
Regulation of Syntaxin3B-Mediated Membrane Fusion by T14, Munc18, and Complexin

{kind=link}
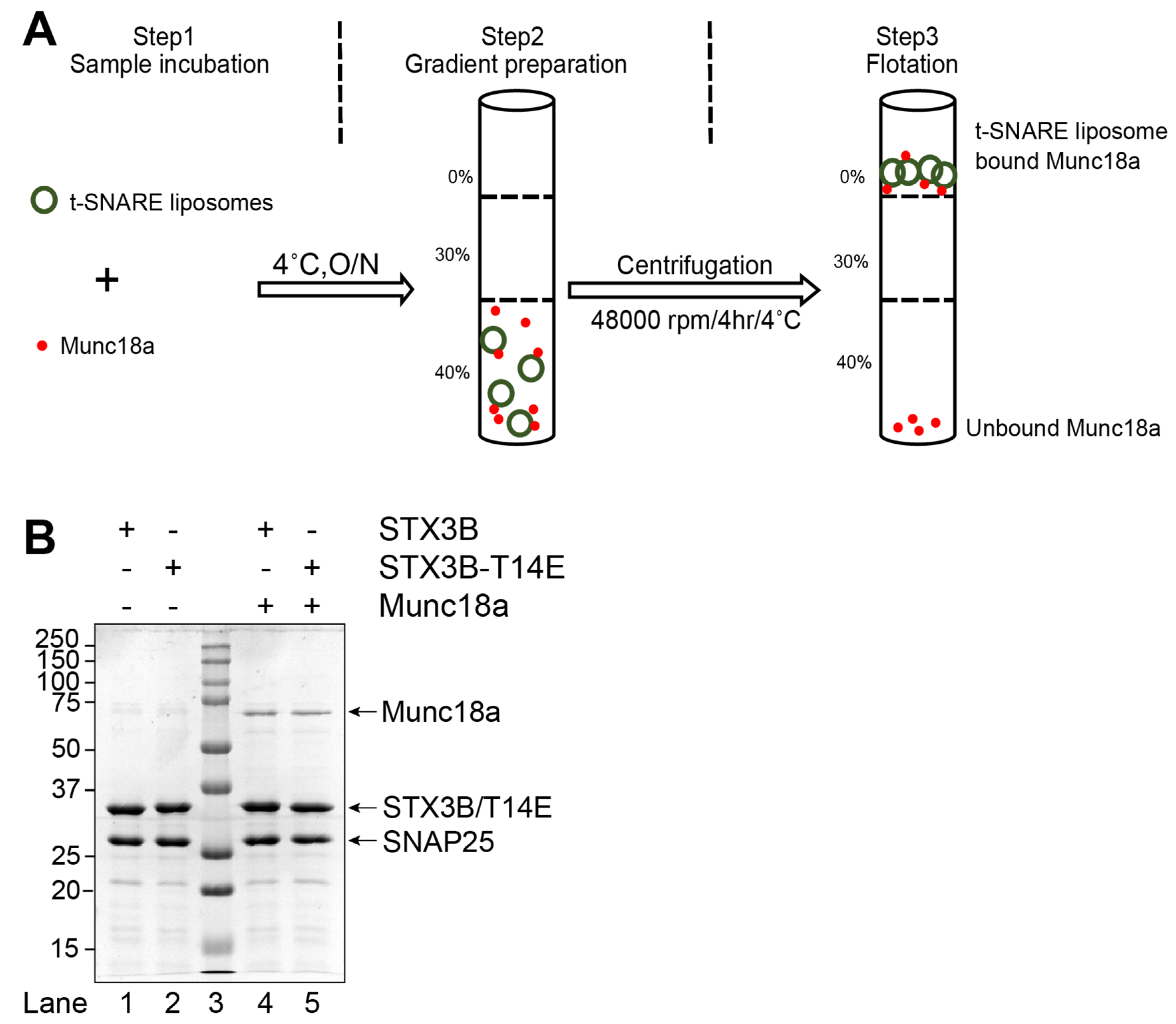{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Biology
2.2. Site-Directed Mutagenesis
2.3. Protein Expression and Purification
2.3.1. Syntaxin 3B and His6SNAP-25
2.3.2. VAMP2
2.3.3. Munc18a
2.3.4. Complexin III and IV
2.4. Liposome Production and Reconstitution
2.5. Gradient Preparation and Flotation
2.6. In Vitro Lipid Mixing Assays
2.7. Statistical Analyses
3. Results
3.1. Syntaxin3B with a Phosphomimetic Mutation at T14 Enhances Membrane Fusion
3.2. Munc18a Interacts with Membrane-Bound t-SNARE Complexes Containing Syntaxin3B
3.3. Syntaxin3B, with a Phosphomimetic Mutation at T14, Exhibits Enhanced Membrane Fusion in the Presence of Munc18a
3.4. Complexin III and Complexin IV Regulate Membrane Fusion Catalyzed by Syntaxin3B-Containing SNARE Complexes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgans, C.W.; Brandstatter, J.H.; Kellerman, J.; Betz, H.; Wassle, H. A SNARE complex containing syntaxin 3 is present in ribbon synapses of the retina. J. Neurosci. 1996, 16, 6713–6721. [Google Scholar] [CrossRef] [PubMed]
- Sherry, D.M.; Mitchell, R.; Standifer, K.M.; du Plessis, B. Distribution of plasma membrane-associated syntaxins 1 through 4 indicates distinct trafficking functions in the synaptic layers of the mouse retina. BMC Neurosci. 2006, 7, 54. [Google Scholar] [CrossRef]
- Curtis, L.B.; Doneske, B.; Liu, X.; Thaller, C.; McNew, J.A.; Janz, R. Syntaxin 3b is a t-SNARE specific for ribbon synapses of the retina. J. Comp. Neurol. 2008, 510, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Curtis, L.; Datta, P.; Liu, X.; Bogdanova, N.; Heidelberger, R.; Janz, R. Syntaxin 3B is essential for the exocytosis of synaptic vesicles in ribbon synapses of the retina. Neuroscience 2010, 166, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Gilliam, J.; Thoreson, W.B.; Janz, R.; Heidelberger, R. Two Pools of Vesicles Associated with Synaptic Ribbons Are Molecularly Prepared for Release. Biophys. J. 2017, 113, 2281–2298. [Google Scholar] [CrossRef]
- Janecke, A.R.; Liu, X.; Adam, R.; Punuru, S.; Viestenz, A.; Strauss, V.; Laass, M.; Sanchez, E.; Adachi, R.; Schatz, M.P.; et al. Pathogenic STX3 variants affecting the retinal and intestinal transcripts cause an early-onset severe retinal dystrophy in microvillus inclusion disease subjects. Hum. Genet. 2021, 140, 1143–1156. [Google Scholar] [CrossRef]
- Kakakhel, M.; Tebbe, L.; Makia, M.S.; Conley, S.M.; Sherry, D.M.; Al-Ubaidi, M.R.; Naash, M.I. Syntaxin 3 is essential for photoreceptor outer segment protein trafficking and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 20615–20624. [Google Scholar] [CrossRef]
