
Effects of Bellamya purificata Cultivation at Different Stocking Densities on the Dynamics and Assembly of Bacterial Communities in Sediment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Experiment
2.2. Sample Collection
2.3. Sediment Properties Determination
2.4. 16S rRNA Gene Amplification and Sequencing
2.5. Data Processing
2.6. Statistical Analysis
3. Results
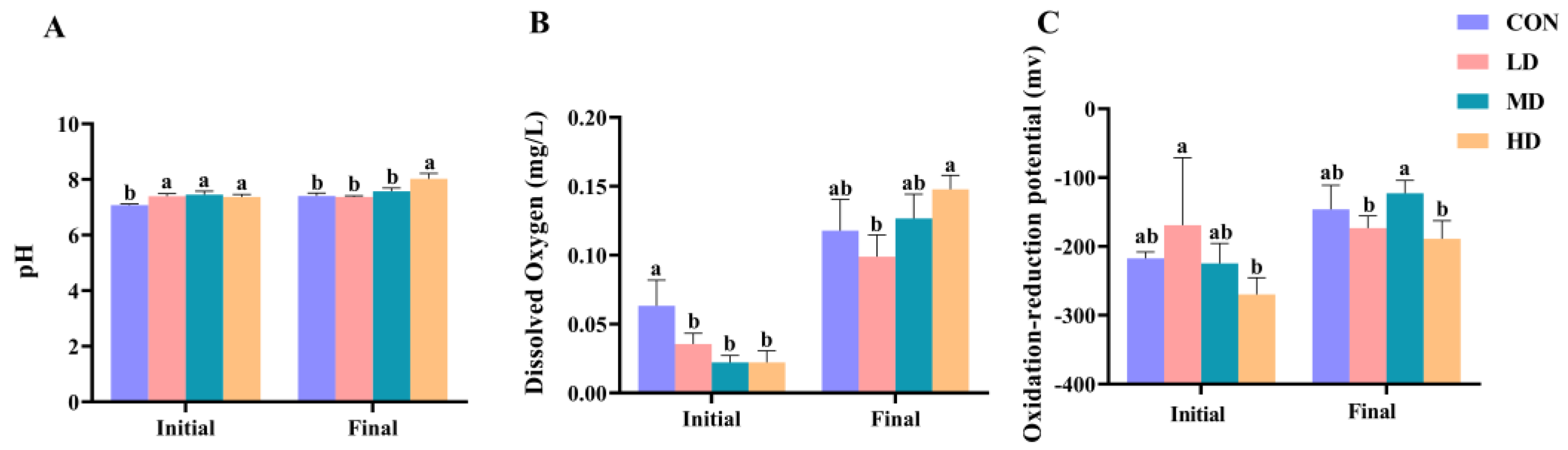
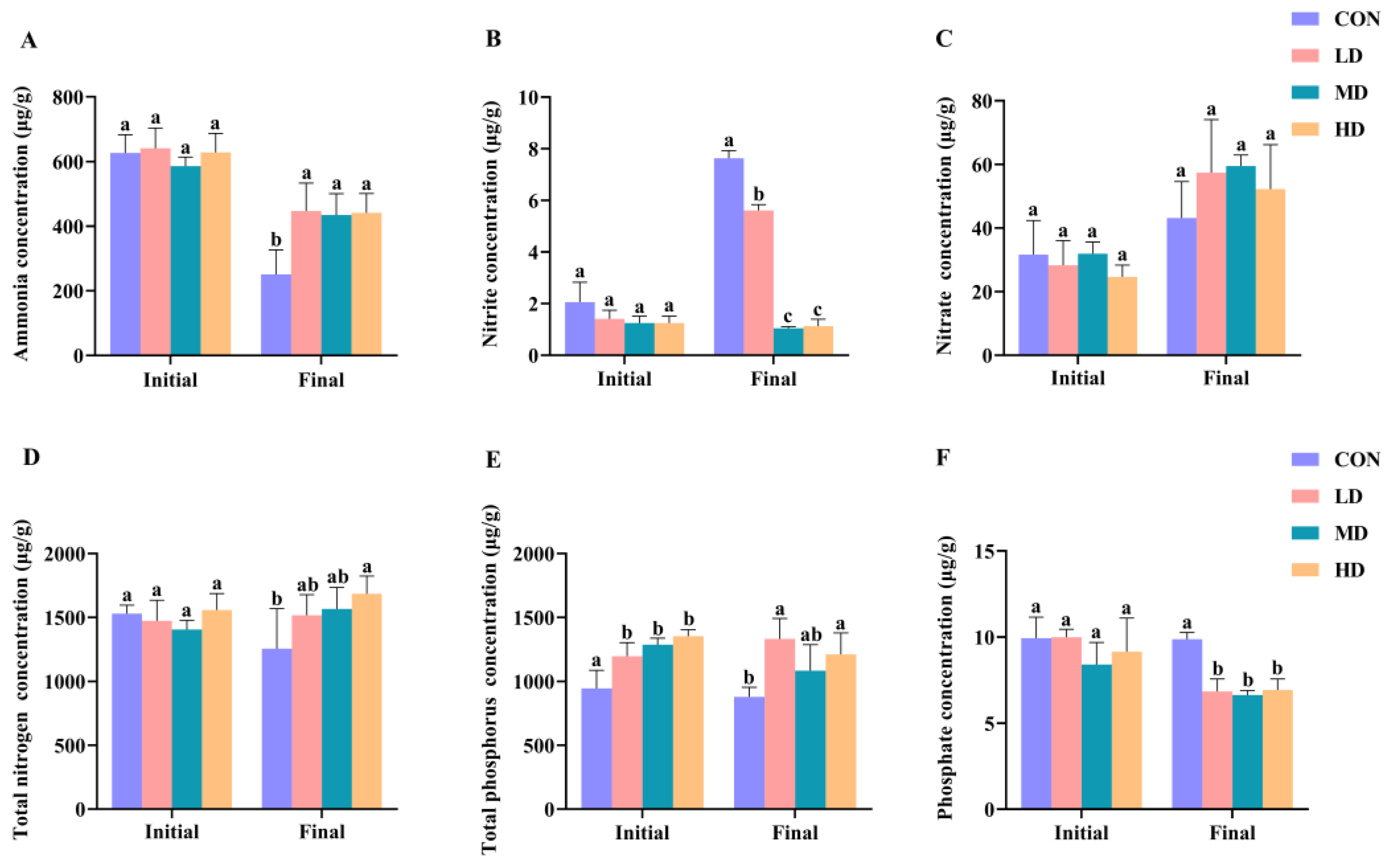
3.1. Sediment Characteristics
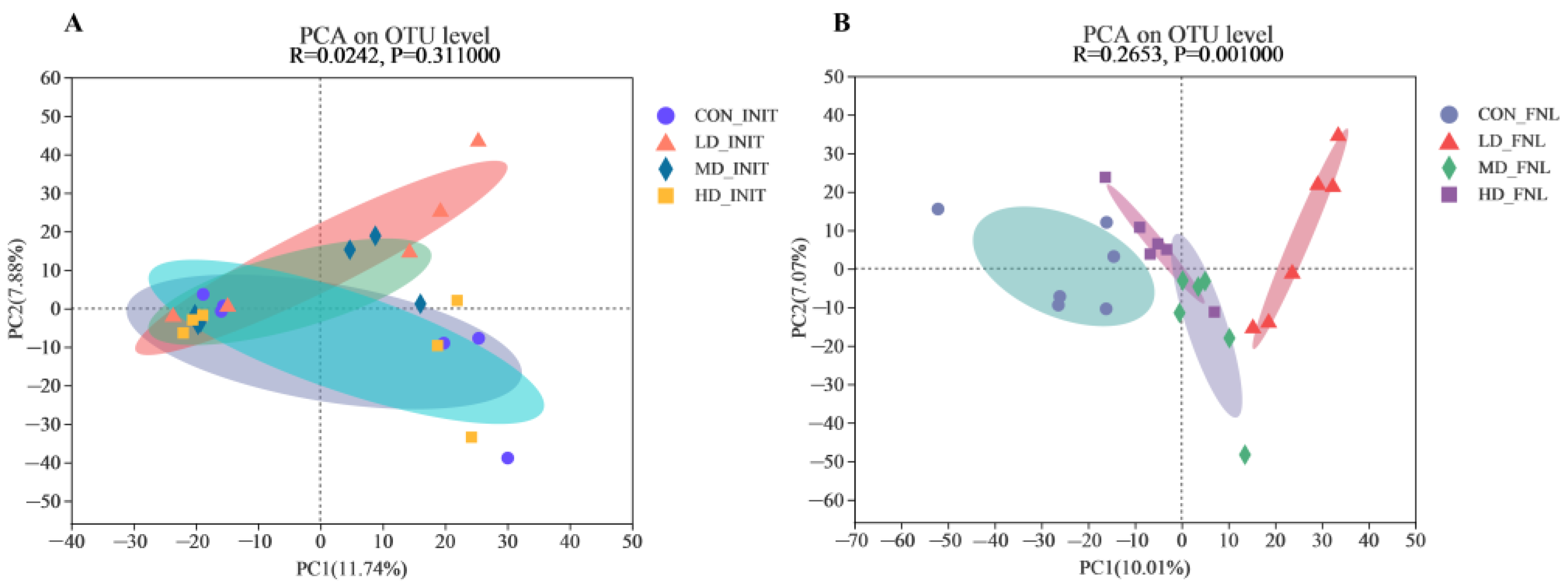
3.2. Bacterial Community Structure
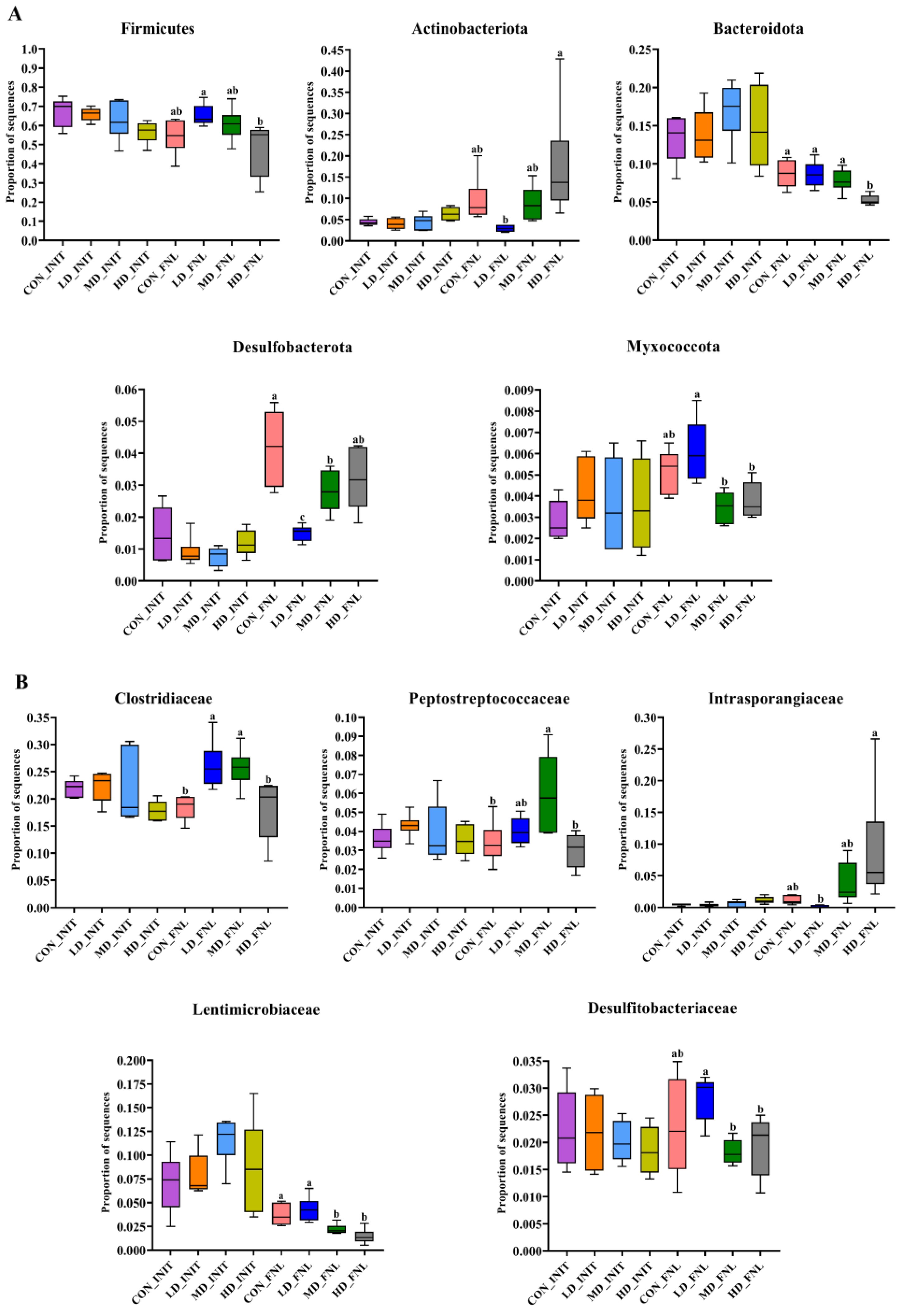
3.3. Bacterial Community Compositions
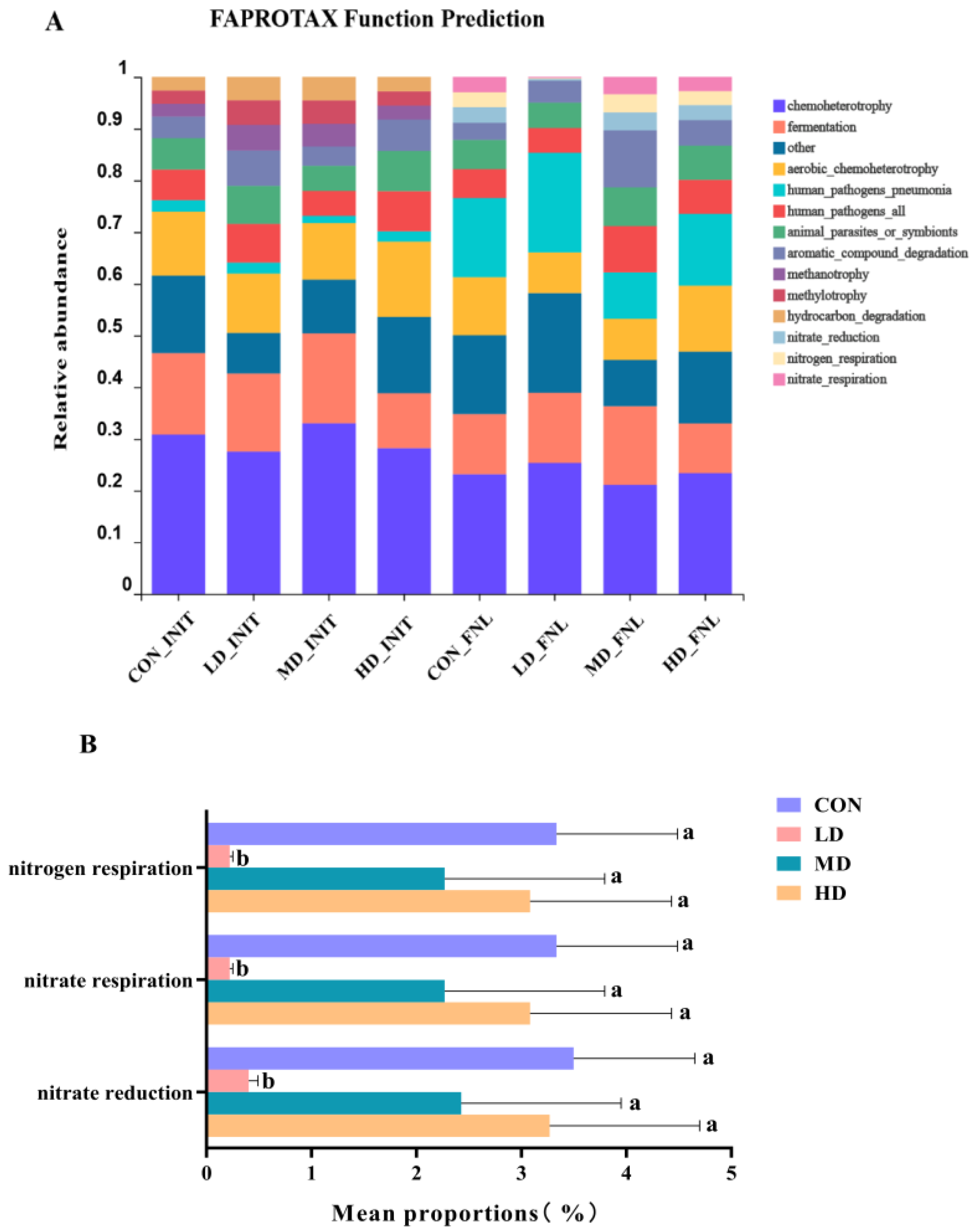
3.4. Bacterial Functional Prediction
3.5. Effect of Environmental Factors on Bacterial Abundance
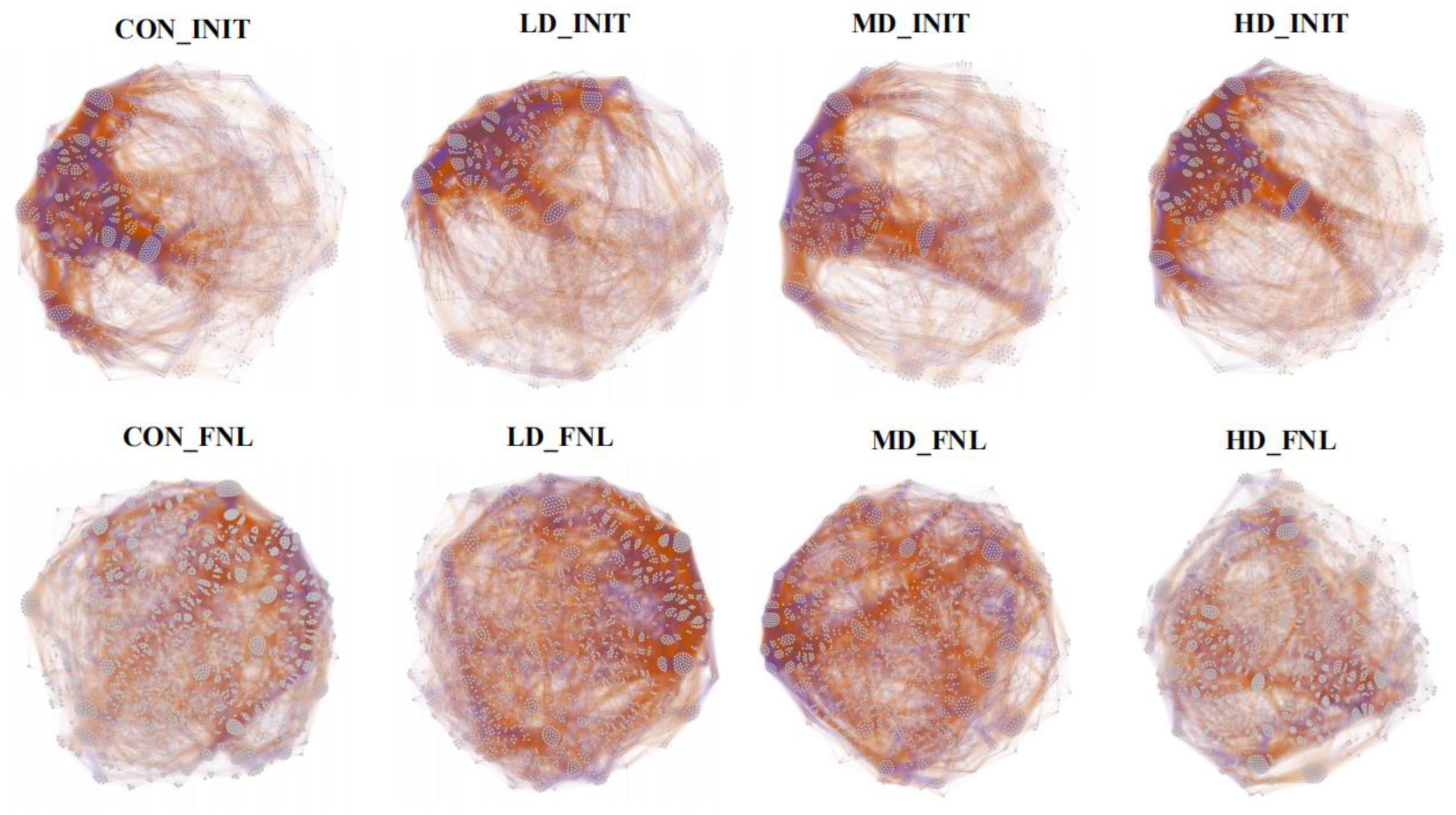
3.6. Co-Occurrence Patterns of Bacterial Taxa during the Culture Cycle
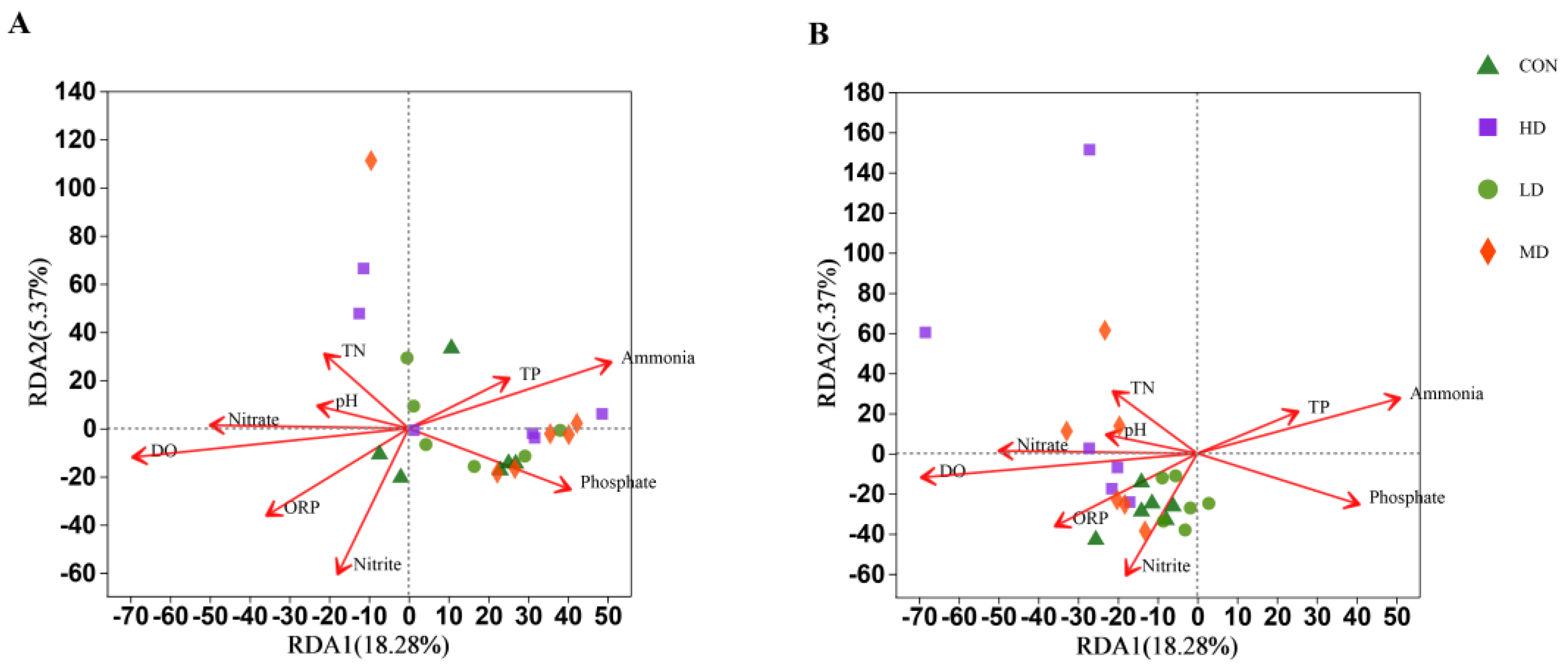
3.7. Mechanisms Involved in Regulating Bacterial Community Structure
4. Discussion
4.1. Bacterial Community Compositions and Functions Affected by B. purificata Cultivation
4.2. Bacterial Co-Occurrence Networks Affected by B. purificata Cultivation
4.3. Environmental Factors and Their Correlations with Bacterial Community Affected by B. purificata Cultivation
4.4. Bacterial Community Assembly Affected by B. purificata Cultivation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costello, C.; Cao, L.; Gelcich, S.; Cisneros-Mata, M.A.; Free, C.M.; Froehlich, H.E.; Golden, C.D.; Ishimura, G.; Maier, J.; Macadam-Somer, I.; et al. The future of food from the sea. Nature 2020, 588, 95–100. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Bureau of Fisheries Ministry of Agriculture and Rural Affairs; National Fisheries Technology Extension Center; China Society of Fisheries. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2021.
- Kalcheva, H.; Nikolova, L.; Terziyski, D.; Stoeva, A.; Kalchev, R. Bacterioplankton dynamics and its relations to productivity in carp fish ponds. Bulg. J. Agric. Sci. 2008, 14, 186–194. [Google Scholar]
