
Resilience and Vulnerability to Stress-Induced Anhedonia: Unveiling Brain Gene Expression and Mitochondrial Dynamics in a Mouse Chronic Stress Depression Model
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Flow
2.3. Chronic Stress Procedure and Determination of Anhedonia
2.4. Sucrose Preference Test
2.5. Tail Suspension Test
2.6. Evaluation of Coat Disintegration
2.7. Splash Test
2.8. Evaluation of Nest Building Performance
2.9. Marble T-Test
2.10. Y-Maze
2.11. Administration of DS
2.12. Culling and Brain Dissection
2.13. RNA Extraction and RT-PCR
2.14. Illumina Gene Expression Profiling and Ingenuity Pathway Analysis
2.15. Statistical Analysis
3. Results
3.1. Sucrose Preference Test
3.2. Tail Suspension Test
3.3. Marble T-Test
3.4. Coat Disintegration and Nest Building Scores
3.5. Splash Test
3.6. Y-Maze Learning
3.7. The mRNA Expression of the Inflammation Markers and Serotonergic System Proteins in Selected Brain Regions
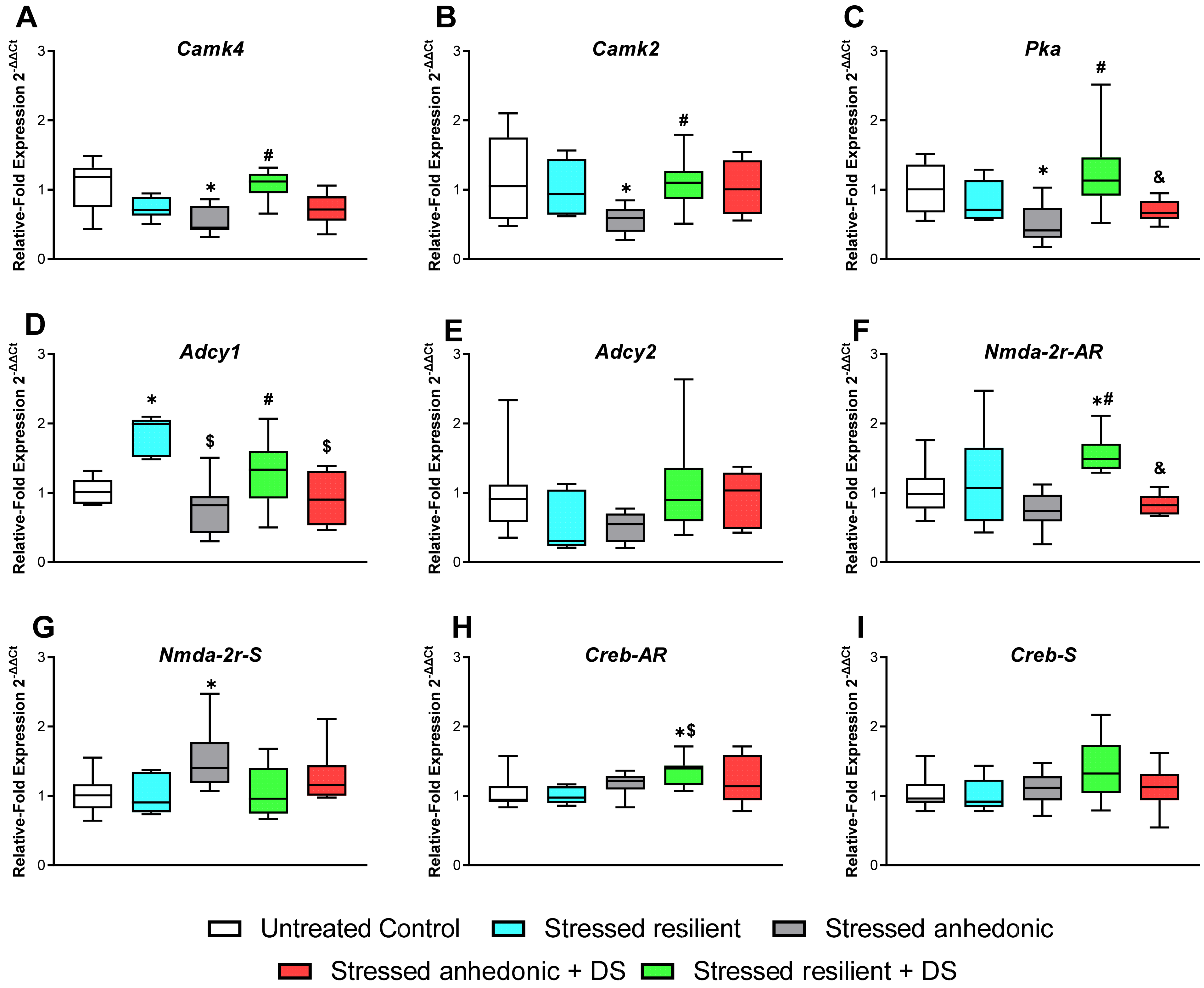
3.8. Hippocampal Expression of the Plasticity Markers
3.9. Illumina Study: Mitochondrial Oxidative Phosphorylation Gene Pathway Enrichment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paul, E.R.; Östman, L.; Heilig, M.; Mayberg, H.S.; Hamilton, J.P. Towards a Multilevel Model of Major Depression: Genes, Immuno-Metabolic Function, and Cortico-Striatal Signaling. Transl. Psychiatry 2023, 13, 171. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Baussan, Y.; Hebert-Chatelain, E. Connecting Dots between Mitochondrial Dysfunction and Depression. Biomolecules 2023, 13, 695. [Google Scholar] [CrossRef] [PubMed]
- Al Omran, A.J.; Watanabe, S.; Hong, E.C.; Skinner, S.G.; Zhang, M.; Zhang, J.; Shao, X.M.; Liang, J. Dihydromyricetin Ameliorates Social Isolation-Induced Anxiety by Modulating Mitochondrial Function, Antioxidant Enzymes, and BDNF. Neurobiol. Stress 2022, 21, 100499. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cui, C.; Xu, P.; Zhu, L.; Xue, H.; Chen, B.; Jiang, P. Targeting PDK2 Rescues Stress-Induced Impaired Brain Energy Metabolism. Mol. Psychiatry 2023. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Filiou, M.D.; Sandi, C. Anxiety and Brain Mitochondria: A Bidirectional Crosstalk. Trends Neurosci. 2019, 42, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Pomytkin, I.; Pinelis, V. Brain Insulin Resistance: Focus on Insulin Receptor-Mitochondria Interactions. Life 2021, 11, 262. [Google Scholar] [CrossRef]
- Fizíková, I.; Dragašek, J.; Račay, P. Mitochondrial Dysfunction, Altered Mitochondrial Oxygen, and Energy Metabolism Associated with the Pathogenesis of Schizophrenia. Int. J. Mol. Sci. 2023, 24, 7991. [Google Scholar] [CrossRef]
- Maiese, K. Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System. Biomolecules 2023, 13, 816. [Google Scholar] [CrossRef]
- Shamabadi, A.; Karimi, H.; Cattarinussi, G.; Moghaddam, H.S.; Akhondzadeh, S.; Sambataro, F.; Schiena, G.; Delvecchio, G. Neuroimaging Correlates of Treatment Response to Transcranial Magnetic Stimulation in Bipolar Depression: A Systematic Review. Brain Sci. 2023, 13, 801. [Google Scholar] [CrossRef]
- You, T.-Y.; Dong, Q.; Cui, M. Emerging Links between Cerebral Blood Flow Regulation and Cognitive Decline: A Role for Brain Microvascular Pericytes. Aging Dis. 2022, 14, 1276–1291. [Google Scholar] [CrossRef]
- Fattal, O.; Link, J.; Quinn, K.; Cohen, B.H.; Franco, K. Psychiatric Comorbidity in 36 Adults with Mitochondrial Cytopathies. CNS Spectr. 2007, 12, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.M.; Mervis, R.F.; Niu, S.L.; Salem, N.; Witters, L.A.; Tseng, V.; Reinhardt, R.; Bondy, C.A. Insulin-like Growth Factor 1 Is Essential for Normal Dendritic Growth. J. Neurosci. Res. 2003, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.L.; Cline, H.T. Insulin Receptor Signaling in the Development of Neuronal Structure and Function. Neural Dev. 2010, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Mielke, J.G.; Taghibiglou, C.; Wang, Y.T. Endogenous Insulin Signaling Protects Cultured Neurons from Oxygen-Glucose Deprivation-Induced Cell Death. Neuroscience 2006, 143, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yao, H.; Douglas, R.M.; Gu, X.Q.; Wang, J.; Haddad, G.G. Insulin/PI3K Signaling Protects Dentate Neurons from Oxygen-Glucose Deprivation in Organotypic Slice Cultures. J. Neurochem. 2010, 112, 377–388. [Google Scholar] [CrossRef] [PubMed]
