

Antimicrobial Potential of Different Isolates of Chaetomium globosum Combined with Liquid Chromatography Tandem Mass Spectrometry Chemical Profiling
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolation and Identification
2.1.1. Collection of Samples
2.1.2. Isolation and Preservation of Mycobiota
2.1.3. Phenotypic Identification of Chaetomium Isolates
2.2. Culture of Chaetomium Isolates and Extraction of Their Active Metabolites
2.2.1. Subcultures of Pure Isolates of C. globosum
2.2.2. Extraction of Active Metabolites
2.3. Assessment of Antimicrobial Activity of Different Isolates of Chaetomium Isolates
2.3.1. Collection of Clinical Samples and Isolation of Microbes
2.3.2. Antibiotics Susceptibility Test for Determination of MDR Isolates
2.3.3. Screening of Antimicrobial Activity of Chaetomium Taxa against MDR Isolates
2.3.4. Determination of Minimum Inhibitory Concentrations (MIC) of Active Isolates
2.4. Metabolic Profiling of Selected Chaetomium Isolates Using LC-MS/MS
3. Results and Discussion
3.1. Fungal Sources and Identification of Chaetomium sp.
3.2. Antimicrobial Assessment of Chaetomium Isolates
3.2.1. Microbial Isolates and Their Antibiotic Resistance Pattern
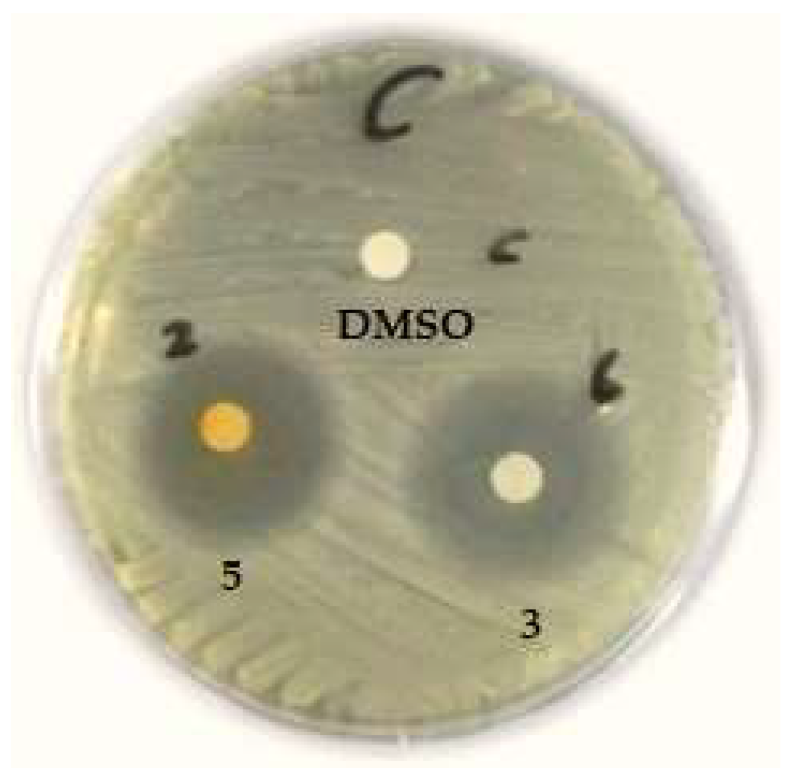
3.2.2. The Antimicrobial Activity of Chaetomium Isolates against MDR Strains
3.2.3. Minimum Inhibitory Concentration (MIC) of Fungal Extracts
3.3. LC-MS/MS Metabolic Profiling of Crude Extracts of C. globosum Isolates 3 and 5
3.3.1. Identification of Natural Metabolites in the Crude Extract of C. globosum Soil Taxon Recovered from Cultivated Soil in Assiut Governorate, Upper Egypt, (3) Using LC-MS/MS Technique
3.3.2. Identification of Natural Metabolites in the Crude Extract of C. globosum Endophytic Taxon Recovered from Zygophyllum album (5), Wadi El-Arbaein, Saint Katherine, South Sinai, Using LC-MS/MS Technique
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badau, E. A One Health Perspective on the Issue of the Antibiotic Resistance. Parasite 2021, 28, 16. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and Combating Antibiotic Resistance from One Health and Global Health Perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Sridhar, S.; Turbett, S.E.; Harris, J.B.; Larocque, R.C. Antimicrobial-Resistant Bacteria in International Travelers. Curr. Opin. Infect. Dis. 2021, 34, 423–431. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645. [Google Scholar] [CrossRef]
- Bate, P.N.N.; Orock, A.E.; Nyongbela, K.D.; Babiaka, S.B.; Kukwah, A.; Ngemenya, M.N. In Vitro Activity against Multi-Drug Resistant Bacteria and Cytotoxicity of Lichens Collected from Mount Cameroon. J. King Saud Univ. -Sci. 2020, 32, 614–619. [Google Scholar] [CrossRef]
- Elkhateeb, W.A. Highlights on Chaetomium Morphology, Secondary Metabolites and Biological Activates. Pharm. Pharmacol. Res. 2021, 4, 1–5. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.M. Taxonomy and Biodiversity of the Genus Chaetomium in Different Habitats-Recent Developments on Genus Chaetomium; Abdel-Azeem, A.M., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 3–77. ISBN 978-3-030-31612-9. [Google Scholar]
- Wijeratne, E.M.K.; Turbyville, T.J.; Fritz, A.; Whitesell, L.; Gunatilaka, A.A.L. A New Dihydroxanthenone from a Plant-Associated Strain of the Fungus Chaetomium globosum Demonstrates Anticancer Activity. Bioorg. Med. Chem. 2006, 14, 7917–7923. [Google Scholar] [CrossRef]
- El-Kassem, L.A.; Hawas, U.W.; El-Souda, S.; Ahmed, E.F.; El-Khateeb, W.; Fayad, W. Anti-HCV Protease Potential of Endophytic Fungi and Cytotoxic Activity. Biocatal. Agric. Biotechnol. 2019, 19, 101170. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. The Natural Functions of Secondary Metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [CrossRef]
- Zhang, J.; Ge, H.M.; Jiao, R.H.; Li, J.; Peng, H.; Wang, Y.R.; Wu, J.H.; Song, Y.C.; Tan, R.X. Cytotoxic Chaetoglobosins from the Endophyte Chaetomium globosum. Planta Med. 2010, 76, 1910–1914. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, H.-Q.; Zong, S.-C.; Gao, J.-M.; Zhang, A.-L. Chemical and Bioactive Diversities of the Genus Chaetomium Secondary Metabolites. Mini-Rev. Med. Chem. 2012, 12, 127–148. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Abdel-Wareth, M.T.A. The Use of Chaetomium Taxa as Biocontrol Agents BT-Recent Developments on Genus Chaetomium; Abdel-Azeem, A.M., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 251–266. ISBN 978-3-030-31612-9. [Google Scholar]
- Zhao, S.S.; Zhang, Y.Y.; Yan, W.; Cao, L.L.; Xiao, Y.; Ye, Y.H. Chaetomium Globosum CDW7, a Potential Biological Control Strain and Its Antifungal Metabolites. FEMS Microbiol. Lett. 2017, 364, fnw287. [Google Scholar] [CrossRef]
- Gao, W.; He, Y.; Li, F.; Chai, C.; Zhang, J.; Guo, J.; Chen, C.; Wang, J.; Zhu, H.; Hu, Z.; et al. Antibacterial Activity against Drug-Resistant Microbial Pathogens of Cytochalasan Alkaloids from the Arthropod-Associated Fungus Chaetomium Globosum TW1-1. Bioorganic Chem. 2019, 83, 98–104. [Google Scholar] [CrossRef]
- Warcup, J.H.; Baker, K.F. Occurrence of Dormant Ascospores in Soil. Nature 1963, 197, 1317–1318. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.M.; Abdel-Azeem, M.A.; Khalil, W.F. Endophytic Fungi as a New Source of Antirheumatoid Metabolites. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases; Academic Press: Cambridge, MA, USA, 2019; pp. 355–384. [Google Scholar] [CrossRef]
- Smith, N.R.; Dawson, V.T. The Bacteriostatic Action of Rose Bengal in Media Used for Plate Counts of Soil Fungi. Soil Sci. 1944, 58, 467–472. [Google Scholar] [CrossRef]