- Ferreira, J.G.; Hawkins, A.; Monteiro, P.; Moore, H.; Service, M.; Pascoe, P.L.; Ramos, L.; Sequeira, A. Integrated assessment of ecosystem-scale carrying capacity in shellfish growing areas. Aquaculture 2008, 275, 138–151. [Google Scholar] [CrossRef]
- Yang, R.; Kaichang, W.U.; Gang, Y.U.; Wen, W.; Chen, X.; Zhao, W.; Le, Y.E. Effects of Culture Model on Growth and Main Environmental Factors in Snail Babylonia arelata. Fish. Sci. 2019, 38, 610–615. [Google Scholar] [CrossRef]
- Cao, Z.; Jiang, X. The influence of environmental factors on Bellamya purificata. Shanghai Fish. Univ. 1998, 7, 200–205. [Google Scholar]
- Zhu, J.; Lu, K.; Liu, X. Can the freshwater snail Bellamya aeruginosa (Mollusca) affect phytoplankton community and water quality? Hydrobiologia 2013, 707, 147–157. [Google Scholar] [CrossRef]
- Yanqing, Y.; Zhang, J.; Liu, L.; Wang, G.; Chen, M.; Zhang, Y.; Tang, X. Experimental study on phosphorus release from sediment with fresh-water snail (Bellamya aeruginosa) bioturbation in eutrophic lakes. J. Soils Sediments 2020, 20, 2526–2536. [Google Scholar] [CrossRef]
- Hou, Y.; Li, B.; Luo, J.; Zhang, C.; He, J.; Zhu, J. Effect of Bellamya purificata on organic matter degradation in surface sediment as revealed by amino acids. Inter Res. Sci. Cent. 2021, 13, 1–12. [Google Scholar] [CrossRef]
- Cheng, Y.F.; Zhou, Q.L.; Xie, J.; Ge, X.P.; Zhu, W.Y.; Liu, B. Microbial community analysis in crab ponds by denaturing gradient gel electrophoresis. World J. Microbiol. Biotechnol. 2010, 26, 825–831. [Google Scholar] [CrossRef]
- Zhao, Z.; Jiang, J.; Pan, Y.; Dong, Y.; Zhou, Z. Temporal dynamics of bacterial communities in the water and sediments of sea cucumber (Apostichopus japonicus) culture ponds. Aquaculture 2020, 528, 735498. [Google Scholar] [CrossRef]
- Guan, X.Y.; Wang, B.; Duan, P.; Tian, J.S.; Dong, Y.; Jiang, J.W.; Sun, B.; Zhou, Z.C. The dynamics of bacterial community in a polyculture aquaculture system of Penaeus chinensis, Rhopilema esculenta and Sinonovacula constricta. Aquac. Res. 2020, 51, 1789–1800. [Google Scholar] [CrossRef]
- Nogaro, G.; Mermillod-Blondin, F.; Valett, M.H.; Francois-Carcaillet, F.; Gaudet, J.-P.; Lafont, M.; Gibert, J. Ecosystem engineering at the sediment-water interface: Bioturbation and consumer-substrate interaction. Oecologia 2009, 161, 125–138. [Google Scholar] [CrossRef]