- Campbell, J.R.; Li, H.; Wang, Y.; Kozhemyakin, M.; Hunt, A.J., Jr.; Liu, X.; Janz, R.; Heidelberger, R. Phosphorylation of the Retinal Ribbon Synapse Specific t-SNARE Protein Syntaxin3B Is Regulated by Light via a Ca(2+)-Dependent Pathway. Front. Cell. Neurosci. 2020, 14, 587072. [Google Scholar] [CrossRef]
- Liu, X.; Heidelberger, R.; Janz, R. Phosphorylation of syntaxin 3B by CaMKII regulates the formation of t-SNARE complexes. Mol. Cell. Neurosci. 2014, 60, 53–62. [Google Scholar] [CrossRef]
- Ma, C.; Su, L.; Seven, A.B.; Xu, Y.; Rizo, J. Reconstitution of the vital functions of Munc18 and Munc13 in neurotransmitter release. Science 2013, 339, 421–425. [Google Scholar] [CrossRef]
- Wang, S.; Ma, C. Neuronal SNARE complex assembly guided by Munc18-1 and Munc13-1. FEBS Open Bio 2022, 12, 1939–1957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hughson, F.M. Chaperoning SNARE Folding and Assembly. Annu. Rev. Biochem. 2021, 90, 581–603. [Google Scholar] [CrossRef] [PubMed]
- Gething, C.; Ferrar, J.; Misra, B.; Howells, G.; Andrzejewski, A.L.; Bowen, M.E.; Choi, U.B. Conformational change of Syntaxin-3b in regulating SNARE complex assembly in the ribbon synapses. Sci. Rep. 2022, 12, 9261. [Google Scholar] [CrossRef]
- Cooper, B.; Hemmerlein, M.; Ammermuller, J.; Imig, C.; Reim, K.; Lipstein, N.; Kalla, S.; Kawabe, H.; Brose, N.; Brandstatter, J.H.; et al. Munc13-independent vesicle priming at mouse photoreceptor ribbon synapses. J. Neurosci. 2012, 32, 8040–8052. [Google Scholar] [CrossRef] [PubMed]
- Risinger, C.; Bennett, M.K. Differential phosphorylation of syntaxin and synaptosome-associated protein of 25 kDa (SNAP-25) isoforms. J. Neurochem. 1999, 72, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Rathore, S.S.; Khandan, L.; Rothman, J.E. SNARE bundle and syntaxin N-peptide constitute a minimal complement for Munc18-1 activation of membrane fusion. J. Cell Biol. 2010, 190, 55. [Google Scholar] [CrossRef] [PubMed]
- Rickman, C.; Duncan, R.R. Munc18/Syntaxin interaction kinetics control secretory vesicle dynamics. J. Biol. Chem. 2009, 285, 3965–3972. [Google Scholar] [CrossRef]
- Burkhardt, P.; Hattendorf, D.A.; Weis, W.I.; Fasshauer, D. Munc18a controls SNARE assembly through its interaction with the syntaxin N-peptide. EMBO J. 2008, 27, 923–933. [Google Scholar] [CrossRef]
- Colbert, K.N.; Hattendorf, D.A.; Weiss, T.M.; Burkhardt, P.; Fasshauer, D.; Weis, W.I. Syntaxin1a variants lacking an N-peptide or bearing the LE mutation bind to Munc18a in a closed conformation. Proc. Natl. Acad. Sci. USA 2013, 110, 12637–12642. [Google Scholar] [CrossRef]
- Christie, M.P.; Whitten, A.E.; King, G.J.; Hu, S.-H.; Jarrott, R.J.; Chen, K.-E.; Duff, A.P.; Callow, P.; Collins, B.M.; James, D.E.; et al. Low-resolution solution structures of Munc18:Syntaxin protein complexes indicate an open binding mode driven by the Syntaxin N-peptide. Proc. Natl. Acad. Sci. USA 2012, 109, 9816. [Google Scholar] [CrossRef]