- Guarro, J. Atlas of Soil Ascomycetes; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2012; p. 486. [Google Scholar]
- Abdel-Azeem, A.M. Recent Developments on Genus Chaetomium; Springer Science and Business Media LLC: Dordrecht, The Netherlands, 2020; p. 453. [Google Scholar] [CrossRef]
- Kirk, P.; Cannon, P.; Stalpers, J.; Minter, D.W. Dictionary of the Fungi, 10th ed.; CABI Publishing: Wallingford, UK, 2008. [Google Scholar]
- Cheng, M.; Zhao, S.; Liu, H.; Liu, Y.; Lin, C.; Song, J.; Thawai, C.; Charoensettasilp, S.; Yang, Q. Functional Analysis of a Chaetoglobosin A Biosynthetic Regulator in Chaetomium globosum. Fungal Biol. 2021, 125, 201–210. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Abdel-hamed, A.R.; Mehanna, E.T.; Hazem, R.M.; Badr, J.M.; Abo-elmatty, D.M.; Abdel-kader, M.S.; Goda, M.S. Plicosepalus Acacia Extract and Its Major Constituents, Methyl Gallate and Quercetin, Potentiate Therapeutic Angiogenesis in Diabetic Hind Limb Ischemia: HPTLC Quantification and LC-MS/MS Metabolic Profiling. Antioxidants 2021, 10, 1701. [Google Scholar] [CrossRef]
- Goda, M.S.; Nafie, M.S.; Awad, B.M.; Abdel-Kader, M.S.; Ibrahim, A.K.; Badr, J.M.; Eltamany, E.E. In Vitro and In Vivo Studies of Anti-Lung Cancer Activity of Artemesia judaica L. Crude Extract Combined with LC-MS/MS Metabolic Profiling, Docking Simulation and HPLC-DAD Quantification. Antioxidants 2021, 11, 17. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Habib, E.S.; Goda, M.S.; Fahim, J.R.; Hassanean, H.A.; Eltamany, E.E.; Ibrahim, A.K.; Aboulmagd, A.M.; Fayez, S.; El-Kader, A.M.A.; et al. Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron Ciliatum. Mar. Drugs 2020, 18, 354. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Elhady, S.S.; Goda, M.S.; Aly, O.M.; Habib, E.S.; Ibrahim, A.K.; Hassanean, H.A.; Abdelmohsen, U.R.; Safo, M.K.; Ahmed, S.A. Chemical Composition of the Red Sea Green Algae Ulva Lactuca: Isolation and in Silico Studies of New Anti-COVID-19 Ceramides. Metabolites 2021, 11, 816. [Google Scholar] [CrossRef]
- Elhady, S.S.; Habib, E.S.; Abdelhameed, R.F.A.; Goda, M.S.; Hazem, R.M.; Mehanna, E.T.; Helal, M.A.; Hosny, K.M.; Diri, R.M.; Hassanean, H.A.; et al. Anticancer Effects of New Ceramides Isolated from the Red Sea Red Algae Hypnea musciformis in a Model of Ehrlich Ascites Carcinoma: LC-HRMS Analysis Profile and Molecular Modeling. Mar. Drugs 2022, 20, 63. [Google Scholar] [CrossRef]
- Mulu, W.; Abera, B.; Yimer, M.; Hailu, T.; Ayele, H.; Abate, D. Bacterial Agents and Antibiotic Resistance Profiles of Infections from Different Sites That Occurred among Patients at Debre Markos Referral Hospital, Ethiopia: A Cross-Sectional Study. BMC Res. Notes 2017, 10, 254. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Abdelkareem, M.Z.; Sayed, M.; Hassuna, N.A.; Mahmoud, M.S.; Abdelwahab, S.F. Multi-Drug-Resistant Enterococcus faecalis among Egyptian Patients with Urinary Tract Infection. J. Chemother. 2017, 29, 74–82. [Google Scholar] [CrossRef]
- Farman, M.; Yasir, M.; Al-Hindi, R.R.; Farraj, S.A.; Jiman-Fatani, A.A.; Alawi, M.; Azhar, E.I. Genomic Analysis of Multidrug-Resistant Clinical Enterococcus faecalis Isolates for Antimicrobial Resistance Genes and Virulence Factors from the Western Region of Saudi Arabia. Antimicrob. Resist. Infect. Control 2019, 8, 55. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mishra, S.; Tiwary, S. Microbial Profile and Antibiogram of Pus Isolate in a Tertiary Care Hospital of Western Odisha. Orig. Res. Artic. J. Evol. Med. Dent. Sci 2020, 9, 1325–1330. [Google Scholar] [CrossRef]
- Khanam, R.A.; Islam, M.R.; Sharif, A.; Parveen, R.; Sharmin, I.; Yusuf, M.A. Bacteriological Profiles of Pus with Antimicrobial Sensitivity Pattern at a Teaching Hospital in Dhaka City. Bangladesh J. Infect. Dis. 2018, 5, 10–14. [Google Scholar] [CrossRef]
- Bazaid, A.S.; Aldarhami, A.; Gattan, H.; Barnawi, H.; Qanash, H.; Alsaif, G.; Alharbi, B.; Alrashidi, A.; Eldrehmy, E.H. Antibiogram of Urinary Tract Infections and Sepsis among Infants in Neonatal Intensive Care Unit. Children 2022, 9, 629. [Google Scholar] [CrossRef]
- Omololu-Aso, J.; Omololu-Aso, O.O.; Adekanye, N.; Alex, T.; Owolabi, T.A.; Shesha, A. Antimicrobial Susceptibility Pattern of Escherichia coli Isolates from Clinical Sources at Tertiary Health Care Setting, Ile Ife, South Western Nigeria. Eur. J. Exp. Biol. 2017, 7, 5. [Google Scholar] [CrossRef]
- Naqid, I.A.; Balatay, A.A.; Hussein, N.R.; Saeed, K.A.; Ahmed, H.A.; Yousif, S.H. Antibiotic Susceptibility Pattern of Escherichia coli Isolated from Various Clinical Samples in Duhok City, Kurdistan Region of Iraq. Int. J. Infect. 2020, 7, e103740. [Google Scholar] [CrossRef]
- World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef]
- Oliveira, R.; Castro, J.; Silva, S.; Oliveira, H.; Saavedra, M.J.; Azevedo, N.F.; Almeida, C. Exploring the Antibiotic Resistance Profile of Clinical Klebsiella pneumoniae Isolates in Portugal. Antibiotics 2022, 11, 1613. [Google Scholar] [CrossRef]
- Mirzaei, A.; Habibi, M.; Bouzari, S.; Karam, M.R.A. Characterization of Antibiotic-Susceptibility Patterns, Virulence Factor Profiles and Clonal Relatedness in Proteus mirabilis Isolates from Patients with Urinary Tract Infection in Iran. Infect. Drug Resist. 2019, 12, 3967–3979. [Google Scholar] [CrossRef]
- Hasan, T.H.; Alasedi, K.; Aljanaby, A. Proteus Mirabilis Virulence Factors: Review. Int. J. Pharm. Res. 2020, 13, 2145–2149. [Google Scholar] [CrossRef]
- Imanah, E.O.; Beshiru, A.; Igbinosa, E.O. Antibiogram Profile of Pseudomonas aeruginosa Isolated from Some Selected Hospital Environmental Drains. Asian Pac. J. Trop. Dis. 2017, 7, 604–609. [Google Scholar] [CrossRef]
