
Organismal Roles of Hsp90

Department of Biological Sciences, The University of North Carolina at Charlotte, Charlotte, NC 28223, USA
Biomolecules 2023, 13(2), 251; https://doi.org/10.3390/biom13020251
Submission received: 20 December 2022
/
Revised: 24 January 2023
/
Accepted: 25 January 2023
/
Published: 29 January 2023
(This article belongs to the Special Issue Hsp90 Structure, Mechanism and Disease)
{kind=link}
{kind=link}
Abstract
:Heat shock protein 90 (Hsp90) is a highly conserved molecular chaperone that assists in the maturation of many client proteins involved in cellular signal transduction. As a regulator of cellular signaling processes, it is vital for the maintenance of cellular proteostasis and adaptation to environmental stresses. Emerging research shows that Hsp90 function in an organism goes well beyond intracellular proteostasis. In metazoans, Hsp90, as an environmentally responsive chaperone, is involved in inter-tissue stress signaling responses that coordinate and safeguard cell nonautonomous proteostasis and organismal health. In this way, Hsp90 has the capacity to influence evolution and aging, and effect behavioral responses to facilitate tissue-defense systems that ensure organismal survival. In this review, I summarize the literature on the organismal roles of Hsp90 uncovered in multicellular organisms, from plants to invertebrates and mammals.
1. Introduction
Cellular protein homeostasis depends on the integrity and function of its proteome, of which molecular chaperones play an indispensable role to maintain it. Heat shock protein 90 (Hsp90) is an essential and evolutionary conserved molecular chaperone, that except for archea, is found in all kingdoms of life [1]. Hsp90 is crucial for the viability and growth of eukaryotic cells and organisms, and it is one of the most abundant cellular proteins known to date, representing ~2% of total protein in a cell [2]. This abundance of Hsp90 is required to sustain the wide range of cellular processes it is implicated in, by chaperoning components involved in cellular signal transduction events such as protein kinases and steroid hormone receptors [3,4,5,6,7,8].
Hsp90 requires ATP function for its activity to help facilitate folding of client proteins, and this function is regulated and controlled by a variety of co-chaperones in a context-specific manner [7,9], as well as post translational modifications [10,11,12,13]. Briefly, each Hsp90 dimer binds ATP in its “open conformation”, which is followed by dimerization of the N-terminal domains of each protomer, allowing ATP hydrolysis. This subsequently leads to release of the folded and mature client protein, resulting in the open conformation of the Hsp90 dimer [7,8,14]. Each step along the Hsp90 chaperone cycle is finetuned by co-chaperones, such as, e.g., Cdc37/p50, which promote binding to kinase clients [15,16,17], or Aha1 and p23 that are involved in the regulation of Hsp90 ATP hydrolysis [14,18,19]. For more detailed information on the Hsp90 structure and regulation by co-chaperones, the reader is referred to articles and reviews specifically discussing this topic, including this Special Issue [8,20,21,22].
Because of its involvement in a wide range of cellular processes, Hsp90 supports an expansive network of more than 300 confirmed client proteins encompassing at least 5% of total proteins in yeast cells, and this number is similarly increased in multicellular organisms and mammals (https://www.picard.ch/downloads/Hsp90interactors.pdf; accessed 20 December 2022). Consequently, it is perhaps not surprising that Hsp90 is involved in almost every cellular process from cell cycle and a multitude of signal transduction pathways to protein trafficking, transcriptional processes and genomic stability [23,24,25,26,27,28,29]. In multicellular organisms, Hsp90’s involvement is expanded accordingly, and evidence in the past two decades has shown that Hsp90’s role reaches far beyond maintenance of signaling proteins: Its involvement ranges from development and evolution to intercellular stress signaling, aging responses and innate immunity, as well as neuronal function and behavior (Figure 1). In this review, I will highlight these organismal roles of Hsp90 which have been uncovered in different multicellular model systems, including plants, invertebrates and mammals.
2. Hsp90 in Organismal Development and Evolution
2.1. Development
The coordination of cell proliferation and differentiation is crucial for proper development. Hsp90’s central role in growth and development is profound, as its client proteins regulate almost all phases of the cell cycle. These include PI-3/AKT, NFkB and MAP kinase pathways, which drive progression through G1/S and G2/M checkpoints through transcriptional routes converging on Cyclin D, and Cyclin B and E [30]. Hsp90 also regulates various key cell cycle regulators directly, including Cdk1, Cdk2, Cdk4 and Cdk6 [31,32,33,34]. Furthermore, check point kinases Wee-1 and Myt-1 depend on Hsp90 function [35,36,37]. Later stages of mitosis and cytokinesis also depend on Hsp90 via mitotic regulators Survivin and Aurora B [38,39]. Because of Hsp90 influencing the cell cycle at multiple levels, either directly or indirectly, Hsp90 function is indispensable not only for organismal development but also for tumor cell progression, which was recognized early on through targeted inhibition of Hsp90 function using ansamycin inhibitors such as geldanamycin and 17-AAG [2,40,41,42]. Due to Hsp90 being involved in signaling pathways promoting cancer cell progression, it has become an attractive and well-established therapeutic cancer target, with Hsp90 inhibitors being continually developed and reviewed in clinical trials in an ongoing basis [43].
Most eukaryotic systems have two different cytosolic Hsp90 isoforms, with the exception of C. elegans that has only one cytosolic isoform (HSP-90/DAF-21). In mammals and yeast, the stress-inducible Hsp90α is encoded by the gene HSP90AA1 in humans (HSP82 in yeast), and the constitutively expressed Hsp90β is encoded by HSP90AB1 in humans and HSC82 in yeast. Although both isoforms share extensive sequence identity, their cellular functions are not completely identical and this is also demonstrated in their developmental requirements in an organism. For example, the Hsp90β knockout mouse shows early embryonic lethality [44], whereas this is not the case for mice lacking Hsp90α, which are viable but exhibit a failure of spermatogenesis and become sterile [45]. Moreover, while both isoforms are mutually expressed in most tissues in the mouse, the heart and muscle were found to harbor reduced levels of Hsp90α compared to Hsp90β [45]. Interestingly, zebrafish contains two Hsp90α genes, called Hsp90a1 and Hsp90a2. Hsp90a1 is crucial for myofibril organization in skeletal muscle development, whereas Hsp90a2 has no effect on muscle development [46]. Coherent with observations in vertebrates, the only cytosolic Hsp90 isoform in the invertebrate C. elegans (HSP-90) is crucial for myosin folding and muscle development, as RNAi-mediated knockdown leads to disrupted myosin filaments and motility defects [47,48]. Mammals such as mice, however, appear to require higher threshold levels of Hsp90 to promote stress adaptation and survival of the organism compared to yeast. This is accomplished through an internal ribosome entry site (IRES) in the 5’UTR of the Hsp90ab1 mRNA that can reprogram Hsp90 translational levels in stressed conditions [49].
As observed in other multicellular model organisms, depletion of Hsp90 by RNAi in C. elegans leads to morphological and transcriptional changes, including developmental changes to the gonad, vulval structures and oocyte development [50]. Indeed, Hsp90 regulates the meiotic prophase to metaphase transition during oocyte development by ensuring wee-1 kinase functionality, which results in reduced fertility in the worm [51]. Interestingly, besides transcriptional changes that demonstrate induction of the heat shock response, Hsp90 RNAi at the whole animal level also leads to induction of an innate immune response, by altering expression levels of innate immune genes primarily expressed in the intestine [50].
The role of Hsp90 in C. elegans development is further highlighted through its involvement in dauer formation, TGFβ- and Notch signaling. The C. elegans TGF-β pathway regulates a decision between reproductive development and arrest at a larval stage known as dauer that is suited for survival under conditions of environmental stress. Hsp90 itself is implicated in dauer formation through its interaction with two components of the TGF-β pathway, TGF-β-RI (DAF-1 in C. elegans) and TGF-β-RII (DAF-4 in C. elegans) [52]. Hsp90 also regulates the functionality of a DAF-11/guanyl cyclase signaling pathway in sensory ciliae and amphid neurons that controls dauer formation in response to environmental cues, in parallel to the TGF-β pathway [53,54], as well as chemosensory behaviors [54,55]. Germline proliferation in C. elegans requires signaling from the somatic gonad to the germline, which is mediated by GLP-1 (a Notch orthologue) [56]. If GLP-1/Notch signaling is defective through mutations in the glp-1 gene, germline stem cells prematurely exit mitosis and enter meiosis to form gametes, resulting in reduced germline proliferation and sterility [57]. Hsp90 has been identified as a regulator of Notch signaling that suppresses defective GLP-1/Notch signaling and promotes germline proliferation [58]. Strikingly, C. elegans depleted for Hsp90 by RNA interference or using an HSP-90(I461N) mutant leads to the formation of a proximal germline tumor, despite its reduced function and reduced GLP-1 signaling [57,58]. Further research in solving this paradox will be required to better illuminate the complex tissue-specific and organismal functions of the Hsp90 chaperone system in C. elegans and other metazoans.
