

KI04 an Aminoglycosides-Derived Molecule Acts as an Inhibitor of Human Connexin46 Hemichannels Expressed in HeLa Cells
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture and Transfections
2.3. Dye Uptake
2.4. Microscopy Images
2.5. Western Blot
2.6. Whole-Cell Electrophysiology
2.7. Molecular Docking
2.8. Statistical Analyses
3. Results
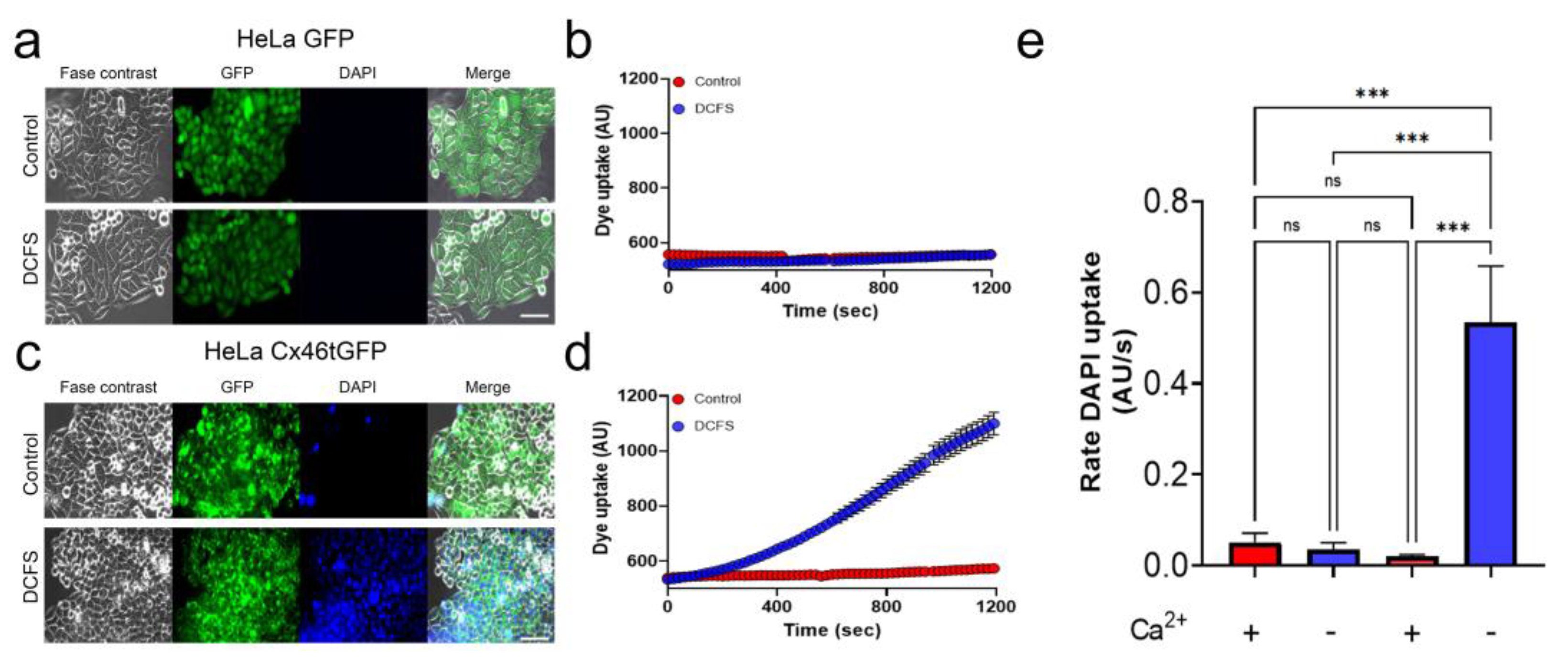
3.1. HeLa Cells Only Showed Hemichannel Activity When Transfected with Cx46tGFP
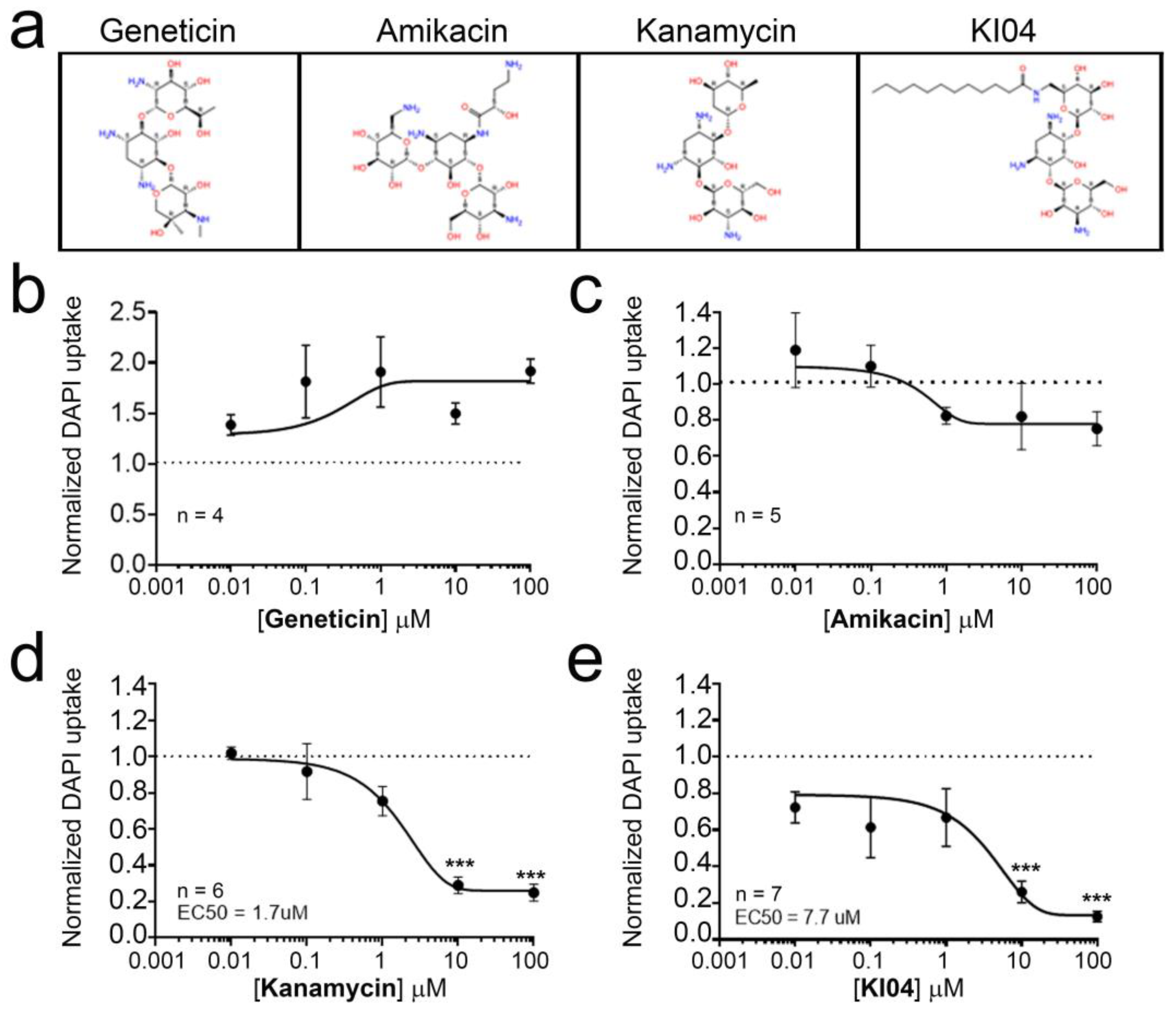
3.2. KI04, a Kanamycin-Derived Aminoglycoside, Acts as Inhibitor of Cx46 Hemichannels
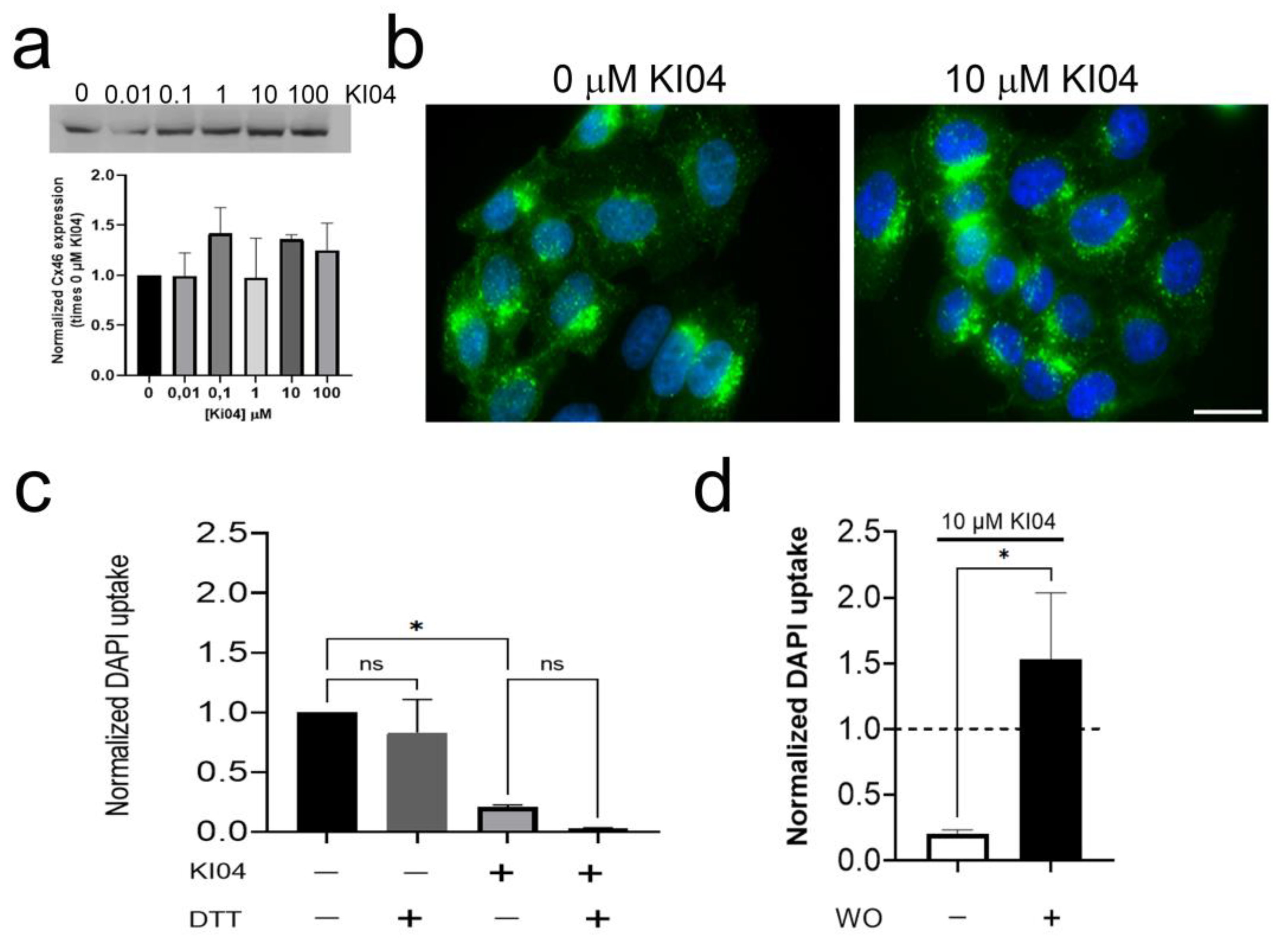
3.3. The KI04 Inhibitory Effect Was Not Associated with Changes in Cx46 Molecular Weight or in Its Cellular Distribution
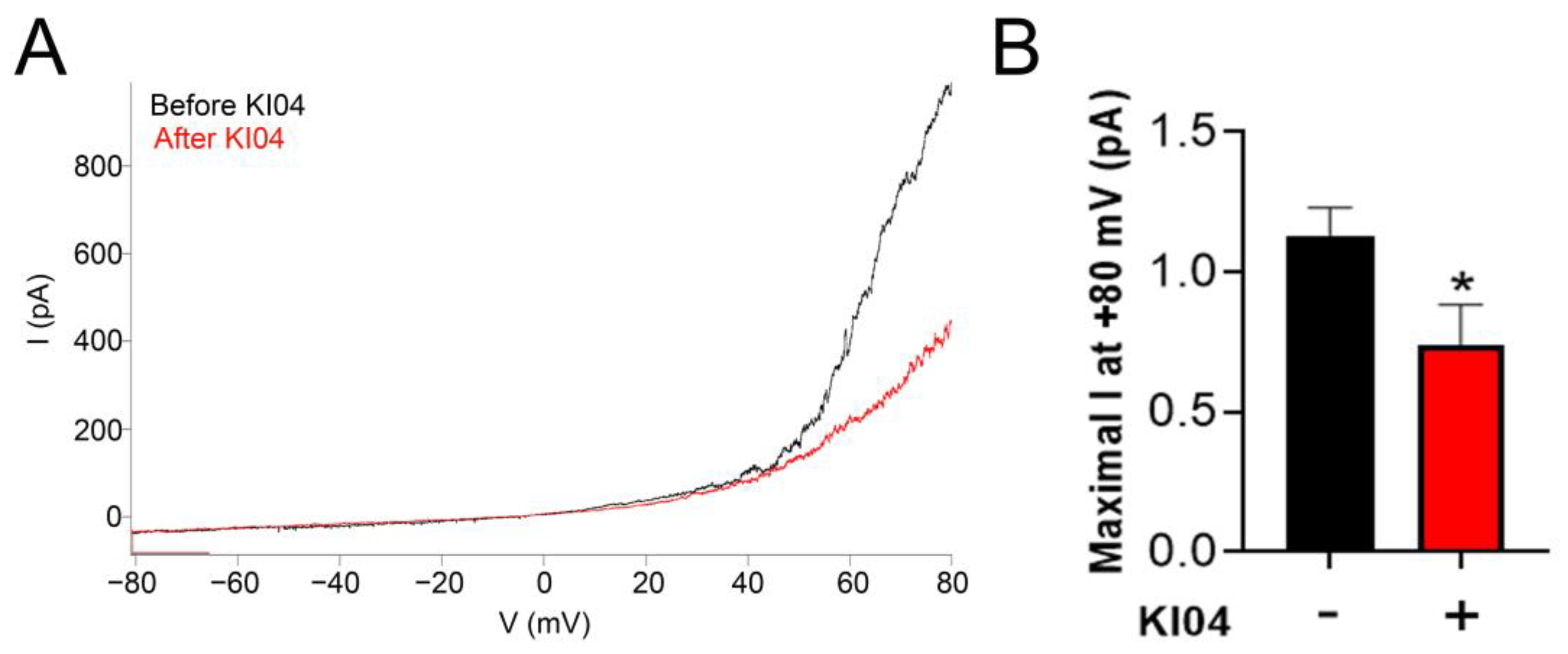
3.4. KI04 Decreases Whole-Cell Cx46 Hemichannel-Mediated Currents
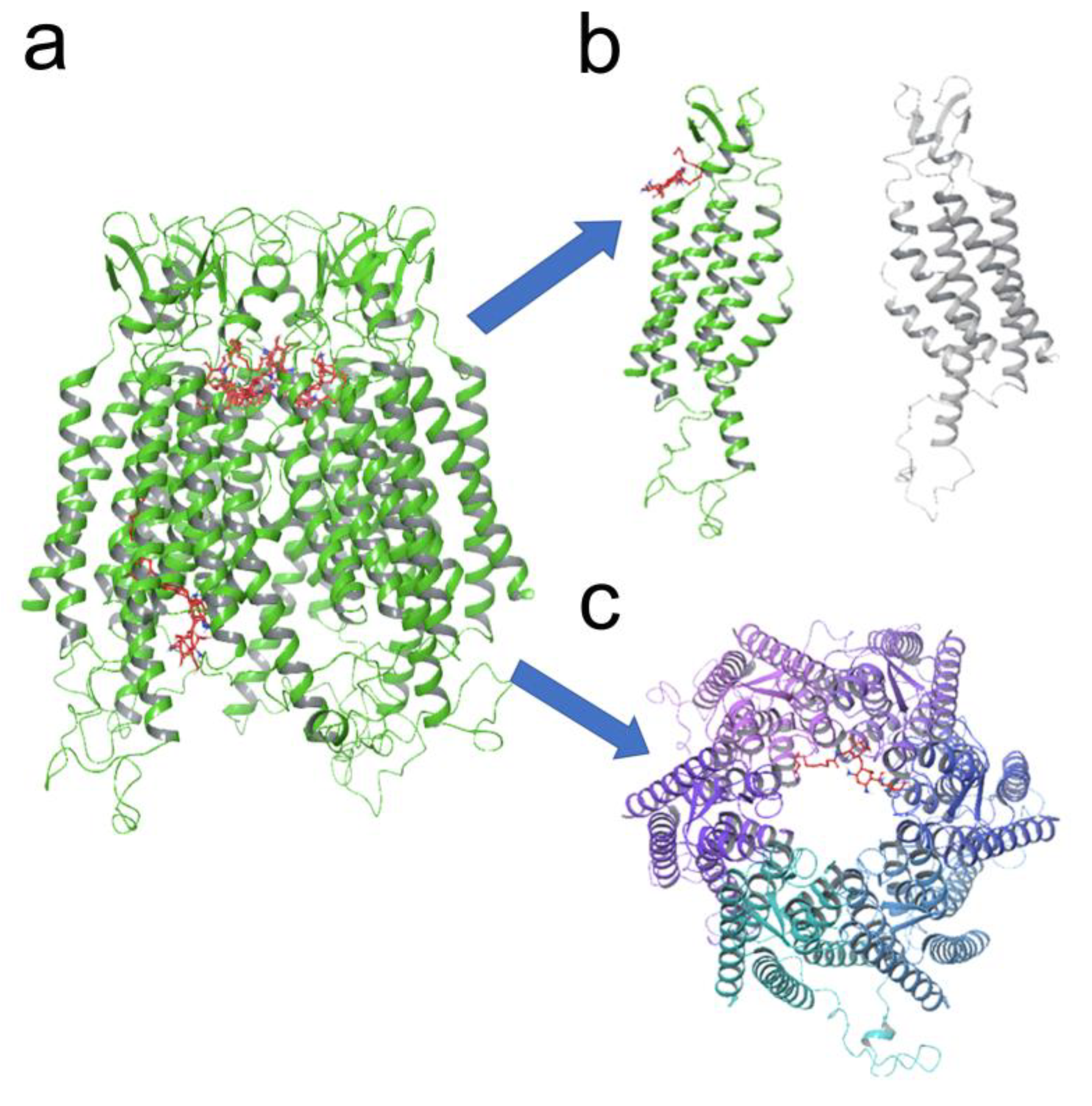
3.5. KI04 Shows Two Putative Binding Sites with Cx46 Hemichannels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sáez, J.C.; Berthoud, V.M.; Brañes, M.C.; Martínez, A.D.; Beyer, E.C. Plasma membrane channels formed by connexins: Their regulation and functions. Physiol. Rev. 2003, 83, 1359–1400. [Google Scholar] [CrossRef] [Green Version]
- Sáez, J.C.; Retamal, M.A.; Basilio, D.; Bukauskas, F.F.; Bennett, M.V.L. Connexin-based gap junction hemichannels: Gating mechanisms. Biochim. Biophys. Acta–Biomembr. 2005, 1711, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stout, C.E.; Costantin, J.L.; Naus, C.C.G.; Charles, A.C. Intercellular Calcium Signaling in Astrocytes via ATP Release through Connexin Hemichannels. J. Biol. Chem. 2002, 277, 10482–10488. [Google Scholar] [CrossRef] [Green Version]