- Golle, A.; Janezic, S.; Rupnik, M. Low Overlap between Carbapenem Resistant Pseudomonas aeruginosa Genotypes Isolated from Hospitalized Patients and Wastewater Treatment Plants. PLoS ONE 2017, 12, e0186736. [Google Scholar] [CrossRef]
- Ghaith, D.M.; Zafer, M.M.; Ismail, D.K.; Al-Agamy, M.H.; Bohol, M.F.F.; Al-Qahtani, A.; Al-Ahdal, M.N.; Elnagdy, S.M.; Mostafa, I.Y. First Reported Nosocomial Outbreak of Serratia Marcescens Harboring Bla IMP-4 and Bla VIM-2 in a Neonatal Intensive Care Unit in Cairo, Egypt. Infect. Drug Resist. 2018, 11, 2211–2217. [Google Scholar] [CrossRef]
- Cristina, M.L.; Sartini, M.; Spagnolo, A.M. Serratia Marcescens Infections in Neonatal Intensive Care Units (NICUs). Int. J. Environ. Res. Public Health 2019, 16, 610. [Google Scholar] [CrossRef]
- Mohammed, A.E.; Sonbol, H.; Alwakeel, S.S.; Alotaibi, M.O.; Alotaibi, S.; Alothman, N.; Suliman, R.S.; Ahmedah, H.T.; Ali, R. Investigation of Biological Activity of Soil Fungal Extracts and LC/MS-QTOF Based Metabolite Profiling. Sci. Rep. 2021, 11, 4760. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Dufossé, L.; Chhipa, H.; Saxena, S.; Mahajan, G.B.; Gupta, M.K. Fungal Endophytes: A Potential Source of Antibacterial Compounds. J. Fungi 2022, 8, 164. [Google Scholar] [CrossRef]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Shi, D.; Ren, L. Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef]
- Prima, S.R.; Elfahmi; Julianti, E.; Fidrianny, I. Evaluation of Antibacterial and Antioxidant Activity of Endophytic Fungi Isolated from Capsicum annuum L. and Allium cepa L. Pharmacogn. J. 2022, 14, 329–334. [Google Scholar] [CrossRef]
- Dramae, A.; Intaraudom, C.; Bunbamrung, N.; Boonyuen, N.; Auncharoen, P.; Pittayakhajonwut, P. Antimicrobial Tanzawaic Acid Derivatives from the Endophytic Penicillium Citrinum BCC71086. Tetrahedron 2022, 106–107, 132645. [Google Scholar] [CrossRef]
- Khan, B.; Yan, W.; Wei, S.; Wang, Z.; Zhao, S.; Cao, L.; Rajput, N.A.; Ye, Y. Nematicidal Metabolites from Endophytic Fungus Chaetomium globosum YSC5. FEMS Microbiol. Lett. 2019, 366, 169. [Google Scholar] [CrossRef]
- Manganyi, M.C.; Ateba, C.N. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms 2020, 8, 1934. [Google Scholar] [CrossRef]
- Johansson, L.; Lindskog, A.; Silfversparre, G.; Cimander, C.; Nielsen, K.F.; Lidén, G. Shikimic Acid Production by a Modified Strain of E. coli (W3110.Shik1) under Phosphate-Limited and Carbon-Limited Conditions. Biotechnol. Bioeng. 2005, 92, 541–552. [Google Scholar] [CrossRef]
- Buiarelli, F.; Di Filippo, P.; Riccardi, C.; Pomata, D.; Rumolo, E.; Giannetti, L.; Neri, B. Analytical Method for the Determination of Mycotoxins in Indoor/Outdoor Airborne Particulate Matter by HPLC-MS-MS. Int. J. Environ. Anal. Chem. 2015, 95, 713–729. [Google Scholar] [CrossRef]
- García-Salas, P.; Gómez-Caravaca, A.M.; Morales-Soto, A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Identification and Quantification of Phenolic and Other Polar Compounds in the Edible Part of Annona Cherimola and Its By-Products by HPLC-DAD-ESI-QTOF-MS. Food Res. Int. 2015, 78, 246–257. [Google Scholar] [CrossRef]
- Kramberger, K.; Barlič-Maganja, D.; Bandelj, D.; Baruca Arbeiter, A.; Peeters, K.; Miklavčič Višnjevec, A.; Pražnikar, Z.J. HPLC-DAD-ESI-QTOF-MS Determination of Bioactive Compounds and Antioxidant Activity Comparison of the Hydroalcoholic and Water Extracts from Two Helichrysum italicum Species. Metabolites 2020, 10, 403. [Google Scholar] [CrossRef]
- Szczesny, D.; Bartosińska, E.; Jacyna, J.; Patejko, M.; Siluk, D.; Kaliszan, R. Quantitative Determination of Trigonelline in Mouse Serum by Means of Hydrophilic Interaction Liquid Chromatography–MS/MS Analysis: Application to a Pharmacokinetic Study. Biomed. Chromatogr. 2018, 32, e4054. [Google Scholar] [CrossRef]
- Liang, S.; Sanchez-Espiridion, B.; Xie, H.; Ma, J.; Wu, X.; Liang, D. Determination of Proline in Human Serum by a Robust LC-MS/MS Method: Application to Identification of Human Metabolites as Candidate Biomarkers for Esophageal Cancer Early Detection and Risk Stratification. Biomed. Chromatogr. 2015, 29, 570–577. [Google Scholar] [CrossRef]
- Guo, Y.P.; Yang, H.; Wang, Y.L.; Chen, X.X.; Zhang, K.; Wang, Y.L.; Sun, Y.F.; Huang, J.; Yang, L.; Wang, J.H. Determination of Flavonoids Compounds of Three Species and Different Harvesting Periods in Crataegi Folium Based on Lc-Ms/Ms. Molecules 2021, 26, 1602. [Google Scholar] [CrossRef]
- Yan, W.; Cao, L.L.; Zhang, Y.Y.; Zhao, R.; Zhao, S.S.; Khan, B.; Ye, Y.H. New Metabolites from Endophytic Fungus Chaetomium globosum CDW7. Molecules 2018, 23, 2873. [Google Scholar] [CrossRef]
- Wang, Y.; Song, M.; Hang, T.; Wen, A.; Yang, L. LC-MS-MS Simultaneous Determination of Niacin, Niacinamide and Nicotinuric Acid in Human Plasma LC-MS-MS and Its Application to a Human Pharmacokinetic Study. Chromatographia 2010, 72, 245–253. [Google Scholar] [CrossRef]
- Varga, E.; Glauner, T.; Berthiller, F.; Krska, R.; Schuhmacher, R.; Sulyok, M. Development and Validation of a (Semi-)Quantitative UHPLC-MS/MS Method for the Determination of 191 Mycotoxins and Other Fungal Metabolites in Almonds, Hazelnuts, Peanuts and Pistachios. Anal. Bioanal. Chem. 2013, 405, 5087–5104. [Google Scholar] [CrossRef]
- Del Mar Contreras, M.; Bribi, N.; Gómez-Caravaca, A.M.; Gálvez, J.; Segura-Carretero, A. Alkaloids Profiling of Fumaria capreolata by Analytical Platforms Based on the Hyphenation of Gas Chromatography and Liquid Chromatography with Quadrupole-Time-of-Flight Mass Spectrometry. Int. J. Anal. Chem. 2017, 2017, 5178729. [Google Scholar] [CrossRef]