2.2. Hsp90 as a Capacitor of Organismal Evolution
The physiological requirement of Hsp90 for the growth and development of model organisms was obvious early on using Saccharomyces cerevisiae [35,59], but was further highlighted using Drosophila melanogaster [60], where point mutations of the Drosophila Hsp90 (Hsp83) gene are lethal as homozygotes [61]. Although heterozygous mutant combinations are viable as adults, they are associated with sterility due to defects in microtubule dynamics during spermatogenesis [61]. Further experiments in Drosophila designed to identify suppressors of signal transduction Sevenless and Raf pathway mutants recovered Hsp90 mutants. Progeny of these Hsp90 mutants resulted in developmental abnormalities that, dependent on the genetic background, affected different morphological structures of the fruit fly [60,62]. This discovery led to further demonstration of Hsp90’s importance in Drosophila spermatogenesis and germline development [61]. It was one of the cornerstones that defined Hsp90’s prominent role in evolution, which was established as the “Hsp90 capacitor hypothesis” by the Lindquist lab [63].
The capacitor hypothesis demonstrated that diverse pathways become sensitive to the effects of genetic variation when Hsp90 function is compromised due to environmental stress, pharmacological inhibition or genetic mutation. It showed that Hsp90 functions in a wide variety of morphogenetic processes that are apparent in all model organisms tested, from yeast to vertebrates [63,64,65]. For example, the diverse phenotypes associated with Hsp90 impairment in Drosophila are deformed eye and thickened wing phenotypes [63], whereas in Arabidopsis thaliana, this leads to altered leaf and cotyledon shapes [64]. Similar consequences were observed in zebrafish upon reduced Hsp90 expression [66]. In C. elegans, the expression level of Hsp90 in particular varies during C. elegans embryonic development, causing embryos with stronger induction of Hsp90 to be less affected by mutation, thus buffering genetic variation [67]. However, individuals of a population with increased stress resistance due to higher Hsp90 levels show a “trade-off” with lowered reproductive potential, whereas worms with lower stress resistance are associated with higher reproductive fitness. The reason for this is thought to be a bet-hedging strategy, which is beneficial in ever-changing environments, that ensures survival of the population as a whole [67,68]. The reduced reproductive fitness due to increased Hsp90 expression perhaps highlights the requirement for Hsp90 expression levels to be tightly regulated due to its important role in germline development [57].
Importantly, phenotypic traits revealed upon temporary Hsp90 impairment can be selected for over several generations and become fixed in following generations, establishing Hsp90’s crucial role in evolution. An example for this in a natural setting was provided by the cavefish Asyanax mexicanus, where cryptic variation in eye size was masked by Hsp90 in the ancestral river but revealed when fish were kept in caves that challenged the Hsp90 system due to low-salinity water [65]. This even plays a role in human disease, as is the case in Fanconi anemia (FA), a complex autosomal recessive human cancer predisposition syndrome that results in point mutations of 19 genes involved in the FA genome maintenance pathway [69]. The function of less severe FANCA mutants was preserved by Hsp90 binding, which maintained FA pathway function but became destabilized and sensitive to genotoxic stress upon Hsp90 impairment [69].
However, while the evolutionary capacitor hypothesis relies on the potential of cytosolic protein instability that can be exposed upon Hsp90 inhibition, other contributions were shown to underlie Hsp90-dependent transcriptional mechanisms and chromatin structure [25,26,27,28,70,71]. While these are seemingly different mechanisms leading to the variety of Hsp90 buffered traits, it is perhaps a combination of multiple Hsp90-dependent genetic as well as epigenetic mechanisms working in concert.
3. Hsp90-Dependent Regulation of Organismal Proteostasis, Stress and Aging
3.1. Hsp90 in the Regulation of Cell Nonautonomous Stress Signaling
Hsp90, together with its co-chaperone machinery, is an integral part of the cellular network that safeguards proteostasis. As with other chaperones, Hsp90 expression is regulated by the stress transcription factor Heat Shock Factor 1 (HSF1) and is increased in response to environmental challenges that initiate the cytosolic heat shock response (HSR) [72]. This is accomplished in a negative feedback mechanism, whereby under normal conditions, HSF1 is sequestered by a multichaperone complex including Hsp90 and Hsp70 in an inactive monomeric form [73]. Proteotoxic stress conditions that increase the amount of misfolded proteins in the cell recruit the chaperones away from HSF1 towards the accumulating pool of misfolded proteins and releases HSF1 monomers. This in turn allows HSF1 monomers to form homotrimers that translocate to the nucleus, where they bind to heat shock elements (HSE) that induce molecular chaperones (heat shock proteins), as well as trafficking and proteolytic genes, in order to restore cytosolic proteostasis. Once the levels of Hsp90 and other chaperones have sufficiently increased in the cytosol to refold damaged proteins, they are recruited back to HSF1.
However, expression of Hsp90 can also, directly or indirectly, be regulated by other transcription factors in addition to HSF-1. For example in C. elegans, the GATA transcription factor PQM-1 responds to local changes in Hsp90 expression levels as a mediator of transcellular chaperone signaling, but also regulates Hsp90 expression itself [74]. In addition to HSF1, Hsp90 also regulates the function of the FOXO orthologue DAF-16 isoform A by facilitating its translocation into the nucleus upon heat stress and reduced ILS [75].
In metazoans, the stress-dependent induction of HSF-1 transcriptional activity also depends on intercellular stress signaling responses. In C. elegans, temperature alterations are sensed by two thermosensory AFD neurons that control temperature-dependent behaviors. This is accomplished through the action of the guanylyl cyclase GCY-8 that is specifically expressed in AFD neurons, and which controls HSF1-dependent induction of the HSR in distal cells in order to restore proteostasis at the organismal level. Neuronal control of proteostasis in response to acute temperature challenges is, however, uncoupled from aging-related responses via a GPCR thermal receptor GTR-1 expressed in chemosensory neurons [76]. The C. elegans nervous system relays the signal to distal organs via the neurotransmitter serotonin, thus involving serotonergic neurocircuitry [77,78]. However, astrocyte-like cells in the nervous system can also regulate the cell-nonautonomous HSR in an HSF-1 dependent manner that does not rely on known neurotransmitters but instead requires small clear vesicle release [79]. Non-neuronal tissues such as muscle and gut cells can equally relay information of temperature changes to thermosensory AFD neurons via estrogen signaling through the nuclear hormone receptor NHR-69 [80], an orthologue of the human HNF4 transcription factors that are clients of Hsp90 [81].
Interestingly, heat shock leads to rapid induction of HSF1 activity in the C. elegans germline [77,82,83] and HSF1 is required for gametogenesis in invertebrates and vertebrates [72,84]. Like Hsp90, HSF1 is required for germline proliferation and fecundity, relying on Insulin/IGF-1 signaling in the soma that nonautonomously activates HSF-1 in the germline [85], although whether Hsp90 is involved in this regulation is currently unknown.