- Gilbert, F.; Bonin, P.; Stora, G. Effect of bioturbation on denitrification in a marine sediment from the West Mediterranean littoral. Hydrobiologia 1995, 304, 49–58. [Google Scholar] [CrossRef]
- Bonaglia, S.; Bartoli, M.; Gunnarsson, J.S.; Rahm, L.; Raymond, C.; Svensson, O.; Yekta, S.S.; Bruchert, V. Effect of reoxygenation and Marenzelleria spp. bioturbation on Baltic Sea sediment metabolism. Mar. Ecol. Prog. Ser. 2013, 482, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Jie, L.; Zheng, X.L.; Lei, C. Notice of Retraction<BR>Estimation of nitrogen and phosphorus release in sediment based on dynamic conditions. In Proceedings of the International Conference on Geoscience & Remote Sensing, Qingdao, China, 28–31 August 2010. [Google Scholar]
- Huang, X.F.; Chen, W.M.; Cai, Q.M. Survey, Observation and Analysis of Lake Ecology; Standard Press of China: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Xing, D.; Huo, T.; Wu, H.; Liu, J.; Liu, C.; Lei, Y. Simultaneous digestion for determination of total phosphorus and total nitrogen in sae water. J. Dalian Fish. Univ. 2006, 21, 219–225. [Google Scholar] [CrossRef]
- Qin, B.; Zhu, G.; Luo, L.; Gao, G.; Gu, B. Estimation of internal nutrient release in large shallow Lake Taihu, China. Sci. China Ser. D Earth Sci. 2006, 49, 38–50. [Google Scholar] [CrossRef]
- Zheng, X.; Tang, J.; Zhang, C.; Qin, J.; Wang, Y. Bacterial composition, abundance and diversity in fish polyculture and mussel-fish integrated cultured ponds in China. Aquac. Res. 2017, 48, 3950–3961. [Google Scholar] [CrossRef]
- Nicholaus, R.; Lukwambe, B.; Zhao, L.; Yang, W.; Zhu, J.; Zheng, Z. Bioturbation of blood clam Tegillarca granosa on benthic nutrient fluxes and microbial community in an aquaculture wastewater treatment system. Int. Biodeterior. Biodegrad. 2019, 142, 73–82. [Google Scholar] [CrossRef]
- Sun, F.; Zheng, Z.-M.; Lu, K.-H.; Zhai, H.-J.; Shao, L.-L. Effects of Snail Bellamya Aeruginosa Bioturbation on Microbial Community in The Sediment of Algal Bloom Water. Oceanol. Limnol. Sin. 2012, 43, 359–361. [Google Scholar] [CrossRef]