- Menéndez-Perdomo, I.M.; Hagel, J.M.; Facchini, P.J. Benzylisoquinoline Alkaloid Analysis Using High-Resolution Orbitrap LC-MSn. J. Mass Spectrom. 2021, 56, e4683. [Google Scholar] [CrossRef]
- Yao, C.; Bai, H.-H.; Zhang, Q.; Qian, X.-Q.; Zhang, X.; Wu, L.-W.; Yang, T. Secondary Metabolites from the Fungus Chaetomium elatum CIB-412. Chem. Nat. Compd. 2019, 55, 899–902. [Google Scholar] [CrossRef]
- Dwibedi, V.; Rath, S.K.; Jain, S.; Martínez-Argueta, N.; Prakash, R.; Saxena, S.; Rios-Solis, L. Key Insights into Secondary Metabolites from Various Chaetomium Species. Appl. Microbiol. Biotechnol. 2023, 107, 1077–1093. [Google Scholar] [CrossRef]
- Li, X.; Tian, Y.; Yang, S.X.; Zhang, Y.M.; Qin, J.C. Cytotoxic Azaphilone Alkaloids from Chaetomium globosum TY1. Bioorganic Med. Chem. Lett. 2013, 23, 2945–2947. [Google Scholar] [CrossRef]
- Dar, P.M.; Abdel-Azeem, A.M.; Hamed, S.R. Chaetomium’s Alkaloids. In Recent Developments on Genus Chaetomium; Springer: Cham, Switzerland, 2020; pp. 205–225. [Google Scholar] [CrossRef]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Handbook of Secondary Fungal Metabolites; Gulf Professional Publishing: Houston, TX, USA, 2003; pp. 1–672. [Google Scholar]
- Hebra, T.; Elie, N.; Poyer, S.; Van Elslande, E.; Touboul, D.; Eparvier, V. Dereplication, Annotation, and Characterization of 74 Potential Antimicrobial Metabolites from Penicillium sclerotiorum Using t-SNE Molecular Networks. Metabolites 2021, 11, 444. [Google Scholar] [CrossRef]
- Hoeksma, J.; Misset, T.; Wever, C.; Kemmink, J.; Kruijtzer, J.; Versluis, K.; Liskamp, R.M.J.; Boons, G.J.; Heck, A.J.R.; Boekhout, T.; et al. A New Perspective on Fungal Metabolites: Identification of Bioactive Compounds from Fungi Using Zebrafish Embryogenesis as Read-Out. Sci. Rep. 2019, 9, 17546. [Google Scholar] [CrossRef]
- Zhou, J.; Chan, L.; Zhou, S. Trigonelline: A Plant Alkaloid with Therapeutic Potential for Diabetes and Central Nervous System Disease. Curr. Med. Chem. 2012, 19, 3523–3531. [Google Scholar] [CrossRef]
- Paulo, D.Q.M.; Barbosa-Filho, J.M.; Lima, E.O.; Maia, R.F.; de Cassia, R.; Barbosa, B.B.C.; Kaplan, M.A.C. Antimicrobial Activity of Benzylisoquinoline Alkaloids from Annona Salzmanii D.C. J. Ethnopharmacol. 1992, 36, 39–41. [Google Scholar] [CrossRef]
- Conceição, R.S.; Perez, C.J.; Branco, A.; Botura, M.B.; Ifa, D.R. Identification of Sassafras Albidum Alkaloids by High-Performance Thin-Layer Chromatography Tandem Mass Spectrometry and Mapping by Desorption Electrospray Ionization Mass Spectrometry Imaging. J. Mass Spectrom. 2021, 56, e4674. [Google Scholar] [CrossRef]
- Al-Ghazzawi, A.M. Anti-Cancer Activity of New Benzyl Isoquinoline Alkaloid from Saudi Plant Annona Squamosa. BMC Chem. 2019, 13, 13. [Google Scholar] [CrossRef]
- Wangchuk, P.; Sastraruji, T.; Taweechotipatr, M.; Keller, P.A.; Pyne, S.G. Anti-Inflammatory, Anti-Bacterial and Anti-Acetylcholinesterase Activities of Two Isoquinoline Alkaloids–Scoulerine and Cheilanthifoline. Nat. Prod. Commun. 2016, 11, 1801–1804. [Google Scholar] [CrossRef]
- Siddiquee, S. Recent Advancements on the Role of Biologically Active Secondary Metabolites from Aspergillus. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 69–94. [Google Scholar] [CrossRef]
- Xu, G.B.; He, G.; Bai, H.H.; Yang, T.; Zhang, G.L.; Wu, L.W.; Li, G.Y. Indole Alkaloids from Chaetomium globosum. J. Nat. Prod. 2015, 78, 1479–1485. [Google Scholar] [CrossRef]
- Chen, C.; Tao, H.; Chen, W.; Yang, B.; Zhou, X.; Luo, X.; Liu, Y. Recent Advances in the Chemistry and Biology of Azaphilones. RSC Adv. 2020, 10, 10197. [Google Scholar] [CrossRef]
- McMullin, D.R.; Sumarah, M.W.; Miller, J.D. Chaetoglobosins and Azaphilones Produced by Canadian Strains of Chaetomium globosum Isolated from the Indoor Environment. Mycotoxin Res. 2013, 29, 47–54. [Google Scholar] [CrossRef]
- Ding, G.; Song, Y.; Chen, J.R.; Xu, C.; Ge, H.; Wang, X.T.; Tan, R. Chaetoglobosin U, a Cytochalasan Alkaloid from Endophytic Chaetomium globosum IFB-E019. J. Nat. Prod. 2006, 69, 302–304. [Google Scholar] [CrossRef]
- Cooper, J.A. Effects of Cytochalasin and Phalloidin on Actin. J. Cell Biol. 1987, 105, 1473–1478. [Google Scholar] [CrossRef]
- Takahashi, J.A.; de Castro, M.C.M.; Souza, G.G.; Lucas, E.M.F.; Bracarense, A.A.P.; Abreu, L.M.; Marriel, I.E.; Oliveira, M.S.; Floreano, M.B.; Oliveira, T.S. Isolation and Screening of Fungal Species Isolated from Brazilian Cerrado Soil for Antibacterial Activity against Escherichia coli, Staphylococcus aureus, Salmonella typhimurium, Streptococcus pyogenes and Listeria Monocytogenes. J. Mycol. Med. 2008, 18, 198–204. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone Promotes Cell Proliferation or Causes Cell Death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef]
- Stanely Mainzen Prince, P.; Rajakumar, S.; Dhanasekar, K. Protective Effects of Vanillic Acid on Electrocardiogram, Lipid Peroxidation, Antioxidants, Proinflammatory Markers and Histopathology in Isoproterenol Induced Cardiotoxic Rats. Eur. J. Pharmacol. 2011, 668, 233–240. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Ibeji, C.U.; Olasehinde, T.A.; Koorbanally, N.A.; Islam, M.S. Vanillin and Vanillic Acid Modulate Antioxidant Defense System via Amelioration of Metabolic Complications Linked to Fe2+-Induced Brain Tissues Damage. Metab. Brain Dis. 2020, 35, 727–738. [Google Scholar] [CrossRef]
- Mourtzinos, I.; Konteles, S.; Kalogeropoulos, N.; Karathanos, V.T. Thermal Oxidation of Vanillin Affects Its Antioxidant and Antimicrobial Properties. Food Chem. 2009, 114, 791–797. [Google Scholar] [CrossRef]