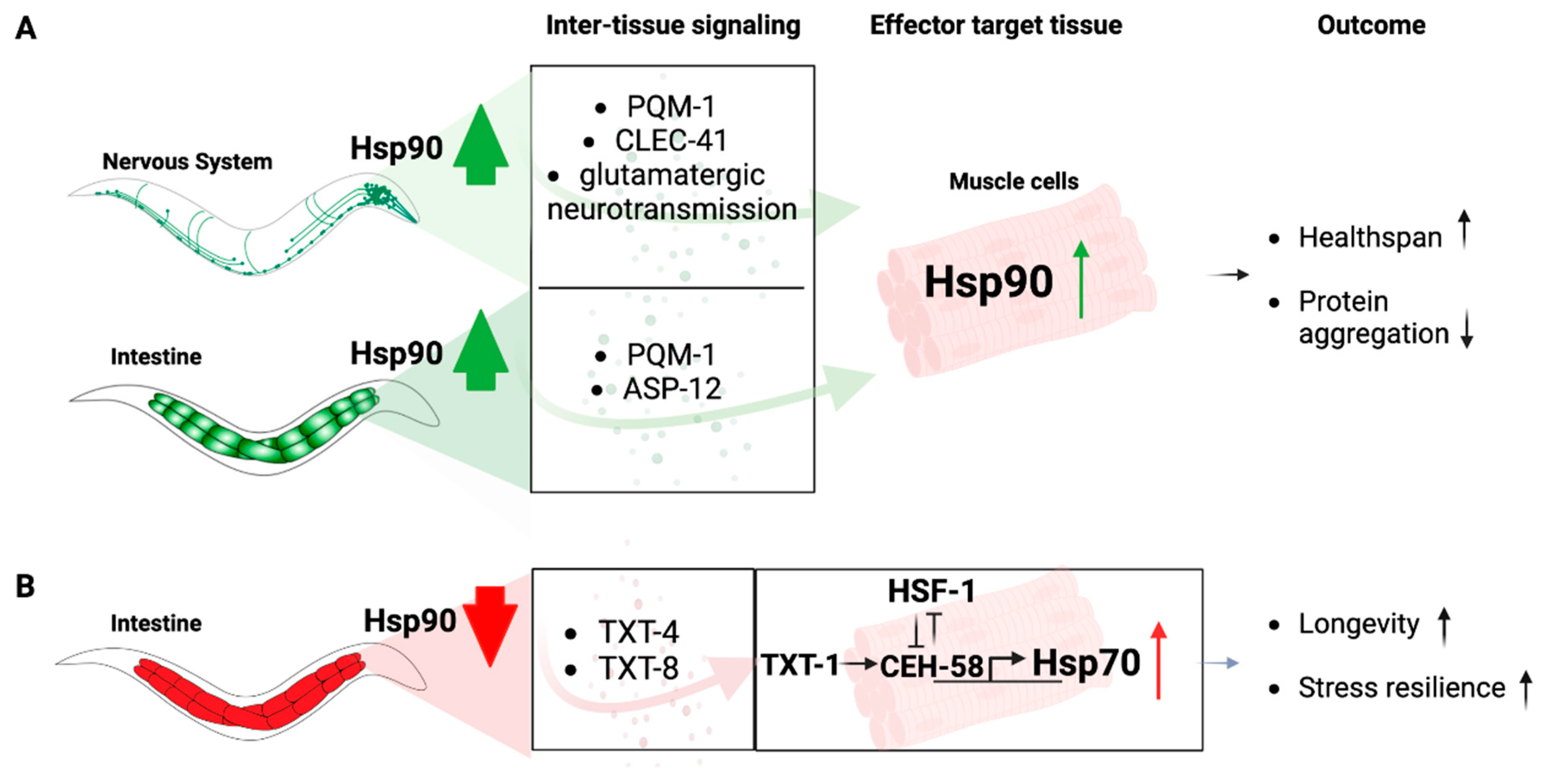
However, Hsp90 is itself involved in relaying signals from one tissue to another, particularly when its expression levels are altered in the gut or the nervous system, an organismal stress signaling response known as Transcellular Chaperone Signaling (Figure 2) [86,87]. Enhancement of Hsp90 capacity in the gut or the neurons leads to a compensatory transcriptional inter-tissue response, regulated via the transcription factor PQM-1, that induces Hsp90 expression in other distal cell types and primarily muscle cells [74,88]. This protects against the age-associated debilitating consequences of misfolded proteins expressed in muscle cells, including human amyloid beta protein or endogenously expressed metastable myosin [74,88]. How this transcriptional response is relayed from one tissue to another, however, depends on tissue context. Transcellular chaperone signaling from neurons to the muscle requires glutamatergic signaling and relies on the c-type lectin clec-41 that associates with AMPA receptor in glutamatergic neurons (Figure 2A) [74]. Increased Hsp90 expression in the gut is relayed via the secreted immune peptide asp-12 which leads to transcriptional upregulation of Hsp90 in muscle cells (Figure 2A) [74].
On the other hand, when Hsp90 levels are reduced by tissue-specific RNA interference in the gut, a compensatory signaling mechanism elevates Hsp70 expression in distant cells (Figure 2B). This is, however, not mediated by a mechanism that relies on HSF1 to activate a canonical HSR, but depends on a homeodomain transcription factor, CEH-58. HSF1 transcriptional activity is suppressed upon gut-specific Hsp90 depletion, and induction of Hsp70 relies on a different intercellular signaling cue involving TXT-1, a membrane-associated guanylate cyclase that relays the signal received from the intestine to the muscle cell nucleus where the homeodomain transcription factor CEH-58 induces Hsp70 expression (Figure 2B) [87]. Thus, there is a difference in intercellular-signaling components which depend on the tissue-type perceiving altered Hsp90 expression levels. This argues for multiple and complex layers of responses that cannot be answered by one particular molecular mechanism, at least not in a multicellular organismal setting [87]. This demonstrates that in metazoans, local Hsp90 capacity can regulate organismal proteostasis and stress resilience via Transcellular Chaperone Signaling.
Comparable organismal effects as a result of local induction of the HSF-1 mediated HSR is also observed in mammals via neuroendocrine signaling. For example, rats undergoing restraint stress have higher cortisol levels secreted by the pituitary gland which signals to activate HSF-1 in the adrenal glands in the kidney to induce Hsp70 expression [89], although how Hsp90 itself could potentially be involved in this response is currently not known and will require further research.
One question that often arises is whether Hsp90 is secreted as part of inter-tissue stress signaling in an organism. Hsp90 secretion has been observed in tissue-culture in response to a variety of stress conditions as well as in cancer cells [90]. Clinically, skin injury promotes Hsp90α secretion and potentiates wound healing in tissue-culture, pigs and dogs [91,92]. However, secreted, extracellular Hsp90 has not been observed as a signaling component itself involved in inter-tissue stress signaling in an organism. In fact, secretion of Hsp90 was not detected in C. elegans overexpressing Hsp90 in different cell types [88]. For a more detailed review on the roles of secreted, extracellular Hsp90, the reader is referred to reviews by Li and colleagues [93] in this Special Issue on Hsp90.
3.2. Hsp90-Dependent Regulation of Lifespan and Aging
Consistent with a growth-promoting role, substantial depletion of Hsp90 by RNAi-mediated knockdown can lead to growth defects and larval arrest, and even shorten lifespan [50,75]. The developmental defects associated with Hsp90 RNAi are morphological changes to the gonad and vulva, induction of the HSR and changes to the muscle ultrastructure [50]. Importantly, however, mild impairment of Hsp90 either by RNAi or pharmacological inhibition leads to lifespan extension and enhances health span [94]. This was shown in a pharmacological geroprotector screen using C. elegans that identified two Hsp90 inhibitors, Tanespimycin and Monorden, that extended lifespan and improved health of the nematode throughout the course of aging [94]. The study found that both inhibitors acted through HSF1 to induce the age-defying and health span-inducing effects in the worm. This is consistent with HSF1’s role in promoting longevity [95,96,97]. Similar to mild Hsp90 impairment by inhibitors, moderate depletion of Hsp90 RNAi in the gut also enhances lifespan and stress resilience in C. elegans without any developmental issues [87]. Similar observations were made in vertebrates, where transient knockdown of Hsp90 during embryonic development in zebrafish results in cold stress resistance in adult animals [98]. Interestingly, the Hs90 co-chaperone p23 acts in key longevity pathways to regulate lifespan in a temperature-dependent manner [99]. At elevated temperatures, p23 mutation extends lifespan through DAF-16 and HSF1 signaling pathways. Short-lived phenotypes depend on the DAF-12 steroid receptor signaling pathway [99], with DAF-12 being a type II nuclear receptor that resembles the human thyroid receptor and is a known client of the p23-HSP90 complex [100]. Apart from being involved in the key longevity pathways, ILS/IGF-1 signaling and HSF-1 signaling pathways, Hsp90 is also involved in the regulation of SIRT1 in both C. elegans and mammalian cells [101]. Thus, Hsp90 is unique, as it is a major facilitator that ensures the efficacy of all signaling processes maintaining organismal health and promoting survival.
4. Pathogen Response and Innate Immunity
The involvement of Hsp90 in immune responses is manifold, as it is implicated in the adaptive as well as innate immunity pathways in almost all organisms. In plants, R proteins are client proteins of Hsp90, which is important for the defense response against microbial pathogens [102,103]. The activation of R proteins results in local cell death to limit pathogen proliferation. Because of this, R protein activation also needs to be tightly controlled to avoid tissue damage, which is regulated by Hsp90 [102,103].
In the invertebrate C. elegans, which does not have an adaptive immune response, Hsp90 plays an important role in the innate immune response via HSF1. For example, mutant Hsp90, as well as heat shock, causes release of HSF1 from Hsp90, resulting in HSF1 initiating expression of antimicrobial peptide genes [104,105]. Coherently, depletion of Hsp90 by RNA interference also induces an innate immune transcriptional response that was proposed to be similar to the immune response after C. elegans exposure to Pseudomona aeruginosa [50]. Similarly, pathogen-infected wax moths treated with Hsp90 inhibitor 17-DMAG were protected by an increased immune response [106]. This breadth of Hsp90 being implicated in a process that mediates innate immunity via HSF1 activation demonstrates the importance and conservation of Hsp90 in the innate immune response. In mammals, Hsp90 is implicated in the presentation of antigen to T-cells and activation of macrophages [107]. Hsp90 mediates antigen presentation in target antigen-presenting cells (APC) by facilitating endocytosis of bound polypeptides [108,109]. These generated antigenic peptides are presented to MHC-I/II by Hsp90 [110]. Extracellular Hsp90 can also bind to peptide antigens to facilitate uptake of the Hsp90 antigen complex by endocytosis [107]. After the antigen is internalized, intracellular Hsp90 facilitates further processing of these peptides to the proteasome for degradation [107]. Interestingly, Hsp90 also regulates the reactivation of the human immunodeficiency virus (HIV-1) via regulation of the PKC/ERK MAPK pathways, which influences replication and gene expression of the virus [111]. In the response to pathogens, extracellular Hsp90 can act as a damage-associated molecular pattern (DAMP) signal that regulates the production of cytokines in response to pathogenic infection and inflammation [112]. This involvement of p38 and ERK MAPK pathways in response to pathogens was shown to require Hsp90 for their function through direct interaction of Hsp90 with MAP kinases p38 and ERK in evolutionary diverse organisms [16,113,114]. Hsp90 also plays an important role in growth, development and virulence of parasitic pathogens itself, such as the parasitic protozoa Plasmodium falciparum [115] and Toxoplasma gondii [116,117]. This makes Hsp90 a high-value drug target to inhibit the parasite’s growth and infection cycle in humans [117]. In summary, the role of Hsp90 in the adaptive and innate immune response is vast, and the reader is referred to specialized reviews on this topic for more detailed information (e.g., [110,118]).
