
Response of Fibroblasts from Menkes’ and Wilson’s Copper Metabolism-Related Disorders to Ionizing Radiation: Influence of the Nucleo-Shuttling of the ATM Protein Kinase


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. X-rays Irradiation
2.3. Copper (II) Sulfate Treatment
2.4. Clonogenic Cell Survival
2.5. Immunofluorescence
2.6. Micronuclei Assay
2.7. Proximity Ligation Assay
2.8. Cell Extracts and Immunoblots
2.9. Statistical Analysis
3. Results
3.1. Cellular Radiosensitivity of MD and WD Fibroblasts
3.2. Abnormally High Levels of Micronuclei in MD and WD Fibroblasts
3.3. Abnormal Number of γH2AX Foci in MD and WD Fibroblasts
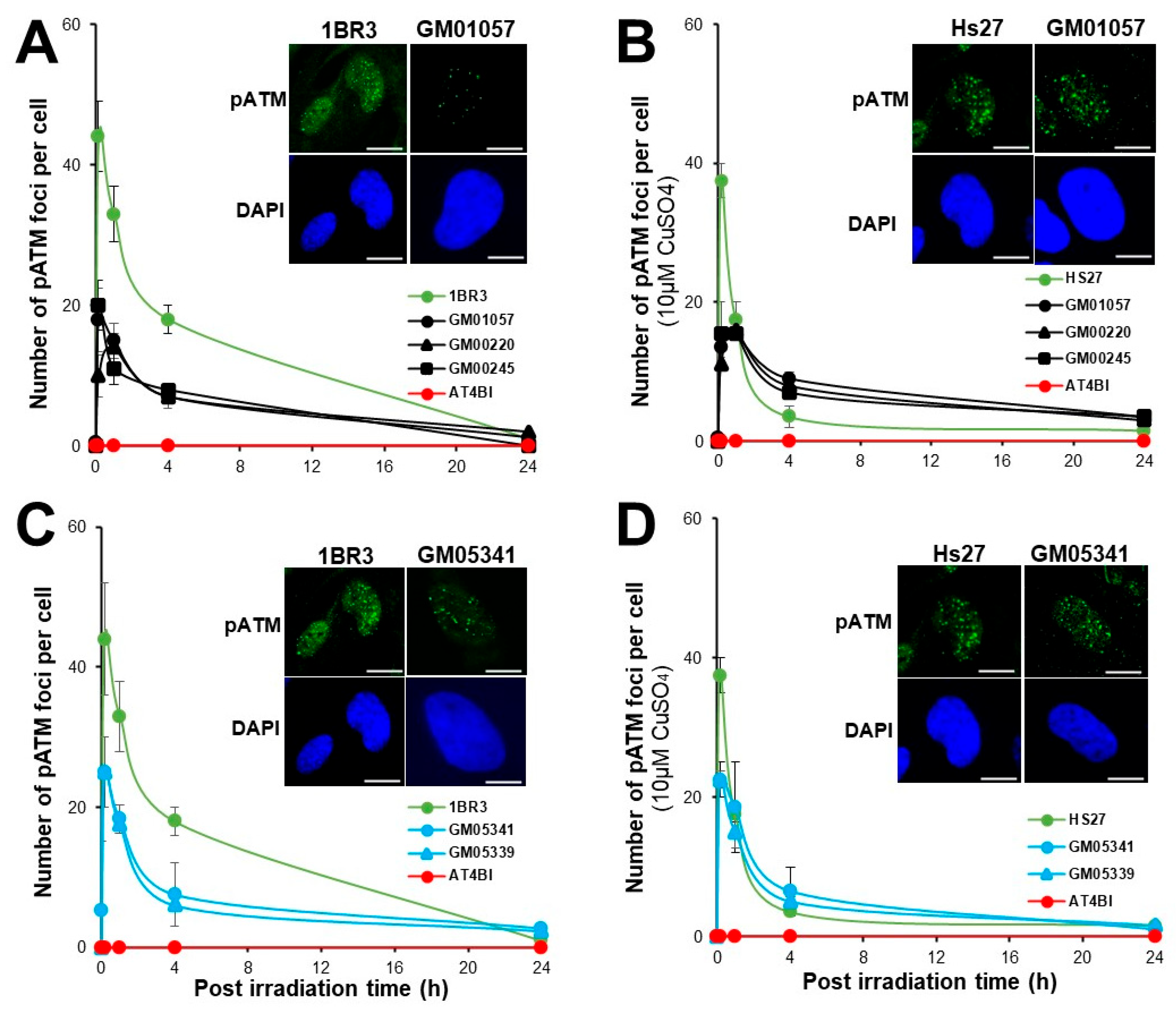
3.4. Abnormal Number of pATM Foci in MD and WD Fibroblasts
3.5. Abnormal Number of MRE11 Foci in MD and WD Fibroblasts
3.6. Expression and Subcellular Localization of the ATP7A and ATP7B Proteins
3.7. Caracterisation of the Transformed MD and WD Cells
4. Discussion
4.1. Justifications of the Approach
- (1)
- a systematic impaired DSB recognition and repair
- (2)
- a systematic delayed RIANS with cytoplasmic complexation between ATM and ATP7B (for MD cells) or between ATM and both ATP7B and ATP7A (for WD cells).
4.2. MD and WD: Towards the Identification of the X-Proteins?
- -