- Qin, Q.-L.; Zhang, X.-Y.; Wang, X.-M.; Liu, G.-M.; Chen, X.-L.; Xie, B.-B.; Dang, H.-Y.; Zhou, B.-C.; Yu, J.; Zhang, Y.-Z. The complete genome of Zunongwangia profunda SM-A87 reveals its adaptation to the deep-sea environment and ecological role in sedimentary organic nitrogen degradation. BMC Genom. 2010, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Mayer, R.E.; Sofill-Mas, S.; Egle, L.; Reischer, G.H.; Schade, M.; Fernandez-Cassi, X.; Fuchs, W.; Mach, R.L.; Lindner, G.; Kirschner, A.; et al. Occurrence of human-associated Bacteroidetes genetic source tracking markers in raw and treated wastewater of municipal and domestic origin and comparison to standard and alternative indicators of faecal pollution. Water Res. 2016, 90, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, A.T.; Stach, J.E.M.; Ward, A.C.; Goodfellow, M. Marine actinobacteria: Perspectives, challenges, future directions. Antonie Leeuwenhoek 2005, 87, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Bonin, P. Anaerobic nitrate reduction to ammonium in two strains isolated from coastal marine sediment: A dissimilatory pathway. Fems Microbiol. Ecol. 1996, 19, 27–38. [Google Scholar] [CrossRef]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration--genes, enzymes and environmental distribution. J. Biotechnol. 2011, 155, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Zhu, G.B.; Singh, B.; Zhu, Y.G. Anaerobic ammonium oxidation in agricultural soils-synthesis and prospective. Environ. Pollut. 2019, 244, 127–134. [Google Scholar] [CrossRef]
- Putz, M.; Schleusner, P.; Rütting, T.; Hallin, S. Relative abundance of denitrifying and DNRA bacteria and their activity determine nitrogen retention or loss in agricultural soil. Soil Biol. Biochem. 2018, 123, 97–104. [Google Scholar] [CrossRef]
- Laverock, B.; Tait, K.; Gilbert, J.A.; Osborn, A.M.; Widdicombe, S. Impacts of bioturbation on temporal variation in bacterial and archaeal nitrogen-cycling gene abundance in coastal sediments. Environ. Microbiol. Rep. 2014, 6, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Lukwambe, B.; Yang, W.; Zheng, Y.; Nicholaus, R.; Zhu, J.; Zheng, Z. Bioturbation by the razor clam (Sinonovacula constricta) on the microbial community and enzymatic activities in the sediment of an ecological aquaculture wastewater treatment system. Sci. Total Environ. 2018, 643, 1098–1107. [Google Scholar] [CrossRef]
