
High-Intensity Focused Ultrasound Decreases Subcutaneous Fat Tissue Thickness by Increasing Apoptosis and Autophagy
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. HIFU System and Applied to Rats
2.3. In Vitro Model
2.4. Nucleus, Cytosol, and Mitochondria Isolation and Protein Preparation
2.5. Western Blot
2.6. Preparation of Paraffin-Embedded Skin and SAT Tissue Sections
2.7. TdT-Mediated dUTP Nick-End Labeling Assay
2.8. Hematoxylin and Eosin Staining
2.9. Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.10. Statistical Analysis
3. Results
3.1. HIFU Decreased the Thickness of the SAT
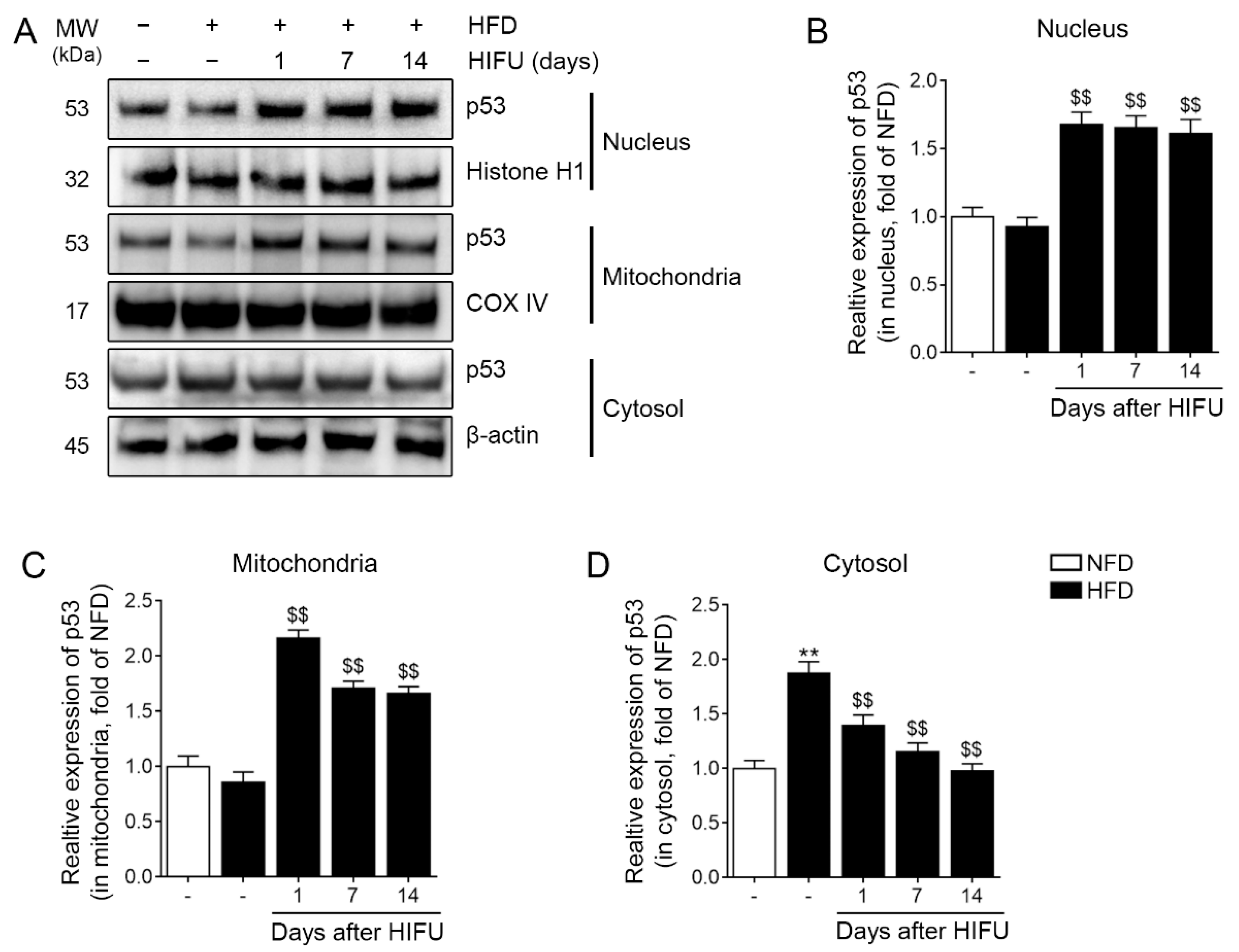
3.2. HIFU Increased the Expression of Nuclear and Mitochondrial p53 and Decreased the Expression of Cytoplasmic p53 in the SAT
3.3. HIFU Decreased BCL2/BCL-xL Expression and Increased BAX/BAK Expression and Cytochrome C Release
3.4. HIFU Decreased BCL2/BCL-xL Expression and Increased BAX/BAK Expression and Cytochrome C Release via Modulation of p53
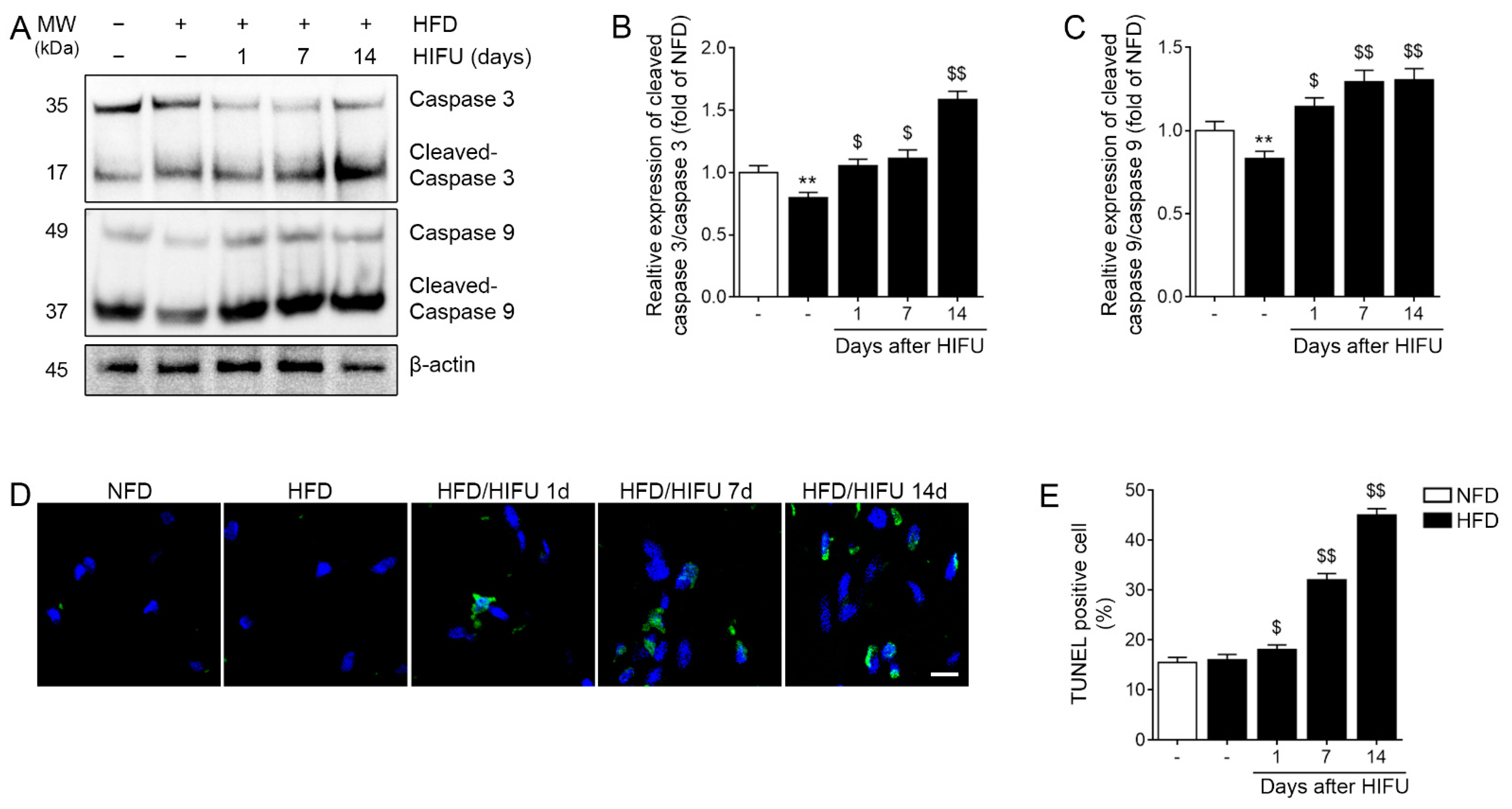
3.5. HIFU Induced Caspase 3/9 Activation and Apoptosis in the SAT
3.6. HIFU Induced p53 Change Involved in Caspase 3/9 Activation
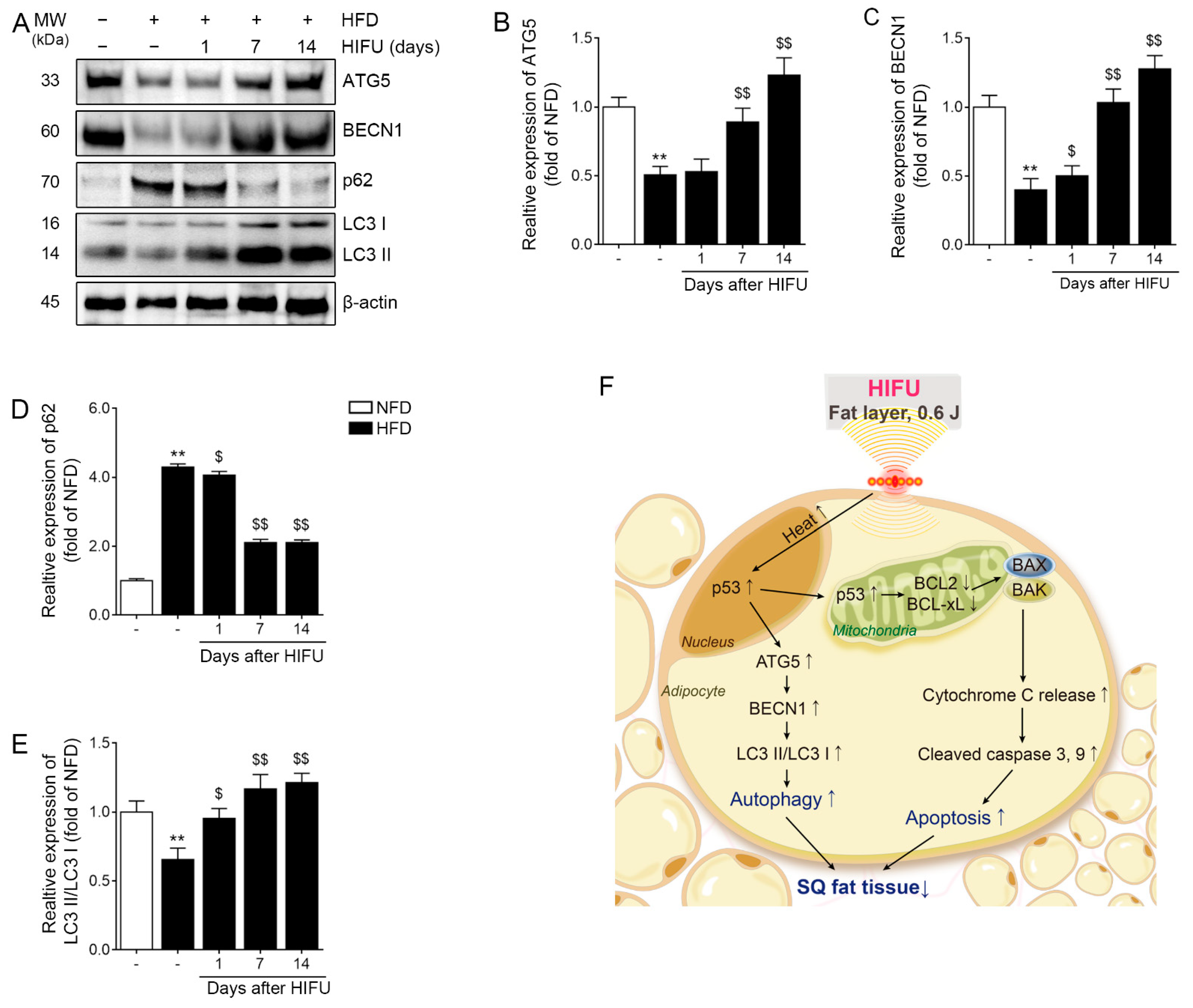
3.7. HIFU Increased Autophagy in the SAT
3.8. HIFU Increased Autophagy via Modulation of p53
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fatemi, A. High-intensity focused ultrasound effectively reduces adipose tissue. Semin. Cutan. Med. Surg. 2009, 28, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Haar, G.T.; Coussios, C. High intensity focused ultrasound: Physical principles and devices. Int. J. Hyperth. 2007, 23, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Jewell, M.L.; Solish, N.J.; Desilets, C.S. Noninvasive body sculpting technologies with an emphasis on high-intensity focused ultrasound. Aesthetic Plast. Surg. 2011, 35, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Xu, G. Damage effect of high-intensity focused ultrasound on breast cancer tissues and their vascularities. World J. Surg. Oncol. 2016, 14, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engin, K. Biological rationale and clinical experience with hyperthermia. Control. Clin. Trials 1996, 17, 316–342. [Google Scholar] [CrossRef]