- Stehberg, J.; Moraga-Amaro, R.; Salazar, C.; Becerra, A.; Echeverría, C.; Orellana, J.A.; Bultynck, G.; Ponsaerts, R.; Leybaert, L.; Simon, F.; et al. Release of gliotransmitters through astroglial connexin 43 hemichannels is necessary for fear memory consolidation in the basolateral amygdala. FASEB J. 2012, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saez, J.C.; Connor, J.A.; Spray, D.C.; Bennett, M.V.L. Hepatocyte gap junctions are permeable to the second messenger, inositol 1,4,5-trisphosphate, and to calcium ions. Proc. Natl. Acad. Sci. USA 1989, 86, 2708–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Retamal, M.A.; Reyes, E.P.; García, I.E.; Pinto, B.; Martínez, A.D.; González, C. Diseases associated with leaky hemichannels. Front. Cell. Neurosci. 2015, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Pal, J.A.Y.D.; Liu, X.; Mackay, D.; Shiels, A.; Berthoud, V.M.; Beyer, E.C.; Ebihara, L.; Jay, D.; Liu, X.; Mackay, D.; et al. Connexin46 mutations linked to congenital cataract show loss of gap junction channel function. Am. J. Physiol. Cell Physiol. 2000, 279, 596–602. [Google Scholar] [CrossRef]
- Maeda, S.; Nakagawa, S.; Suga, M.; Yamashita, E.; Oshima, A.; Fujiyoshi, Y.; Tsukihara, T. Structure of the connexin 26 gap junction channel at 3.5 Å resolution. Nature 2009, 458, 597–602. [Google Scholar] [CrossRef]
- Abrams, C.K.; Peinado, A.; Mahmoud, R.; Bocarsly, M.; Zhang, H.; Chang, P.; Botello-Smith, W.M.; Freidin, M.M.; Luo, Y. Alterations at Arg 76 of human connexin 46, a residue associated with cataract formation, cause loss of gap junction formation but preserve hemichannel function. Am. J. Physiol. Physiol. 2018, 315, C623–C635. [Google Scholar] [CrossRef] [Green Version]