- Shen, J.; Tareste, D.C.; Paumet, F.; Rothman, J.E.; Melia, T.J. Selective activation of cognate SNAREpins by Sec1/Munc18 proteins. Cell 2007, 128, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Choi, U.B.; Leitz, J.; Rhee, H.J.; Lee, C.; Altas, B.; Zhao, M.; Pfuetzner, R.A.; Wang, A.L.; Brose, N.; et al. Molecular Mechanisms of Synaptic Vesicle Priming by Munc13 and Munc18. Neuron 2017, 95, 591–607.e10. [Google Scholar] [CrossRef] [PubMed]
- Stepien, K.P.; Xu, J.; Zhang, X.; Bai, X.C.; Rizo, J. SNARE assembly enlightened by cryo-EM structures of a synaptobrevin-Munc18-1-syntaxin-1 complex. Sci. Adv. 2022, 8, eabo5272. [Google Scholar] [CrossRef]
- Rodkey, T.L.; Liu, S.; Barry, M.; McNew, J.A. Munc18a scaffolds SNARE assembly to promote membrane fusion. Mol. Biol. Cell 2008, 19, 5422–5434. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.L.; Van Komen, J.S.; Liu, S.; Weber, T.; Melia, T.J.; McNew, J.A. Liposome fusion assay to monitor intracellular membrane fusion machines. Methods Enzymol. 2003, 372, 274–300. [Google Scholar] [CrossRef]
- Moser, T.; Grabner, C.P.; Schmitz, F. Sensory Processing at Ribbon Synapses in the Retina and the Cochlea. Physiol. Rev. 2020, 100, 103–144. [Google Scholar] [CrossRef]
- Sokal, I.; Haeseleer, F. Insight into the role of Ca2+-binding protein 5 in vesicle exocytosis. Invest. Ophthalmol. Vis. Sci. 2011, 52, 9131–9141. [Google Scholar] [CrossRef]
- Ullrich, B.; Sudhof, T.C. Distribution of synaptic markers in the retina: Implications for synaptic vesicle traffic in ribbon synapses. J. Physiol. Paris. 1994, 88, 249–257. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, L.; Wang, L.; Wang, C.; Liu, C.; Guo, A.; Liu, M.; Zhang, L.; Ma, C.; Zhang, X.; et al. p.His16Arg of STXBP1 (MUNC18-1) Associated With Syntaxin 3B Causes Autosomal Dominant Congenital Nystagmus. Front. Cell Dev. Biol. 2020, 8, 591781. [Google Scholar] [CrossRef]
- Tellam, J.T.; Macaulay, S.L.; McIntosh, S.; Hewish, D.R.; Ward, C.W.; James, D.E. Characterization of Munc-18c and syntaxin-4 in 3T3-L1 adipocytes. Putative role in insulin-dependent movement of GLUT-4. J. Biol. Chem. 1997, 272, 6179–6186. [Google Scholar] [CrossRef]
- Hata, Y.; Slaughter, C.A.; Sudhof, T.C. Synaptic vesicle fusion complex contains unc-18 homologue bound to syntaxin. Nature 1993, 366, 347–351. [Google Scholar] [CrossRef]
- Pevsner, J.; Hsu, S.C.; Scheller, R.H. n-Sec1: A neural-specific syntaxin-binding protein. Proc. Natl. Acad. Sci. USA 1994, 91, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Verhage, M.; Maia, A.S.; Plomp, J.J.; Brussaard, A.B.; Heeroma, J.H.; Vermeer, H.; Toonen, R.F.; Hammer, R.E.; van den Berg, T.K.; Missler, M.; et al. Synaptic assembly of the brain in the absence of neurotransmitter secretion. Science 2000, 287, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tomchick, D.R.; Kovrigin, E.; Arac, D.; Machius, M.; Sudhof, T.C.; Rizo, J. Three-dimensional structure of the complexin/SNARE complex. Neuron 2002, 33, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Reim, K.; Mansour, M.; Varoqueaux, F.; McMahon, H.T.; Sudhof, T.C.; Brose, N.; Rosenmund, C. Complexins regulate a late step in Ca2+-dependent neurotransmitter release. Cell 2001, 104, 71–81. [Google Scholar] [CrossRef]