- McNay, E.C.; Recknagel, A.K. Brain Insulin Signaling: A Key Component of Cognitive Processes and a Potential Basis for Cognitive Impairment in Type 2 Diabetes. Neurobiol. Learn. Mem. 2011, 96, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, S.; Heni, M.; Hallschmid, M.; Fritsche, A.; Preissl, H.; Häring, H.U. Brain Insulin Resistance at the Crossroads of Metabolic and Cognitive Disorders in Humans. Physiol. Rev. 2016, 96, 1169–1209. [Google Scholar] [CrossRef]
- Persiyantseva, N.A.; Storozhevykh, T.P.; Senilova, Y.E.; Gorbacheva, L.R.; Pinelis, V.G.; Pomytkin, I.A. Mitochondrial H2O2 as an Enable Signal for Triggering Autophosphorylation of Insulin Receptor in Neurons. J. Mol. Signal. 2013, 8, 11. [Google Scholar] [CrossRef]
- Morella, I.M.; Brambilla, R.; Morè, L. Emerging Roles of Brain Metabolism in Cognitive Impairment and Neuropsychiatric Disorders. Neurosci. Biobehav. Rev. 2022, 142, 104892. [Google Scholar] [CrossRef]
- Pomytkin, I.; Costa-Nunes, J.P.; Kasatkin, V.; Veniaminova, E.; Demchenko, A.; Lyundup, A.; Lesch, K.-P.; Ponomarev, E.D.; Strekalova, T. Insulin Receptor in the Brain: Mechanisms of Activation and the Role in the CNS Pathology and Treatment. CNS Neurosci. Ther. 2018, 24, 763–774. [Google Scholar] [CrossRef]
- Chigogora, S.; Zaninotto, P.; Kivimaki, M.; Steptoe, A.; Batty, G.D. Insulin-like Growth Factor 1 and Risk of Depression in Older People: The English Longitudinal Study of Ageing. Transl. Psychiatry 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Bou Khalil, R. Is Insulin Growth Factor-1 the Future for Treating Autism Spectrum Disorder and/or Schizophrenia? Med. Hypotheses 2017, 99, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Łastowska, M.; Al-Afghani, H.; Al-Balool, H.H.; Sheth, H.; Mercer, E.; Coxhead, J.M.; Redfern, C.P.F.; Peters, H.; Burt, A.D.; Santibanez-Koref, M.; et al. Identification of a Neuronal Transcription Factor Network Involved in Medulloblastoma Development. Acta Neuropathol. Commun. 2013, 1, 35. [Google Scholar] [CrossRef] [PubMed]
- Pini, G.; Congiu, L.; Benincasa, A.; DiMarco, P.; Bigoni, S.; Dyer, A.H.; Mortimer, N.; Della-Chiesa, A.; O’Leary, S.; McNamara, R.; et al. Illness Severity, Social and Cognitive Ability, and EEG Analysis of Ten Patients with Rett Syndrome Treated with Mecasermin (Recombinant Human IGF-1). Autism Res. Treat. 2016, 2016, 5073078. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, L.; Enam, S.; Lassak, A.; Wang, J.Y.; Croul, S.; Khalili, K.; Reiss, K. Insulin-like Growth Factor I Receptor Activity in Human Medulloblastomas. Clin. Cancer Res. 2002, 8, 1822–1830. [Google Scholar]
- Park, H.J.; Kim, S.K.; Kang, W.S.; Park, J.K.; Kim, Y.J.; Nam, M.; Kim, J.W.; Chung, J.H. Association between IRS1 Gene Polymorphism and Autism Spectrum Disorder: A Pilot Case-Control Study in Korean Males. Int. J. Mol. Sci. 2016, 17, 1227. [Google Scholar] [CrossRef]
- Du, M.; Wu, C.; Yu, R.; Cheng, Y.; Tang, Z.; Wu, B.; Fu, J.; Tan, W.; Zhou, Q.; Zhu, Z.; et al. A Novel Circular RNA, CircIgfbp2, Links Neural Plasticity and Anxiety through Targeting Mitochondrial Dysfunction and Oxidative Stress-Induced Synapse Dysfunction after Traumatic Brain Injury. Mol. Psychiatry 2022, 27, 4575–4589. [Google Scholar] [CrossRef]
- Pomytkin, I.A. H2O2 Signalling Pathway: A Possible Bridge between Insulin Receptor and Mitochondria. Curr. Neuropharmacol. 2012, 10, 311–320. [Google Scholar] [CrossRef]
- Nibber, A.; Singh, H.; Burnet, P.; Lennox, B.; Minichino, A. Investigating the Pro-Cognitive and Anti-Depressant Efficacy of Metformin: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. J. Affect. Disord. 2022, 310, 52–59. [Google Scholar] [CrossRef]