- JS, A.; DB, H.; MS, N.; N, M. Isoform-specific phosphorylation-dependent regulation of connexin hemichannels. J. Neurophysiol. 2015, 114, 3014–3022. [Google Scholar] [CrossRef] [Green Version]
- Alaga, K.C.; Crawford, M.; Dagnino, L.; Laird, D.W. Aberrant Cx43 Expression and Mislocalization in Metastatic Human Melanomas. J. Cancer 2017, 8, 1123–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, P.D.; Lau, A.F. Regulation of Gap Junctions by Phosphorylation of Connexins. Arch. Biochem. Biophys. 2000, 384, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Retamal, M.A.; García, I.E.; Pinto, B.I.; Pupo, A.; Báez, D.; Stehberg, J.; Del Rio, R.; González, C. Extracellular cysteine in connexins: Role as redox sensors. Front. Physiol. 2016, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.M.A. Connexin and Pannexin hemichannels are regulated by redox potential. Front. Physiol. 2014, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Guan, Q.W.; Zhou, X.Y.; Xia, Q.X.; Yin, X.X.; Zhou, H.H.; Mao, X.Y. The mutual interplay of redox signaling and connexins. J. Mol. Med. 2021, 99, 933–941. [Google Scholar] [CrossRef]
- Slavi, N.; Wang, Z.; Harvey, L.; Schey, K.L.; Srinivas, M. Identification and Functional Assessment of Age-Dependent Truncations to Cx46 and Cx50 in the Human Lens. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5714. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.L.; Garcia, C.J.; Akpovi, C.D.; Pelletier, R.-M. Distinctive actions of connexin 46 and connexin 50 in anterior pituitary folliculostellate cells. PLoS ONE 2017, 12, e0182495. [Google Scholar] [CrossRef] [Green Version]
