

Evolution and Role of Proteases in Campylobacter jejuni Lifestyle and Pathogenesis

Abstract
:1. Introduction
2. Proteases
2.1. Serine Proteases
2.1.1. Serine Protease HtrA (Cj1228c)
2.1.2. Serine Protease Cj1365c
2.1.3. Serine Protease Cj0511
2.2. AAA+ Group Proteases
2.2.1. ClpP Protease (Cj0192c)
2.2.2. Lon (Cj1073c)
2.2.3. FtsH (Cj1116c)
2.3. PqqE Protease (Cj0805)
3. Peptidases
3.1. Proline Aminopeptidase PepP (Cj0653c)
3.2. Oligopeptidase PepF (Cj1099)
3.3. C26 Peptidase (Cj1417c)
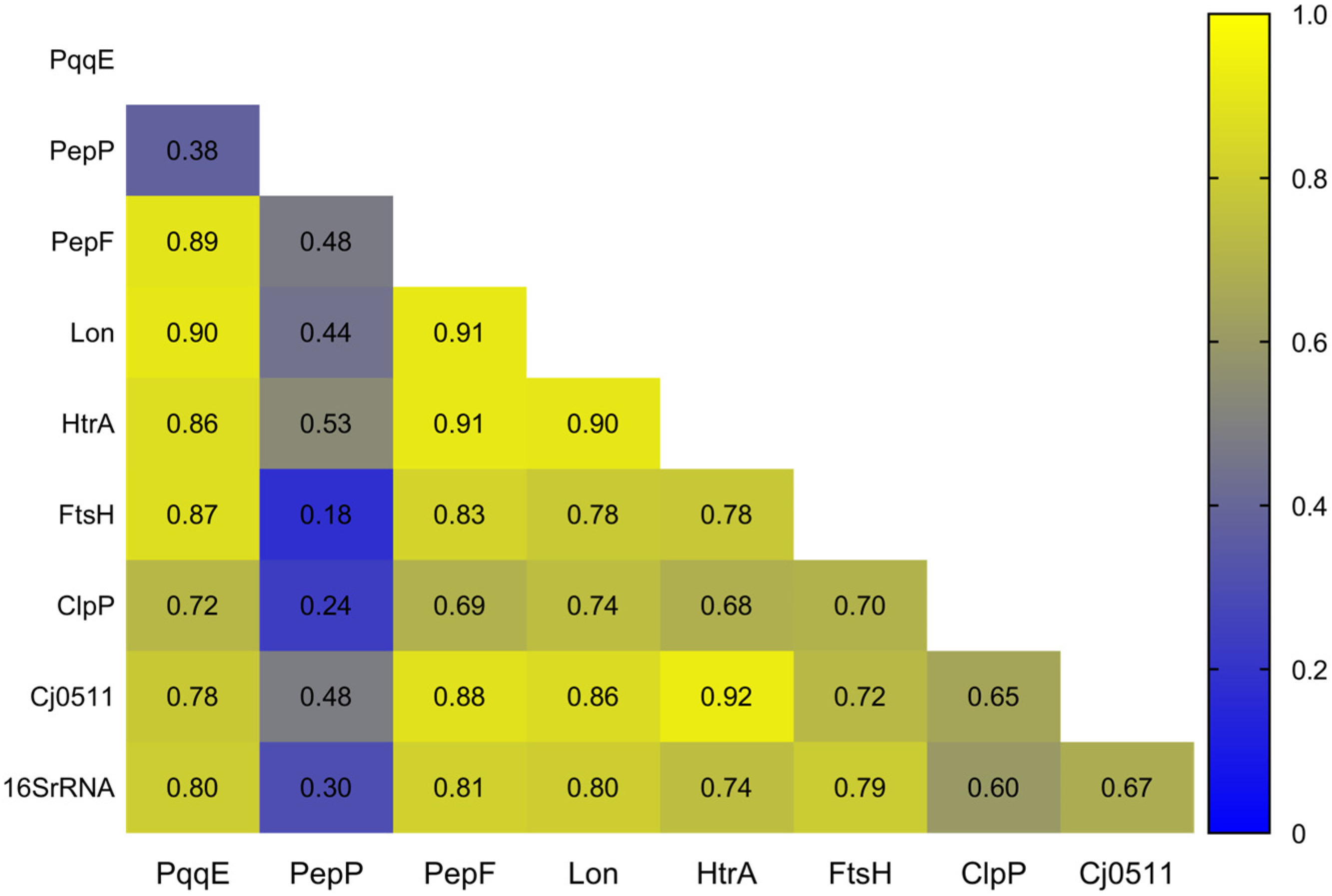
4. Evolution of Proteases and Peptidases in the Campylobacter Genus
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- European Food Safety, Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. Efsa J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Laughlin, M.E.; Chatham-Stephens, K.; Geissler, A.L. Campylobacteriosis. In CDC Yellow Book 2020: Health Information for International Travel; Brunette, G.W., Nemhauser, J.B., Eds.; Oxford University Press: New York, NY, USA, 2019. [Google Scholar]
- Blaser, M.J. Epidemiologic and clinical features of Campylobacter jejuni infections. J. Infect. Dis. 1997, 176, S103–S105. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human Campylobacteriosis-A Serious Infectious Threat in a One Health Perspective. In Fighting Campylobacter Infections; Current Topics in Microbiology and Immunology; Springer: Cham, Switzerland, 2021; Volume 431, pp. 1–23. [Google Scholar] [CrossRef]
- Burnham, P.M.; Hendrixson, D.R. Campylobacter jejuni: Collective components promoting a successful enteric lifestyle. Nat. Rev. Microbiol. 2018, 16, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Murine Models for the Investigation of Colonization Resistance and Innate Immune Responses in Campylobacter jejuni Infections. Curr. Top. Microbiol. Immunol. 2021, 431, 233–263. [Google Scholar] [CrossRef] [PubMed]
- Wakerley, B.R.; Uncini, A.; Yuki, N.; Group, G.B.S.C.; Group, G.B.S.C. Guillain-Barre and Miller Fisher syndromes--new diagnostic classification. Nat. Rev. Neurol. 2014, 10, 537–544. [Google Scholar] [CrossRef]
- de Sa, F.D.L.; Schulzke, J.D.; Bucker, R. Diarrheal Mechanisms and the Role of Intestinal Barrier Dysfunction in Campylobacter Infections. Curr. Top. Microbiol. Immunol. 2021, 431, 203–231. [Google Scholar] [CrossRef]
- Mozina, S.S.; Kurincic, M.; Klancnik, A.; Mavri, A. Campylobacter and its multi-resistance in the food chain. Trends Food Sci. Technol. 2011, 22, 91–98. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.I.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Hofreuter, D.; Novik, V.; Galan, J.E. Metabolic diversity in Campylobacter jejuni enhances specific tissue colonization. Cell Host Microbe 2008, 4, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Cox, N.A.; Richardson, L.J.; Buhr, R.J.; Fedorka-Cray, P.J. Campylobacter species occurrence within internal organs and tissues of commercial caged Leghorn laying hens. Poult. Sci. 2009, 88, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Croinin, T.O.; Backert, S. Host epithelial cell invasion by Campylobacter jejuni: Trigger or zipper mechanism? Front. Cell. Infect. Microbiol. 2012, 2, 25. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Sharafutdinov, I.; Harrer, A.; Soltan Esmaeili, D.; Linz, B.; Backert, S. Campylobacter Virulence Factors and Molecular Host-Pathogen Interactions. Curr. Top. Microbiol. Immunol. 2021, 431, 169–202. [Google Scholar] [CrossRef]
- Mourkas, E.; Taylor, A.J.; Meric, G.; Bayliss, S.C.; Pascoe, B.; Mageiros, L.; Calland, J.K.; Hitchings, M.D.; Ridley, A.; Vidal, A.; et al. Agricultural intensification and the evolution of host specialism in the enteric pathogen Campylobacter jejuni. Proc. Natl. Acad. Sci. USA 2020, 117, 11018–11028. [Google Scholar] [CrossRef]
- Young, G.M.; Schmiel, D.H.; Miller, V.L. A new pathway for the secretion of virulence factors by bacteria: The flagellar export apparatus functions as a protein-secretion system. Proc. Natl. Acad. Sci. USA 1999, 96, 6456–6461. [Google Scholar] [CrossRef]
- Lindmark, B.; Rompikuntal, P.K.; Vaitkevicius, K.; Song, T.; Mizunoe, Y.; Uhlin, B.E.; Guerry, P.; Wai, S.N. Outer membrane vesicle-mediated release of cytolethal distending toxin (CDT) from Campylobacter jejuni. BMC Microbiol. 2009, 9, 220. [Google Scholar] [CrossRef]
- Boehm, M.; Hoy, B.; Rohde, M.; Tegtmeyer, N.; Baek, K.T.; Oyarzabal, O.A.; Brondsted, L.; Wessler, S.; Backert, S. Rapid paracellular transmigration of Campylobacter jejuni across polarized epithelial cells without affecting TER: Role of proteolytic-active HtrA cleaving E-cadherin but not fibronectin. Gut Pathog. 2012, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Sharafutdinov, I.; Esmaeili, D.S.; Harrer, A.; Tegtmeyer, N.; Sticht, H.; Backert, S. Campylobacter jejuni Serine Protease HtrA Cleaves the Tight Junction Component Claudin-8. Front. Cell. Infect. Microbiol. 2020, 10, 590186. [Google Scholar] [CrossRef] [PubMed]
- Boehm, M.; Simson, D.; Escher, U.; Schmidt, A.-M.; Bereswill, S.; Tegtmeyer, N.; Backert, S.; Heimesaat, M.M. Function of Serine Protease HtrA in the Lifecycle of the Foodborne Pathogen Campylobacter jejuni. Eur. J. Microbiol. Immunol. 2018, 8, 70–77. [Google Scholar] [CrossRef]