- Toba-Oluboka, T.; Vochosková, K.; Hajek, T. Are the Antidepressant Effects of Insulin-Sensitizing Medications Related to Improvements in Metabolic Markers? Transl. Psychiatry 2022, 12, 469. [Google Scholar] [CrossRef]
- Storozhevykh, T.P.; Senilova, Y.E.; Persiyantseva, N.A.; Pinelis, V.G.; Pomytkin, I.A. Mitochondrial Respiratory Chain Is Involved in Insulin-Stimulated Hydrogen Peroxide Production and Plays an Integral Role in Insulin Receptor Autophosphorylation in Neurons. BMC Neurosci. 2007, 8, 84. [Google Scholar] [CrossRef]
- Storozheva, Z.I.; Proshin, A.T.; Sherstnev, V.V.; Storozhevykh, T.P.; Senilova, Y.E.; Persiyantseva, N.A.; Pinelis, V.G.; Semenova, N.A.; Zakharova, E.I.; Pomytkin, I.A. Dicholine Salt of Succinic Acid, a Neuronal Insulin Sensitizer, Ameliorates Cognitive Deficits in Rodent Models of Normal Aging, Chronic Cerebral Hypoperfusion, and Beta-Amyloid Peptide-(25-35)-Induced Amnesia. BMC Pharmacol. 2008, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Hirata, A.; Yamaguchi, H.; Jimbu, Y.; Tominaga, M. Pioglitazone Reduces Atherogenic Outcomes in Type 2 Diabetic Patients. J. Atheroscler. Thromb. 2008, 15, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Patzer, A.; Herdegen, T.; Gohlke, P.; Culman, J. Activation of Cerebral Peroxisome Proliferator-Activated Receptors Gamma Promotes Neuroprotection by Attenuation of Neuronal Cyclooxygenase-2 Overexpression after Focal Cerebral Ischemia in Rats. FASEB J. 2006, 20, 1162–1175. [Google Scholar] [CrossRef] [PubMed]
- Strum, J.C.; Shehee, R.; Virley, D.; Richardson, J.; Mattie, M.; Selley, P.; Ghosh, S.; Nock, C.; Saunders, A.; Roses, A. Rosiglitazone Induces Mitochondrial Biogenesis in Mouse Brain. J. Alzheimers Dis. 2007, 11, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Eissa Ahmed, A.A.; Al-Rasheed, N.M.; Al-Rasheed, N.M. Antidepressant-like Effects of Rosiglitazone, a PPARγ Agonist, in the Rat Forced Swim and Mouse Tail Suspension Tests. Behav. Pharmacol. 2009, 20, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Malhotra, S.; Pandhi, P.; Kaur, I.; Dogra, S. Efficacy and Safety of Combination Acitretin and Pioglitazone Therapy in Patients with Moderate to Severe Chronic Plaque-Type Psoriasis: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Arch. Dermatol. 2009, 145, 387–393. [Google Scholar] [CrossRef]
- Rasgon, N.L.; Kenna, H.A.; Williams, K.E.; Powers, B.; Wroolie, T.; Schatzberg, A.F. Rosiglitazone Add-on in Treatment of Depressed Patients with Insulin Resistance: A Pilot Study. Sci. World J. 2010, 10, 321–328. [Google Scholar] [CrossRef]
- Kemp, D.E.; Ismail-Beigi, F.; Ganocy, S.J.; Conroy, C.; Gao, K.; Obral, S.; Fein, E.; Findling, R.L.; Calabrese, J.R. Use of Insulin Sensitizers for the Treatment of Major Depressive Disorder: A Pilot Study of Pioglitazone for Major Depression Accompanied by Abdominal Obesity. J. Affect. Disord. 2012, 136, 1164–1173. [Google Scholar] [CrossRef]
- Saubermann, L.J.; Nakajima, A.; Wada, K.; Zhao, S.; Terauchi, Y.; Kadowaki, T.; Aburatani, H.; Matsuhashi, N.; Nagai, R.; Blumberg, R.S. Peroxisome Proliferator-Activated Receptor Gamma Agonist Ligands Stimulate a Th2 Cytokine Response and Prevent Acute Colitis. Inflamm. Bowel Dis. 2002, 8, 330–339. [Google Scholar] [CrossRef]
- Ali, S.; Stone, M.A.; Peters, J.L.; Davies, M.J.; Khunti, K. The Prevalence of Co-Morbid Depression in Adults with Type 2 Diabetes: A Systematic Review and Meta-Analysis. Diabet. Med. 2006, 23, 1165–1173. [Google Scholar] [CrossRef]
- Asghar, S.; Hussain, A.; Ali, S.M.K.; Khan, A.K.A.; Magnusson, A. Prevalence of Depression and Diabetes: A Population-Based Study from Rural Bangladesh. Diabet. Med. 2007, 24, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Sadaghiani, M.; Javadi-Paydar, M.; Gharedaghi, M.H.; Zandieh, A.; Heydarpour, P.; Yousefzadeh-fard, Y.; Dehpour, A.R. NMDA Receptor Involvement in Antidepressant-like Effect of Pioglitazone in the Forced Swimming Test in Mice. Psychopharmacology 2012, 223, 345–355. [Google Scholar] [CrossRef]