- Lepock, J.R. Role of nuclear protein denaturation and aggregation in thermal radiosensitization. Int. J. Hyperth. 2004, 20, 115–130. [Google Scholar] [CrossRef]
- Roti Roti, J.L. Cellular responses to hyperthermia (40-46 degrees C): Cell killing and molecular events. Int. J. Hyperth. 2008, 24, 3–15. [Google Scholar] [CrossRef]
- Gu, Z.T.; Wang, H.; Li, L.; Liu, Y.S.; Deng, X.B.; Huo, S.F.; Yuan, F.F.; Liu, Z.F.; Tong, H.S.; Su, L. Heat stress induces apoptosis through transcription-independent p53-mediated mitochondrial pathways in human umbilical vein endothelial cell. Sci. Rep. 2014, 4, 4469. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Kim, Y.C.; Song, S.B.; Kim, Y.S. Stabilization and translocation of p53 to mitochondria is linked to Bax translocation to mitochondria in simvastatin-induced apoptosis. Biochem. Biophys. Res. Commun. 2010, 391, 1592–1597. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Green, D.R. Dissecting p53-dependent apoptosis. Cell Death Differ. 2006, 13, 994–1002. [Google Scholar] [CrossRef] [Green Version]
- Vaseva, A.V.; Moll, U.M. The mitochondrial p53 pathway. Biochim. Biophys. Acta 2009, 1787, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Kroemer, G. Cytoplasmic functions of the tumour suppressor p53. Nature 2009, 458, 1127–1130. [Google Scholar] [CrossRef] [Green Version]
- Szegezdi, E.; Macdonald, D.C.; Ní Chonghaile, T.; Gupta, S.; Samali, A. Bcl-2 family on guard at the ER. Am. J. Physiol. Cell Physiol. 2009, 296, C941–C953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, J.C. Proapoptotic multidomain Bcl-2/Bax-family proteins: Mechanisms, physiological roles, and therapeutic opportunities. Cell Death Differ. 2006, 13, 1378–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef] [Green Version]