- Lebrun, I.; Marques-Porto, R.; Pereira, A.S.; Pereira, A.; Perpetuo, E.A. Bacterial toxins: An overview on bacterial proteases and their action as virulence factors. Mini Rev. Med. Chem. 2009, 9, 820–828. [Google Scholar] [CrossRef]
- Solanki, P.; Putatunda, C.; Kumar, A.; Bhatia, R.; Walia, A. Microbial proteases: Ubiquitous enzymes with innumerable uses. 3 Biotech 2021, 11, 428. [Google Scholar] [CrossRef]
- Viana, F.; Peringathara, S.S.; Rizvi, A.; Schroeder, G.N. Host manipulation by bacterial type III and type IV secretion system effector proteases. Cell. Microbiol. 2021, 23, e13384. [Google Scholar] [CrossRef]
- Mohiuddin, S.G.; Ghosh, S.; Ngo, H.G.; Sensenbach, S.; Karki, P.; Dewangan, N.K.; Angardi, V.; Orman, M.A. Cellular Self-Digestion and Persistence in Bacteria. Microorganisms 2021, 9, 2269. [Google Scholar] [CrossRef]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Butcher, J.; Stintzi, A. Nutrient acquisition and metabolism by Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2012, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Bittner, L.M.; Arends, J.; Narberhaus, F. ATP-dependent proteases in bacteria. Biopolymers 2016, 105, 505–517. [Google Scholar] [CrossRef]
- Frees, D.; Brondsted, L.; Ingmer, H. Bacterial proteases and virulence. Subcell Biochem. 2013, 66, 161–192. [Google Scholar] [CrossRef]
- Backert, S.; Bernegger, S.; Skorko-Glonek, J.; Wessler, S. Extracellular HtrA serine proteases: An emerging new strategy in bacterial pathogenesis. Cell. Microbiol. 2018, 20, e12845. [Google Scholar] [CrossRef] [PubMed]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2000, 82, 427–446. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kim, K.K. Structure and function of HtrA family proteins, the key players in protein quality control. J. Biochem. Mol. Biol. 2005, 38, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Krojer, T.; Sawa, J.; Huber, R.; Clausen, T. HtrA proteases have a conserved activation mechanism that can be triggered by distinct molecular cues. Nat. Struct. Mol. Biol. 2010, 17, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Zarzecka, U.; Grinzato, A.; Kandiah, E.; Cysewski, D.; Berto, P.; Skorko-Glonek, J.; Zanotti, G.; Backert, S. Functional analysis and cryo-electron microscopy of Campylobacter jejuni serine protease HtrA. Gut Microbes 2020, 12, e1810532. [Google Scholar] [CrossRef] [PubMed]
- Brondsted, L.; Andersen, M.T.; Parker, M.; Jorgensen, K.; Ingmer, H. The HtrA protease of Campylobacter jejuni is required for heat and oxygen tolerance and for optimal interaction with human epithelial cells. Appl. Environ. Microbiol. 2005, 71, 3205–3212. [Google Scholar] [CrossRef]
- Baek, K.T.; Vegge, C.S.; Skorko-Glonek, J.; Brondsted, L. Different contributions of HtrA protease and chaperone activities to Campylobacter jejuni stress tolerance and physiology. Appl. Environ. Microbiol. 2011, 77, 57–66. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Bereswill, S. Murine infection models for the investigation of Campylobacter jejuni-host interactions and pathogenicity. Berl. Munch Tierarztl. Wochenschr. 2015, 128, 98–103. [Google Scholar] [PubMed]
- Heimesaat, M.M.; Alutis, M.; Grundmann, U.; Fischer, A.; Tegtmeyer, N.; Bohm, M.; Kuhl, A.A.; Gobel, U.B.; Backert, S.; Bereswill, S. The role of serine protease HtrA in acute ulcerative enterocolitis and extra-intestinal immune responses during Campylobacter jejuni infection of gnotobiotic IL-10 deficient mice. Front. Cell. Infect. Microbiol. 2014, 4, 77. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Escher, U.; Mousavi, S.; Boehm, M.; Backert, S.; Bereswill, S.; Heimesaat, M.M. Protease Activity of Campylo-bacter jejuni HtrA Modulates Distinct Intestinal and Systemic Immune Responses in Infected Secondary Abiotic IL-10 Deficient Mice. Front. Cell. Infect. Microbiol. 2019, 9, 79. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Fischer, A.; Alutis, M.; Grundmann, U.; Boehm, M.; Tegtmeyer, N.; Gobel, U.B.; Kuhl, A.A.; Bereswill, S.; Backert, S. The impact of serine protease HtrA in apoptosis, intestinal immune responses and extra-intestinal histopathology during Campylobacter jejuni infection of infant mice. Gut Pathog. 2014, 6, 16. [Google Scholar] [CrossRef]
- Baek, K.T.; Vegge, C.S.; Brondsted, L. HtrA chaperone activity contributes to host cell binding in Campylobacter jejuni. Gut Pathog. 2011, 3, 13. [Google Scholar] [CrossRef]
- Boehm, M.; Haenel, I.; Hoy, B.; Brondsted, L.; Smith, T.G.; Hoover, T.; Wessler, S.; Tegtmeyer, N. Extracellular secretion of protease HtrA from Campylobacter jejuni is highly efficient and independent of its protease activity and flagellum. Eur. J. Microbiol. Immunol. 2013, 3, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Boehm, M.; Lind, J.; Backert, S.; Tegtmeyer, N. Campylobacter jejuni serine protease HtrA plays an important role in heat tolerance, oxygen resistance, host cell adhesion, invasion, and transmigration. Eur. J. Microbiol. Immunol. 2015, 5, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Simson, D.; Boehm, M.; Backert, S. HtrA-dependent adherence and invasion of Campylobacter jejuni in human vs avian cells. Lett. Appl. Microbiol. 2020, 70, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Elmi, A.; Watson, E.; Sandu, P.; Gundogdu, O.; Mills, D.C.; Inglis, N.F.; Manson, E.; Imrie, L.; Bajaj-Elliott, M.; Wren, B.W.; et al. Campylobacter jejuni outer membrane vesicles play an important role in bacterial interactions with human intestinal epithelial cells. Infect. Immun. 2012, 80, 4089–4098. [Google Scholar] [CrossRef]