- Sharma, A.N.; Elased, K.M.; Lucot, J.B. Rosiglitazone Treatment Reversed Depression- but Not Psychosis-like Behavior of Db/Db Diabetic Mice. J. Psychopharmacol. 2012, 26, 724–732. [Google Scholar] [CrossRef]
- Costa-Nunes, J.P.; Cline, B.H.; Araújo-Correia, M.; Valencą, A.; Markova, N.; Dolgov, O.; Kubatiev, A.; Yeritsyan, N.; Steinbusch, H.W.M.; Strekalova, T. Animal Models of Depression and Drug Delivery with Food as an Effective Dosing Method: Evidences from Studies with Celecoxib and Dicholine Succinate. Biomed. Res. Int. 2015, 2015, 596126. [Google Scholar] [CrossRef] [PubMed]
- Cline, B.H.; Steinbusch, H.W.M.; Malin, D.; Revishchin, A.V.; Pavlova, G.V.; Cespuglio, R.; Strekalova, T. The Neuronal Insulin Sensitizer Dicholine Succinate Reduces Stress-Induced Depressive Traits and Memory Deficit: Possible Role of Insulin-like Growth Factor 2. BMC Neurosci. 2012, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Cline, B.H.; Costa-Nunes, J.P.; Cespuglio, R.; Markova, N.; Santos, A.I.; Bukhman, Y.V.; Kubatiev, A.; Steinbusch, H.W.M.; Lesch, K.-P.; Strekalova, T. Dicholine Succinate, the Neuronal Insulin Sensitizer, Normalizes Behavior, REM Sleep, Hippocampal PGSK3 Beta and MRNAs of NMDA Receptor Subunits in Mouse Models of Depression. Front. Behav. Neurosci. 2015, 9, 37. [Google Scholar] [CrossRef]
- Strekalova, T.; Costa-Nunes, J.P.; Veniaminova, E.; Kubatiev, A.; Lesch, K.-P.; Chekhonin, V.P.; Evans, M.C.; Steinbusch, H.W.M. Insulin Receptor Sensitizer, Dicholine Succinate, Prevents Both Toll-like Receptor 4 (TLR4) Upregulation and Affective Changes Induced by a High-Cholesterol Diet in Mice. J. Affect. Disord. 2016, 196, 109–116. [Google Scholar] [CrossRef]
- Shomaker, L.B.; Tanofsky-Kraff, M.; Young-Hyman, D.; Han, J.C.; Yanoff, L.B.; Brady, S.M.; Yanovski, S.Z.; Yanovski, J.A. Psychological Symptoms and Insulin Sensitivity in Adolescents. Pediatr. Diabetes 2010, 11, 417–423. [Google Scholar] [CrossRef]
- Al-Harbi, K.S. Treatment-Resistant Depression: Therapeutic Trends, Challenges, and Future Directions. Patient Prefer. Adherence 2012, 6, 369. [Google Scholar] [CrossRef]
- Kasahara, T.; Kato, T. What Can Mitochondrial DNA Analysis Tell Us About Mood Disorders? Biol. Psychiatry 2018, 83, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Petschner, P.; Gonda, X.; Baksa, D.; Eszlari, N.; Trivaks, M.; Juhasz, G.; Bagdy, G. Genes Linking Mitochondrial Function, Cognitive Impairment and Depression Are Associated with Endophenotypes Serving Precision Medicine. Neuroscience 2018, 370, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Cathomas, F.; Murrough, J.W.; Nestler, E.J.; Han, M.H.; Russo, S.J. Neurobiology of Resilience: Interface Between Mind and Body. Biol. Psychiatry 2019, 86, 410–420. [Google Scholar] [CrossRef]
- Dudek, K.A.; Dion-Albert, L.; Kaufmann, F.N.; Tuck, E.; Lebel, M.; Menard, C. Neurobiology of Resilience in Depression: Immune and Vascular Insights from Human and Animal Studies. Eur. J. Neurosci. 2021, 53, 183–221. [Google Scholar] [CrossRef] [PubMed]
- Verhallen, A.M.; Alonso-Martínez, S.; Renken, R.J.; Marsman, J.B.C.; ter Horst, G.J. Depressive Symptom Trajectory Following Romantic Relationship Breakup and Effects of Rumination, Neuroticism and Cognitive Control. Stress Health 2022, 38, 653–665. [Google Scholar] [CrossRef]
- Isella, C.; Gasparini, A.; Lucca, G.; Ielmini, M.; Caselli, I.; Poloni, N.; Dajelli Ermolli, C.; Caravati, F.; Castiglioni, B.; De Ponti, R.; et al. Resilience, Cardiological Outcome, and Their Correlations with Anxious-Depressive Symptoms and Quality of Life in Patients with an Implantable Cardioverter Defibrillator. Front. Psychiatry 2021, 12, 763726. [Google Scholar] [CrossRef]
- Strekalova, T.; Liu, Y.; Kiselev, D.; Khairuddin, S.; Chiu, J.L.Y.; Lam, J.; Chan, Y.-S.; Pavlov, D.; Proshin, A.; Lesch, K.-P.; et al. Chronic Mild Stress Paradigm as a Rat Model of Depression: Facts, Artifacts, and Future Perspectives. Psychopharmacology 2022, 239, 663–693. [Google Scholar] [CrossRef]