- Márquez-Rosado, L.; Singh, D.; Rincón-Arano, H.; Solan, J.L.; Lampe, P.D. CASK (LIN2) interacts with Cx43 in wounded skin and their coexpression affects cell migration. J. Cell Sci. 2012, 125, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Solan, J.L.; Taffet, S.M.; Javier, R.; Lampe, P.D. Connexin 43 interacts with zona occludens-1 and -2 proteins in a cell cycle stage-specific manner. J. Biol. Chem. 2005, 280, 30416–30421. [Google Scholar] [CrossRef] [Green Version]
- Willebrords, J.; Maes, M.; Crespo Yanguas, S.; Vinken, M. Inhibitors of connexin and pannexin channels as potential therapeutics. Pharmacol. Ther. 2017, 180, 144. [Google Scholar] [CrossRef] [Green Version]
- Alfindee, M.N.; Subedi, Y.P.; Fiori, M.C.; Krishnan, S.; Kjellgren, A.; Altenberg, G.A.; Chang, C.W.T. Inhibition of Connexin Hemichannels by New Amphiphilic Aminoglycosides without Antibiotic Activity. ACS Med. Chem. Lett. 2018, 9, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Fiori, M.C.; Krishnan, S.; Kjellgren, A.; Cuello, L.G.; Altenberg, G.A. Inhibition by commercial aminoglycosides of human connexin hemichannels expressed in bacteria. Molecules 2017, 22, 2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subedi, Y.P.; Kjellgren, A.; Roberts, P.; Montgomery, H.; Thackeray, N.; Fiori, M.C.; Altenberg, G.A.; Chang, C.W.T. Amphiphilic aminoglycosides with increased selectivity for inhibition of connexin 43 (Cx43) hemichannels. Eur. J. Med. Chem. 2020, 203, 112602. [Google Scholar] [CrossRef] [PubMed]
- Gerdtzen, Z.P. Modeling metabolic networks for mammalian cell systems: General considerations, modeling strategies, and available tools. Adv. Biochem. Eng. Biotechnol. 2012, 127, 71–108. [Google Scholar] [CrossRef] [PubMed]