5. Neuronal Signaling and Behavior
Considering the wide range of client proteins dependent on Hsp90 function, it is perhaps unsurprising, but nevertheless fascinating, to find it involved in neuronal signaling and function. Some of the first experimental evidence demonstrating a role for Hsp90 in neuronal function stems from research in C. elegans. Hsp90 is crucial for the function of specific chemosensory amphid neurons required to sense pheromones and other attractants. It was proposed that Hsp90 accomplishes this through interaction and stabilization of the transmembrane guanylyl cyclase DAF-11, which regulates cGMP levels, a prominent second messenger in C. elegans chemosensory transduction [54,55].
In mice, Hsp90 is required for the constitutive trafficking of glutamatergic AMPA-type receptors into synapses during their continuous cycling between synaptic and non-synaptic sites, as well as efficient neurotransmitter release at the presynaptic terminal [119]. In addition to its role in neuronal signaling, Hsp90 chaperones the pro-regenerative dual leucine zipper kinase (DLK), a critical neuronal sensor that drives axon regeneration, degeneration and neurological disease in Drosophila and mammalian neurons [120]. This suggests a vital role for Hsp90 in axon injury signaling, as well as neuronal function that is evolutionary conserved in both vertebrates and invertebrates.
With this importance for neuronal signaling, is it possible that Hsp90 could be involved in the regulation of behavioral responses that facilitate survival during stress conditions? There is at least one example in C. elegans that provides direct evidence supporting such a role. Exposure of nematodes to high concentrations of volatile compounds, such as benzaldehyde and diacetyl, induces toxicity and food avoidance behavior [121]. However, preconditioning with benzaldehyde activates stress responses mediated via DAF-16, SKN-1 and HSP-90 in non-neuronal cells that confer increased stress resilience and behavioral tolerance [121]. Another example is provided by the heat stress-induced activation of HSF1, which regulates behavioral responses through estrogen signaling from non-neuronal cells to thermosensory neurons [80]. hsf-1 mutants are defective in their thermotactic response towards temperature, i.e., migration towards cultivation temperature. Expression of wild type HSF-1 in muscle or intestinal cells rescued this behavioral defect via activation of the NHR-69 nuclear hormone receptor involved in estrogen-like signaling [80], which is a client of Hsp90, as mentioned earlier. Thus, Hsp90, through its role in multiple stress-responsive signaling pathways, may influence behavioral outputs in order to promote survival during environmental stress conditions.
6. Outlook and Conclusions
As a chaperone safeguarding the functionality of clients involved in almost every cellular signaling process, Hsp90 is essential for cellular homeostasis. At the organismal level, intercellular signaling processes that require the involvement of Hsp90 may be underlying the organismal coordination of extra- and intracellular signaling networks between and across different tissues and organs. Especially at the organismal level, many open questions remain to fully comprehend the organismal biology of Hsp90, particularly with regard to intercellular stress signaling.
For example, (1) is there is a tissue map or tissue hierarchy allowing highly coordinated signaling responses to occur? We know that stress signaling can be regulated via both the nervous system and non-neuronal cell types, with, e.g., muscle and gut cells transmitting feedback information to the nervous system or even suppressing stress responses in different cell types and organs. (2) What is the tissue-specific Hsp90 interactome in an organism and how are potential Hsp90 interactors of these tissue-specific networks contributing to intercellular stress signaling? (3) Is there a role of extracellular Hsp90 in intercellular signaling processes? (4) If the tissue-specific expression levels of Hsp90 can affect stress responses in distant tissues, is there a naturally occurring/physiological condition that alters Hsp90 expression levels to induce transcellular chaperone signaling? (5) As Hsp90 function is tightly regulated by co-chaperones and post translational modifications [13], we currently do not know how co-chaperones of the Hsp90 machinery and its PTMs are involved in organismal proteostasis. For example, it can be envisioned that stress responses and intercellular stress signaling pathways are similarly influenced and perhaps finetuned through tissue-specific PTMs and co-chaperone networks. (6) How do the organismal roles of Hsp90 affect diseases, including neurodegenerative diseases and cancer, in a tissue- and disease-specific context?
Thus, the involvement of Hsp90 in almost all aspects of organismal biology, from development to aging, stress adaptation, evolution and different diseases including cancer and neurodegenerative diseases, places it at the nexus of a plethora of cell nonautonomous signaling processes. The challenge for future research will be to navigate through these inter-tissue signaling pathways in a comprehensive manner to understand their increased complexity in the multicellular setting of an organism.
Funding
This work was supported by laboratory start-up funds to P.v.O.-H. from the University of North Carolina at Charlotte.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The Figure was created using Biorender.com.
Conflicts of Interest
The author declares no conflict of interest.
References
- Johnson, J.L. Evolution and Function of Diverse Hsp90 Homologs and Cochaperone Proteins. Biochim. Biophys. Acta 2012, 1823, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitesell, L.; Lindquist, S.L. HSP90 and the Chaperoning of Cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Picard, D. Chaperoning Steroid Hormone Action. Trends Endocrinol. Metab. 2006, 17, 229–235. [Google Scholar] [CrossRef]
- Picard, D.; Khursheed, B.; Garabedian, M.J.; Fortin, M.G.; Lindquist, S.; Yamamoto, K.R. Reduced Levels of Hsp90 Compromise Steroid Receptor Action in Vivo. Nature 1990, 348, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, P.C.; Picard, D. Molecular Chaperones, Essential Partners of Steroid Hormone Receptors for Activity and Mobility. Biochim. Biophys. Acta 2010, 1803, 641–649. [Google Scholar] [CrossRef]
- Riggs, D.L.; Roberts, P.J.; Chirillo, S.C.; Cheung-Flynn, J.; Prapapanich, V.; Ratajczak, T.; Gaber, R.; Picard, D.; Smith, D.F. The Hsp90-Binding Peptidylprolyl Isomerase FKBP52 Potentiates Glucocorticoid Signaling in Vivo. EMBO J. 2003, 22, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, P.; Rohrberg, J.; Biebl, M.M.; Rutz, D.A.; Buchner, J. The Plasticity of the Hsp90 Co-Chaperone System. Mol. Cell 2017, 67, 947–961.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodromou, C.; Bjorklund, D.M. Advances towards Understanding the Mechanism of Action of the Hsp90 Complex. Biomolecules 2022, 12, 600. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L. Mutations in Hsp90 Cochaperones Result in a Wide Variety of Human Disorders. Front. Mol. Biosci. 2021, 8, 787260. [Google Scholar] [CrossRef]