- Elmi, A.; Nasher, F.; Jagatia, H.; Gundogdu, O.; Bajaj-Elliott, M.; Wren, B.; Dorrell, N. Campylobacter jejuni outer membrane vesicle-associated proteolytic activity promotes bacterial invasion by mediating cleavage of intestinal epithelial cell E-cadherin and occludin. Cell. Microbiol. 2016, 18, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Neddermann, M.; Backert, S. How many protein molecules are secreted by single Helicobacter pylori cells: Quantification of serine protease HtrA. Cell. Microbiol. 2019, 21, e13022. [Google Scholar] [CrossRef]
- Harrer, A.; Bucker, R.; Boehm, M.; Zarzecka, U.; Tegtmeyer, N.; Sticht, H.; Schulzke, J.D.; Backert, S. Campylobacter jejuni enters gut epithelial cells and impairs intestinal barrier function through cleavage of occludin by serine protease HtrA. Gut Pathog. 2019, 11, 4. [Google Scholar] [CrossRef]
- Hoy, B.; Geppert, T.; Boehm, M.; Reisen, F.; Plattner, P.; Gadermaier, G.; Sewald, N.; Ferreira, F.; Briza, P.; Schneider, G.; et al. Distinct roles of secreted HtrA proteases from gram-negative pathogens in cleaving the junctional protein and tumor suppressor E-cadherin. J. Biol. Chem. 2012, 287, 10115–10120. [Google Scholar] [CrossRef]
- Backert, S.; Boehm, M.; Wessler, S.; Tegtmeyer, N. Transmigration route of Campylobacter jejuni across polarized intestinal epithelial cells: Paracellular, transcellular or both? Cell Commun. Signal. 2013, 11, 72. [Google Scholar] [CrossRef]
- Eucker, T.P.; Konkel, M.E. The cooperative action of bacterial fibronectin-binding proteins and secreted proteins promote maximal Campylobacter jejuni invasion of host cells by stimulating membrane ruffling. Cell. Microbiol. 2012, 14, 226–238. [Google Scholar] [CrossRef]
- Monteville, M.R.; Konkel, M.E. Fibronectin-facilitated invasion of T84 eukaryotic cells by Campylobacter jejuni occurs preferentially at the basolateral cell surface. Infect. Immun. 2002, 70, 6665–6671. [Google Scholar] [CrossRef] [Green Version]
- van Alphen, L.B.; Bleumink-Pluym, N.M.; Rochat, K.D.; van Balkom, B.W.; Wosten, M.M.; van Putten, J.P. Active migration into the subcellular space precedes Campylobacter jejuni invasion of epithelial cells. Cell. Microbiol. 2008, 10, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, L.I.; Niewold, P.; van Putten, J.P. Basolateral invasion and trafficking of Campylobacter jejuni in polarized epithelial cells. PLoS ONE 2013, 8, e54759. [Google Scholar] [CrossRef] [PubMed]
- Krause-Gruszczynska, M.; Boehm, M.; Rohde, M.; Tegtmeyer, N.; Takahashi, S.; Buday, L.; Oyarzabal, O.A.; Backert, S. The signaling pathway of Campylobacter jejuni-induced Cdc42 activation: Role of fibronectin, integrin beta1, tyrosine kinases and guanine exchange factor Vav2. Cell Commun. Signal. 2011, 9, 32. [Google Scholar] [CrossRef]
- Sharafutdinov, I.; Tegtmeyer, N.; Musken, M.; Backert, S. Campylobacter jejuni Serine Protease HtrA Induces Paracellular Transmigration of Microbiota across Polarized Intestinal Epithelial Cells. Biomolecules 2022, 12, 521. [Google Scholar] [CrossRef] [PubMed]
- De Sa, F.D.L.; Butkevych, E.; Nattramilarasu, P.K.; Fromm, A.; Mousavi, S.; Moos, V.; Golz, J.C.; Stingl, K.; Kittler, S.; Seinige, D.; et al. Curcumin Mitigates Immune-Induced Epithelial Barrier Dysfunction by Campylobacter jejuni. Int. J. Mol. Sci. 2019, 20, 4830. [Google Scholar] [CrossRef]
- De Sa, F.D.L.; Heimesaat, M.M.; Bereswill, S.; Nattramilarasu, P.K.; Schulzke, J.D.; Bucker, R. Resveratrol Prevents Campylobacter jejuni-Induced Leaky gut by Restoring Occludin and Claudin-5 in the Paracellular Leak Pathway. Front. Pharmacol. 2021, 12, 640572. [Google Scholar] [CrossRef]
- Perna, A.M.; Rodrigues, T.; Schmidt, T.P.; Bohm, M.; Stutz, K.; Reker, D.; Pfeiffer, B.; Altmann, K.H.; Backert, S.; Wessler, S.; et al. Fragment-Based de novo Design Reveals a Small-Molecule Inhibitor of Helicobacter pylori HtrA. Angew. Chem. 2015, 54, 10244–10248. [Google Scholar] [CrossRef]
- Schmidt, T.P.; Perna, A.M.; Fugmann, T.; Bohm, M.; Hiss, J.; Haller, S.; Gotz, C.; Tegtmeyer, N.; Hoy, B.; Rau, T.T.; et al. Identification of E-cadherin signature motifs functioning as cleavage sites for Helicobacter pylori HtrA. Sci. Rep. 2016, 6, 23264. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Backert, S. Genetic and functional analyses reveal high conservation and crucial role of Helicobacter pylori serine protease HtrA. Helicobacter 2016, 21, 121. [Google Scholar]
- Wessler, S.; Schneider, G.; Backert, S. Bacterial serine protease HtrA as a promising new target for antimicrobial therapy? Cell Commun. Signal. 2017, 15, 4. [Google Scholar] [CrossRef]
- Benz, I.; Schmidt, M.A. Structures and functions of autotransporter proteins in microbial pathogens. Int. J. Med. Microbiol. 2011, 301, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmi, A.; Dorey, A.; Watson, E.; Jagatia, H.; Inglis, N.F.; Gundogdu, O.; Bajaj-Elliott, M.; Wren, B.W.; Smith, D.G.E.; Dorrell, N. The bile salt sodium taurocholate induces Campylobacter jejuni outer membrane vesicle production and increases OMV-associated proteolytic activity. Cell. Microbiol. 2018, 20, e12814. [Google Scholar] [CrossRef]