- Molina, S.A.; Takemoto, D.J. The role of Connexin 46 promoter in lens and other hypoxic tissues. Commun. Integr. Biol. 2012, 5, 114–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, E.C.; Berthoud, V.M. Connexin hemichannels in the lens. Front. Physiol. 2014, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Hitomi, M.; Deleyrolle, L.P.; Mulkearns-Hubert, E.E.; Jarrar, A.; Li, M.; Sinyuk, M.; Otvos, B.; Brunet, S.; Flavahan, W.A.; Hubert, C.G.; et al. Differential connexin function enhances self-renewal in glioblastoma. Cell Rep. 2015, 11, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, D.; Gakhar, G.; Madgwick, D.; Hurt, A.; Takemoto, D.; Nguyen, T.A. A novel role of gap junction connexin46 protein to protect breast tumors from hypoxia. Int. J. Cancer 2010, 127, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Acuña, R.A.; Varas-Godoy, M.; Herrera-Sepulveda, D.; Retamal, M.A. Connexin46 Expression Enhances Cancer Stem Cell and Epithelial-to-Mesenchymal Transition Characteristics of Human Breast Cancer MCF-7 Cells. Int. J. Mol. Sci. 2021, 22, 12604. [Google Scholar] [CrossRef]
- Paemeleire, K.; Martin, P.E.M.; Coleman, S.L.; Fogarty, K.E.; Carrington, W.A.; Leybaert, L.; Tuft, R.A.; Evans, W.H.; Sanderson, M.J. Intercellular Calcium Waves in HeLa Cells Expressing GFP-labeled Connexin 43, 32, or 26. Mol. Biol. Cell 2000, 11, 1815. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Fiori, M.C.; Fernandez-Olivares, A.; Linsambarth, S.; Peña, F.; Quintana, D.; Stehberg, J.; Altenberg, G.A. 4-Hydroxynonenal induces Cx46 hemichannel inhibition through its carbonylation. Biochim. Biophys. Acta–Mol. Cell Biol. Lipids 2020, 1865, 158705. [Google Scholar] [CrossRef] [PubMed]
