
Ploidy Status of Ovarian Cancer Cell Lines and Their Association with Gene Expression Profiles
Abstract
:1. Introduction
2. Materials and Methods
2.1. Online Tools
2.2. Cell Lines
2.3. G-Banding
2.4. Flow Cytometry
2.5. RNA Sequencing
2.6. Identification of DEGs and Enrichment Analysis
2.7. Statistical Analysis
3. Results
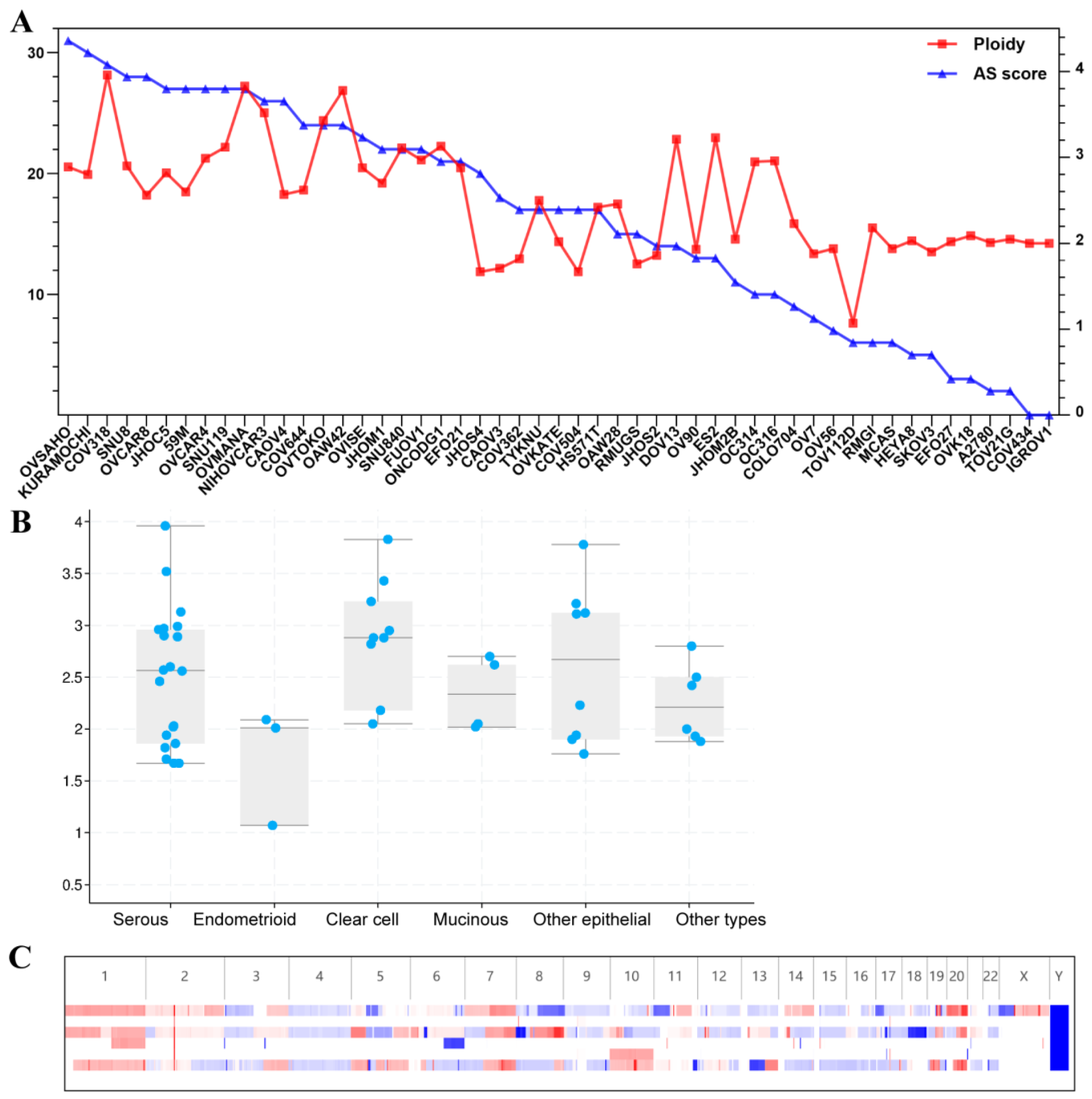