- Schaub, J.R.; Lu, X.; Doneske, B.; Shin, Y.K.; McNew, J.A. Hemifusion arrest by complexin is relieved by Ca2+-synaptotagmin I. Nat. Struct. Mol. Biol. 2006, 13, 748–750. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, F.; Su, Z.; McNew, J.A.; Shin, Y.K. Hemifusion in SNARE-mediated membrane fusion. Nat. Struct. Mol. Biol. 2005, 12, 417–422. [Google Scholar] [CrossRef]
- Reim, K.; Wegmeyer, H.; Brandstatter, J.H.; Xue, M.; Rosenmund, C.; Dresbach, T.; Hofmann, K.; Brose, N. Structurally and functionally unique complexins at retinal ribbon synapses. J. Cell Biol. 2005, 169, 669–680. [Google Scholar] [CrossRef]
- Bhoi, J.D.; Zhang, Z.; Janz, R.; You, Y.; Wei, H.; Wu, J.; Ribelayga, C.P. The SNARE regulator Complexin 3 is a target of the cone circadian clock. J. Comp. Neurol. 2021, 529, 1066–1080. [Google Scholar] [CrossRef]
- Babai, N.; Sendelbeck, A.; Regus-Leidig, H.; Fuchs, M.; Mertins, J.; Reim, K.; Brose, N.; Feigenspan, A.; Brandstatter, J.H. Functional Roles of Complexin 3 and Complexin 4 at Mouse Photoreceptor Ribbon Synapses. J. Neurosci. 2016, 36, 6651–6667. [Google Scholar] [CrossRef]
- Mortensen, L.S.; Park, S.J.H.; Ke, J.B.; Cooper, B.H.; Zhang, L.; Imig, C.; Lowel, S.; Reim, K.; Brose, N.; Demb, J.B.; et al. Complexin 3 Increases the Fidelity of Signaling in a Retinal Circuit by Regulating Exocytosis at Ribbon Synapses. Cell Rep. 2016, 15, 2239–2250. [Google Scholar] [CrossRef] [PubMed]
- Vaithianathan, T.; Henry, D.; Akmentin, W.; Matthews, G. Functional roles of complexin in neurotransmitter release at ribbon synapses of mouse retinal bipolar neurons. J. Neurosci. 2015, 35, 4065–4070. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Zemelman, B.V.; McNew, J.A.; Westermann, B.; Gmachl, M.; Parlati, F.; Sollner, T.H.; Rothman, J.E. SNAREpins: Minimal machinery for membrane fusion. Cell 1998, 92, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Betancourt-Solis, M.A.; McNew, J.A. Detergent-Assisted Reconstitution of Recombinant Drosophila Atlastin into Liposomes for Lipid-Mixing Assays. J. Vis. Exp. 2019, 149, e59867. [Google Scholar] [CrossRef]
- Orso, G.; Pendin, D.; Liu, S.; Tosetto, J.; Moss, T.J.; Faust, J.E.; Micaroni, M.; Egorova, A.; Martinuzzi, A.; McNew, J.A.; et al. Homotypic fusion of ER membranes requires the dynamin-like GTPase atlastin. Nature 2009, 460, 978–983. [Google Scholar] [CrossRef]
- Trimbuch, T.; Rosenmund, C. Should I stop or should I go? The role of complexin in neurotransmitter release. Nat. Rev. Neurosci. 2016, 17, 118–125. [Google Scholar] [CrossRef]
