. Gene 2019, 687, 39–46. [Google Scholar] [CrossRef]
- Bedoui, S.; Herold, M.J.; Strasser, A. Emerging connectivity of programmed cell death pathways and its physiological impli-cations. Nat. Rev. Mol. Cell Biol. 2020, 21, 678–695. [Google Scholar] [CrossRef]
- Gratton, R.; Tricarico, P.M.; Agrelli, A.; Colaço da Silva, H.V.; Coêlho Bernardo, L.; Crovella, S.; Campos Coelho, A.V.; Ro-drigues de Moura, R.; Cavalcanti Brandão, L.A. In Vitro Zika Virus Infection of Human Neural Progenitor Cells: Meta-Analysis of RNA-Seq Assays. Microorganisms 2020, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- McLaren, P.J.; Gawanbacht, A.; Pyndiah, N.; Krapp, C.; Hotter, D.; Kluge, S.F.; Götz, N.; Heilmann, J.; Mack, K.; Sauter, D.; et al. Identification of potential HIV restriction factors by combining evolutionary genomic signatures with functional analyses. Retrovirology 2015, 12, 41. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Innate immune recognition of viral infection. Nat. Immunol. 2006, 7, 131–137. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Chiang, N.J.; Hou, Y.C.; Tan, K.T.; Tsai, H.W.; Lin, Y.J.; Yeh, Y.C.; Chen, L.T.; Hou, Y.F.; Chen, M.H.; Shan, Y.S. The immune microenvironment features and response to immunotherapy in EBV-associated lymphoepithelioma-like cholangiocarcinoma. Hepatol. Int. 2022, 16, 1137–1149. [Google Scholar] [CrossRef]
- Granai, M.; Lazzi, S.; Mancini, V.; Akarca, A.; Santi, R.; Vergoni, F.; Sorrentino, E.; Guazzo, R.; Mundo, L.; Cevenini, G.; et al. Burkitt lymphoma with a granulomatous reaction: An M1/Th1-polarised microenvironment is associated with controlled growth and spontaneous regression. Histopathology 2022, 80, 430–442. [Google Scholar] [CrossRef]
- Barros, M.H.M.; Segges, P.; Vera-Lozada, G.; Hassan, R.; Niedobitek, G. Macrophage Polarization Reflects T Cell Composition of Tumor Microenvironment in Pediatric Classical Hodgkin Lymphoma and Has Impact on Survival. PLoS ONE 2015, 10, e0124531. [Google Scholar] [CrossRef]
- Tian, J.-J.; Lei, C.-X.; Ji, H.; Zhou, J.-S.; Yu, H.-B.; Li, Y.; Yu, E.-M.; Xie, J. Dietary arachidonic acid decreases the expression of transcripts related to adipocyte development and chronic inflammation in the adipose tissue of juvenile grass carp, Ctenopharyngodon idella. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 122–132. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistics | Crude Analysis | Adjust Analysis | |||
|---|---|---|---|---|---|
| OR (95%CI) | p Value a | OR (95%CI) | p Value a | ||
| APOL6 expression b | |||||
| Low | 31 (50.0%) | 1 | 1 | ||
| High | 31 (50.0%) | 9.35 (2.63, 33.26) | <0.01 | 12.46 (3.11, 50.01) | <0.01 c |
| Treatment regimen | |||||
| PD1 | 35 (56.5%) | 1 | |||
| PD1 plus CTLA4 | 27 (43.5%) | 2.95 (0.96, 9.08) | 0.06 | ||
| Biopsy Site | |||||