- Weger, M.; Alpern, D.; Cherix, A.; Ghosal, S.; Grosse, J.; Russeil, J.; Gruetter, R.; de Kloet, E.R.; Deplancke, B.; Sandi, C. Mitochondrial Gene Signature in the Prefrontal Cortex for Differential Susceptibility to Chronic Stress. Sci. Rep. 2020, 10, 18308. [Google Scholar] [CrossRef]
- Levone, B.R.; Cryan, J.F.; O’Leary, O.F. Role of Adult Hippocampal Neurogenesis in Stress Resilience. Neurobiol. Stress 2014, 1, 147–155. [Google Scholar] [CrossRef]
- Tang, M.; Huang, H.; Li, S.; Zhou, M.; Liu, Z.; Huang, R.; Liao, W.; Xie, P.; Zhou, J. Hippocampal Proteomic Changes of Susceptibility and Resilience to Depression or Anxiety in a Rat Model of Chronic Mild Stress. Transl. Psychiatry 2019, 9, 260. [Google Scholar] [CrossRef]
- Albrecht, A.; Segal, M.; Stork, O. Allostatic Gene Regulation of Inhibitory Synaptic Factors in the Rat Ventral Hippocampus in a Juvenile/Adult Stress Model of Psychopathology. Eur. J. Neurosci. 2022, 55, 2142–2153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Hong, X.; Li, X.; Meshul, C.K.; Moore, C.; Yang, Y.; Han, Y.; Li, W.G.; Qi, X.; et al. NG2 Glia-Derived GABA Release Tunes Inhibitory Synapses and Contributes to Stress-Induced Anxiety. Nat. Commun. 2021, 12, 5740. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Murrough, J.W.; Han, M.H.; Charney, D.S.; Nestler, E.J. Neurobiology of Resilience. Nat. Neurosci. 2012, 15, 1475–1484. [Google Scholar] [CrossRef]
- Dantzer, R.; Cohen, S.; Russo, S.J.; Dinan, T.G. Resilience and Immunity. Brain Behav. Immun. 2018, 74, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.F. Letter: Pathophysiology of Depressive Syndromes. Biol. Psychiatry 1974, 8, 119–120. [Google Scholar] [PubMed]
- Hamilton, M. Development of a Rating Scale for Primary Depressive Illness. Br. J. Social. Clin. Psychol. 1967, 6, 278–296. [Google Scholar] [CrossRef]
- Willner, P.; Belzung, C. Treatment-Resistant Depression: Are Animal Models of Depression Fit for Purpose? Psychopharmacology 2015, 232, 3473–3495. [Google Scholar] [CrossRef]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of Sucrose Preference by Chronic Unpredictable Mild Stress, and Its Restoration by a Tricyclic Antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef]
- Strekalova, T.; Steinbusch, H.W.M. Measuring Behavior in Mice with Chronic Stress Depression Paradigm. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 348–361. [Google Scholar] [CrossRef]
- Gorinski, N.; Bijata, M.; Prasad, S.; Wirth, A.; Abdel Galil, D.; Zeug, A.; Bazovkina, D.; Kondaurova, E.; Kulikova, E.; Ilchibaeva, T.; et al. Attenuated Palmitoylation of Serotonin Receptor 5-HT1A Affects Receptor Function and Contributes to Depression-like Behaviors. Nat. Commun. 2019, 10, 3924. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.A.; Gass, P. Stress-Induced Anhedonia in Mice Is Associated with Deficits in Forced Swimming and Exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef]
- Strekalova, T.; Couch, Y.; Kholod, N.; Boyks, M.; Malin, D.; Leprince, P.; Steinbusch, H.M.W. Update in the Methodology of the Chronic Stress Paradigm: Internal Control Matters. Behav. Brain Funct. 2011, 7, 9. [Google Scholar] [CrossRef]
- Strekalova, T.; Gorenkova, N.; Schunk, E.; Dolgov, O.; Bartsch, D. Selective Effects of Citalopram in a Mouse Model of Stress-Induced Anhedonia with a Control for Chronic Stress. Behav. Pharmacol. 2006, 17, 271–287. [Google Scholar] [CrossRef]
- Malatynska, E.; Steinbusch, H.W.M.; Redkozubova, O.; Bolkunov, A.; Kubatiev, A.; Yeritsyan, N.B.; Vignisse, J.; Bachurin, S.; Strekalova, T. Anhedonic-like Traits and Lack of Affective Deficits in 18-Month-Old C57BL/6 Mice: Implications for Modeling Elderly Depression. Exp. Gerontol. 2012, 47, 552–564. [Google Scholar] [CrossRef]