- Brunger, A.T.; Leitz, J. The Core Complex of the Ca(2+)-Triggered Presynaptic Fusion Machinery. J. Mol. Biol. 2023, 435, 167853. [Google Scholar] [CrossRef]
- Rizo, J. Molecular Mechanisms Underlying Neurotransmitter Release. Annu. Rev. Biophys. 2022, 51, 377–408. [Google Scholar] [CrossRef]
- Parlati, F.; Weber, T.; McNew, J.A.; Westermann, B.; Söllner, T.H.; Rothman, J.E. Rapid and efficient fusion of phospholipid vesicles by the alpha-helical core of a SNARE complex in the absence of an N-terminal regulatory domain. Proc. Natl. Acad. Sci. USA 1999, 96, 12565–12570. [Google Scholar] [CrossRef]
- Schaffner, W.; Weissmann, C. A rapid, sensitive, and specific method for the determination of protein in dilute solution. Anal. Biochem. 1973, 56, 502–514. [Google Scholar] [CrossRef]
- Lai, Y.; Diao, J.; Cipriano, D.J.; Zhang, Y.; Pfuetzner, R.A.; Padolina, M.S.; Brunger, A.T. Complexin inhibits spontaneous release and synchronizes Ca2+-triggered synaptic vesicle fusion by distinct mechanisms. eLife 2014, 3, e03756. [Google Scholar] [CrossRef]
- Rigaud, J.L.; Levy, D. Reconstitution of membrane proteins into liposomes. Methods Enzymol. 2003, 372, 65–86. [Google Scholar] [CrossRef]
- Sanchez, E.; Gonzalez, E.A.; Moreno, D.S.; Cardenas, R.A.; Ramos, M.A.; Davalos, A.J.; Manllo, J.; Rodarte, A.I.; Petrova, Y.; Moreira, D.C.; et al. Syntaxin 3, but not syntaxin 4, is required for mast cell-regulated exocytosis, where it plays a primary role mediating compound exocytosis. J. Biol. Chem. 2019, 294, 3012–3023. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, S.; Riento, K.; Olkkonen, V.M.; Lehtonen, E. Syntaxin 3 and Munc-18-2 in epithelial cells during kidney development. Kidney Int. 1999, 56, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Torkko, J.M.; Manninen, A.; Schuck, S.; Simons, K. Depletion of apical transport proteins perturbs epithelial cyst formation and ciliogenesis. J. Cell Sci. 2008, 121, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Koo, E.; Kwan, E.; Kang, Y.; Park, S.; Xie, H.; Sugita, S.; Gaisano, H.Y. Syntaxin-3 regulates newcomer insulin granule exocytosis and compound fusion in pancreatic beta cells. Diabetologia 2013, 56, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Brochetta, C.; Suzuki, R.; Vita, F.; Soranzo, M.R.; Claver, J.; Madjene, L.C.; Attout, T.; Vitte, J.; Varin-Blank, N.; Zabucchi, G.; et al. Munc18-2 and syntaxin 3 control distinct essential steps in mast cell degranulation. J. Immunol. 2014, 192, 41–51. [Google Scholar] [CrossRef]
- Wiegerinck, C.L.; Janecke, A.R.; Schneeberger, K.; Vogel, G.F.; van Haaften-Visser, D.Y.; Escher, J.C.; Adam, R.; Thoni, C.E.; Pfaller, K.; Jordan, A.J.; et al. Loss of syntaxin 3 causes variant microvillus inclusion disease. Gastroenterology 2014, 147, 65–68.e10. [Google Scholar] [CrossRef]