3.1. Ploidy Analysis from the CCLE for 51 Ovarian Cancer Cell Lines
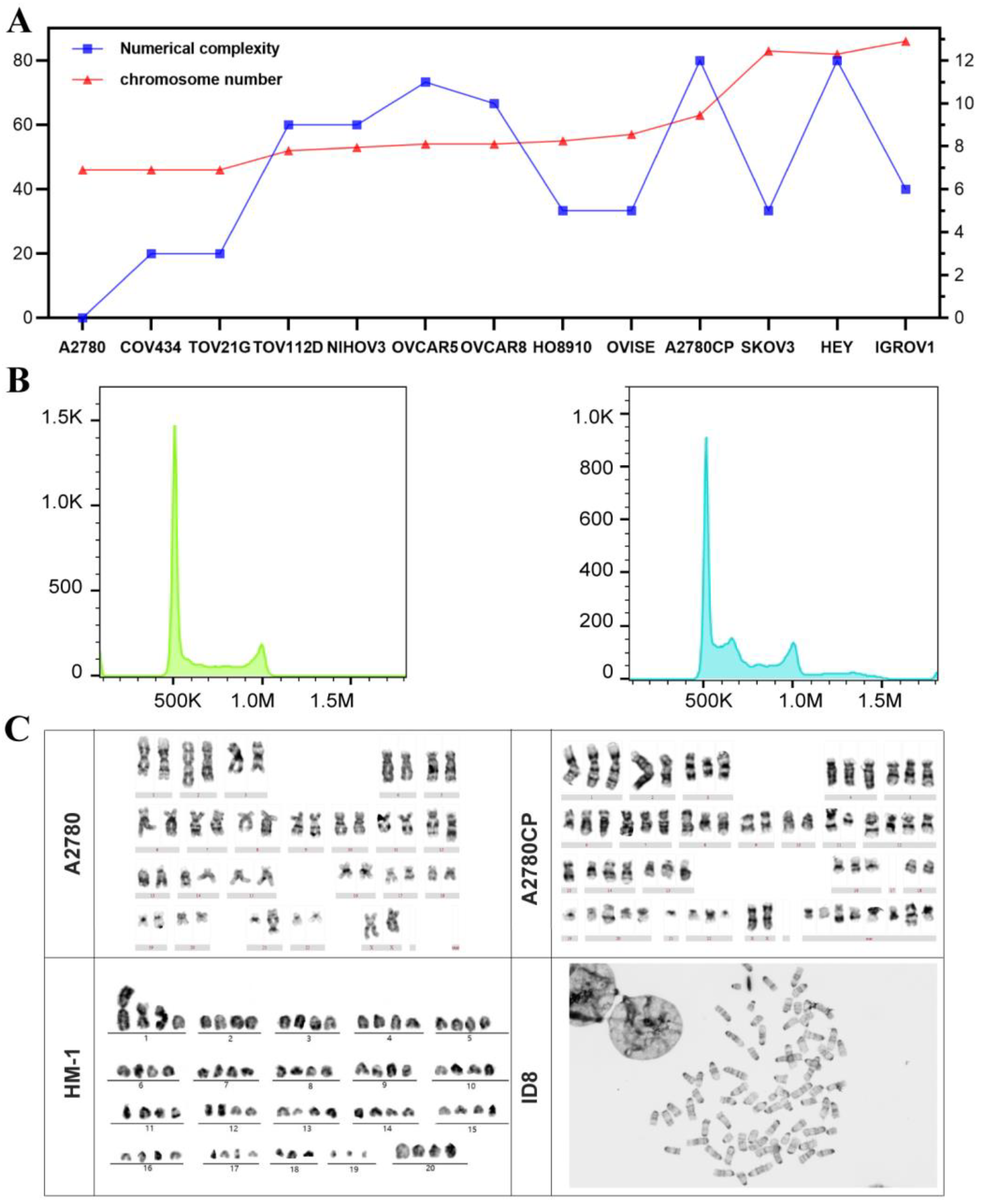
3.2. Karyotype Numerical Complexity and Integrated Ploidy Information for Ovarian Cancer Cell Lines