- Contreras, J.E.; Saez, J.C.; Bukauskas, F.F.; Bennett, M.V.L. Gating and regulation of connexin 43 (Cx43) hemichannels. Proc. Natl. Acad. Sci. USA 2003, 100, 11388–11393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfahnl, A.; Dahl, G. Gating of cx46 gap junction hemichannels by calcium and voltage. Pflug. Arch. Eur. J. Physiol. 1999, 437, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Solan, J.L.; Lampe, P.D. Connexin43 phosphorylation: Structural changes and biological effects. Biochem. J. 2009, 419, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, D.L.; Ebihara, L.; Takemoto, L.J.; Swenson, K.I.; Goodenough, D.A. Connexin46, a novel lens gap junction protein, induces voltage-gated currents in nonjunctional plasma membrane of Xenopus oocytes. J. Cell Biol. 1991, 115, 1077–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.X.; Paul, D.L.; Goodenough, D.A. Posttranslational phosphorylation of lens fiber connexin46: A slow occurrence. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3558–3565. [Google Scholar]
- Bao, X.; Reuss, L.; Altenberg, G.A. Regulation of Purified and Reconstituted Connexin 43 Hemichannels by Protein Kinase C-mediated Phosphorylation of Serine 368. J. Biol. Chem. 2004, 279, 20058–20066. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.T.; Maldonado-Pérez, D.; Hollins, L.; Riccardi, D. Aminoglycosides induce acute cell signaling and chronic cell death in renal cells that express the calcium-sensing receptor. J. Am. Soc. Nephrol. 2005, 16, 1236–1244. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.T.; McLarnon, S.J.; Riccardi, D. Aminoglycosides increase intracellular calcium levels and ERK activity in proximal tubular OK cells expressing the extracellular calcium-sensing receptor. J. Am. Soc. Nephrol. 2002, 13, 1481–1489. [Google Scholar] [CrossRef] [Green Version]