- Ul Amin, F.; Shah, S.A.; Kim, M.O. Vanillic Acid Attenuates Aβ1-42-Induced Oxidative Stress and Cognitive Impairment in Mice. Sci. Rep. 2017, 7, 40753. [Google Scholar] [CrossRef]
- Mardirossian, M.; Sola, R.; Beckert, B.; Collis, D.W.P.; Di Stasi, A.; Armas, F.; Hilpert, K.; Wilson, D.N.; Scocchi, M. Proline-Rich Peptides with Improved Antimicrobial Activity against E. coli, K. pneumoniae, and A. baumannii. ChemMedChem 2019, 14, 2025. [Google Scholar] [CrossRef]
- Xing, X.R.; Liao, Z.; Tan, F.; Zhu, Z.Y.; Jiang, Y.; Cao, Y.Y. Effect of Nicotinamide against Candida albicans. Front. Microbiol. 2019, 10, 595. [Google Scholar] [CrossRef]
- Kiefer, A.; Arnholdt, M.; Grimm, V.; Geske, L.; Groß, J.; Vierengel, N.; Opatz, T.; Erkel, G. Structure Elucidation and Biological Activities of Perylenequinones from an Alternaria Species. Mycotoxin Res. 2023, 1, 303–316. [Google Scholar] [CrossRef]
- Mahmoud, M.M.; Abdel-Razek, A.S.; Soliman, H.S.M.; Ponomareva, L.V.; Thorson, J.S.; Shaaban, K.A.; Shaaban, M. Diverse Polyketides from the Marine Endophytic Alternaria sp. LV52: Structure Determination and Cytotoxic Activities. Biotechnol. Rep. 2021, 33, e00628. [Google Scholar] [CrossRef]
- Kezimana, P.; Dmitriev, A.A.; Kudryavtseva, A.V.; Romanova, E.V.; Melnikova, N.V. Secoisolariciresinol Diglucoside of Flaxseed and Its Metabolites: Biosynthesis and Potential for Nutraceuticals. Front. Genet. 2018, 9, 641. [Google Scholar] [CrossRef]
- De Carvalho, M.; Weich, H.; Abraham, W.-R. Macrocyclic Trichothecenes as Antifungal and Anticancer Compounds. Curr. Med. Chem. 2016, 23, 23–35. [Google Scholar] [CrossRef]
- Bodet, C.A.; Jorgensen, J.H.; Drutz, D.J. Antibacterial Activities of Antineoplastic Agents. Antimicrob. Agents Chemother. 1985, 28, 437. [Google Scholar] [CrossRef]
- Peiris, V.; Oppenheim, B.A. Antimicrobial activity of cytotoxic drugs may influence isolation of bacteria and fungi from blood cultures Influence Isolation. J. Clin. Pathol. 1993, 1124–1125. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, B.; Feng, X.; Rubens, J.R.; Huang, R.; Hicks, L.M.; Pakrasi, H.B.; Tang, Y.J. Alternative Isoleucine Synthesis Pathway in Cyanobacterial Species. Microbiology 2010, 156, 596–602. [Google Scholar] [CrossRef]
- Peng, J.; Xie, J.; Shi, S.; Luo, L.; Li, K.; Xiong, P.; Cai, W. Diagnostic Fragment-Ion-Based for Rapid Identification of Chlorogenic Acids Derivatives in Inula Cappa Using UHPLC-Q-Exactive Orbitrap Mass Spectrometry. J. Anal. Methods Chem. 2021, 2021, 6393246. [Google Scholar] [CrossRef]
- Kumar, R.; Kundu, A.; Dutta, A.; Saha, S.; Das, A.; Bhowmik, A. Chemo-Profiling of Bioactive Metabolites from Chaetomium globosum for Biocontrol of Sclerotinia Rot and Plant Growth Promotion. Fungal Biol. 2021, 125, 167–176. [Google Scholar] [CrossRef]
- Duffield, A.M.; Budzikiewicz, H.; Williams, D.H.; Djerassi, C. Mass Spectrometry in Structural and Stereochemical Problems. LXIV. A Study of the Fragmentation Processes of Some Cyclic Amines. J. Am. Chem. Soc. 1965, 87, 810–816. [Google Scholar] [CrossRef]
- Mahrous, F.; Mohammed, H.; Sabour, R.; Ismail, L. LC-ESI-QTOF-MS/MS of Holoptelea integrifolia (Roxb.) Planch. Leaves and In Silico Study of Phenolic Compounds’ Antiviral Activity against the HSV1 Virus. Azhar Int. J. Pharm. Med. Sci. 2021, 1, 91–101. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Tan, X.M.; Yu, M.; Yang, J.; Sun, B.; Qin, J.C.; Guo, L.P.; Ding, G. Bioactive Metabolites from the Desert Plant-Associated Endophytic Fungus Chaetomium globosum (Chaetomiaceae). Phytochemistry 2021, 185, 112701. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, H.J.; Huang, X.L.; Shao, L.J.; Xie, X.L.; Wang, F.; Yang, J.; Pei, P.; Zhang, Z.Q.; Zhai, Y.C.; et al. A Novel LC-MS/MS Assay for Vitamin B 1, B 2 and B 6 Determination in Dried Blood Spots and Its Application in Children. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1112, 33–40. [Google Scholar] [CrossRef]
- Kanazawa, H.; Atsumi, R.; Matsushima, Y.; Kizu, J. Determination of Theophylline and Its Metabolites in Biological Samples by Liquid Chromatography-Mass Spectrometry. J. Chromatogr. A 2000, 870, 87–96. [Google Scholar] [CrossRef]
- Husain, K.; Abid, M.; Azam, A. Synthesis, Characterization and Antiamoebic Activity of New Indole-3-Carboxaldehyde Thiosemicarbazones and Their Pd(II) Complexes. Eur. J. Med. Chem. 2007, 42, 1300–1308. [Google Scholar] [CrossRef]
- Uenoyama, R.; Miyazaki, M.; Miyazaki, T.; Shigeno, Y.; Tokairin, Y.; Konno, H.; Yamashita, T. LC-ESI-MS/MS Quantification of Carnosine, Anserine, and Balenine in Meat Samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1132, 121826. [Google Scholar] [CrossRef]
- Gachumi, G.; El-Aneed, A. Mass Spectrometric Approaches for the Analysis of Phytosterols in Biological Samples. J. Agric. Food Chem. 2017, 65, 10141–10156. [Google Scholar] [CrossRef]
- Ravindernath, A.; Reddy, M.S. Synthesis and Evaluation of Anti-Inflammatory, Antioxidant and Antimicrobial Activities of Densely Functionalized Novel Benzo [d] Imidazolyl Tetrahydropyridine Carboxylates. Arab. J. Chem. 2017, 10, S1172–S1179. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, J.; Shi, M.; Han, X.; Li, W.; Zhang, X.; Wen, X. Pitaya: A Potential Plant Resource of Citramalic Acid. CyTA-J. Food 2020, 18, 249–256. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Bu, Q.; Zhong, K.; Gao, H. Comparative Study on Antibacterial Mechanism of Shikimic Acid and Quinic Acid against Staphylococcus aureus through Transcriptomic and Metabolomic Approaches. LWT 2022, 153, 112441. [Google Scholar] [CrossRef]