- Fang, J.; Jiang, W.; Meng, S.; He, W.; Wang, G.; Guo, E.; Yan, Y. Polychaete Bioturbation Alters the Taxonomic Structure, Co-occurrence Network, and Functional Groups of Bacterial Communities in the Intertidal Flat. Microb. Ecol. 2022, 1–15. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Ma, B.; Wang, Y.; Ye, S.; Liu, S.; Stirling, E.; Gilbert, J.A.; Faust, K.; Knight, R.; Jansson, J.K.; Cardona, C.; et al. Earth microbial co-occurrence network reveals interconnection pattern across microbiomes. Microbiome 2020, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Li, B.; Xu, G.; Li, D.; Zhang, C.; Jia, R.; Li, Q.; Zhu, J. Dynamic and Assembly of Benthic Bacterial Community in an Industrial-Scale In-Pond Raceway Recirculating Culture System. Front. Microbiol. 2021, 12, 797817. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Jia, R.; Li, B.; Zhu, J. Apex Predators Enhance Environmental Adaptation but Reduce Community Stability of Bacterioplankton in Crustacean Aquaculture Ponds. Int. J. Mol. Sci. 2022, 23, 10785. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Xu, H.; Zhao, D.; Zeng, J.; Wu, Q.L. Aquaculture drives distinct patterns of planktonic and sedimentary bacterial communities: Insights into co-occurrence pattern and assembly processes. Environ. Microbiol. 2022, 24, 4079–4093. [Google Scholar] [CrossRef]
- Hunt, D.E.; Ward, C.S. A network-based approach to disturbance transmission through microbial interactions. Front. Microbiol. 2015, 6, 1182. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.M.; Henry, L.M.; Cornwallis, C.K.; Kiers, E.T.; West, S.A. The evolution of host-symbiont dependence. Nat. Commun. 2017, 8, 15973. [Google Scholar] [CrossRef] [Green Version]
- Decaestecker, E.; Gaba, S.; Raeymaekers, J.A.M.; Stoks, R.; Van Kerckhoven, L.; Ebert, D.; De Meester, L. Host-parasite ‘Red Queen’ dynamics archived in pond sediment. Nature 2007, 450, 870–873. [Google Scholar] [CrossRef]