- Rybak, L.P.; Brenner, M.J. Aminoglycoside-Induced Oxidative Stress: Pathways and Protection. Free. Radic. ENT Pathol. 2015, 195–216. [Google Scholar] [CrossRef]
- Fernández-Olivares, A.; Durán-Jara, E.; Verdugo, D.A.; Fiori, M.C.; Altenberg, G.A.; Stehberg, J.; Alfaro, I.; Calderón, J.F.; Retamal, M.A. Extracellular cysteines are critical to form functional Cx46 hemichannels. Int. J. Mol. Sci. 2022, 23, 7252. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, V.A.; Retamal, M.A.; Cea, L.A.; Salas, J.D.; Vargas, A.A.; Verdugo, C.A.; Jara, O.; Martínez, A.D.; Sáez, J.C. Extracellular gentamicin reduces the activity of connexin hemichannels and interferes with purinergic Ca2+ signaling in HeLa cells. Front. Cell. Neurosci. 2014, 8, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.B.; Haddad, B.G.; O’Neill, S.E.; Chorev, D.S.; Yoshioka, C.C.; Robinson, C.V.; Zuckerman, D.M.; Reichow, S.L. Structure of native lens connexin 46/50 intercellular channels by cryo-EM. Nature 2018, 564, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.C.; Purdy, M.D.; Baker, K.A.; Acharya, C.; Mcintire, W.E.; Stevens, R.C.; Zhang, Q.; Harris, A.L.; Abagyan, R.; Yeager, M. An electrostatic mechanism for Ca2+-mediated regulation of gap junction channels. Nat. Commun. 2016, 7, 8770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hao, H. Conserved glycine at position 45 of major cochlear connexins constitutes a vital component of the Ca2+ sensor for gating of gap junction hemichannels. Biochem. Biophys. Res. Commun. 2013, 436, 424–429. [Google Scholar] [CrossRef]
- Takeichi, T.; Sugiura, K.; Hsu, C.K.; Nomura, T.; Takama, H.; Simpson, M.A.; Shimizu, H.; McGrath, J.A.; Akiyama, M. Erythrokeratoderma variabilis caused by p.Gly45Glu in connexin 31: Importance of the first extracellular loop glycine residue for gap junction function. Acta Derm. Venereol. 2016, 96, 557–559. [Google Scholar] [CrossRef] [Green Version]