- Antonsson, B.; Montessuit, S.; Lauper, S.; Eskes, R.; Martinou, J.C. Bax oligomerization is required for channel-forming activity in liposomes and to trigger cytochrome c release from mitochondria. Biochem. J. 2000, 345 Pt 2, 271–278. [Google Scholar] [CrossRef]
- Wolter, K.G.; Hsu, Y.T.; Smith, C.L.; Nechushtan, A.; Xi, X.G.; Youle, R.J. Movement of Bax from the cytosol to mitochondria during apoptosis. J. Cell Biol. 1997, 139, 1281–1292. [Google Scholar] [CrossRef]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a human protein homologous to C. elegans CED-4, participates in cytochrome c-dependent activation of caspase-3. Cell 1997, 90, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zou, H.; Slaughter, C.; Wang, X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 1997, 89, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, E. Autophagy and p53. Cold Spring Harb. Perspect. Med. 2016, 6, a026120. [Google Scholar] [CrossRef]
- Yip, K.W.; Reed, J.C. Bcl-2 family proteins and cancer. Oncogene 2008, 27, 6398–6406. [Google Scholar] [CrossRef] [Green Version]
- Maiuri, M.C.; Le Toumelin, G.; Criollo, A.; Rain, J.C.; Gautier, F.; Juin, P.; Tasdemir, E.; Pierron, G.; Troulinaki, K.; Tavernarakis, N.; et al. Functional and physical interaction between Bcl-X(L) and a BH3-like domain in Beclin-1. EMBO J. 2007, 26, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Rosenbluth, J.M.; Mays, D.J.; Pino, M.F.; Tang, L.J.; Pietenpol, J.A. A gene signature-based approach identifies mTOR as a regulator of p73. Mol. Cell. Biol. 2008, 28, 5951–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbluth, J.M.; Pietenpol, J.A. mTOR regulates autophagy-associated genes downstream of p73. Autophagy 2009, 5, 114–116. [Google Scholar] [CrossRef] [Green Version]
- Summers, C.M.; Valentine, R.J. Acute Heat Exposure Alters Autophagy Signaling in C2C12 Myotubes. Front. Physiol. 2020, 10, 1521. [Google Scholar] [CrossRef]