- Vignisse, J.; Steinbusch, H.W.M.; Bolkunov, A.; Nunes, J.; Santos, A.I.; Grandfils, C.; Bachurin, S.; Strekalova, T. Dimebon Enhances Hippocampus-Dependent Learning in Both Appetitive and Inhibitory Memory Tasks in Mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Vignisse, J.; Sambon, M.; Gorlova, A.; Pavlov, D.; Caron, N.; Malgrange, B.; Shevtsova, E.; Svistunov, A.; Anthony, D.C.; Markova, N.; et al. Thiamine and Benfotiamine Prevent Stress-Induced Suppression of Hippocampal Neurogenesis in Mice Exposed to Predation without Affecting Brain Thiamine Diphosphate Levels. Mol. Cell. Neurosci. 2017, 82, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Trofimov, A.; Markova, N.; Nikolenko, V.; Steinbusch, H.W.; Chekhonin, V.; Schroeter, C.; Lesch, K.P.; Anthony, D.C.; Strekalova, T. Low-Dose Lipopolysaccharide (LPS) Inhibits Aggressive and Augments Depressive Behaviours in a Chronic Mild Stress Model in Mice. J. Neuroinflamm. 2016, 13, 108. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Anthony, D.C.; Dolgov, O.; Revischin, A.; Festoff, B.; Santos, A.I.; Steinbusch, H.W.; Strekalova, T. Microglial Activation, Increased TNF and SERT Expression in the Prefrontal Cortex Define Stress-Altered Behaviour in Mice Susceptible to Anhedonia. Brain Behav. Immun. 2013, 29, 136–146. [Google Scholar] [CrossRef]
- Costa-Nunes, J.P.; Gorlova, A.; Pavlov, D.; Cespuglio, R.; Gorovaya, A.; Proshin, A.; Umriukhin, A.; Ponomarev, E.D.; Kalueff, A.V.; Strekalova, T.; et al. Ultrasound Stress Compromises the Correlates of Emotional-like States and Brain AMPAR Expression in Mice: Effects of Antioxidant and Anti-Inflammatory Herbal Treatment. Stress 2020, 23, 481–495. [Google Scholar] [CrossRef]
- Li, X.Y.; Qi, W.W.; Zhang, Y.X.; Jiang, S.Y.; Yang, B.; Xiong, L.; Tong, J.C. Helicid Ameliorates Learning and Cognitive Ability and Activities CAMP/PKA/CREB Signaling in Chronic Unpredictable Mild Stress Rats. Biol. Pharm. Bull. 2019, 42, 1146–1154. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, J.; Li, L.; Qin, Y.; Wei, Y.; Pan, S.; Jiang, Y.; Chen, J.; Xie, Y. PKA-CREB-BDNF Signaling Pathway Mediates Propofol-Induced Long-Term Learning and Memory Impairment in Hippocampus of Rats. Brain Res. 2018, 1691, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Chen, Y.; Shen, Q.; Guo, X.; Zhang, Y.; Chen, G. Neural Plasticity Associated with Hippocampal PKA-CREB and NMDA Signaling Is Involved in the Antidepressant Effect of Repeated Low Dose of Yueju Pill on Chronic Mouse Model of Learned Helplessness. Neural Plast. 2017, 2017, 9160515. [Google Scholar] [CrossRef] [PubMed]
- Giese, K.P. The Role of CaMKII Autophosphorylation for NMDA Receptor-Dependent Synaptic Potentiation. Neuropharmacology 2021, 193, 108616. [Google Scholar] [CrossRef]
- Sałaciak, K.; Koszałka, A.; Żmudzka, E.; Pytka, K. The Calcium/Calmodulin-Dependent Kinases II and IV as Therapeutic Targets in Neurodegenerative and Neuropsychiatric Disorders. Int. J. Mol. Sci. 2021, 22, 4307. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yang, L.; Wei, W. Role of Fto on CaMKII/CREB Signaling Pathway of Hippocampus in Depressive-like Behaviors Induced by Chronic Restraint Stress Mice. Behav. Brain Res. 2021, 406, 113227. [Google Scholar] [CrossRef]
- Yang, M.; Ding, Q.; Zhang, M.; Moon, C.; Wang, H. Forebrain Overexpression of Type 1 Adenylyl Cyclase Promotes Molecular Stability and Behavioral Resilience to Physical Stress. Neurobiol. Stress 2020, 13, 100237. [Google Scholar] [CrossRef]
- Cline, B.H.; Anthony, D.C.; Lysko, A.; Dolgov, O.; Anokhin, K.; Schroeter, C.; Malin, D.; Kubatiev, A.; Steinbusch, H.W.; Lesch, K.P.; et al. Lasting Downregulation of the Lipid Peroxidation Enzymes in the Prefrontal Cortex of Mice Susceptible to Stress-Induced Anhedonia. Behav. Brain Res. 2015, 276, 118–129. [Google Scholar] [CrossRef]
- Strekalova, T.; Moskvin, O.; Jain, A.Y.; Gorbunov, N.; Gorlova, A.; Sadovnik, D.; Umriukhin, A.; Cespuglio, R.; Yu, W.S.; Tse, A.C.K.; et al. Molecular signature of excessive female aggression: Study of stressed mice with genetic inactivation of neuronal serotonin synthesis. J. Neural Transm. 2023, 130, 1113–1132. [Google Scholar] [CrossRef]
- Pothion, S.; Bizot, J.-C.; Trovero, F.; Belzung, C. Strain Differences in Sucrose Preference and in the Consequences of Unpredictable Chronic Mild Stress. Behav. Brain Res. 2004, 155, 135–146. [Google Scholar] [CrossRef]