- Mollapour, M.; Tsutsumi, S.; Truman, A.W.; Xu, W.; Vaughan, C.K.; Beebe, K.; Konstantinova, A.; Vourganti, S.; Panaretou, B.; Piper, P.W.; et al. Threonine 22 Phosphorylation Attenuates Hsp90 Interaction with Cochaperones and Affects Its Chaperone Activity. Mol. Cell 2011, 41, 672–681. [Google Scholar] [CrossRef]
- Woodford, M.R.; Truman, A.W.; Dunn, D.M.; Jensen, S.M.; Cotran, R.; Bullard, R.; Abouelleil, M.; Beebe, K.; Wolfgeher, D.; Wierzbicki, S.; et al. Mps1 Mediated Phosphorylation of Hsp90 Confers Renal Cell Carcinoma Sensitivity and Selectivity to Hsp90 Inhibitors. Cell Rep. 2016, 14, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollapour, M.; Bourboulia, D.; Beebe, K.; Woodford, M.R.; Polier, S.; Hoang, A.; Chelluri, R.; Li, Y.; Guo, A.; Lee, M.-J.; et al. Asymmetric Hsp90 N Domain SUMOylation Recruits Aha1 and ATP-Competitive Inhibitors. Mol. Cell 2014, 53, 317–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backe, S.J.; Sager, R.A.; Woodford, M.R.; Makedon, A.M.; Mollapour, M. Post-Translational Modifications of Hsp90 and Translating the Chaperone Code. J. Biol. Chem. 2020, 295, 11099–11117. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.U.; Roe, S.M.; Vaughan, C.K.; Meyer, P.; Panaretou, B.; Piper, P.W.; Prodromou, C.; Pearl, L.H. Crystal Structure of an Hsp90–Nucleotide–P23/Sba1 Closed Chaperone Complex. Nature 2006, 440, 1013–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawle, P.; Siepmann, M.; Harst, A.; Siderius, M.; Reusch, H.P.; Obermann, W.M.J. The Middle Domain of Hsp90 Acts as a Discriminator between Different Types of Client Proteins. Mol. Cell. Biol. 2006, 26, 8385–8395. [Google Scholar] [CrossRef] [Green Version]
- Hawle, P.; Horst, D.; Bebelman, J.P.; Yang, X.X.; Siderius, M.; van der Vies, S.M. Cdc37p Is Required for Stress-Induced High-Osmolarity Glycerol and Protein Kinase C Mitogen-Activated Protein Kinase Pathway Functionality by Interaction with Hog1p and Slt2p (Mpk1p). Eukaryot. Cell 2007, 6, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, C.K.; Gohlke, U.; Sobott, F.; Good, V.M.; Ali, M.M.U.; Prodromou, C.; Robinson, C.V.; Saibil, H.R.; Pearl, L.H. Structure of an Hsp90-Cdc37-Cdk4 Complex. Mol. Cell 2006, 23, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, P.; Prodromou, C.; Liao, C.; Hu, B.; Roe, S.M.; Vaughan, C.K.; Vlasic, I.; Panaretou, B.; Piper, P.W.; Pearl, L.H. Structural Basis for Recruitment of the ATPase Activator Aha1 to the Hsp90 Chaperone Machinery. EMBO J. 2004, 23, 1402–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotz, G.P.; Lin, H.; Harst, A.; Obermann, W.M.J. Aha1 Binds to the Middle Domain of Hsp90, Contributes to Client Protein Activation, and Stimulates the ATPase Activity of the Molecular Chaperone. J. Biol. Chem. 2003, 278, 17228–17235. [Google Scholar] [CrossRef] [Green Version]
- Pearl, L.H.; Prodromou, C. Structure and Mechanism of the Hsp90 Molecular Chaperone Machinery. Annu. Rev. Biochem. 2006, 75, 271–294. [Google Scholar] [CrossRef]
- Maiti, S.; Picard, D. Cytosolic Hsp90 Isoform-Specific Functions and Clinical Significance. Biomolecules 2022, 12, 1166. [Google Scholar] [CrossRef] [PubMed]
- Biebl, M.M.; Buchner, J. Structure, Function, and Regulation of the Hsp90 Machinery. Cold Spring Harb. Perspect. Biol. 2019, 11, a034017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mankovich, A.G.; Freeman, B.C. Regulation of Protein Transport Pathways by the Cytosolic Hsp90s. Biomolecules 2022, 12, 1077. [Google Scholar] [CrossRef] [PubMed]
- McClellan, A.J.; Xia, Y.; Deutschbauer, A.M.; Davis, R.W.; Gerstein, M.; Frydman, J. Diverse Cellular Functions of the Hsp90 Molecular Chaperone Uncovered Using Systems Approaches. Cell 2007, 131, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echtenkamp, F.J.; Gvozdenov, Z.; Adkins, N.L.; Zhang, Y.; Lynch-Day, M.; Watanabe, S.; Peterson, C.L.; Freeman, B.C. Hsp90 and P23 Molecular Chaperones Control Chromatin Architecture by Maintaining the Functional Pool of the RSC Chromatin Remodeler. Mol. Cell 2016, 64, 888–899. [Google Scholar] [CrossRef] [Green Version]
- DeZwaan, D.C.; Toogun, O.A.; Echtenkamp, F.J.; Freeman, B.C. The Hsp82 Molecular Chaperone Promotes a Switch between Unextendable and Extendable Telomere States. Nat. Struct. Mol. Biol. 2009, 16, 711–716. [Google Scholar] [CrossRef]
- Sawarkar, R.; Paro, R. Hsp90@chromatin.Nucleus: An Emerging Hub of a Networker. Trends Cell Biol. 2013, 23, 193–201. [Google Scholar] [CrossRef]
- Antonova, A.; Hummel, B.; Khavaran, A.; Redhaber, D.M.; Aprile-Garcia, F.; Rawat, P.; Gundel, K.; Schneck, M.; Hansen, E.C.; Mitschke, J.; et al. Heat-Shock Protein 90 Controls the Expression of Cell-Cycle Genes by Stabilizing Metazoan-Specific Host-Cell Factor HCFC1. Cell Rep. 2019, 29, 1645–1659.e9. [Google Scholar] [CrossRef] [Green Version]
- Gvozdenov, Z.; Bendix, L.D.; Kolhe, J.; Freeman, B.C. The Hsp90 Molecular Chaperone Regulates the Transcription Factor Network Controlling Chromatin Accessibility. J. Mol. Biol. 2019, 431, 4993–5003. [Google Scholar] [CrossRef]
- Muise-Helmericks, R.C.; Grimes, H.L.; Bellacosa, A.; Malstrom, S.E.; Tsichlis, P.N.; Rosen, N. Cyclin D Expression Is Controlled Post-Transcriptionally via a Phosphatidylinositol 3-Kinase/Akt-Dependent Pathway *. J. Biol. Chem. 1998, 273, 29864–29872. [Google Scholar] [CrossRef]
- Bedin, M.; Gaben, A.-M.; Saucier, C.; Mester, J. Geldanamycin, an Inhibitor of the Chaperone Activity of HSP90, Induces MAPK-Independent Cell Cycle Arrest. Int. J. Cancer 2004, 109, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Mahony, D.; Parry, D.A.; Lees, E. Active Cdk6 Complexes Are Predominantly Nuclear and Represent Only a Minority of the Cdk6 in T Cells. Oncogene 1998, 16, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, L.; Leng, X.; Parker, S.B.; Harper, J.W. Mammalian P50Cdc37 Is a Protein Kinase-Targeting Subunit of Hsp90 That Binds and Stabilizes Cdk4. Genes Dev. 1996, 10, 1491–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, M.J.; Jimenez, J. Genetic Interactions between Hsp90 and the Cdc2 Mitotic Machinery in the Fission Yeast Schizosaccharomyces Pombe. Mol. Gen. Genet 1999, 261, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Aligue, R.; Akhavan-Niak, H.; Russell, P. A Role for Hsp90 in Cell Cycle Control: Wee1 Tyrosine Kinase Activity Requires Interaction with Hsp90. EMBO J. 1994, 13, 6099–6106. [Google Scholar] [CrossRef] [PubMed]
- Goes, F.S.; Martin, J. Hsp90 Chaperone Complexes Are Required for the Activity and Stability of Yeast Protein Kinases Mik1, Wee1 and Swe1. Eur. J. Biochem. 2001, 268, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Mollapour, M.; Tsutsumi, S.; Neckers, L. Hsp90 Phosphorylation, Wee1 and the Cell Cycle. Cell Cycle 2010, 9, 2310–2316. [Google Scholar] [CrossRef] [PubMed]
- Fortugno, P.; Beltrami, E.; Plescia, J.; Fontana, J.; Pradhan, D.; Marchisio, P.C.; Sessa, W.C.; Altieri, D.C. Regulation of Survivin Function by Hsp90. Proc. Natl. Acad. Sci. USA 2003, 100, 13791–13796. [Google Scholar] [CrossRef] [Green Version]
- Lange, B.M.H.; Rebollo, E.; Herold, A.; González, C. Cdc37 Is Essential for Chromosome Segregation and Cytokinesis in Higher Eukaryotes. EMBO J. 2002, 21, 5364–5374. [Google Scholar] [CrossRef] [Green Version]