- Group I that gathers radioresistant individuals whose cells elicit a normal (rapid) RIANS. This group gathers individuals with low risk of radiation-induced cancer or aging disease. After two Gy X-rays, in the group I cells, all the DSB are recognized by non-homologous end-joining (NHEJ) pathway, the most predominant DSB repair pathway in humans. The group I may represent up to 75–85% individuals [29,48].
- -
- Group II that gathers individuals whose cells elicit a delayed RIANS caused by overexpression of cytoplasmic proteins, substrates of ATM and called X-proteins. This is the group of patients at risk of cancer or else aging diseases both associated with moderate but significant radiosensitivity. Particularly, this group gathers the cancer patients at risk of post-radiotherapy radiotoxicities (with CTCAE grade higher than two but lower than five). The group II may represent up to 10–25% individuals and the syndromes are generally caused by heterozygous mutations (that generally lead to over-expression of the mutated protein) [29,48].
- -
- Group III that gathers all the individuals suffering from hyper-radiosensitivity either caused by a gross DSB recognition defect (ex: homozygous ATM mutations; group IIIa) or by a gross DSB repair defect (ex: homozygous LIG4 mutations; group IIIb. The group III gathers the very severe pediatric syndromes associated with a risk of fatal radiotherapy (CTCAE grade 5). From the above calculations, the group III may represent less than 1% individuals [29,37,48] and is generally composed of very rare recessive disorders caused by the loss of an important function.