- Paulina, F.; Nadia, D.; Ana, T.; Leonardo, L.; Fernando, U. The microbial food web structure of a hypertrophic warm-temperate shallow lake, as affected by contrasting zooplankton assemblages. Hydrobiologia 2013, 714, 115–130. [Google Scholar] [CrossRef]
- Rath, K.M.; Fierer, N.; Murphy, D.V.; Rousk, J. Linking bacterial community composition to soil salinity along environmental gradients. ISME J. 2019, 13, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Harris, J.K.; Wilcox, J.; Spear, J.R.; Miller, S.R.; Bebout, B.M.; Maresca, J.A.; Bryant, D.A.; Sogin, M.L.; Pace, N.R. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl. Environ. Microbiol. 2006, 72, 3685–3695. [Google Scholar] [CrossRef] [Green Version]
- Webster, G.; Parkes, R.J.; Cragg, B.A.; Newberry, C.J.; Weightman, A.J.; Fry, J.C. Prokaryotic community composition and biogeochemical processes in deep subseafloor sediments from the Peru Margin. Fems Microbiol. Ecol. 2006, 58, 65–85. [Google Scholar] [CrossRef] [Green Version]
- Marmen, S.; Fadeev, E.; Al Ashhab, A.; Benet-Perelberg, A.; Naor, A.; Patil, H.J.; Cytryn, E.; Viner-Mozzini, Y.; Sukenik, A.; Lalzar, M.; et al. Seasonal Dynamics Are the Major Driver of Microbial Diversity and Composition in Intensive Freshwater Aquaculture. Front. Microbiol. 2021, 12, 679743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhu, H.; Wang, J.; Lin, X.; Wang, J.; Huang, Y.; Li, B.; Mou, H.; Ma, X.; Wang, R. Monitoring Bacterial Community Dynamics in Abalone (Haliotis discus hannai) and the Correlations Associated with Aquatic Diseases. Water 2022, 14, 1769. [Google Scholar] [CrossRef]
- Urakawa, H.; Yoshida, T.; Nishimura, M.; Ohwada, K. Characterization of depth-related population variation in microbial communities of a coastal marine sediment using 16S rDNA-based approaches and quinone profiling. Environ. Microbiol. 2000, 2, 542–554. [Google Scholar] [CrossRef]