- Julia, J.; Shui, V.; Mittal, N.; Heim-Hall, J.; Blanco, C.L. Microvillus inclusion disease, a diagnosis to consider when abnormal stools and neurological impairments run together due to a rare syntaxin 3 gene mutation. J. Neonatal Perinat. Med. 2019, 12, 313–319. [Google Scholar] [CrossRef]
- Gutierrez, B.A.; Chavez, M.A.; Rodarte, A.I.; Ramos, M.A.; Dominguez, A.; Petrova, Y.; Davalos, A.J.; Costa, R.M.; Elizondo, R.; Tuvim, M.J.; et al. Munc18-2, but not Munc18-1 or Munc18-3, controls compound and single-vesicle-regulated exocytosis in mast cells. J. Biol. Chem. 2018, 293, 7148–7159. [Google Scholar] [CrossRef]
- Yang, X.; Kaeser-Woo, Y.J.; Pang, Z.P.; Xu, W.; Sudhof, T.C. Complexin clamps asynchronous release by blocking a secondary Ca(2+) sensor via its accessory alpha helix. Neuron 2010, 68, 907–920. [Google Scholar] [CrossRef] [PubMed]
- Bera, M.; Ramakrishnan, S.; Coleman, J.; Krishnakumar, S.S.; Rothman, J.E. Molecular determinants of complexin clamping and activation function. eLife 2022, 11, e71938. [Google Scholar] [CrossRef] [PubMed]
- Zdanowicz, R.; Kreutzberger, A.; Liang, B.; Kiessling, V.; Tamm, L.K.; Cafiso, D.S. Complexin Binding to Membranes and Acceptor t-SNAREs Explains Its Clamping Effect on Fusion. Biophys J 2017, 113, 1235–1250. [Google Scholar] [CrossRef]
- Brose, N. For better or for worse: Complexins regulate SNARE function and vesicle fusion. Traffic 2008, 9, 1403–1413. [Google Scholar] [CrossRef]
- Hobson, R.J.; Liu, Q.; Watanabe, S.; Jorgensen, E.M. Complexin maintains vesicles in the primed state in C. elegans. Curr. Biol. 2011, 21, 106–113. [Google Scholar] [CrossRef]
- Martin, J.A.; Hu, Z.; Fenz, K.M.; Fernandez, J.; Dittman, J.S. Complexin has opposite effects on two modes of synaptic vesicle fusion. Curr. Biol. 2011, 21, 97–105. [Google Scholar] [CrossRef]
- Maximov, A.; Tang, J.; Yang, X.; Pang, Z.P.; Sudhof, T.C. Complexin controls the force transfer from SNARE complexes to membranes in fusion. Science 2009, 323, 516–521. [Google Scholar] [CrossRef]
- Xue, M.; Craig, T.K.; Xu, J.; Chao, H.T.; Rizo, J.; Rosenmund, C. Binding of the complexin N terminus to the SNARE complex potentiates synaptic-vesicle fusogenicity. Nat. Struct. Mol. Biol. 2010, 17, 568–575. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishad, R.; Betancourt-Solis, M.; Dey, H.; Heidelberger, R.; McNew, J.A. Regulation of Syntaxin3B-Mediated Membrane Fusion by T14, Munc18, and Complexin. Biomolecules 2023, 13, 1463. https://doi.org/10.3390/biom13101463
Nishad R, Betancourt-Solis M, Dey H, Heidelberger R, McNew JA. Regulation of Syntaxin3B-Mediated Membrane Fusion by T14, Munc18, and Complexin. Biomolecules. 2023; 13(10):1463. https://doi.org/10.3390/biom13101463
Chicago/Turabian StyleNishad, Rajkishor, Miguel Betancourt-Solis, Himani Dey, Ruth Heidelberger, and James A. McNew. 2023. "Regulation of Syntaxin3B-Mediated Membrane Fusion by T14, Munc18, and Complexin" Biomolecules 13, no. 10: 1463. https://doi.org/10.3390/biom13101463