- Champion, O.L.; Gaunt, M.W.; Gundogdu, O.; Elmi, A.; Witney, A.A.; Hinds, J.; Dorrell, N.; Wren, B.W. Comparative phylogenomics of the food-borne pathogen Campylobacter jejuni reveals genetic markers predictive of infection source. Proc. Natl. Acad. Sci. USA 2005, 102, 16043–16048. [Google Scholar] [CrossRef]
- Zautner, A.E.; Herrmann, S.; Corso, J.; Tareen, A.M.; Alter, T.; Gross, U. Epidemiological association of different Campylobacter jejuni groups with metabolism-associated genetic markers. Appl. Environ. Microbiol. 2011, 77, 2359–2365. [Google Scholar] [CrossRef]
- Karlyshev, A.V.; Thacker, G.; Jones, M.A.; Clements, M.O.; Wren, B.W. Campylobacter jejuni gene cj0511 encodes a serine peptidase essential for colonisation. FEBS Open Bio 2014, 4, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Young, N.M.; Brisson, J.R.; Kelly, J.; Watson, D.C.; Tessier, L.; Lanthier, P.H.; Jarrell, H.C.; Cadotte, N.; St Michael, F.; Aberg, E.; et al. Structure of the N-linked glycan present on multiple glycoproteins in the Gram-negative bacterium, Campylobacter jejuni. J. Biol. Chem. 2002, 277, 42530–42539. [Google Scholar] [CrossRef] [PubMed]
- Sommerfield, A.G.; Darwin, A.J. Bacterial Carboxyl-Terminal Processing Proteases Play Critical Roles in the Cell Envelope and Beyond. J. Bacteriol. 2022, 204, e0062821. [Google Scholar] [CrossRef]
- Saoud, J.; Mani, T.; Faucher, S.P. The Tail-Specific Protease Is Important for Legionella pneumophila To Survive Thermal Stress in Water and inside Amoebae. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef]
- Nash, Z.M.; Cotter, P.A. Regulated, sequential processing by multiple proteases is required for proper maturation and release of Bordetella filamentous hemagglutinin. Mol. Microbiol. 2019, 112, 820–836. [Google Scholar] [CrossRef]
- Roy, R.; You, R.I.; Chang, C.H.; Yang, C.Y.; Lin, N.T. Carboxy-Terminal Processing Protease Controls Production of Outer Membrane Vesicles and Biofilm in Acinetobacter baumannii. Microorganisms 2021, 9, 1336. [Google Scholar] [CrossRef]
- Bandara, A.B.; Sriranganathan, N.; Schurig, G.G.; Boyle, S.M. Carboxyl-terminal protease regulates Brucella suis morphology in culture and persistence in macrophages and mice. J. Bacteriol. 2005, 187, 5767–5775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandara, A.B.; DeShazer, D.; Inzana, T.J.; Sriranganathan, N.; Schurig, G.G.; Boyle, S.M. A disruption of ctpA encoding carboxy-terminal protease attenuates Burkholderia mallei and induces partial protection in CD1 mice. Microb. Pathog. 2008, 45, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Darwin, A.J. The Pseudomonas aeruginosa periplasmic protease CtpA can affect systems that impact its ability to mount both acute and chronic infections. Infect. Immun. 2013, 81, 4561–4570. [Google Scholar] [CrossRef] [PubMed]
- Carroll, R.K.; Rivera, F.E.; Cavaco, C.K.; Johnson, G.M.; Martin, D.; Shaw, L.N. The lone S41 family C-terminal processing protease in Staphylococcus aureus is localized to the cell wall and contributes to virulence. Microbiology 2014, 160, 1737–1748. [Google Scholar] [CrossRef]
- Seal, B.S.; Hiett, K.L.; Kuntz, R.L.; Woolsey, R.; Schegg, K.M.; Ard, M.; Stintzi, A. Proteomic analyses of a robust versus a poor chicken gastrointestinal colonizing isolate of Campylobacter jejuni. J. Proteome Res. 2007, 6, 4582–4591. [Google Scholar] [CrossRef]
- Jowiya, W.; Brunner, K.; Abouelhadid, S.; Hussain, H.A.; Nair, S.P.; Sadiq, S.; Williams, L.K.; Trantham, E.K.; Stephenson, H.; Wren, B.W.; et al. Pancreatic amylase is an environmental signal for regulation of biofilm formation and host interaction in Campylobacter jejuni. Infect. Immun. 2015, 83, 4884–4895. [Google Scholar] [CrossRef]
- Gao, B.; Vorwerk, H.; Huber, C.; Lara-Tejero, M.; Mohr, J.; Goodman, A.L.; Eisenreich, W.; Galan, J.E.; Hofreuter, D. Metabolic and fitness determinants for in vitro growth and intestinal colonization of the bacterial pathogen Campylobacter jejuni. PLoS Biol. 2017, 15, e2001390. [Google Scholar] [CrossRef]
- Sauer, R.T.; Baker, T.A. AAA+ proteases: ATP-fueled machines of protein destruction. Annu. Rev. Biochem. 2011, 80, 587–612. [Google Scholar] [CrossRef]
- Kedzierska-Mieszkowska, S.; Arent, Z. AAA+ Molecular Chaperone ClpB in Leptospira interrogans: Its Role and Significance in Leptospiral Virulence and Pathogenesis of Leptospirosis. Int. J. Mol. Sci. 2020, 21, 6645. [Google Scholar] [CrossRef]
- Lourdault, K.; Cerqueira, G.M.; Wunder, E.A., Jr.; Picardeau, M. Inactivation of clpB in the pathogen Leptospira interrogans reduces virulence and resistance to stress conditions. Infect. Immun. 2011, 79, 3711–3717. [Google Scholar] [CrossRef]
- Ju, Y.; An, Q.; Zhang, Y.; Sun, K.; Bai, L.; Luo, Y. Recent advances in Clp protease modulation to address virulence, resistance and persistence of MRSA infection. Drug Discov. Today 2021, 26, 2190–2197. [Google Scholar] [CrossRef]
- Vegge, C.S.; Brondsted, L.; Ligowska-Marzeta, M.; Ingmer, H. Natural transformation of Campylobacter jejuni occurs beyond limits of growth. PLoS ONE 2012, 7, e45467. [Google Scholar] [CrossRef]
- Cohn, M.T.; Ingmer, H.; Mulholland, F.; Jorgensen, K.; Wells, J.M.; Brondsted, L. Contribution of conserved ATP-dependent proteases of Campylobacter jejuni to stress tolerance and virulence. Appl. Environ. Microbiol. 2007, 73, 7803–7813. [Google Scholar] [CrossRef]