- Oh, S.; Kim, H.M.; Batsukh, S.; Sun, H.J.; Kim, T.; Kang, D.; Son, K.H.; Byun, K. High-Intensity Focused Ultrasound Induces Adipogenesis via Control of Cilia in Adipose-Derived Stem Cells in Subcutaneous Adipose Tissue. Int. J. Mol. Sci. 2022, 23, 8866. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, X.; Wang, Y.; Guo, Y. Endoplasmic reticulum stress and autophagy are involved in adipocyte-induced fibrosis in hepatic stellate cells. Mol. Cell. Biochem. 2021, 476, 2527–2538. [Google Scholar] [CrossRef]
- Roshanzadeh, A.; Yadav, A.K.; Pydi, S.P.; Kimura, T.; Jang, B.C. Expression and role of β3-adrenergic receptor during the differentiation of 3T3-L1 preadipocytes into adipocytes. Biology 2022, 11, 772. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Ding, Z.; Ming, J. Interleukin 7 inhibit autophagy via P53 regulated AMPK/mTOR signaling pathway in non-small cell lung cancer. Sci. Rep. 2022, 12, 11208. [Google Scholar] [CrossRef]
- Yoshii, S.R.; Mizushima, N. Monitoring and Measuring Autophagy. Int. J. Mol. Sci. 2017, 18, 1865. [Google Scholar] [CrossRef]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell. Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, H.; Ge, H.; Yang, Z. AG-1031 induced autophagic cell death and apoptosis in C6 glioma cells associated with Notch-1 signaling pathway. J. Cell. Biochem. 2018, 119, 5893–5903. [Google Scholar] [CrossRef] [PubMed]
- Lefort, K.; Mandinova, A.; Ostano, P.; Kolev, V.; Calpini, V.; Kolfschoten, I.; Devgan, V.; Lieb, J.; Raffoul, W.; Hohl, D.; et al. Notch1 is a p53 target gene involved in human keratinocyte tumor suppression through negative regulation of ROCK1/2 and MRCKalpha kinases. Genes Dev. 2007, 21, 562–577. [Google Scholar] [CrossRef] [Green Version]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, C.; Tao, Y.; Jiang, L.; Wen, X.; Liao, J. Assay technologies for apoptosis and autophagy. Med. Drug Discov. 2021, 11, 100100. [Google Scholar] [CrossRef]