- Jansen, S.; Walpersdorf, E.; Werner, U.; Billerbeck, M.; Bottcher, M.E.; de Beer, D. Functioning of intertidal flats inferred from temporal and spatial dynamics of O2, H2S and pH in their surface sediment. Ocean. Dyn. 2009, 59, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Laverock, B.; Smith, C.J.; Tait, K.; Osborn, A.M.; Widdicombe, S.; Gilbert, J.A. Bioturbating shrimp alter the structure and diversity of bacterial communities in coastal marine sediments. ISME J. 2010, 4, 1531–1544. [Google Scholar] [CrossRef] [Green Version]
- Boyd, C.E.; Pippopinyo, S. Factors affecting respiration in dry pond bottom soils. Aquaculture 1994, 120, 283–293. [Google Scholar] [CrossRef]
- Aller, R. Transport and reactions in the bioirrigated zone. In The Benthic Boundary Layer: Transport Processes and Biogeochemistry; Boudreau, B.P., Jørgensen, B.B., Eds.; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Grenz, C.; Denis, L.; Boucher, G.; Chauvaud, L.; Clavier, J.; Fichez, R.; Pringault, O. Spatial variability in Sediment Oxygen Consumption under winter conditions in a lagoonal system in New Caledonia (South Pacific). J. Exp. Mar. Biol. Ecol. 2003, 285, 33–47. [Google Scholar] [CrossRef]
- Li, Y.; Hua, X.; Mao, D.; Zheng, F.; Dong, D.; Liang, D. Effects of Tubificids and Their Bioturbation on pH and Dissolved Oxygen in the Microenvironment of Surface Sediment. J. Jilin Univ. Sci. Ed. 2015, 53, 1334–1340. [Google Scholar] [CrossRef]
- Foshtomi, M.Y.; Braeckman, U.; Derycke, S.; Sapp, M.; Van Gansbeke, D.; Sabbe, K.; Willems, A.; Vincx, M.; Vanaverbeke, J. The link between microbial diversity and nitrogen cycling in marine sediments is modulated by macrofaunal bioturbation. PLoS ONE 2015, 10, e0130116. [Google Scholar] [CrossRef] [Green Version]
- Kolukirik, M.; Ince, O.; Cetecioglu, Z.; Celikkol, S.; Ince, B.K. Spatial and temporal changes in microbial diversity of the Marmara Sea Sediments. Mar. Pollut. Bull. 2011, 62, 2384–2394. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Gong, Y.; Zhou, C.; Zhang, J.; Wang, Z.; Ning, K. Comparison and Interpretation of Taxonomical Structure of Bacterial Communities in Two Types of Lakes on Yun-Gui plateau of China. Sci. Rep. 2016, 6, 30616. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, L.; Zhu, Y.; Zhu, T.; Wu, X.; Yang, D. Microbial Community Structure and Its Driving Environmental Factors in Black Carp (Mylopharyngodon piceus) Aquaculture Pond. Water 2021, 13, 3089. [Google Scholar] [CrossRef]