- Jameel, A.; Skilton, R.A.; Campbell, T.A.; Chander, S.K.; Coombes, R.C.; Luqmani, Y.A. Clinical and Biological Significance of HSP89 Alpha in Human Breast Cancer. Int. J. Cancer 1992, 50, 409–415. [Google Scholar] [CrossRef]
- Whitesell, L.; Mimnaugh, E.G.; De Costa, B.; Myers, C.E.; Neckers, L.M. Inhibition of Heat Shock Protein HSP90-Pp60v-Src Heteroprotein Complex Formation by Benzoquinone Ansamycins: Essential Role for Stress Proteins in Oncogenic Transformation. Proc. Natl. Acad. Sci. USA 1994, 91, 8324–8328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neckers, L. Hsp90 Inhibitors as Novel Cancer Chemotherapeutic Agents. Trends Mol. Med. 2002, 8, S55–S61. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, Y. Recent Advances in the Discovery of Novel HSP90 Inhibitors: An Update from 2014. Curr. Drug Targets 2020, 21, 302–317. [Google Scholar] [CrossRef]
- Voss, A.K.; Thomas, T.; Gruss, P. Mice Lacking HSP90beta Fail to Develop a Placental Labyrinth. Development 2000, 127, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Grad, I.; Cederroth, C.R.; Walicki, J.; Grey, C.; Barluenga, S.; Winssinger, N.; Massy, B.D.; Nef, S.; Picard, D. The Molecular Chaperone Hsp90α Is Required for Meiotic Progression of Spermatocytes beyond Pachytene in the Mouse. PLoS ONE 2010, 5, e15770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.J.; Li, H.; Bian, Y.; Zhong, Y. Heat-Shock Protein 90α1 Is Required for Organized Myofibril Assembly in Skeletal Muscles of Zebrafish Embryos. Proc. Natl. Acad. Sci. USA 2008, 105, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaiser, A.M.; Kaiser, C.J.O.; Haslbeck, V.; Richter, K. Downregulation of the Hsp90 System Causes Defects in Muscle Cells of Caenorhabditis Elegans. PLoS ONE 2011, 6, e25485. [Google Scholar] [CrossRef] [Green Version]
- Frumkin, A.; Dror, S.; Pokrzywa, W.; Bar-Lavan, Y.; Karady, I.; Hoppe, T.; Ben-Zvi, A. Challenging Muscle Homeostasis Uncovers Novel Chaperone Interactions in Caenorhabditis Elegans. Front. Mol. Biosci. 2014, 1, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, K.; Maiti, S.; Zahoran, S.; Weidenauer, L.; Hany, D.; Wider, D.; Bernasconi, L.; Quadroni, M.; Collart, M.; Picard, D. Translational Reprogramming in Response to Accumulating Stressors Ensures Critical Threshold Levels of Hsp90 for Mammalian Life. Nat. Commun. 2022, 13, 6271. [Google Scholar] [CrossRef] [PubMed]
- Eckl, J.; Sima, S.; Marcus, K.; Lindemann, C.; Richter, K. Hsp90-Downregulation Influences the Heat-Shock Response, Innate Immune Response and Onset of Oocyte Development in Nematodes. PLoS ONE 2017, 12, e0186386. [Google Scholar] [CrossRef]
- Inoue, T.; Hirata, K.; Kuwana, Y.; Fujita, M.; Miwa, J.; Roy, R.; Yamaguchi, Y. Cell Cycle Control by Daf-21/Hsp90 at the First Meiotic Prophase/Metaphase Boundary during Oogenesis in Caenorhabditis Elegans. Dev. Growth Differ. 2006, 48, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tewari, M.; Hu, P.J.; Ahn, J.S.; Ayivi-Guedehoussou, N.; Vidalain, P.-O.; Li, S.; Milstein, S.; Armstrong, C.M.; Boxem, M.; Butler, M.D.; et al. Systematic Interactome Mapping and Genetic Perturbation Analysis of a C. Elegans TGF-β Signaling Network. Mol. Cell 2004, 13, 469–482. [Google Scholar] [CrossRef]
- Murakami, M.; Koga, M.; Ohshima, Y. DAF-7/TGF-Beta Expression Required for the Normal Larval Development in C. Elegans Is Controlled by a Presumed Guanylyl Cyclase DAF-11. Mech. Dev. 2001, 109, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Birnby, D.A.; Link, E.M.; Vowels, J.J.; Tian, H.; Colacurcio, P.L.; Thomas, J.H. A Transmembrane Guanylyl Cyclase (DAF-11) and Hsp90 (DAF-21) Regulate a Common Set of Chemosensory Behaviors in Caenorhabditis Elegans. Genetics 2000, 155, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Vowels, J.J.; Thomas, J.H. Multiple Chemosensory Defects in Daf-11 and Daf-21 Mutants of Caenorhabditis Elegans. Genetics 1994, 138, 303–316. [Google Scholar] [CrossRef]
- Greenwald, I.; Kovall, R. Notch Signaling: Genetics and Structure. WormBook 2013, 1–28. [Google Scholar] [CrossRef]
- Qiao, L.; Lissemore, J.L.; Shu, P.; Smardon, A.; Gelber, M.B.; Maine, E.M. Enhancers of Glp-1, a Gene Required for Cell-Signaling in Caenorhabditis Elegans, Define a Set of Genes Required for Germline Development. Genetics 1995, 141, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Lissemore, J.L.; Connors, E.; Liu, Y.; Qiao, L.; Yang, B.; Edgley, M.L.; Flibotte, S.; Taylor, J.; Au, V.; Moerman, D.G.; et al. The Molecular Chaperone HSP90 Promotes Notch Signaling in the Germline of Caenorhabditis Elegans. G3 (Bethesda) 2018, 8, 1535–1544. [Google Scholar] [CrossRef] [Green Version]
- Borkovich, K.A.; Farrelly, F.W.; Finkelstein, D.B.; Taulien, J.; Lindquist, S. Hsp82 Is an Essential Protein That Is Required in Higher Concentrations for Growth of Cells at Higher Temperatures. Mol. Cell. Biol. 1989, 9, 3919–3930. [Google Scholar] [CrossRef]
- Cutforth, T.; Rubin, G.M. Mutations in Hsp83 and Cdc37 Impair Signaling by the Sevenless Receptor Tyrosine Kinase in Drosophila. Cell 1994, 77, 1027–1036. [Google Scholar] [CrossRef]
- Yue, L.; Karr, T.L.; Nathan, D.F.; Swift, H.; Srinivasan, S.; Lindquist, S. Genetic Analysis of Viable Hsp90 Alleles Reveals a Critical Role in Drosophila Spermatogenesis. Genetics 1999, 151, 1065–1079. [Google Scholar] [CrossRef]
- van der Straten, A.; Rommel, C.; Dickson, B.; Hafen, E. The Heat Shock Protein 83 (Hsp83) Is Required for Raf-Mediated Signalling in Drosophila. EMBO J. 1997, 16, 1961–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, S.L.; Lindquist, S. Hsp90 as a Capacitor for Morphological Evolution. Nature 1998, 396, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Queitsch, C.; Sangster, T.A.; Lindquist, S. Hsp90 as a Capacitor of Phenotypic Variation. Nature 2002, 417, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Rohner, N.; Jarosz, D.F.; Kowalko, J.E.; Yoshizawa, M.; Jeffery, W.R.; Borowsky, R.L.; Lindquist, S.; Tabin, C.J. Cryptic Variation in Morphological Evolution: HSP90 as a Capacitor for Loss of Eyes in Cavefish. Science 2013, 342, 1372–1375. [Google Scholar] [CrossRef] [Green Version]
- Yeyati, P.L.; Bancewicz, R.M.; Maule, J.; van Heyningen, V. Hsp90 Selectively Modulates Phenotype in Vertebrate Development. PLoS Genet 2007, 3, e43. [Google Scholar] [CrossRef] [Green Version]
- Burga, A.; Casanueva, M.O.; Lehner, B. Predicting Mutation Outcome from Early Stochastic Variation in Genetic Interaction Partners. Nature 2011, 480, 250–253. [Google Scholar] [CrossRef]
- Casanueva, M.O.; Burga, A.; Lehner, B. Fitness Trade-Offs and Environmentally Induced Mutation Buffering in Isogenic C. Elegans. Science 2012, 335, 82–85. [Google Scholar] [CrossRef]
- Karras, G.I.; Yi, S.; Sahni, N.; Fischer, M.; Xie, J.; Vidal, M.; D’Andrea, A.D.; Whitesell, L.; Lindquist, S. HSP90 Shapes the Consequences of Human Genetic Variation. Cell 2017, 168, 856–866.e12. [Google Scholar] [CrossRef] [Green Version]
- Hummel, B.; Hansen, E.C.; Yoveva, A.; Aprile-Garcia, F.; Hussong, R.; Sawarkar, R. The Evolutionary Capacitor HSP90 Buffers the Regulatory Effects of Mammalian Endogenous Retroviruses. Nat. Struct. Mol. Biol. 2017, 24, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Sollars, V.; Lu, X.; Xiao, L.; Wang, X.; Garfinkel, M.D.; Ruden, D.M. Evidence for an Epigenetic Mechanism by Which Hsp90 Acts as a Capacitor for Morphological Evolution. Nat. Genet 2003, 33, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Anckar, J.; Sistonen, L. Regulation of HSF1 Function in the Heat Stress Response: Implications in Aging and Disease. Annu. Rev. Biochem. 2011, 80, 1089–1115. [Google Scholar] [CrossRef]