4.3. Potential Influence of Cu in the Response to Ionizing Radiation of MD and WD Cells
4.4. MD and WD: At the Edge of Cancer and Degeneration?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crichton, R.R.; Pierre, J.L. Old iron, young copper: From Mars to Venus. Biometals 2001, 14, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Daniel, K.G.; Harbach, R.H.; Guida, W.C.; Dou, Q.P. Copper storage diseases: Menkes, Wilsons, and cancer. Front. Biosci. 2004, 9, 2652–2662. [Google Scholar] [CrossRef] [PubMed]
- Scheiber, I.; Dringen, R.; Mercer, J.F.B. Copper: Effects of Deficiency and Overload. In Interrelations between Essential Metal Ions and Human Diseases; Sigel, A., Sigel, H., Sigel, R.K.O., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 359–387. [Google Scholar]
- Kaler, S.G. Inborn errors of copper metabolism. Handb. Clin. Neurol. 2013, 113, 1745–1754. [Google Scholar] [CrossRef]
- La Fontaine, S.; Ackland, M.L.; Mercer, J.F. Mammalian copper-transporting P-type ATPases, ATP7A and ATP7B: Emerging roles. Int. J. Biochem. Cell Biol. 2010, 42, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Barry, A.N.; Shinde, U.; Lutsenko, S. Structural organization of human Cu-transporting ATPases: Learning from building blocks. J. Biol. Inorg. Chem. 2010, 15, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.H.; Yang, N.; Bothe, J.; Tonelli, M.; Nokhrin, S.; Dolgova, N.V.; Braiterman, L.; Lutsenko, S.; Dmitriev, O.Y. The metal chaperone Atox1 regulates the activity of the human copper transporter ATP7B by modulating domain dynamics. J. Biol. Chem. 2017, 292, 18169–18177. [Google Scholar] [CrossRef] [PubMed]
- Tümer, Z.; Møller, L.B. Menkes disease. Eur. J. Hum. Genet. 2010, 18, 511–518. [Google Scholar] [CrossRef]
- Tümer, Z. An Overview and Update of ATP7A Mutations Leading to Menkes Disease and Occipital Horn Syndrome. Hum. Mutat. 2013, 34, 417–429. [Google Scholar] [CrossRef]
- Kaler, S.G.; Gallo, L.K.; Proud, V.K.; Percy, A.K.; Mark, Y.; Segal, N.A.; Goldstein, D.S.; Holmes, C.S.; Gahl, W.A. Occipital horn syndrome and a mild Menkes phenotype associated with splice site mutations at the MNK locus. Nat. Genet. 1994, 8, 195–202. [Google Scholar] [CrossRef]
- Caicedo-Herrera, G.; Candelo, E.; Pinilla, J.; Vidal, A.; Cruz, S.; Pachajoa, H.M. Novel ATP7A gene mutation in a patient with Menkes disease. Appl. Clin. Genet. 2018, 11, 151–155. [Google Scholar] [CrossRef]
- de Bie, P.; Muller, P.; Wijmenga, C.; Klomp, L.W. Molecular pathogenesis of Wilson and Menkes disease: Correlation of mutations with molecular defects and disease phenotypes. J. Med. Genet. 2007, 44, 673–688. [Google Scholar] [CrossRef] [PubMed]
- Kaler, S.G. Wilson’s Disease. In Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Oxford, UK, 2014. [Google Scholar]
- Compston, A. Progressive lenticular degeneration: A familial nervous disease associated with cirrhosis of the liver, by S. A. Kinnier Wilson, (From the National Hospital, and the Laboratory of the National Hospital, Queen Square, London) Brain 1912: 34; 295–509. Brain 2009, 132, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Bull, P.C.; Thomas, G.R.; Rommens, J.M.; Forbes, J.R.; Cox, D.W. The Wilson disease gene is a putative copper transporting P-type ATPase similar to the Menkes gene. Nat. Genet. 1993, 5, 327–337. [Google Scholar] [CrossRef]