- Kim, S.; Jeong, J.; Lee, H.; Lee, J.; Lee, S.; Ha, J.; Choi, Y.; Yoon, Y.; Choi, K.H. Kinetic Behavior of Campylobacter jejuni in Beef Tartare at Cold Temperatures and Transcriptomes Related to Its Survival. J. Food Prot. 2017, 80, 2127–2131. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.; Kim, S.; Ha, J.; Lee, J.; Choi, Y.; Oh, H.; Kim, Y.; Lee, Y.; Yoon, Y. The risk of aerotolerant Campylobacter jejuni strains in poultry meat distribution and storage. Microb. Pathog. 2019, 134, 103537. [Google Scholar] [CrossRef]
- Ghunaim, H.; Behnke, J.M.; Aigha, I.; Sharma, A.; Doiphode, S.H.; Deshmukh, A.; Abu-Madi, M.M. Analysis of resistance to antimicrobials and presence of virulence/stress response genes in Campylobacter isolates from patients with severe diarrhoea. PLoS ONE 2015, 10, e0119268. [Google Scholar] [CrossRef] [PubMed]
- Poli, V.F.; Thorsen, L.; Olesen, I.; Wik, M.T.; Jespersen, L. Differentiation of the virulence potential of Campylobacter jejuni strains by use of gene transcription analysis and a Caco-2 assay. Int. J. Food Microbiol. 2012, 155, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wurfel, S.D.R.; Prates, D.D.; Kleinubing, N.R.; Dalla Vecchia, J.; Vaniel, C.; Haubert, L.; Dellagostin, O.A.; da Silva, W.P. Comprehensive characterization reveals antimicrobial-resistant and potentially virulent Campylobacter isolates from poultry meat products in Southern Brazil. LWT-Food Sci. Technol. 2021, 149, 111831. [Google Scholar] [CrossRef]
- Thies, F.L.; Hartung, H.P.; Giegerich, G. Cloning and expression of the Campylobacter jejuni lon gene detected by RNA arbitrarily primed PCR. FEMS Microbiol. Lett. 1998, 165, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Langklotz, S.; Baumann, U.; Narberhaus, F. Structure and function of the bacterial AAA protease FtsH. Biochim. Biophys. Acta 2012, 1823, 40–48. [Google Scholar] [CrossRef]
- Wang, W.; Jiang, J.; Chen, H.; Zhang, Y.; Liu, Q. FtsH is required for protein secretion homeostasis and full bacterial virulence in Edwardsiella piscicida. Microb. Pathog. 2021, 161, 105194. [Google Scholar] [CrossRef]
- Van Hoang, K.; Stern, N.J.; Saxton, A.M.; Xu, F.; Zeng, X.; Lin, J. Prevalence, development, and molecular mechanisms of bacteriocin resistance in Campylobacter. Appl. Environ. Microbiol. 2011, 77, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.H.; Kim, I.J.; Wen, Y.; Park, N.Y.; Park, J.; Lee, K.W.; Koh, A.; Lee, J.H.; Koo, S.H.; Kim, K.S. Vibrio vulnificus Secretes an Insulin-degrading Enzyme That Promotes Bacterial Proliferation in vivo. J. Biol. Chem. 2015, 290, 18708–18720. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.G.; Lim, J.M.; Weinberg, M.V.; Wells, L.; Hoover, T.R. Direct analysis of the extracellular proteome from two strains of Helicobacter pylori. Proteomics 2007, 7, 2240–2245. [Google Scholar] [CrossRef] [PubMed]
- Mullaney, E.; Brown, P.A.; Smith, S.M.; Botting, C.H.; Yamaoka, Y.Y.; Terres, A.M.; Kelleher, D.P.; Windle, H.J. Proteomic and functional characterization of the outer membrane vesicles from the gastric pathogen Helicobacter pylori. Proteom. Clin. Appl. 2009, 3, 785–796. [Google Scholar] [CrossRef]
- Marques, M.S.; Costa, A.C.; Osorio, H.; Pinto, M.L.; Relvas, S.; Dinis-Ribeiro, M.; Carneiro, F.; Leite, M.; Figueiredo, C. Helicobacter pylori PqqE is a new virulence factor that cleaves junctional adhesion molecule A and disrupts gastric epithelial integrity. Gut Microbes 2021, 13, e1921928. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Gressmann, H.; Linz, B.; Ghai, R.; Pleissner, K.P.; Schlapbach, R.; Yamaoka, Y.; Kraft, C.; Suerbaum, S.; Meyer, T.F.; Achtman, M. Gain and loss of multiple genes during the evolution of Helicobacter pylori. PLoS Genet. 2005, 1, e43. [Google Scholar] [CrossRef]
- Peng, C.T.; Liu, L.; Li, C.C.; He, L.H.; Li, T.; Shen, Y.L.; Gao, C.; Wang, N.Y.; Xia, Y.; Zhu, Y.B.; et al. Structure-Function Relationship of Aminopeptidase P from Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 2385. [Google Scholar] [CrossRef] [PubMed]
- Feinbaum, R.L.; Urbach, J.M.; Liberati, N.T.; Djonovic, S.; Adonizio, A.; Carvunis, A.R.; Ausubel, F.M. Genome-wide identification of Pseudomonas aeruginosa virulence-related genes using a Caenorhabditis elegans infection model. PLoS Pathog. 2012, 8, e1002813. [Google Scholar] [CrossRef]
- Omar, M.N.; Abd Rahman, R.N.Z.R.; Noor, N.D.M.; Latip, W.; Knight, V.F.; Ali, M.S.M. Structure-Function and Industrial Relevance of Bacterial Aminopeptidase P. Catalysts 2021, 11, 1157. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Schmidt, A.M.; Mousavi, S.; Escher, U.; Tegtmeyer, N.; Wessler, S.; Gadermaier, G.; Briza, P.; Hofreuter, D.; Bereswill, S.; et al. Peptidase PepP is a novel virulence factor of Campylobacter jejuni contributing to murine campylobacteriosis. Gut Microbes 2020, 12, e1770017. [Google Scholar] [CrossRef] [PubMed]
- Lowther, W.T.; Matthews, B.W. Metalloaminopeptidases: Common functional themes in disparate structural surroundings. Chem. Rev. 2002, 102, 4581–4608. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.N.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef]