- Zeng, J.; Lin, Y.; Zhao, D.; Huang, R.; Xu, H.; Jiao, C. Seasonality overwhelms aquacultural activity in determining the composition and assembly of the bacterial community in Lake Taihu, China. Sci. Total Environ. 2019, 683, 427–435. [Google Scholar] [CrossRef]
- Qin, M.; Xu, H.; Zeng, J.; Zhao, D.; Yu, Z.; Wu, Q.L. Composition and assembly of bacterial communities in surface and deeper sediments from aquaculture-influenced sites in Eastern Lake Taihu, China. Aquat. Sci. 2020, 82, 80. [Google Scholar] [CrossRef]
- Dongwei, H.; Renjun, Z.; Shenzheng, Z.; Dongdong, W.; Xisha, D.; Chengguang, X.; Shaoping, W.; Jianguo, H.; Zhijian, H. Stochastic processes shape the bacterial community assembly in shrimp cultural pond sediments. Appl. Microbiol. Biotechnol. 2021, 105, 5013–5022. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial | Final | |||||||
|---|---|---|---|---|---|---|---|---|
| CON | LD | MD | HD | CON | LD | MD | HD | |
| Nodes | 1462 | 1469 | 1392 | 1513 | 1961 | 1859 | 1809 | 1829 |
| Edges | 117,327 | 107,979 | 97,268 | 127,572 | 145,023 | 137,972 | 126,824 | 131,918 |
| Positive edge ratio | 65.92% | 63.16% | 60.55% | 63.90% | 59.99% | 59.24% | 65.17% | 68.89% |
| Negative edge ratio | 34.08% | 36.84% | 39.45% | 36.10% | 40.01% | 40.76% | 34.83% | 31.11% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Hou, Y.; Jia, R.; Li, B.; Zhu, J. Effects of Bellamya purificata Cultivation at Different Stocking Densities on the Dynamics and Assembly of Bacterial Communities in Sediment. Biomolecules 2023, 13, 254. https://doi.org/10.3390/biom13020254
Zhou M, Hou Y, Jia R, Li B, Zhu J. Effects of Bellamya purificata Cultivation at Different Stocking Densities on the Dynamics and Assembly of Bacterial Communities in Sediment. Biomolecules. 2023; 13(2):254. https://doi.org/10.3390/biom13020254
Chicago/Turabian StyleZhou, Mengmeng, Yiran Hou, Rui Jia, Bing Li, and Jian Zhu. 2023. "Effects of Bellamya purificata Cultivation at Different Stocking Densities on the Dynamics and Assembly of Bacterial Communities in Sediment" Biomolecules 13, no. 2: 254. https://doi.org/10.3390/biom13020254