- Veniaminova, E.; Cespuglio, R.; Cheung, C.W.; Umriukhin, A.; Markova, N.; Shevtsova, E.; Lesch, K.-P.; Anthony, D.C.; Strekalova, T. Autism-Like Behaviours and Memory Deficits Result from a Western Diet in Mice. Neural Plast. 2017, 2017, 9498247. [Google Scholar] [CrossRef]
- Veniaminova, E.; Cespuglio, R.; Chernukha, I.; Schmitt-Boehrer, A.G.; Morozov, S.; Kalueff, A.V.; Kuznetsova, O.; Anthony, D.C.; Lesch, K.P.; Strekalova, T. Metabolic, Molecular, and Behavioral Effects of Western Diet in Serotonin Transporter-Deficient Mice: Rescue by Heterozygosity? Front. Neurosci. 2020, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- de Munter, J.; Babaevskaya, D.; Wolters, E.C.; Pavlov, D.; Lysikova, E.; Kalueff, A.V.; Gorlova, A.; Oplatchikova, M.; Pomytkin, I.A.; Proshin, A.; et al. Molecular and Behavioural Abnormalities in the FUS-Tg Mice Mimic Frontotemporal Lobar Degeneration: Effects of Old and New Anti-Inflammatory Therapies. J. Cell. Mol. Med. 2020, 24, 10251–10257. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, D.; Bettendorff, L.; Gorlova, A.; Olkhovik, A.; Kalueff, A.V.; Ponomarev, E.D.; Inozemtsev, A.; Chekhonin, V.; Lesch, K.P.; Anthony, D.C.; et al. Neuroinflammation and Aberrant Hippocampal Plasticity in a Mouse Model of Emotional Stress Evoked by Exposure to Ultrasound of Alternating Frequencies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 90, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Lu, P.; Zeng, X.; Jin, S.; Chen, X. Study on the Mechanism for SIRT1 during the Process of Exercise Improving Depression. Brain Sci. 2023, 13, 719. [Google Scholar] [CrossRef] [PubMed]
- Carreras, I.; Jung, Y.; Lopez-Benitez, J.; Tognoni, C.M.; Dedeoglu, A. Fingolimod Mitigates Memory Loss in a Mouse Model of Gulf War Illness amid Decreasing the Activation of Microglia, Protein Kinase R, and NFκB. Neurotoxicology 2023, 96, 197–206. [Google Scholar] [CrossRef]
- Pomytkin, I.A.; Cline, B.H.; Anthony, D.C.; Steinbusch, H.W.; Lesch, K.P.; Strekalova, T. Endotoxaemia Resulting from Decreased Serotonin Tranporter (5-HTT) Function: A Reciprocal Risk Factor for Depression and Insulin Resistance? Behav. Brain Res. 2015, 276, 111–117. [Google Scholar] [CrossRef]
- Durand-De Cuttoli, R.; Martínez-Rivera, F.J.; Li, L.; Minier-Toribio, A.; Holt, L.M.; Cathomas, F.; Yasmin, F.; Elhassa, S.O.; Shaikh, J.F.; Ahmed, S.; et al. Distinct Forms of Regret Linked to Resilience versus Susceptibility to Stress Are Regulated by Region-Specific CREB Function in Mice. Sci. Adv. 2022, 8, eadd5579. [Google Scholar] [CrossRef]
- Rezin, G.T.; Cardoso, M.R.; Gonçalves, C.L.; Scaini, G.; Fraga, D.B.; Riegel, R.E.; Comim, C.M.; Quevedo, J.; Streck, E.L. Inhibition of Mitochondrial Respiratory Chain in Brain of Rats Subjected to an Experimental Model of Depression. Neurochem. Int. 2008, 53, 395–400. [Google Scholar] [CrossRef]
- Gong, Y.; Chai, Y.; Ding, J.H.; Sun, X.L.; Hu, G. Chronic Mild Stress Damages Mitochondrial Ultrastructure and Function in Mouse Brain. Neurosci. Lett. 2011, 488, 76–80. [Google Scholar] [CrossRef]
- Ghosh, S.; Castillo, E.; Frias, E.S.; Swanson, R.A. Bioenergetic Regulation of Microglia. Glia 2018, 66, 1200–1212. [Google Scholar] [CrossRef]
- Carlezon, W.A.; Duman, R.S.; Nestler, E.J. The Many Faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef]
- Mons, N.; Guillou, J.L.; Decorte, L.; Jaffard, R. Spatial Learning Induces Differential Changes in Calcium/Calmodulin-Stimulated (ACI) and Calcium-Insensitive (ACII) Adenylyl Cyclases in the Mouse Hippocampus. Neurobiol. Learn. Mem. 2003, 79, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ferguson, G.D.; Pineda, V.V.; Cundiff, P.E.; Storm, D.R. Overexpression of Type-1 Adenylyl Cyclase in Mouse Forebrain Enhances Recognition Memory and LTP. Nat. Neurosci. 2004, 7, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.V.; Woo, N.H. Regulation of Hippocampal Synaptic Plasticity by Cyclic AMP-Dependent Protein Kinases. Prog. Neurobiol. 2003, 71, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.; Rudolph-Correia, S.; Sheng, M. Developmentally Regulated NMDA Receptor-Dependent Dephosphorylation of CAMP Response Element-Binding Protein (CREB) in Hippocampal Neurons. J. Neurosci. 2000, 20, 3529–3536. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Hirose, E.; Ishida, T.; Hori, A.; Nagai, T.; Kobayashi, Y.; Kiyohara, H.; Oikawa, T.; Hanawa, T.; Odaguchi, H. Kososan, a Kampo Medicine, prevents a Social Avoidance Behavior and Attenuates Neuroinflammation in Socially Defeated Mice. J. Neuroinflamm. 2017, 14, 98. [Google Scholar] [CrossRef]