- Zou, L.; Wu, D.; Zang, X.; Wang, Z.; Wu, Z.; Chen, D. Construction of a Germline-Specific RNAi Tool in C. Elegans. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, D.; Jones, L.M.; Good, S.; Miles, J.; Vijayabaskar, M.S.; Aston, R.; Smith, C.E.; Westhead, D.R.; van Oosten-Hawle, P. A PQM-1-Mediated Response Triggers Transcellular Chaperone Signaling and Regulates Organismal Proteostasis. Cell Rep. 2018, 23, 3905–3919. [Google Scholar] [CrossRef] [PubMed]
- Somogyvári, M.; Gecse, E.; Sőti, C. DAF-21/Hsp90 Is Required for C. Elegans Longevity by Ensuring DAF-16/FOXO Isoform A Function. Sci. Rep. 2018, 8, 12048. [Google Scholar] [CrossRef] [Green Version]
- Maman, M.; Marques, F.C.; Volovik, Y.; Dubnikov, T.; Bejerano-Sagie, M.; Cohen, E. A Neuronal GPCR Is Critical for the Induction of the Heat Shock Response in the Nematode C. Elegans. J. Neurosci. 2013, 33, 6102–6111. [Google Scholar] [CrossRef] [Green Version]
- Tatum, M.C.; Ooi, F.K.; Chikka, M.R.; Chauve, L.; Martinez-Velazquez, L.A.; Steinbusch, H.W.M.; Morimoto, R.I.; Prahlad, V. Neuronal Serotonin Release Triggers the Heat Shock Response in C. Elegans in the Absence of Temperature Increase. Curr. Biol. 2015, 25, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prahlad, V.; Cornelius, T.; Morimoto, R.I. Regulation of the Cellular Heat Shock Response in Caenorhabditis Elegans by Thermosensory Neurons. Science 2008, 320, 811–814. [Google Scholar] [CrossRef] [Green Version]
- Gildea, H.K.; Frankino, P.A.; Tronnes, S.U.; Pender, C.L.; Durieux, J.; Dishart, J.G.; Choi, H.O.; Hunter, T.D.; Cheung, S.S.; Frakes, A.E.; et al. Glia of C. Elegans Coordinate a Protective Organismal Heat Shock Response Independent of the Neuronal Thermosensory Circuit. Sci. Adv. 2022, 8, eabq3970. [Google Scholar] [CrossRef]
- Sugi, T.; Nishida, Y.; Mori, I. Regulation of Behavioral Plasticity by Systemic Temperature Signaling in Caenorhabditis Elegans. Nat. Neurosci. 2011, 14, 984–992. [Google Scholar] [CrossRef]
- Jing, R.; Duncan, C.B.; Duncan, S.A. A Small-Molecule Screen Reveals That HSP90β Promotes the Conversion of Induced Pluripotent Stem Cell-Derived Endoderm to a Hepatic Fate and Regulates HNF4A Turnover. Development 2017, 144, 1764–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chauve, L.; Phelps, G.; Brielmann, R.M.; Morimoto, R.I. E2F Coregulates an Essential HSF Developmental Program That Is Distinct from the Heat-Shock Response. Genes Dev. 2016, 30, 2062–2075. [Google Scholar] [CrossRef] [Green Version]
- Morton, E.A.; Lamitina, T. Caenorhabditis Elegans HSF-1 Is an Essential Nuclear Protein That Forms Stress Granule-like Structures Following Heat Shock. Aging Cell 2013, 12, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abane, R.; Mezger, V. Roles of Heat Shock Factors in Gametogenesis and Development. FEBS J. 2010, 277, 4150–4172. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.L.; Erdenebat, P.; Morphis, A.C.; Kumar, L.; Wang, L.; Chamera, T.; Georgescu, C.; Wren, J.D.; Li, J. Insulin/IGF-1 Signaling and Heat Stress Differentially Regulate HSF1 Activities in Germline Development. Cell. Rep. 2021, 36, 109623. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.; Oosten-Hawle, P. van Tissue-Specific RNAi Tools to Identify Components for Systemic Stress Signaling. JoVE (J. Vis. Exp. ) 2020, 159, e61357. [Google Scholar] [CrossRef]
- Miles, J.; Townend, S.; Smith, W.; Westhead, D.R.; van Oosten-Hawle, P. Transcellular Chaperone Signaling Is an Intercellular Stress-Response Distinct from the HSF-1 Mediated HSR. bioRxiv 2022, 2022.03.17.484707. [Google Scholar]
- van Oosten-Hawle, P.; Porter, R.S.; Morimoto, R.I. Regulation of Organismal Proteostasis by Transcellular Chaperone Signaling. Cell 2013, 153, 1366–1378. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, T.W.; Sylvester, S.L.; Sarge, K.D.; Morimoto, R.I.; Holbrook, N.J. Effects of Neurohormonal Stress and Aging on the Activation of Mammalian Heat Shock Factor 1. J. Biol. Chem. 1994, 269, 32272–32278. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.-F.; Jin, Z.-G.; Baas, A.S.; Daum, G.; Gygi, S.P.; Aebersold, R.; Berk, B.C. Purification and Identification of Secreted Oxidative Stress-Induced Factors from Vascular Smooth Muscle Cells *. J. Biol. Chem. 2000, 275, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, A.; O’Brien, K.; Guo, J.; Lincoln, V.; Kajiwara, C.; Chen, M.; Woodley, D.T.; Udono, H.; Li, W. Extracellular and Non-Chaperone Function of Heat Shock Protein-90α Is Required for Skin Wound Healing. J. Invest. Derm. 2018, 138, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sahu, D.; Tsen, F. Secreted Heat Shock Protein-90 (Hsp90) in Wound Healing and Cancer. Biochim. Et Biophys. Acta (BBA)—Mol. Cell Res. 2012, 1823, 730–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jay, D.; Luo, Y.; Li, W. Extracellular Heat Shock Protein-90 (EHsp90): Everything You Need to Know. Biomolecules 2022, 12, 911. [Google Scholar] [CrossRef] [PubMed]
- Janssens, G.E.; Lin, X.-X.; Millan-Ariño, L.; Kavšek, A.; Sen, I.; Seinstra, R.I.; Stroustrup, N.; Nollen, E.A.A.; Riedel, C.G. Transcriptomics-Based Screening Identifies Pharmacological Inhibition of Hsp90 as a Means to Defer Aging. Cell Rep. 2019, 27, 467–480.e6. [Google Scholar] [CrossRef] [Green Version]
- Hsu, A.-L.; Murphy, C.T.; Kenyon, C. Regulation of Aging and Age-Related Disease by DAF-16 and Heat-Shock Factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [Green Version]
- Douglas, P.M.; Baird, N.A.; Simic, M.S.; Uhlein, S.; McCormick, M.A.; Wolff, S.C.; Kennedy, B.K.; Dillin, A. Heterotypic Signals from Neural HSF-1 Separate Thermotolerance from Longevity. Cell Rep. 2015, 12, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morley, J.F.; Morimoto, R.I. Regulation of Longevity in Caenorhabditis Elegans by Heat Shock Factor and Molecular Chaperones. Mol. Biol. Cell 2004, 15, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Luo, J.; Jiang, P.; Li, Y.; Wang, Q.; Bai, Y.; Chen, J.; Wang, J.; Zhang, J. Inhibition of Embryonic HSP 90 Function Promotes Variation of Cold Tolerance in Zebrafish. Front. Genet. 2020, 11, 541944. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, M.; Sural, S.; Hsu, A.-L.; Antebi, A. Co-Chaperone P23 Regulates C. Elegans Lifespan in Response to Temperature. PLoS Genet 2015, 11, e1005023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, B.C.; Felts, S.J.; Toft, D.O.; Yamamoto, K.R. The P23 Molecular Chaperones Act at a Late Step in Intracellular Receptor Action to Differentially Affect Ligand Efficacies. Genes Dev. 2000, 14, 422–434. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Somogyvári, M.; Sőti, C. Hsp90 Stabilizes SIRT1 Orthologs in Mammalian Cells and C. Elegans. Int. J. Mol. Sci. 2018, 19, 3661. [Google Scholar] [CrossRef] [PubMed]