- Langner, C.; Denk, H. Wilson Disease. Virchows Arch. 2004, 445, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.J.; Hahn, S.H. The genetics of Wilson Disease. Handb. Clin. Neurol. 2017, 142, 19–34. [Google Scholar] [CrossRef]
- Sandahl, T.D.; Laursen, T.L.; Munk, D.E.; Vilstrup, H.; Weiss, K.H.; Ott, P. The Prevalence of Wilson’s Disease: An Update. Hepatology 2020, 71, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, O.Y.; Bhattacharjee, A.; Nokhrin, S.; Uhlemann, E.M.; Lutsenko, S. Difference in stability of the N-domain underlies distinct intracellular properties of the E1064A and H1069Q mutants of copper-transporting ATPase ATP7B. J. Biol. Chem. 2011, 286, 16355–16362. [Google Scholar] [CrossRef]
- Scheiber, I.F.; Brůha, R.; Dušek, P. Pathogenesis of Wilson Disease. Handb. Clin. Neurol. 2017, 142, 43–55. [Google Scholar]
- Walshe, J.M.; Waldenström, E.; Sams, V.; Nordlinder, H.; Westermark, K. Abdominal malignancies in patients with Wilson’s disease. QJM 2003, 96, 657–662. [Google Scholar] [CrossRef]
- Xu, R.; Hajdu, C.H. Wilson Disease and Hepatocellular Carcinoma. Gastroenterol. Hepatol. 2008, 4, 438–439. [Google Scholar]
- Ohkoshi-Yamada, M.; Kamimura, K.; Kamimura, H.; Terai, S. Rare complication of hepatocellular carcinoma in Wilson’s disease. JGH Open 2021, 5, 1220–1222. [Google Scholar] [CrossRef] [PubMed]
- Okui, T.; Endoh, D.; Arai, S.; Hayashi, M. A high frequency of induction of chromosome aberrations in the fibroblasts of LEC strain rats by X-irradiation. J. Vet. Med. Sci. 1996, 58, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Okui, T.; Hayashi, M.; Endoh, D.; Sato, F.; Kasai, N.; Watanabe, T.; Namioka, S. A high frequency of induction of chromosome aberrations in the bone marrow cells of LEC strain rats by X-irradiation. Mutat. Res. 1994, 324, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Endoh, D.; Kon, Y.; Yamashita, T.; Sato, F.; Kasai, N.; Namioka, S. Hypersensitivity of LEC strain rats in radiation-induced acute bone marrow death. J. Vet. Med. Sci. 1993, 55, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Endoh, D.; Kon, Y.; Yamashita, T.; Hashimoto, N.; Sato, F.; Kasai, N.; Namioka, S. Higher sensitivity of LEC strains rats in radio-induced acute intestinal death. J. Vet. Med. Sci. 1992, 54, 269–273. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, X.; Yu, M.; Lin, B.; Yu, C. Radiation-induced liver injury and hepatocyte senescence. Cell Death Discov. 2021, 7, 244. [Google Scholar] [CrossRef]
- Foray, N.; Bourguignon, M.; Hamada, N. Individual response to ionizing radiation. Mutat. Res. Rev. Mutat. Res. 2016, 770, 369–386. [Google Scholar] [CrossRef]
- Combemale, P.; Sonzogni, L.; Devic, C.; Bencokova, Z.; Ferlazzo, M.L.; Granzotto, A.; Burlet, S.F.; Pinson, S.; Amini-Adle, M.; Al-Choboq, J.; et al. Individual Response to Radiation of Individuals with Neurofibromatosis Type I: Role of the ATM Protein and Influence of Statins and Bisphosphonates. Mol. Neurobiol. 2022, 59, 556–573. [Google Scholar] [CrossRef]
- Al-Choboq, J.; Ferlazzo, M.L.; Sonzogni, L.; Granzotto, A.; El-Nachef, L.; Maalouf, M.; Berthel, E.; Foray, N. Usher Syndrome Belongs to the Genetic Diseases Associated with Radiosensitivity: Influence of the ATM Protein Kinase. Int. J. Mol. Sci. 2022, 23, 1570. [Google Scholar] [CrossRef]