- Martin, S.J.; Henry, C.M.; Cullen, S.P. A perspective on mammalian caspases as positive and negative regulators of inflammation. Mol. Cell 2012, 46, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocco, R.E.; Ucker, D.S. Distinct modes of macrophage recognition for apoptotic and necrotic cells are not specified exclusively by phosphatidylserine exposure. Mol. Biol. Cell 2001, 12, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Birge, R.B.; Ucker, D.S. Innate apoptotic immunity: The calming touch of death. Cell Death Differ. 2008, 15, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol. Rev. 2010, 90, 1383–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, D.R.; Levine, B. To be or not to be? How selective autophagy and cell death govern cell fate. Cell 2014, 157, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Yin, B.; Wang, X.; Xia, G.; Shen, Z.; Gu, W.; Wu, M. Effects of the Notch1 signaling pathway on human lung cancer A549 cells. Exp. Lung Res. 2017, 43, 208–216. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byun, K.-A.; Park, H.J.; Oh, S.; Batsukh, S.; Sun, H.J.; Kim, T.; Kim, S.; Kang, D.; Son, K.H.; Byun, K. High-Intensity Focused Ultrasound Decreases Subcutaneous Fat Tissue Thickness by Increasing Apoptosis and Autophagy. Biomolecules 2023, 13, 392. https://doi.org/10.3390/biom13020392
Byun K-A, Park HJ, Oh S, Batsukh S, Sun HJ, Kim T, Kim S, Kang D, Son KH, Byun K. High-Intensity Focused Ultrasound Decreases Subcutaneous Fat Tissue Thickness by Increasing Apoptosis and Autophagy. Biomolecules. 2023; 13(2):392. https://doi.org/10.3390/biom13020392
Chicago/Turabian StyleByun, Kyung-A, Hyun Jun Park, Seyeon Oh, Sosorburam Batsukh, Hye Jin Sun, Taehui Kim, Sunggeun Kim, Donghwan Kang, Kuk Hui Son, and Kyunghee Byun. 2023. "High-Intensity Focused Ultrasound Decreases Subcutaneous Fat Tissue Thickness by Increasing Apoptosis and Autophagy" Biomolecules 13, no. 2: 392. https://doi.org/10.3390/biom13020392