- Mansur, F.J.; Takahara, S.; Yamamoto, M.; Shimatani, M.; Minnatul Karim, M.; Noiri, Y.; Ebisu, S.; Azakami, H. Purification and characterization of hemolysin from periodontopathogenic bacterium Eikenella corrodens strain 1073. Biosci. Biotechnol. Biochem. 2017, 81, 1246–1253. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Evolutionary families of metallopeptidases. Methods Enzymol. 1995, 248, 183–228. [Google Scholar] [CrossRef] [PubMed]
- Chao, S.H.; Cheng, T.H.; Shaw, C.Y.; Lee, M.H.; Hsu, Y.H.; Tsai, Y.C. Characterization of a novel PepF-like oligopeptidase secreted by Bacillus amyloliquefaciens 23-7A. Appl. Environ. Microbiol. 2006, 72, 968–971. [Google Scholar] [CrossRef]
- Mierau, I.; Kunji, E.R.; Leenhouts, K.J.; Hellendoorn, M.A.; Haandrikman, A.J.; Poolman, B.; Konings, W.N.; Venema, G.; Kok, J. Multiple-peptidase mutants of Lactococcus lactis are severely impaired in their ability to grow in milk. J. Bacteriol. 1996, 178, 2794–2803. [Google Scholar] [CrossRef] [PubMed]
- Kunji, E.R.; Mierau, I.; Poolman, B.; Konings, W.N.; Venema, G.; Kok, J. Fate of peptides in peptidase mutants of Lactococcus lactis. Mol. Microbiol. 1996, 21, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.G.; Livny, J.; Dirita, V.J. High-throughput sequencing of Campylobacter jejuni insertion mutant libraries reveals mapA as a fitness factor for chicken colonization. J. Bacteriol. 2014, 196, 1958–1967. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Szczepanska, B.; Andrzejewska, M.; Spica, D.; Klawe, J.J. Prevalence and antimicrobial resistance of Campylobacter jejuni and Campylobacter coli isolated from children and environmental sources in urban and suburban areas. BMC Microbiol. 2017, 17, 80. [Google Scholar] [CrossRef]
- von Graevenitz, A. Revised nomenclature of Campylobacter laridis, Enterobacter intermedium, and “Flavobacterium branchiophila”. Int. J. Syst. Bacteriol. 1990, 40, 211. [Google Scholar] [CrossRef]
- Benjamin, J.; Leaper, S.; Owen, R.J.; Skirrow, M.B. Description of Campylobacter laridis, a New Species Comprising the Nalidixic-Acid Resistant Thermophilic Campylobacter (NARTC) Group. Curr. Microbiol. 1983, 8, 231–238. [Google Scholar] [CrossRef]
- Debruyne, L.; On, S.L.; De Brandt, E.; Vandamme, P. Novel Campylobacter lari-like bacteria from humans and molluscs: Description of Campylobacter peloridis sp. nov., Campylobacter lari subsp. concheus subsp. nov. and Campylobacter lari subsp. lari subsp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1126–1132. [Google Scholar] [CrossRef]
- Veron, M.; Chatelain, R. Taxonomic Study of Genus Campylobacter Sebald and Veron and Designation of Neotype Strain for Type Species, Campylobacter fetus (Smith and Taylor) Sebald and Veron. Int. J. Syst. Bacteriol. 1973, 23, 122–134. [Google Scholar] [CrossRef]
- Sandstedt, K.; Ursing, J. Description of Campylobacter upsaliensis sp-nov Previously Known as the CNW Group. Syst. Appl. Microbiol. 1991, 14, 39–45. [Google Scholar] [CrossRef]
- Boukerb, A.M.; Penny, C.; Serghine, J.; Walczak, C.; Cauchie, H.M.; Miller, W.G.; Losch, S.; Ragimbeau, C.; Mossong, J.; Megraud, F.; et al. Campylobacter armoricus sp. nov., a novel member of the Campylobacter lari group isolated from surface water and stools from humans with enteric infection. Int. J. Syst. Evol. Microbiol. 2019, 69, 3969–3979. [Google Scholar] [CrossRef]
- Roop, R.M.; Smibert, R.M.; Johnson, J.L.; Krieg, N.R. Designation of the Neotype Strain for Campylobacter sputorum (Prevot) Veron and Chatelain 1973. Int. J. Syst. Bacteriol. 1986, 36, 348. [Google Scholar] [CrossRef]
- Tanner, A.C.R.; Badger, S.; Lai, C.H.; Listgarten, M.A.; Visconti, R.A.; Socransky, S.S. Wolinella gen. nov., Wolinella succinogenes (Vibrio succinogenes Wolin et al.) comb. nov., and Description of Bacteroides gracilis sp. nov., Wolinella recta sp. nov., Campylobacter concisus sp. nov., and Eikenella corrodens from Humans with Periodontal Disease. Int. J. Syst. Bacteriol. 1981, 31, 432–445. [Google Scholar] [CrossRef]
- Cornelius, A.J.; Miller, W.G.; Lastovica, A.J.; On, S.L.W.; French, N.P.; Vandenberg, O.; Biggs, P.J. Complete Genome Sequence of Campylobacter concisus ATCC 33237(T) and Draft Genome Sequences for an Additional eight Well-Characterized C. concisus Strains. Genome Announc. 2017, 5, e00711-17. [Google Scholar] [CrossRef] [Green Version]
- Tanner, A.C.R.; Listgarten, M.A.; Ebersole, J.L. Wolinella curva sp nov-Vibrio succinogenes of Human Origin. Int. J. Syst. Bacteriol. 1984, 34, 275–282. [Google Scholar] [CrossRef]
- Vandamme, P.; Falsen, E.; Rossau, R.; Hoste, B.; Segers, P.; Tytgat, R.; De Ley, J. Revision of Campylobacter, Helicobacter, and Wolinella taxonomy: Emendation of generic descriptions and proposal of Arcobacter gen. nov. Int. J. Syst. Bacteriol. 1991, 41, 88–103. [Google Scholar] [CrossRef]
- Miller, W.G.; Yee, E. Complete Genome Sequence of Campylobacter gracilis ATCC 33236T. Genome Announc. 2015, 3, e01087-15. [Google Scholar] [CrossRef]
- Logan, J.M.; Burnens, A.; Linton, D.; Lawson, A.J.; Stanley, J. Campylobacter lanienae sp. nov., a new species isolated from workers in an abattoir. Int. J. Syst. Evol. Microbiol. 2000, 50, 865–872. [Google Scholar] [CrossRef]
- Etoh, Y.; Dewhirst, F.E.; Paster, B.J.; Yamamoto, A.; Goto, N. Campylobacter showae sp. nov., isolated from the human oral cavity. Int. J. Syst. Bacteriol. 1993, 43, 631–639. [Google Scholar] [CrossRef]