- Eren, B.; Fen, Ü.; Dergisi, B.; Ercan, L.; Doğru, M. Antioxidant and Antimicrobial Capacity of Quinic Acid. Bitlis Eren Üniversitesi Fen Bilim. Derg. 2022, 11, 1018–1025. [Google Scholar] [CrossRef]
- Park, J.H.; Gyung, J.C.; Kyoung, S.J.; He, K.L.; Heung, T.K.; Kwang, Y.C.; Kim, J.C. Antifungal Activity against Plant Pathogenic Fungi of Chaetoviridins Isolated from Chaetomium globosum. FEMS Microbiol. Lett. 2005, 252, 309–313. [Google Scholar] [CrossRef]
- Makrerougras, M.; Coffinier, R.; Oger, S.; Chevalier, A.; Sabot, C.; Franck, X. Total Synthesis and Structural Revision of Chaetoviridins A. Org. Lett. 2017, 19, 4146–4149. [Google Scholar] [CrossRef]
- Awad, N.E.; Kassem, H.A.; Hamed, M.A.; El-Naggar, M.A.A.; El-Feky, A.M.M. Bioassays Guided Isolation of Compounds from Chaetomium globosum. J. Mycol. Med. 2014, 24, e35–e42. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Shi, Y.; Pan, X.; Lu, Y.; Cao, P. Antibacterial Effects of Cinnamon (Cinnamomum zeylanicum) Bark Essential Oil on Porphyromonas gingivalis. Microb. Pathog. 2018, 116, 26–32. [Google Scholar] [CrossRef]
- Bakhtiari, S.; Jafari, S.; Taheri, J.B.; Kashi, T.S.J.; Namazi, Z.; Iman, M.; Poorberafeyi, M. The Effects of Cinnamaldehyde (Cinnamon Derivatives) and Nystatin on Candida albicans and Candida glabrata. Open Access Maced. J. Med. Sci. 2019, 7, 1067–1070. [Google Scholar] [CrossRef]
- Ahgilan, A.; Sabaratnam, V.; Periasamy, V. Antimicrobial Properties of Vitamin B2. Int. J. Food Prop. 2016, 19, 1173–1181. [Google Scholar] [CrossRef]
- Tonnetti, L.; Thorp, A.M.; Reddy, H.L.; Keil, S.D.; Goodrich, R.P.; Leiby, D.A. Riboflavin and Ultraviolet Light Reduce the Infectivity of Babesia Microti in Whole Blood. Transfusion 2013, 53, 860–867. [Google Scholar] [CrossRef]
- Abdel-Lateff, A. Chaetominedione, a New Tyrosine Kinase Inhibitor Isolated from the Algicolous Marine Fungus Chaetomium sp. Tetrahedron Lett. 2008, 49, 6398–6400. [Google Scholar] [CrossRef]
- Xing, L.; Chee, M.E.; Zhang, H.; Zhang, W.; Mine, Y. Carnosine—A Natural Bioactive Dipeptide: Bioaccessibility, Bioavailability and Health Benefits. J. Food Bioact. 2019, 5, 8–17. [Google Scholar] [CrossRef]
- Chmiel, M.; Stompor-Gorący, M. The Spectrum of Pharmacological Actions of Syringetin and Its Natural Derivatives—A Summary Review. Nutrients 2022, 14, 5157. [Google Scholar] [CrossRef]
- Villa-De La Torre, F.; Kinscherf, R.; Bonaterra, G.; Arana-Argaez, V.E.; Méndez-González, M.; Cáceres-Farfán, M.; Borges-Argáez, R. Anti-Inflammatory and Immunomodulatory Effects of Critonia Aromatisans Leaves: Downregulation of pro-Inflammatory Cytokines. J. Ethnopharmacol. 2016, 190, 174–182. [Google Scholar] [CrossRef]
- Wen, S.; He, L.; Zhong, Z.; Zhao, R.; Weng, S.; Mi, H.; Liu, F. Stigmasterol Restores the Balance of Treg/Th17 Cells by Activating the Butyrate-PPARγ Axis in Colitis. Front. Immunol. 2021, 12, 741934. [Google Scholar] [CrossRef]
- Zheng, C.J.; Liao, H.X.; Mei, R.Q.; Huang, G.L.; Yang, L.J.; Zhou, X.M.; Shao, T.M.; Chen, G.Y.; Wang, C.Y. Two New Benzophenones and One New Natural Amide Alkaloid Isolated from a Mangrove-Derived Fungus Penicillium citrinum. Nat. Prod. Res. 2019, 33, 1127–1134. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | Site | GPS | |
|---|---|---|---|
| North | East | ||
| 1- Desert Soil | Saint Katherine Mountain | 28°30′23″ | 34°02′00″ |
| Nuwieba | 28.9°73′44.4″ | 34.6°53′43.3″ | |
| Wadi El-Arbaein | 28°33′00″ | 33°58′11″ | |
| El-Arish | 31°11′10″ | 33°50′25″ | |
| El-Kantara | 30°51′01″ | 32°16′33″ | |
| 2- Cultivated Soil | Ismailia Governorate | 30°40′00″ | 32°20′21″ |
| El-Sharkia Governorate | 30°45′00″ | 31°50′12″ | |
| Assiut Governorate | 27.1°80′96″ | 31.18°36′8″ | |
| Giza Governorate | 30°00′20″ | 31°10′37″ | |
| Cairo Governorate | 30°01′11″ | 31°20′34″ | |
| Host plants | Site | GPS | |
|---|---|---|---|
| North | North | ||
| 1- Medicinal plants in SKP | Gebel Ahmar | 28°52′83″ | 33°61′83″ |
| Wadi El-Arbaein | 28°54′54″ | 33°55′36″ | |
| Wadi Talaa | 28°37′22″ | 33°52′48″ | |
| 2- Mangrove plants | Port Fouad | 31°14′60.00″ | 32°18′60.00″ |
| Safaga | 26°43′59.99″ | 33°55′59.99″ | |
| Nuwieba | 28.9°73′44.4″ | 34.6°53′43.3″ | |
| Code | Strain | Interpretation |
|---|---|---|
| 1 | Chaetomium globosum | Soil taxon recovered from the desert in Nuwieba, South Sinai, Egypt. |
| 2 | Chaetomium globosum | Soil taxon isolated from cultivated rice straw soil, Ismailia Governorate (Suez Canal area), Egypt. |
| 3 | Chaetomium globosum | Soil taxon recovered from cultivated soil in Assiut Governorate (Upper Egypt). |
| 4 | Chaetomium globosum | Endophytic taxon recovered from Thymus decussatus, Gebel Ahmer, Saint Katherine, South Sinai, Egypt. |
| 5 | Chaetomium globosum | Endophytic taxon recovered from Zygophyllum album, Wadi El-Arbaein, Saint Katherine, South Sinai, Egypt. |
| 6 | Chaetomium globosum | Endophytic taxon recovered from Chiliadenus montanus, Wadi El-Arbaein, Saint Katherine, South Sinai, Egypt. |
| 7 | Chaetomium iranianum | Endophytic taxon recovered from Origanum syriacum, Wadi El-Arbaein, Saint Katherine, South Sinai, Egypt. |
| 8 | Chaetomium globosum | Endophytic taxon recovered from Adiantum capillus-veneris, Wadi Talaa, Saint Katherine, South Sinai, Egypt. |
| 9 | Chaetomium globosum | Endophytic taxon recovered from Adiantum capillus-veneris, Port Fouad, Port Said Governorate (Mediterranean area), Egypt. |
| 10 | Chaetomium madrasense | Endophytic taxon recovered from Ziziphus spinosa, Nuwieba, South Sinai, Egypt. |
| Organisms | No of Isolates |
|---|---|
| Klebsiella pneumoniea | 30 |
| Staphylococcus aureus | 21 |