- Kamimura, Y.; Kuwagaki, E.; Hamano, S.; Kobayashi, M.; Yamada, Y.; Takahata, Y.; Yoshimoto, W.; Morimoto, H.; Yasukawa, T.; Uozumi, Y.; et al. Reproducible Induction of Depressive-like Behavior in C57BL/6J Mice Exposed to Chronic Social Defeat Stress with a Modified Sensory Contact Protocol. Life Sci. 2021, 282, 119821. [Google Scholar] [CrossRef] [PubMed]
- Strekalova, T.; Evans, M.; Chernopiatko, A.; Couch, Y.; Costa-Nunes, J.; Cespuglio, R.; Chesson, L.; Vignisse, J.; Steinbusch, H.W.; Anthony, D.C.; et al. Deuterium content of water increases depression susceptibility: The potential role of a serotonin-related mechanism. Behav. Brain Res. 2015, 277, 237–244. [Google Scholar] [CrossRef]
- Blackburn, T.P. Depressive Disorders: Treatment Failures and Poor Prognosis over the Last 50 Years. Pharmacol. Res. Perspect. 2019, 7, e00472. [Google Scholar] [CrossRef]
- Santomauro, D.F.; Mantilla Herrera, A.M.; Shadid, J.; Zheng, P.; Ashbaugh, C.; Pigott, D.M.; Abbafati, C.; Adolph, C.; Amlag, J.O.; Aravkin, A.Y.; et al. Global Prevalence and Burden of Depressive and Anxiety Disorders in 204 Countries and Territories in 2020 Due to the COVID-19 Pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- Gabarrell-Pascuet, A.; Varga, T.V.; Moneta, M.V.; Ayuso-Mateos, J.L.; Lara, E.; Olaya, B.; Haro, J.M.; Domènech-Abella, J. What Factors Explain the Changes in Major Depressive Disorder Symptoms by Age Group during the COVID-19 Pandemic? A Longitudinal Study. J. Affect. Disord. 2023, 328, 72–80. [Google Scholar] [CrossRef]
- Proudman, D.; Greenberg, P.; Nellesen, D. The Growing Burden of Major Depressive Disorders (MDD): Implications for Researchers and Policy Makers. Pharmacoeconomics 2021, 39, 619. [Google Scholar] [CrossRef]
- Karrouri, R.; Hammani, Z.; Otheman, Y.; Benjelloun, R. Major Depressive Disorder: Validated Treatments and Future Challenges. World J. Clin. Cases 2021, 9, 9350. [Google Scholar] [CrossRef]
- Haddad, M. Depression in Adults with a Chronic Physical Health Problem: Treatment and Management. Int. J. Nurs. Stud. 2009, 46, 1411–1414. [Google Scholar] [CrossRef] [PubMed]
- Elias, E.; Zhang, A.Y.; Manners, M.T. Novel Pharmacological Approaches to the Treatment of Depression. Life 2022, 12, 196. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strekalova, T.; Svirin, E.; Gorlova, A.; Sheveleva, E.; Burova, A.; Khairetdinova, A.; Sitdikova, K.; Zakharova, E.; Dudchenko, A.M.; Lyundup, A.; et al. Resilience and Vulnerability to Stress-Induced Anhedonia: Unveiling Brain Gene Expression and Mitochondrial Dynamics in a Mouse Chronic Stress Depression Model. Biomolecules 2023, 13, 1782. https://doi.org/10.3390/biom13121782
Strekalova T, Svirin E, Gorlova A, Sheveleva E, Burova A, Khairetdinova A, Sitdikova K, Zakharova E, Dudchenko AM, Lyundup A, et al. Resilience and Vulnerability to Stress-Induced Anhedonia: Unveiling Brain Gene Expression and Mitochondrial Dynamics in a Mouse Chronic Stress Depression Model. Biomolecules. 2023; 13(12):1782. https://doi.org/10.3390/biom13121782
Chicago/Turabian StyleStrekalova, Tatyana, Evgeniy Svirin, Anna Gorlova, Elizaveta Sheveleva, Alisa Burova, Adel Khairetdinova, Kseniia Sitdikova, Elena Zakharova, Alexander M. Dudchenko, Aleksey Lyundup, and et al. 2023. "Resilience and Vulnerability to Stress-Induced Anhedonia: Unveiling Brain Gene Expression and Mitochondrial Dynamics in a Mouse Chronic Stress Depression Model" Biomolecules 13, no. 12: 1782. https://doi.org/10.3390/biom13121782