- Lawson, A.J.; Linton, D.; Stanley, J. 16S rRNA gene sequences of ‘Candidatus Campylobacter hominis’, a novel uncultivated species, are found in the gastrointestinal tract of healthy humans. Microbiology 1998, 144, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Kaur, T.; Singh, J.; Huffman, M.A.; Petrzelkova, K.J.; Taylor, N.S.; Xu, S.; Dewhirst, F.E.; Paster, B.J.; Debruyne, L.; Vandamme, P.; et al. Campylobacter troglodytis sp. nov., isolated from feces of human-habituated wild chimpanzees (Pan troglodytes schweinfurthii) in Tanzania. Appl. Environ. Microbiol. 2011, 77, 2366–2373. [Google Scholar] [CrossRef] [PubMed]
- Koziel, M.; Lucid, A.; Bullman, S.; Corcoran, G.D.; Lucey, B.; Sleator, R.D. Draft Genome Sequence of Campylobacter corcagiensis Strain CIT045T, a Representative of a Novel Campylobacter Species Isolated from Lion-Tailed Macaques (Macaca silenus). Genome Announc. 2014, 2, e00248-14. [Google Scholar] [CrossRef]
- Jensen, A.N.; Dalsgaard, A.; Baggesen, D.L.; Nielsen, E.M. The occurrence and characterization of Campylobacter jejuni and C. coli in organic pigs and their outdoor environment. Vet. Microbiol. 2006, 116, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lawson, G.H.K.; Leaver, J.L.; Pettigrew, G.W.; Rowland, A.C. Some features of Campylobacter sputorum subsp mucosalis subsp nov, nom-rev and their taxonomic significance. Int. J. Syst. Bacteriol. 1981, 31, 385–391. [Google Scholar] [CrossRef]
- Roop, R.M.; Smibert, R.M.; Johnson, J.L.; Krieg, N.R. Campylobacter mucosalis (Lawson, Leaver, Pettigrew, and Rowland 1981) comb-nov-emended description. Int. J. Syst. Bacteriol. 1985, 35, 189–192. [Google Scholar] [CrossRef]
- Gebhart, C.J.; Edmonds, P.; Ward, G.E.; Kurtz, H.J.; Brenner, D.J. “Campylobacter hyointestinalis” sp. nov.: A new species of Campylobacter found in the intestines of pigs and other animals. J. Clin. Microbiol. 1985, 21, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.S.; Orcutt, M.; Little, R.B. Vibrios (Vibrio jejuni, N. Sp.) Associated with Intestinal Disorders of Cows and Calves. J. Exp. Med. 1931, 53, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.; Holmes, B.; Steigerwalt, A.G.; Lawson, P.A.; Thorne, P.; Byrer, D.E.; Ross, H.M.; Xerry, J.; Thompson, P.M.; Collins, M.D. Campylobacter insulaenigrae sp. nov., isolated from marine mammals. Int. J. Syst. Evol. Microbiol. 2004, 54, 2369–2373. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.J.; Miller, W.G.; Leger, J.S.; Chapman, M.H.; Timmerman, A.J.; Duim, B.; Foster, G.; Wagenaar, J.A. Campylobacter pinnipediorum sp. nov., isolated from pinnipeds, comprising Campylobacter pinnipediorum subsp. pinnipediorum subsp. nov. and Campylobacter pinnipediorum subsp. caledonicus subsp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 1961–1968. [Google Scholar] [CrossRef]
- Rossi, M.; Debruyne, L.; Zanoni, R.G.; Manfreda, G.; Revez, J.; Vandamme, P. Campylobacter avium sp. nov., a hippurate-positive species isolated from poultry. Int. J. Syst. Evol. Microbiol. 2009, 59, 2364–2369. [Google Scholar] [CrossRef]
- Van, T.T.H.; Elshagmani, E.; Gor, M.C.; Scott, P.C.; Moore, R.J. Campylobacter hepaticus sp. nov., isolated from chickens with spotty liver disease. Int. J. Syst. Evol. Microbiol. 2016, 66, 4518–4524. [Google Scholar] [CrossRef]
- Stanley, J.; Burnens, A.P.; Linton, D.; On, S.L.; Costas, M.; Owen, R.J. Campylobacter helveticus sp. nov., a new thermophilic species from domestic animals: Characterization, and cloning of a species-specific DNA probe. J. Gen. Microbiol. 1992, 138, 2293–2303. [Google Scholar] [CrossRef]
- Miller, W.G.; Yee, E.; Bono, J.L. Complete Genome Sequence of the Campylobacter helveticus Type Strain ATCC 51209. Genome Announc. 2017, 5, e00398-17. [Google Scholar] [CrossRef]
- Parisi, A.; Chiara, M.; Caffara, M.; Mion, D.; Miller, W.G.; Caruso, M.; Manzari, C.; Florio, D.; Capozzi, L.; D’Erchia, A.M.; et al. Campylobacter vulpis sp. nov. isolated from wild red foxes. Syst. Appl. Microbiol. 2021, 44, 126204. [Google Scholar] [CrossRef]
- Zanoni, R.G.; Debruyne, L.; Rossi, M.; Revez, J.; Vandamme, P. Campylobacter cuniculorum sp. nov., from rabbits. Int. J. Syst. Evol. Microbiol. 2009, 59, 1666–1671. [Google Scholar] [CrossRef]
- Aydin, F.; Abay, S.; Kayman, T.; Karakaya, E.; Mustak, H.K.; Mustak, I.B.; Bilgen, N.; Goncuoglu, M.; Duzler, A.; Guran, O.; et al. Campylobacter anatolicus sp. nov., a novel member of the genus Campylobacter isolated from feces of Anatolian Ground Squirrel (Spermophilus xanthoprymnus) in Turkey. Syst. Appl. Microbiol. 2021, 44, 126265. [Google Scholar] [CrossRef]
- Du, J.; Luo, J.; Huang, J.; Wang, C.; Li, M.; Wang, B.; Wang, B.; Chang, H.; Ji, J.; Sen, K.; et al. Emergence of Genetic Diversity and Multi-Drug Resistant Campylobacter jejuni From Wild Birds in Beijing, China. Front. Microbiol. 2019, 10, 2433. [Google Scholar] [CrossRef]
- Debruyne, L.; Broman, T.; Bergstrom, S.; Olsen, B.; On, S.L.W.; Vandamme, P. Campylobacter volucris sp. nov., isolated from black-headed gulls (Larus ridibundus). Int. J. Syst. Evol. Microbiol. 2010, 60, 1870–1875. [Google Scholar] [CrossRef]
- Debruyne, L.; Broman, T.; Bergstrom, S.; Olsen, B.; On, S.L.W.; Vandamme, P. Campylobacter subantarcticus sp nov., isolated from birds in the sub-Antarctic region. Int. J. Syst. Evol. Microbiol. 2010, 60, 815–819. [Google Scholar] [CrossRef]