- Ferlazzo, M.; Berthel, E.; Granzotto, A.; Devic, C.; Sonzogni, L.; Bachelet, J.T.; Pereira, S.; Bourguignon, M.; Sarasin, A.; Mezzina, M.; et al. Some mutations in the xeroderma pigmentosum D gene may lead to moderate but significant radiosensitivity associated with a delayed radiation-induced ATM nuclear localization. Int. J. Radiat. Biol. 2020, 96, 394–410. [Google Scholar] [CrossRef]
- Ferlazzo, M.L.; Bach-Tobdji, M.K.E.; Djerad, A.; Sonzogni, L.; Devic, C.; Granzotto, A.; Bodgi, L.; Bachelet, J.T.; Djefal-Kerrar, A.; Hennequin, C.; et al. Radiobiological Characterization of Tuberous Sclerosis: A Delay in the Nucleo-Shuttling of ATM May Be Responsible for Radiosensitivity. Mol. Neurobiol. 2018, 55, 4973–4983. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, M.L.; Sonzogni, L.; Granzotto, A.; Bodgi, L.; Lartin, O.; Devic, C.; Vogin, G.; Pereira, S.; Foray, N. Mutations of the Huntington’s disease protein impact on the ATM-dependent signaling and repair pathways of the radiation-induced DNA double-strand breaks: Corrective effect of statins and bisphosphonates. Mol. Neurobiol. 2014, 49, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Al-Choboq, J.; Nehal, M.; Sonzogni, L.; Granzotto, A.; El Nachef, L.; Restier-Verlet, J.; Maalouf, M.; Berthel, E.; Aral, B.; Corradini, N.; et al. Molecular and Cellular Responses to Ionization Radiation in Untransformed Fibroblasts from the Rothmund & Thomson Syndrome: Influence of the Nucleo-Shuttling of the ATM Protein Kinase. Radiation 2023, 3, 21–38. [Google Scholar]
- Le Reun, E.; Bodgi, L.; Granzotto, A.; Sonzogni, L.; Ferlazzo, M.L.; Al-Choboq, J.; El-Nachef, L.; Restier-Verlet, J.; Berthel, E.; Devic, C.; et al. Quantitative Correlations between Radiosensitivity Biomarkers Show That the ATM Protein Kinase Is Strongly Involved in the Radiotoxicities Observed after Radiotherapy. Int. J. Mol. Sci. 2022, 23, 10434. [Google Scholar] [CrossRef] [PubMed]
- Granzotto, A.; Benadjaoud, M.A.; Vogin, G.; Devic, C.; Ferlazzo, M.L.; Bodgi, L.; Pereira, S.; Sonzogni, L.; Forcheron, F.; Viau, M.; et al. Influence of Nucleoshuttling of the ATM Protein in the Healthy Tissues Response to Radiation Therapy: Toward a Molecular Classification of Human Radiosensitivity. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 450–460. [Google Scholar] [CrossRef]
- Foray, N.; Arlett, C.F.; Malaise, E.P. Radiation-induced DNA double-strand breaks and the radiosensitivity of human cells: A closer look. Biochimie 1997, 79, 567–575. [Google Scholar] [CrossRef]
- Fertil, B.; Malaise, E.P. Inherent cellular radiosensitivity as a basic concept for human tumor radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 1981, 7, 621–629. [Google Scholar] [CrossRef]
- Grote, S.J.; Joshi, G.P.; Revell, S.H.; Shaw, C.A. Observations of radiation-induced chromosome fragment loss in live mammalian cells in culture, and its effect on colony-forming ability. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1981, 39, 395–408. [Google Scholar] [CrossRef]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. 2000, 455, 81–95. [Google Scholar] [CrossRef]
- Ristic, M.; Brockly, F.; Piechaczyk, M.; Bossis, G. Detection of Protein-Protein Interactions and Posttranslational Modifications Using the Proximity Ligation Assay: Application to the Study of the SUMO Pathway. Methods Mol. Biol. 2016, 1449, 279–290. [Google Scholar] [CrossRef]