| Subcutaneous | 38 (61.3%) | 1 | |||
| Lymph node | 17 (27.4%) | 0.74 (0.23, 2.41) | 0.62 | ||
| Others | 7 (11.3%) | 1.30 (0.22, 7.64) | 0.77 | ||
| Age | 62.3 ± 14.1 | 1.00 (0.96, 1.04) | 0.95 | ||
| Sex | |||||
| Male | 39 (62.9%) | 1 | |||
| Female | 23 (37.1%) | 1.43 (0.48, 4.28) | 0.52 | ||
| Treatment Regimen | APOL6 Expression a | Numbers of Patients | OR (95%CI) b |
|---|---|---|---|
| Paclitaxel | Low | 17 | Ref. |
| High | 17 | 1.00 (0.24, 4.08) | |
| Durvalumab, Olaparib and Paclitaxel | Low | 36 | 0.61 (0.18, 2.13) |
| High | 35 | 2.44 (0.74, 8.11) | |
| pvalue for interaction | 0.12 | ||
| Statistics | Crude Analysis | Adjust Analysis | |||
|---|---|---|---|---|---|
| HR (95%CI) | p Value a | HR (95%CI) | p Value a | ||
| PFS status | |||||
| APOL6 expressionb | |||||
| Low | 39 (53.4%) | 1 | 1 | ||
| High | 34 (46.6%) | 0.27 (0.14, 0.54) | <0.01 | 0.26 (0.13, 0.51) | <0.01 c |
| Treatment regimen | |||||
| PD1 | 41 (56.2%) | 1 | |||
| PD1 plus CTLA4 | 32 (43.8%) | 0.52 (0.27, 1.01) | 0.05 | ||
| Biopsy Site | |||||
| Subcutaneous | 47 (64.4%) | 1 | |||
| Lymph node | 19 (26.0%) | 0.84 (0.41, 1.71) | 0.63 | ||
| Others | 7 (9.6%) | 0.40 (0.10, 1.68) | 0.21 | ||
| Age | 61.6 ± 13.8 | 1.00 (0.97, 1.02) | 0.71 | ||
| Sex | |||||
| Male | 47 (64.4%) | 1 | |||
| Female | 26 (35.6%) | 0.73 (0.38, 1.40) | 0.34 | ||
| OS status | |||||
| APOL6 expressionb | |||||
| Low | 39 (53.4%) | 1 | 1 | ||
| High | 34 (46.6%) | 0.21 (0.09, 0.49) | <0.01 | 0.20 (0.08, 0.48) | <0.01 c |
| Treatment regimen | |||||
| PD1 | 41 (56.2%) | 1 | |||
| PD1 plus CTLA4 | 32 (43.8%) | 0.28(0.10, 0.74) | 0.01 | ||
| Biopsy Site | |||||
| Subcutaneous | 47 (64.4%) | 1 | |||
| Lymph node | 19 (26.0%) | 0.90 (0.38, 2.11) | 0.80 | ||
| Others | 7 (9.6%) | 0.32 (0.04, 2.38) | 0.27 | ||
| Age | 61.6 ± 13.8 | 1.01 (0.98, 1.04) | 0.51 | ||
| Sex | |||||
| Male | 47 (64.4%) | 1 | |||
| Female | 26 (35.6%) | 0.52 (0.22, 1.23) | 0.14 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Chen, Y.; Li, B.; Li, Y.; Liang, X.; Lin, H.; Luo, L.; Chen, T.; Dai, Y.; Pang, W.; et al. Upregulation of Apolipoprotein L6 Improves Tumor Immunotherapy by Inducing Immunogenic Cell Death. Biomolecules 2023, 13, 415. https://doi.org/10.3390/biom13030415
Liu K, Chen Y, Li B, Li Y, Liang X, Lin H, Luo L, Chen T, Dai Y, Pang W, et al. Upregulation of Apolipoprotein L6 Improves Tumor Immunotherapy by Inducing Immunogenic Cell Death. Biomolecules. 2023; 13(3):415. https://doi.org/10.3390/biom13030415
Chicago/Turabian StyleLiu, Kecheng, Yutong Chen, Bixiang Li, Yaning Li, Xinyue Liang, Hai Lin, Lisi Luo, Tianliang Chen, Yalan Dai, Wenzheng Pang, and et al. 2023. "Upregulation of Apolipoprotein L6 Improves Tumor Immunotherapy by Inducing Immunogenic Cell Death" Biomolecules 13, no. 3: 415. https://doi.org/10.3390/biom13030415