| Escherichia coli | 11 |
| Pseudomonas aeruginosa | 9 |
| Candida albicans | 8 |
| Acinetobacter baumannii | 7 |
| Enterococcus faecalis | 5 |
| Proteus mirabilis | 5 |
| Streptococcus pyogenes | 2 |
| Serratia marcescens | 2 |
| Total | 100 |
| Antibiotics’ Classes | Antibiotic Discs | S. aureus | E. faecalis | S. pyogenes | A. baumannii | E. coli | K. pneumoniea | Pr. mirabilis | P. aeruginosa | S. marcescens |
|---|---|---|---|---|---|---|---|---|---|---|
| Penicillin | Amoxycillin/Clavulanic acid (30 µg) | 100 | 60 | 100 | 57.1 | 90.9 | 100 | 60 | 100 | 100 |
| Ampicillin/Sulbactam (30 µg) | 100 | 100 | 100 | 85.7 | 90.9 | 100 | 80 | 88.9 | 100 | |
| Cephalosporin | Cefepime (30 µg) | 100 | 100 | 100 | 85.7 | 54.5 | 70 | 60 | 100 | 50 |
| Cefotaxime (10 µg) | 100 | 100 | 100 | 71.4 | 81.8 | 90 | 100 | 100 | 50 | |
| Ceftazidime (30 µg) | 80.9 | 100 | 100 | 85.7 | 90.9 | 73.3 | 60 | 66.7 | 100 | |
| Cefoxitin (30 µg) | 0 | 100 | 100 | 100 | 100 | 90 | 100 | 88.9 | 50 | |
| Fluoroquinolone | Ciprofloxacin (5 µg) | 71.4 | 80 | 50 | 85.7 | 81.8 | 80 | 80 | 88.9 | 50 |
| Levofloxacin (5 µg) | 76.2 | 100 | 50 | 57.1 | 54.5 | 100 | 80 | 88.9 | 100 | |
| Aminoglycosides | Gentamicin (10 µg) | 66.7 | 100 | 100 | 71.4 | 90.9 | 70 | 80 | 55.6 | 100 |
| Amikacin (30 µg) | 76.2 | 100 | 50 | 71.4 | 54.5 | 73.3 | 40 | 66.7 | 50 | |
| Carbapenem | Imipenem (10 µg) | 28.6 | 100 | 0 | 14.3 | 63.6 | 30 | 20 | 33.3 | 0 |
| Meropenem (10 µg) | 33.3 | 100 | 0 | 28.6 | 63.6 | 33.3 | 20 | 11.1 | 0 | |
| Tetracycline | Tetracycline (30 µg) | 38.0 | 60 | 50.0 | 57.1 | 63.6 | 80 | 60 | 100 | 50 |
| Doxycycline (5 µg) | 76.2 | 80 | 50 | 100 | 72.7 | 60 | 60 | 100 | 50 | |
| Tigecycline (15 µg) | 19.0 | 0 | 0 | 14.3 | 36.4 | 60 | 20 | 33.3 | 0 |
| Crude Extracts | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Microorganism | Mean of Zone Inhibition in mm (Mean ± SD) | |||||||||
| Staphylococcus aureus | 0 | 0 | 19.5 ± 1.36 | 18.4 ± 1.4 | 17.2 ± 1.48 | 16.0 ± 1.28 | 0 | 14.8 ± 0.77 | 0 | 25.7 ± 1.23 |
| Enterococcus faecalis | 0 | 14.6 ± 0.55 | 0 | 0 | 0 | 10.4 ± 0.55 | 0 | 0 | 0 | 11.6 ± 0.55 |
| Streptococcus pyogenes | 0 | 0 | 21.0 ± 0.0 | 14.5 ± 0.7 | 0 | 15.0 ± 0.0 | 0 | 0 | 0 | 26.0 ± 1.4 |
| Acinetobacter baumannii | 0 | 0 | 0 | 0 | 0 | 15.4 ± 1.27 | 0 | 0 | 0 | 0 |
| Escherichia coli | 21.2 ± 2.09 | 19.8 ± 2.09 | 24.0 ± 2.19 | 16.0 ± 1.48 | 25.4 ± 2.46 | 15.4 ± 1.12 | 18.6 ± 2.46 | 15.4 ± 1.03 | 16.3 ± 0.65 | 10.5 ± 0.93 |
| Klebsiella pneumoniea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Proteus mirabilis | 0 | 0 | 0 | 0 | 21.6 ± 1.34 | 0 | 0 | 0 | 19.6 ± 1.34 | 0 |
| Pseudomonas aeruginosa | 26.0 ± 0.87 | 25.4 ± 1.24 | 26.3 ± 0.5 | 17.0 ± 1.1 | 21.7 ± 2.5 | 16.3 ± 1.1 | 25.2 ± 1.3 | 17.3 ± 0.7 | 21.7 ± 2.0 | 18.4 ± 1.5 |
| Serratia marcescens | 18.0 ± 0.0 | 16.5 ± 0.7 | 22.5 ± 2.1 | 14.5 ± 0.7 | 24.0 ± 0.0 | 0 | 26.0 ± 0.0 | 0 | 26.5 ± 0.7 | 12.0 ± 0.0 |
| Candida albicans | 0 | 0 | 24.5 ± 0.76 | 18.8 ± 1.49 | 25.6 ± 0.74 | 0 | 0 | 11.3 ± 0.7 | 0 | 15.5 ± 0.76 |
| Crude Extracts | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Microorganism | MIC µg/mL (Mean ± SD) | |||||||||
| Staphylococcus aureus | - | - | 29.8 ± 4.7 | 5.0 ± 3.5 | 30.5 ± 3.4 | 62.5 ± 0.0 | - | 15.0 ± 2.6 | - | 3.9 ± 0.0 |
| Enterococcus faecalis | - | 14.0 ± 3.5 | - | - | - | 15.6 ± 0.0 | - | - | - | 31.2 ± 0.0 |
| Streptococcus pyogenes | - | - | 31.2 ± 0.0 | 15.6 ± 0.0 | - | 15.6 ± 0.0 | - | - | - | 3.9 ± 0.0 |
| Acinetobacter baumannii | - | - | - | - | - | 31.2 ± 0.0 | - | - | - | - |
| Escherichia coli | 4.6 ± 1.6 | 8.2 ± 3.5 | 5.7 ± 2.0 | 31.2 ± 0.0 | 5.0 ± 1.8 | 29.8 ± 4.7 | 14.2 ± 3.2 | 12.8 ± 3.9 | 15.6 ± 0.0 | 14.2 ± 3.1 |
| Klebsiella pneumoniea | - | - | - | - | - | - | - | - | - | - |
| Proteus mirabilis | - | - | - | - | 5.5 ± 2.1 | - | - | - | 7.0 ± 1.7 | - |
| Pseudomonas aeruginosa | 4.3 ± 1.3 | 4.7 ± 1.7 | 3.9 ± 0.0 | 8.7 ± 2.6 | 5.2 ± 1.9 | 14.7 ± 2.6 | 6.0 ± 3.9 | 8.7 ± 2.6 | 15.6 ± 0.0 | 9.5 ± 3.4 |
| Serratia marcescens | 15.6 ± 0.0 | 15.6 ± 0.0 | 3.9 ± 0.0 | 15.6 ± 0.0 | 3.9 ± 0.0 | - | 3.9 ± 0.0 | - | 3.9 ± 0.0 | 31.2 ± 0.0 |
| Candida albicans | - | - | 3.9 ± 0.0 | 8.8 ± 2.7 | 4.9 ± 1.8 | - | - | 31.2 ± 0.0 | - | 14.6 ± 2.8 |
| No | Polarity Mode | MZmine ID | Ret. Time (min) | Observed m/z | Calculated m/z | Mass Error (ppm) | Adduct | Molecular Formula | MS/MS Spectrum | Deduced Compound | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Negative | 522 | 1.10 | 173.0448 | 173.0450 | −1.16 | [M − H] − | C7H10O5 | 173, 155, 111 | Shikimic acid | [54] |
| 2 | Positive | 201 | 1.11 | 319.1526 | 319.1545 | −5.95 | [M + H] + | C18H22O5 | 319, 301, 283, 230 | Zearalenone | [55] |
| 3 | Negative | 663 | 1.15 | 329.0897 | 329.0862 | 10.64 | [M − H] − | C14H18O9 | 329, 167 | Vanillic acid hexoside | [56,57] |
| 4 | Positive | 885 | 1.34 | 138.0549 | 138.0555 | −4.35 | [M + H] + | C7H7NO2 | 138, 94, 92 | Trigonelline | [58] |
| 5 | Positive | 1116 | 1.38 | 116.071 | 116.0712 | −1.72 | [M + H] + | C5H9NO2 | 116, 70 | L-Proline | [59] |
| 6 | Negative | 1889 | 1.41 | 309.1176 | 309.1200 | −7.76 | [M − H] − | C12H22O9 | 309, 89 | Hex-2-ulofuranosyl-4,6- dideoxyhexopyranoside | [60] |
| 7 | Positive | 1730 | 1.76 | 297.1083 | 297.1099 | −5.39 | [M + Na] + | C16H18O4 | 297, 274 | Prochaetoviridin A | [61] |
| 8 | Positive | 1924 | 1.89 | 123.0544 | 123.0558 | −11.38 | [M + H] + | C6H6N2O | 123, 80 | Nicotinamide (Niacinamide) | [62] |
| 9 | Negative | 2601 | 3.71 | 351.0866 | 351.0869 | −0.85 | [M − H] − | C20H16O6 | 351, 263 | Altertoxin I | [63] |
| 10 | Positive | 2529 | 4.94 | 314.1368 | 314.1381 | −4.14 | [M + H] + | C18H19NO4 | 314, 283, 268, 251, 233, 223 | Demethyl isoboldine | [64] |