- Bodgi, L.; Granzotto, A.; Devic, C.; Vogin, G.; Lesne, A.; Bottollier-Depois, J.F.; Victor, J.M.; Maalouf, M.; Fares, G.; Foray, N. A single formula to describe radiation-induced protein relocalization: Towards a mathematical definition of individual radiosensitivity. J. Theor. Biol. 2013, 333, 135–145. [Google Scholar] [CrossRef]
- Viau, M.; Sonzogni, L.; Ferlazzo, M.L.; Berthel, E.; Pereira, S.; Bodgi, L.; Granzotto, A.; Devic, C.; Fervers, B.; Charlet, L.; et al. DNA Double-Strand Breaks Induced in Human Cells by Twelve Metallic Species: Quantitative Inter-Comparisons and Influence of the ATM Protein. Biomolecules 2021, 11, 1462. [Google Scholar] [CrossRef]
- Onishi, T.; Inubushi, H.; Tokugawa, S.; Muramatsu, M.; Nishikawa, K.; Suzuki, Y.; Miyao, M. Abnormal Copper Metabolism in Menkes Cultured Fibroblasts. Eur. J. Pediatr. 1980, 134, 205–210. [Google Scholar] [CrossRef]
- Rothkamm, K.; Löbrich, M. Evidence for a lack of DNA double-strand break repair in human cells exposed to very low x-ray doses. Proc. Natl. Acad. Sci. USA 2003, 100, 5057–5062. [Google Scholar] [CrossRef] [PubMed]
- Joubert, A.; Zimmerman, K.M.; Bencokova, Z.; Gastaldo, J.; Chavaudra, N.; Favaudon, V.; Arlett, C.F.; Foray, N. DNA double-strand break repair defects in syndromes associated with acute radiation response: At least two different assays to predict intrinsic radiosensitivity? Int. J. Radiat. Biol. 2008, 84, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Berthel, E.; Foray, N.; Ferlazzo, M.L. The Nucleoshuttling of the ATM Protein: A Unified Model to Describe the Individual Response to High- and Low-Dose of Radiation? Cancers 2019, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- El-Nachef, L.; Al-Choboq, J.; Restier-Verlet, J.; Granzotto, A.; Berthel, E.; Sonzogni, L.; Ferlazzo, M.L.; Bouchet, A.; Leblond, P.; Combemale, P.; et al. Human radiosensitivity and radiosusceptibility: What are the differences? Int. J. Mol. Sci. 2021, 22, 7158. [Google Scholar] [CrossRef] [PubMed]
- Oka, A.; Takashima, S. Expression of the ataxia-telangiectasia gene (ATM) product in human cerebellar neurons during development. Neurosci. Lett. 1998, 252, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.S.; Kirsch, D.G.; Canman, C.E.; Ahn, J.H.; Ziv, Y.; Newman, L.S.; Darnell, R.B.; Shiloh, Y.; Kastan, M.B. ATM binds to beta-adaptin in cytoplasmic vesicles. Proc. Natl. Acad. Sci. USA 1998, 95, 10146–10151. [Google Scholar] [CrossRef]
- Guo, Z.; Kozlov, S.; Lavin, M.F.; Person, M.D.; Paull, T.T. ATM activation by oxidative stress. Science 2010, 330, 517–521. [Google Scholar] [CrossRef]
- Paull, T.T. Mechanisms of ATM Activation. Annu. Rev. Biochem. 2015, 84, 711–738. [Google Scholar] [CrossRef] [PubMed]
- Foray, N.; Marot, D.; Gabriel, A.; Randrianarison, V.; Carr, A.M.; Perricaudet, M.; Ashworth, A.; Jeggo, P. A subset of ATM- and ATR-dependent phosphorylation events requires the BRCA1 protein. EMBO J. 2003, 22, 2860–2871. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Origin | Syndrome | Genetic Features * | Clinical Features * |
|---|---|---|---|---|
| 1BR3 | ECACC | - | Apparently healthy | Apparently healthy radioresistant |
| MRC5 | ECACC | - | Apparently healthy | Apparently healthy radioresistant |
| Hs27 | ECACC | - | Apparently healthy | Apparently healthy radioresistant |
| AT4BI | COPERNIC | AT | ATM mutations | Hyper-radiosensitive |