- Griffith, A.J.; Yang, Y.; Pryor, S.P.; Park, H.J.; Jabs, E.W.; Nadol, J.B.; Russell, L.J.; Wasserman, D.I.; Richard, G.; Adams, J.C.; et al. Cochleosaccular dysplasia associated with a connexin 26 mutation in keratitis-ichthyosis-deafness syndrome. Laryngoscope 2006, 116, 1404–1408. [Google Scholar] [CrossRef]
- Tong, X.; Aoyama, H.; Tsukihara, T.; Bai, D. Charge at the 46th residue of connexin 50 is crucial for the gap-junctional unitary conductance and transjunctional voltage-dependent gating. J. Physiol. 2014, 592, 5187. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, V.M.; Ngezahayo, A. Focus on Lens Connexins; BioMed Central Ltd.: London, UK, 2017; Volume 18, p. 6. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-W.T.; Poudyal, N.; Verdugo, D.A.; Peña, F.; Stehberg, J.; Retamal, M.A. KI04 an Aminoglycosides-Derived Molecule Acts as an Inhibitor of Human Connexin46 Hemichannels Expressed in HeLa Cells. Biomolecules 2023, 13, 411. https://doi.org/10.3390/biom13030411
Chang C-WT, Poudyal N, Verdugo DA, Peña F, Stehberg J, Retamal MA. KI04 an Aminoglycosides-Derived Molecule Acts as an Inhibitor of Human Connexin46 Hemichannels Expressed in HeLa Cells. Biomolecules. 2023; 13(3):411. https://doi.org/10.3390/biom13030411
Chicago/Turabian StyleChang, Cheng-Wei T., Naveena Poudyal, Daniel A. Verdugo, Francisca Peña, Jimmy Stehberg, and Mauricio A. Retamal. 2023. "KI04 an Aminoglycosides-Derived Molecule Acts as an Inhibitor of Human Connexin46 Hemichannels Expressed in HeLa Cells" Biomolecules 13, no. 3: 411. https://doi.org/10.3390/biom13030411