| 11 | Positive | 2541 | 5.02 | 286.1434 | 286.1443 | −3.15 | [M + H] + | C17H19NO3 | 286, 269, 237, 209, 175, 145, 143, 107 | Coclaurine | [64] |
| 12 | Positive | 2609 | 5.34 | 328.156 | 328.1549 | 3.35 | [M + H] + | C19H21NO4 | 328, 178, 151 | Scoulerine | [65] |
| 13 | Positive | 2687 | 5.66 | 545.2042 | 545.2052 | −1.83 | [M + Na] + | C32H30N2O5 | 522.60 | Chaetoglobinol B | [66] |
| 14 | Positive | 2747 | 6.08 | 314.1765 | 314.1763 | 0.64 | [M + H] + | C19H24NO3+ | 314, 299, 298, 269, 237, 209, 175, 121, 107 | N,N-Dimethyl coclaurine | [64] |
| 15 | Positive | 2777 | 6.18 | 500.1878 | 500.1840 | 7.60 | [M + H] + | C27H30ClO6N | 500 | Chaetomugilide A | [67,68] |
| 16 | Positive | 2887 | 6.50 | 523.219 | 523.2233 | −8.22 | [M + H] + | C32H30N2O5 | 523 | Chaetoglobinol A | [66,69] |
| 17 | Positive | 2883 | 6.50 | 300.1592 | 300.1600 | −2.67 | [M + H] + | C18H21NO3 | 300, 269, 237, 209, 177, 175, 145, 107 | N-Methyl coclaurine | [65] |
| 18 | Positive | 2916 | 6.67 | 496.2383 | 496.2335 | 9.67 | [M + H] + | C28H33NO7 | 496 | Penochalasin A | [69] |
| 19 | Positive | 2921 | 6.69 | 548.2684 | 548.2648 | 6.57 | [M + H] + | C32H37NO7 | 548 | Cytochalasin L | [70] |
| 20 | Negative | 2847 | 6.84 | 523.2174 | 523.2177 | −0.57 | [M − H] − | C26H36O11 | 523, 361, 346 | Secoisolariciresinol -β-D-hexoside | [56] |
| 21 | Positive | 3216 | 8.95 | 391.1363 | 391.1312 | 13.04 | [M + H] + | C21H23ClO5 | 391, 363, 147 | Sclerotiorin | [71] |
| 22 | Positive | 3337 | 13.47 | 485.2115 | 485.2175 | 12.37 | [M + H] + | C27H32O8 | 485 | Verrucarin J | [72] |
| No | Polarity Mode | MZmine ID | Ret. Time (min) | Obseved m/z | Calculated m/z | Mass Error (ppm) | Adduct | Molecular Formula | MS/MS Spectrum | Deduced Compound | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Negative | 56 | 1.08 | 147.0313 | 147.0293 | 13.60 | [M − H] − | C5H8O5 | 147, 85, 57 | Citramalic acid | [98] |
| 2 | Positive | 361 | 1.12 | 319.1508 | 319.1545 | −11.59 | [M + H] + | C18H22O5 | 319, 301, 283, 230 | Zearalenone | [55] |
| 3 | Negative | 207 | 1.12 | 191.0542 | 191.0556 | −7.33 | [M − H] − | C7H12O6 | 191, 173, 111, 85 | Quinic acid | [99] |
| 4 | Positive | 1098 | 1.37 | 433.1741 | 433.1762 | −4.85 | [M + H] + | C23H25ClO6 | 433, 296 | Chaetoviridin A | [100] |
| 5 | Positive | 1322 | 1.40 | 138.0553 | 138.0555 | −1.45 | [M + H] + | C7H7NO2 | 138, 94, 92 | Trigonelline | [58] |
| 6 | Negative | 2534 | 1.77 | 309.1176 | 309.1200 | −7.76 | [M − H] − | C12H22O9 | 309, 179, 119, 89 | Hex-2-ulofuranosyl-4,6- dideoxyhexopyranoside | [60] |
| 7 | Positive | 1941 | 1.78 | 86.09572 | 86.0970 | 14.87 | [M + H] + | C5H11N | 86, 56 | Piperidine | [101] |
| 8 | Positive | 2050 | 1.80 | 133.1041 | 133.1023 | 13.52 | [M + H] + | C9H8O | 133, 115, 105, 79 | Cinnamaldehyde | [102] |
| 9 | Positive | 2298 | 1.90 | 402.1937 | 402.1919 | 4.48 | [M + H] + | C22H27NO6 | 402 | Globosumin | [103] |
| 10 | Positive | 2811 | 3.59 | 286.1442 | 286.1443 | −0.35 | [M + H] + | C17H19NO3 | 286, 269, 237, 175, 145, 143, 107 | Coclaurine | [64] |
| 11 | Positive | 2959 | 4.41 | 314.1372 | 314.1381 | −2.87 | [M + H] + | C18H19NO4 | 314, 283, 268, 251, 233, 223 | Demethyl isoboldine | [64] |
| 12 | Positive | 3163 | 5.30 | 377.149 | 377.1461 | 7.69 | [M + H] + | C17H20N4O6 | 377, 243 | Riboflavin | [104] |
| 13 | Positive | 3358 | 6.07 | 314.1752 | 314.1763 | −3.50 | [M + H] + | C19H24NO3+ | 413, 299, 298, 269, 209, 175, 121, 107 | N,N-Dimethyl coclaurine | [64] |
| 14 | Positive | 3449 | 6.42 | 197.07 | 196.0675 | 12.69 | [M + H] + | C7H8N4O3 | 197, 169 | 1,3-Dimethyl urate | [105] |
| 15 | Positive | 3505 | 6.54 | 300.1621 | 300.1600 | 6.99 | [M + H] + | C18H21NO3 | 300, 269, 237, 209, 177, 175, 145, 107 | N-Methyl coclaurine | [65] |
| 16 | Positive | 4060 | 8.10 | 146.0613 | 146.0606 | 4.79 | [M + H] + | C9H7NO | 146, 117 | 3-Formylindole | [106] |
| 17 | Positive | 4609 | 10.15 | 331.0695 | 331.0695 | 0 | [M + Na] + | C17H12N2O4 | 331, 308 | Chaetominedione | [69] |
| 18 | Positive | 4620 | 10.40 | 227.1166 | 227.1144 | 9.69 | [M + H] + | C9H14N4O3 | 227, 181, 156, | Carnosine | [107] |
| 19 | Positive | 4628 | 10.51 | 509.2021 | 509.2022 | −0.20 | [M + H] + | C23H24O13 | 509, 347 | Syringetin 3-O-galactoside | [102] |
| 20 | Positive | 5497 | 22.47 | 395.3634 | 395.3678 | −11.13 | [M + H − H2O] + | C29H48O | 395, 147 | Stigmasterol | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goda, M.S.; El-Kattan, N.; Abdel-Azeem, M.A.; Allam, K.A.M.; Badr, J.M.; Nassar, N.A.; Almalki, A.J.; Alharbi, M.; Elhady, S.S.; Eltamany, E.E. Antimicrobial Potential of Different Isolates of Chaetomium globosum Combined with Liquid Chromatography Tandem Mass Spectrometry Chemical Profiling. Biomolecules 2023, 13, 1683. https://doi.org/10.3390/biom13121683
Goda MS, El-Kattan N, Abdel-Azeem MA, Allam KAM, Badr JM, Nassar NA, Almalki AJ, Alharbi M, Elhady SS, Eltamany EE. Antimicrobial Potential of Different Isolates of Chaetomium globosum Combined with Liquid Chromatography Tandem Mass Spectrometry Chemical Profiling. Biomolecules. 2023; 13(12):1683. https://doi.org/10.3390/biom13121683
Chicago/Turabian StyleGoda, Marwa S., Noura El-Kattan, Mohamed A. Abdel-Azeem, Kamilia A. M. Allam, Jihan M. Badr, Nourelhuda Ahmed Nassar, Ahmad J. Almalki, Majed Alharbi, Sameh S. Elhady, and Enas E. Eltamany. 2023. "Antimicrobial Potential of Different Isolates of Chaetomium globosum Combined with Liquid Chromatography Tandem Mass Spectrometry Chemical Profiling" Biomolecules 13, no. 12: 1683. https://doi.org/10.3390/biom13121683