- Hubert, D.A.; Tornero, P.; Belkhadir, Y.; Krishna, P.; Takahashi, A.; Shirasu, K.; Dangl, J.L. Cytosolic HSP90 Associates with and Modulates the Arabidopsis RPM1 Disease Resistance Protein. EMBO J. 2003, 22, 5679–5689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangster, T.A.; Queitsch, C. The HSP90 Chaperone Complex, an Emerging Force in Plant Development and Phenotypic Plasticity. Curr. Opin. Plant Biol. 2005, 8, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Aballay, A. Heat-Shock Transcription Factor (HSF)-1 Pathway Required for Caenorhabditis Elegans Immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 13092–13097. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Aballay, A. Heat Shock and Genetic Activation of HSF-1 Enhance Immunity to Bacteria. Cell Cycle 2006, 5, 2443–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojda, I.; Kowalski, P. Galleria Mellonella Infected with Bacillus Thuringiensis Involves Hsp90. Open Life Sci. 2013, 8, 561–569. [Google Scholar] [CrossRef]
- Oura, J.; Tamura, Y.; Kamiguchi, K.; Kutomi, G.; Sahara, H.; Torigoe, T.; Himi, T.; Sato, N. Extracellular Heat Shock Protein 90 Plays a Role in Translocating Chaperoned Antigen from Endosome to Proteasome for Generating Antigenic Peptide to Be Cross-Presented by Dendritic Cells. Int. Immunol. 2011, 23, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Murshid, A.; Gong, J.; Calderwood, S.K. Heat Shock Protein 90 Mediates Efficient Antigen Cross Presentation through the Scavenger Receptor Expressed by Endothelial Cells-I. J. Immunol. 2010, 185, 2903–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, A.; Gong, J.; Calderwood, S.K. Hsp90–Peptide Complexes Stimulate Antigen Presentation through the Class II Pathway after Binding Scavenger Receptor SREC-I. Immunobiology 2014, 219, 924–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graner, M.W. Chapter Eight - HSP90 and Immune Modulation in Cancer. In Advances in Cancer Research; Hsp90 in Cancer: Beyond the Usual Suspects; Isaacs, J., Whitesell, L., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 129, pp. 191–224. [Google Scholar]
- Anderson, I.; Low, J.S.; Weston, S.; Weinberger, M.; Zhyvoloup, A.; Labokha, A.A.; Corazza, G.; Kitson, R.A.; Moody, C.J.; Marcello, A.; et al. Heat Shock Protein 90 Controls HIV-1 Reactivation from Latency. Proc. Natl. Acad. Sci. USA 2014, 111, E1528–E1537. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.; Calderwood, S.K.; Coates, A.R.M.; Cohen, I.; van Eden, W.; Lehner, T.; Pockley, A.G. Caught with Their PAMPs Down? The Extracellular Signalling Actions of Molecular Chaperones Are Not Due to Microbial Contaminants. Cell Stress Chaperones 2010, 15, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.-W.; et al. A Conserved P38 MAP Kinase Pathway in Caenorhabditis Elegans Innate Immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, B.; Karin, M. Mitogen-Activated Protein Kinase Cascades and Regulation of Gene Expression. Curr. Opin. Immunol. 1996, 8, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Banumathy, G.; Singh, V.; Pavithra, S.R.; Tatu, U. Heat Shock Protein 90 Function Is Essential for Plasmodium Falciparum Growth in Human Erythrocytes. J. Biol. Chem. 2003, 278, 18336–18345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angel, S.O.; Figueras, M.J.; Alomar, M.L.; Echeverria, P.C.; Deng, B. Toxoplasma Gondii Hsp90: Potential Roles in Essential Cellular Processes of the Parasite. Parasitology 2014, 141, 1138–1147. [Google Scholar] [CrossRef] [Green Version]
- Echeverria, P.C.; Matrajt, M.; Harb, O.S.; Zappia, M.P.; Costas, M.A.; Roos, D.S.; Dubremetz, J.F.; Angel, S.O. Toxoplasma Gondii Hsp90 Is a Potential Drug Target Whose Expression and Subcellular Localization Are Developmentally Regulated. J. Mol. Biol. 2005, 350, 723–734. [Google Scholar] [CrossRef]
- Corigliano, M.G.; Sander, V.A.; Sánchez López, E.F.; Ramos Duarte, V.A.; Mendoza Morales, L.F.; Angel, S.O.; Clemente, M. Heat Shock Proteins 90 KDa: Immunomodulators and Adjuvants in Vaccine Design Against Infectious Diseases. Front. Bioeng. Biotechnol. 2021, 8, 622186. [Google Scholar] [CrossRef]
- Gerges, N.Z.; Tran, I.C.; Backos, D.S.; Harrell, J.M.; Chinkers, M.; Pratt, W.B.; Esteban, J.A. Independent Functions of Hsp90 in Neurotransmitter Release and in the Continuous Synaptic Cycling of AMPA Receptors. J. Neurosci. 2004, 24, 4758–4766. [Google Scholar] [CrossRef] [Green Version]
- Karney-Grobe, S.; Russo, A.; Frey, E.; Milbrandt, J.; DiAntonio, A. HSP90 Is a Chaperone for DLK and Is Required for Axon Injury Signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E9899–E9908. [Google Scholar] [CrossRef] [Green Version]
- Hajdú, G.; Gecse, E.; Taisz, I.; Móra, I.; Sőti, C. Toxic Stress-Specific Cytoprotective Responses Regulate Learned Behavioral Decisions in C. Elegans. BMC Biol. 2021, 19, 26. [Google Scholar] [CrossRef] [PubMed]
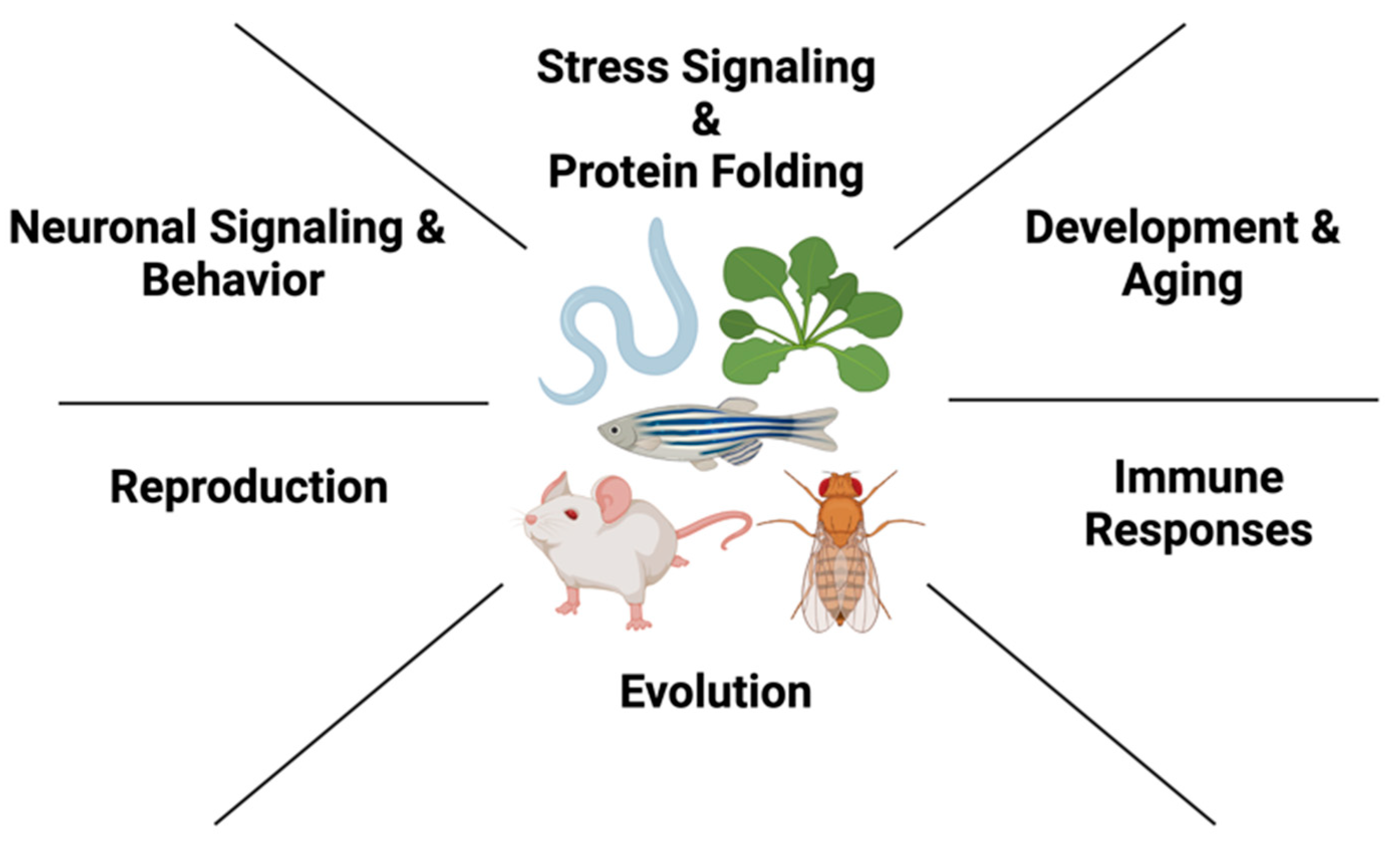
Figure 1.
Organismal roles of Hsp90 in different multicellular model systems. In metazoans, such as C. elegans, Mus musculus, D. melanogaster, Danio rerio and A. thaliana, Hsp90 acts in diverse biological processes to ensure organismal proteostasis.
Figure 1.
Organismal roles of Hsp90 in different multicellular model systems. In metazoans, such as C. elegans, Mus musculus, D. melanogaster, Danio rerio and A. thaliana, Hsp90 acts in diverse biological processes to ensure organismal proteostasis.

Figure 2.
Transcellular chaperone signaling pathways. (A) Overexpression of Hsp90 in the nervous system mediates upregulation of Hsp90 in muscle cells via PQM-1, CLEC-41 and glutamatergic neurotransmission. Overexpression of Hsp90 in the intestine relays the signal to upregulate Hsp90 in muscle cells via PQM-1 and ASP-12. The transcription factor regulating Hsp90 in muscle cells in response to TCS has not been determined. The organismal consequences are increased health span and reduced protein aggregation in the muscle tissue. (B) Knockdown of Hsp90 in the intestine relays the signal to muscle cells via the secreted lipases TXT-4 and TXT-8. There, TXT-1 signals to the transcription factor CEH-58 to induce Hsp70 expression, resulting in increased longevity and stress resilience. HSF-1 functions as a suppressor of this process.
Figure 2.
Transcellular chaperone signaling pathways. (A) Overexpression of Hsp90 in the nervous system mediates upregulation of Hsp90 in muscle cells via PQM-1, CLEC-41 and glutamatergic neurotransmission. Overexpression of Hsp90 in the intestine relays the signal to upregulate Hsp90 in muscle cells via PQM-1 and ASP-12. The transcription factor regulating Hsp90 in muscle cells in response to TCS has not been determined. The organismal consequences are increased health span and reduced protein aggregation in the muscle tissue. (B) Knockdown of Hsp90 in the intestine relays the signal to muscle cells via the secreted lipases TXT-4 and TXT-8. There, TXT-1 signals to the transcription factor CEH-58 to induce Hsp70 expression, resulting in increased longevity and stress resilience. HSF-1 functions as a suppressor of this process.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
van Oosten-Hawle, P. Organismal Roles of Hsp90. Biomolecules 2023, 13, 251. https://doi.org/10.3390/biom13020251
AMA Style
van Oosten-Hawle P. Organismal Roles of Hsp90. Biomolecules. 2023; 13(2):251. https://doi.org/10.3390/biom13020251
Chicago/Turabian Stylevan Oosten-Hawle, Patricija. 2023. "Organismal Roles of Hsp90" Biomolecules 13, no. 2: 251. https://doi.org/10.3390/biom13020251
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.