- Bloomfield, S.; Wilkinson, D.; Rogers, L.; Biggs, P.; French, N.; Mohan, V.; Savoian, M.; Venter, P.; Midwinter, A. Campylobacter novaezeelandiae sp. nov., isolated from birds and water in New Zealand. Int. J. Syst. Evol. Microbiol. 2020, 70, 3775–3784. [Google Scholar] [CrossRef]
- Bryant, E.; Shen, Z.; Mannion, A.; Patterson, M.; Buczek, J.; Fox, J.G. Campylobacter taeniopygiae sp. nov., Campylobacter aviculae sp. nov., and Campylobacter estrildidarum sp. nov., Novel Species Isolated from Laboratory-Maintained Zebra Finches. Avian Dis. 2020, 64, 457–466. [Google Scholar] [CrossRef]
- Caceres, A.; Munoz, I.; Iraola, G.; Diaz-Viraque, F.; Collado, L. Campylobacter ornithocola sp. nov., a novel member of the Campylobacter lari group isolated from wild bird faecal samples. Int. J. Syst. Evol. Microbiol. 2017, 67, 1643–1649. [Google Scholar] [CrossRef]
- Harvey, S.; Greenwood, J.R. Isolation of Campylobacter fetus from a pet turtle. J. Clin. Microbiol. 1985, 21, 260–261. [Google Scholar] [CrossRef]
- Gilbert, M.J.; Kik, M.; Timmerman, A.J.; Severs, T.T.; Kusters, J.G.; Duim, B.; Wagenaar, J.A. Occurrence, diversity, and host association of intestinal Campylobacter, Arcobacter, and Helicobacter in reptiles. PLoS ONE 2014, 9, e101599. [Google Scholar] [CrossRef] [Green Version]
- Piccirillo, A.; Niero, G.; Calleros, L.; Perez, R.; Naya, H.; Iraola, G. Campylobacter geochelonis sp. nov. isolated from the western Hermann’s tortoise (Testudo hermanni hermanni). Int. J. Syst. Evol. Microbiol. 2016, 66, 3468–3476. [Google Scholar] [CrossRef]
- Gilbert, M.J.; Kik, M.; Miller, W.G.; Duim, B.; Wagenaar, J.A. Campylobacter iguaniorum sp. nov., isolated from reptiles. Int. J. Syst. Evol. Microbiol. 2015, 65, 975–982. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Protease Locus Cj | HtrA 1228c | 0511 | 1365c | ClpP 0192c | Lon 1073c | FtsH 1116c | PepP 0653c | PepF 1099 | PqqE 0805 | C26 1417c |
|---|---|---|---|---|---|---|---|---|---|---|
| Predicted cellular location | P | CM, P | OM, EC | C | C | CM | C | C | C, CM, P | C |
| C. jejuni | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. armoricus | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. vulpis | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. fetus | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. upsaliensis | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. coli | Y | Y | Y | Y | Y | Y | Y | Y | Y | Y |
| C. troglodytis | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. lari | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. cuniculorum | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. helveticus | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. insulaenigrae | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. subantarcticus | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. sputorum | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. novaezeelandiae | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| C. peloridis | Y | Y | Y | Y | Y | Y | Y | Y | Y | - |
| C. lanienae | Y | Y | Y | Y | Y | Y | Y | Y | Y | - |
| C. hyointestinalis | Y | Y | Y | Y | Y | Y | Y | Y | Y | - |
| C. volucris | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. concisus | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. corcagiensis | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. curvus | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. gracilis | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. pinnipediorum | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. rectus | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. showae | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. mucosalis | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. avium | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. geochelonis | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. hepaticus | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. hominis | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. iguaniorum | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. taeniopygiae | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. anatolicus | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| C. ornithocola | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| H. pylori | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| H. hepaticus | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| W. succinogenes | Y | Y | - | Y | Y | Y | Y | Y | Y | Y |
| S. arsenophilum | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
| N. profundicola | Y | Y | - | Y | Y | Y | Y | Y | Y | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linz, B.; Sharafutdinov, I.; Tegtmeyer, N.; Backert, S. Evolution and Role of Proteases in Campylobacter jejuni Lifestyle and Pathogenesis. Biomolecules 2023, 13, 323. https://doi.org/10.3390/biom13020323
Linz B, Sharafutdinov I, Tegtmeyer N, Backert S. Evolution and Role of Proteases in Campylobacter jejuni Lifestyle and Pathogenesis. Biomolecules. 2023; 13(2):323. https://doi.org/10.3390/biom13020323
Chicago/Turabian StyleLinz, Bodo, Irshad Sharafutdinov, Nicole Tegtmeyer, and Steffen Backert. 2023. "Evolution and Role of Proteases in Campylobacter jejuni Lifestyle and Pathogenesis" Biomolecules 13, no. 2: 323. https://doi.org/10.3390/biom13020323