| GM04068 | Coriell Institute | Relative of MD patients | abnormal metallothionein gene regulation in response to Cu | Mother of GM00245 |
| GM01057 | Coriell Institute | MD | abnormal metallothionein gene regulation in response to Cu | Fibroblasts exhibit elevated Cu concentration |
| GM00220 | Coriell Institute | MD | abnormal metallothionein gene regulation in response to Cu; culture shows qualitative changes in Mc1 mRNA | Fibroblasts exhibit elevated Cu levels; positive family history; similarly affected brother and cousin |
| GM00245 | Coriell Institute | MD | abnormal metallothionein gene regulation in response to Cu | Fibroblasts exhibit elevated Cu levels |
| GM05762 | Coriell Institute | Relative of MD patients | - | No evidence of WD; rheumatoid arthritis since age 21; mother of the 2 affected daughters GM05339, GM05341 |
| GM05339 | Coriell Institute | WD | 46,XX,1qh+.arr Xq13.2q21.1(72592523-77910651)x2 hmz,15q11.1q11.2(20601541-21939811)x3 | Onset at age 13 with jaundice, evidence of chronic liver disease, presence of Kayser-Fleischer rings, & low ceruloplasmin |
| GM05341 | Coriell Institute | WD | - | Diagnosed as asymptomatic at age 12 with low serum ceruloplasmin, elevated urinary & hepatic Cu levels; no evidence of liver disease or central nervous system dysfunction; |
| MD Cells | WD Cells | |
|---|---|---|
| Gene | ATP7A mutations | ATP7B mutations |
| Major clinical feature | Abnormally high Cu accumulation in other tissues than liver | Abnormally high Cu accumulation in the liver |
| Cell survival (SF2) | 30–60% (Radioresistance SF2 > 60%) | 58–70% (Radioresistance SF2 > 60%) |
| Micronuclei yield | Not significant Increases with Cu-pretreatment | |
| Number of γH2AX foci | Lower at 10 min post-irradiation than controls—Delayed RIANS | |
| Number of pATM foci | Lower at 10 min post-irradiation than controls—Delayed RIANS | |
| Number of MRE11 foci | Lower than controls. Early MRE11 foci after Cu pre-treatment | Lower than controls. Early and late MRE11 foci after Cu pre-treatment |
| Gene expression | ATP7A: No/low expression ATP7B: very high expression | ATP7A: Cytoplasmic expression ATP7B: very high expression |
| Cytoplasmic protein–protein interactions | ATP7B-ATM complexes | ATP7B-ATM complexes Some ATP7A-ATM complexes |
| Cellular transformation in culture | Spontaneous immortalization from passages 28–31 | No signs of immortalization or senescence observed up to passages 28–31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Nachef, L.; Al-Choboq, J.; Bourguignon, M.; Foray, N. Response of Fibroblasts from Menkes’ and Wilson’s Copper Metabolism-Related Disorders to Ionizing Radiation: Influence of the Nucleo-Shuttling of the ATM Protein Kinase. Biomolecules 2023, 13, 1746. https://doi.org/10.3390/biom13121746
El Nachef L, Al-Choboq J, Bourguignon M, Foray N. Response of Fibroblasts from Menkes’ and Wilson’s Copper Metabolism-Related Disorders to Ionizing Radiation: Influence of the Nucleo-Shuttling of the ATM Protein Kinase. Biomolecules. 2023; 13(12):1746. https://doi.org/10.3390/biom13121746
Chicago/Turabian StyleEl Nachef, Laura, Joëlle Al-Choboq, Michel Bourguignon, and Nicolas Foray. 2023. "Response of Fibroblasts from Menkes’ and Wilson’s Copper Metabolism-Related Disorders to Ionizing Radiation: Influence of the Nucleo-Shuttling of the ATM Protein Kinase" Biomolecules 13, no. 12: 1746. https://doi.org/10.3390/biom13121746