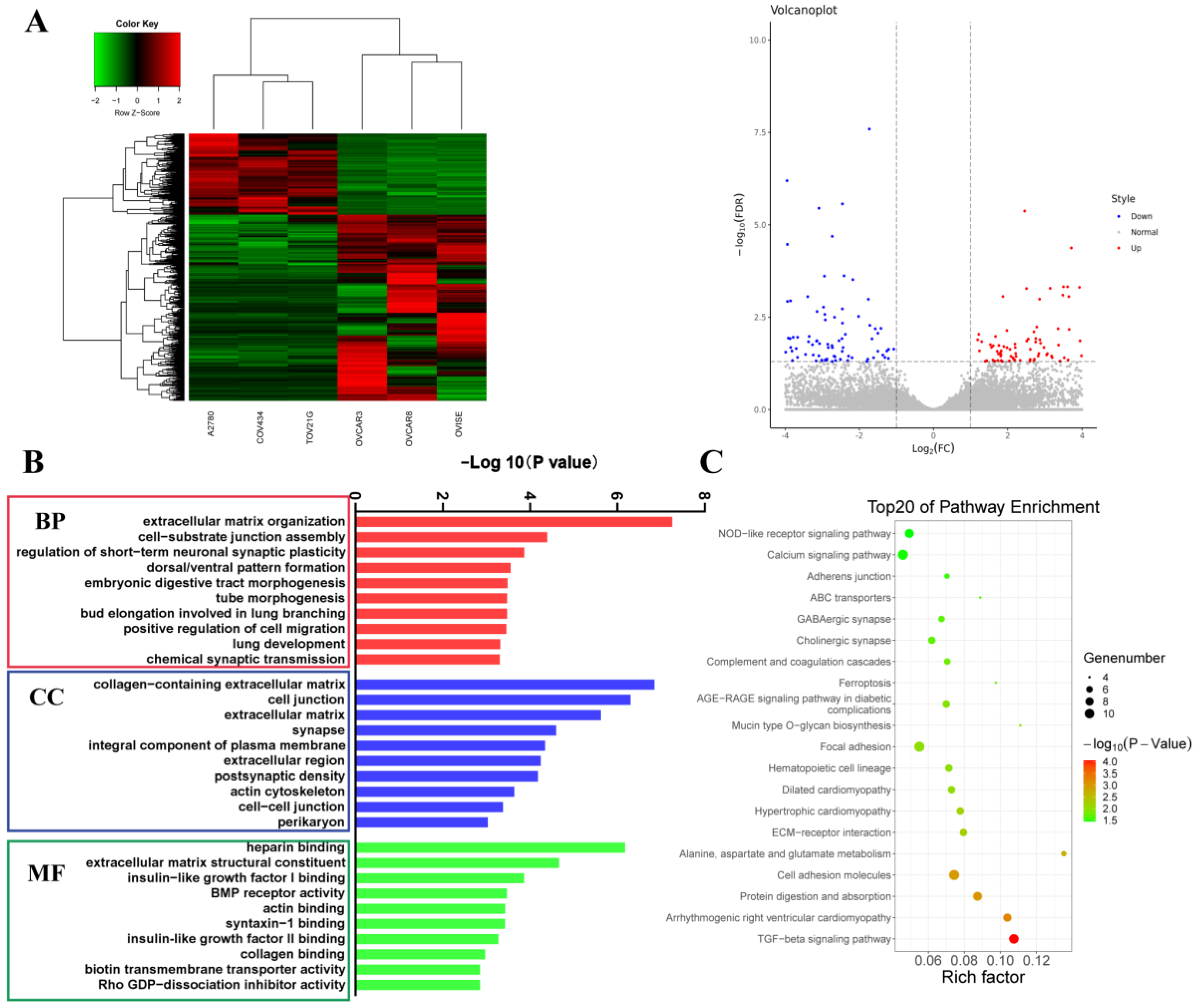
3.3. Difference in Expression Profiles between Human Diploid and Aneuploid Ovarian Cancer Cell Lines
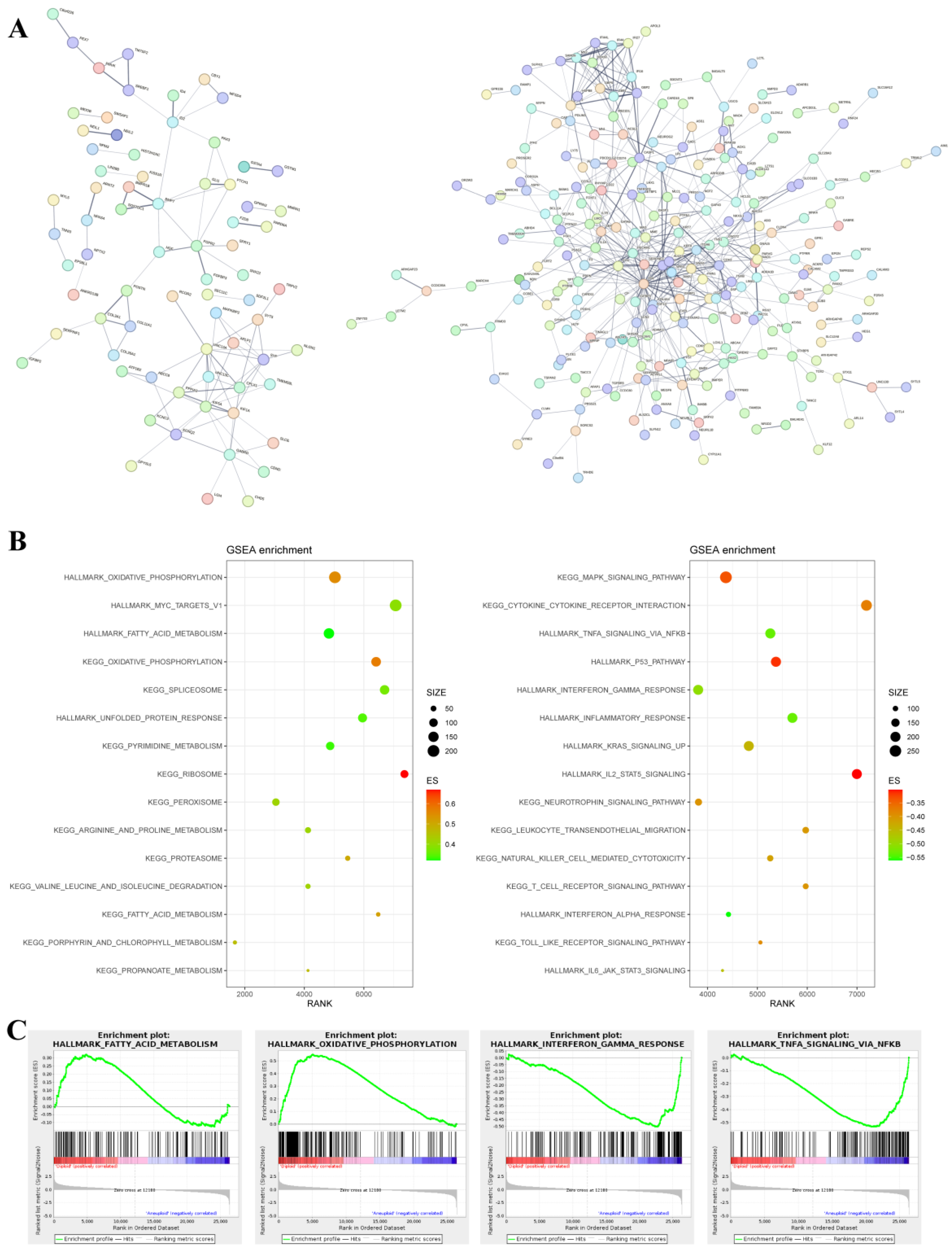
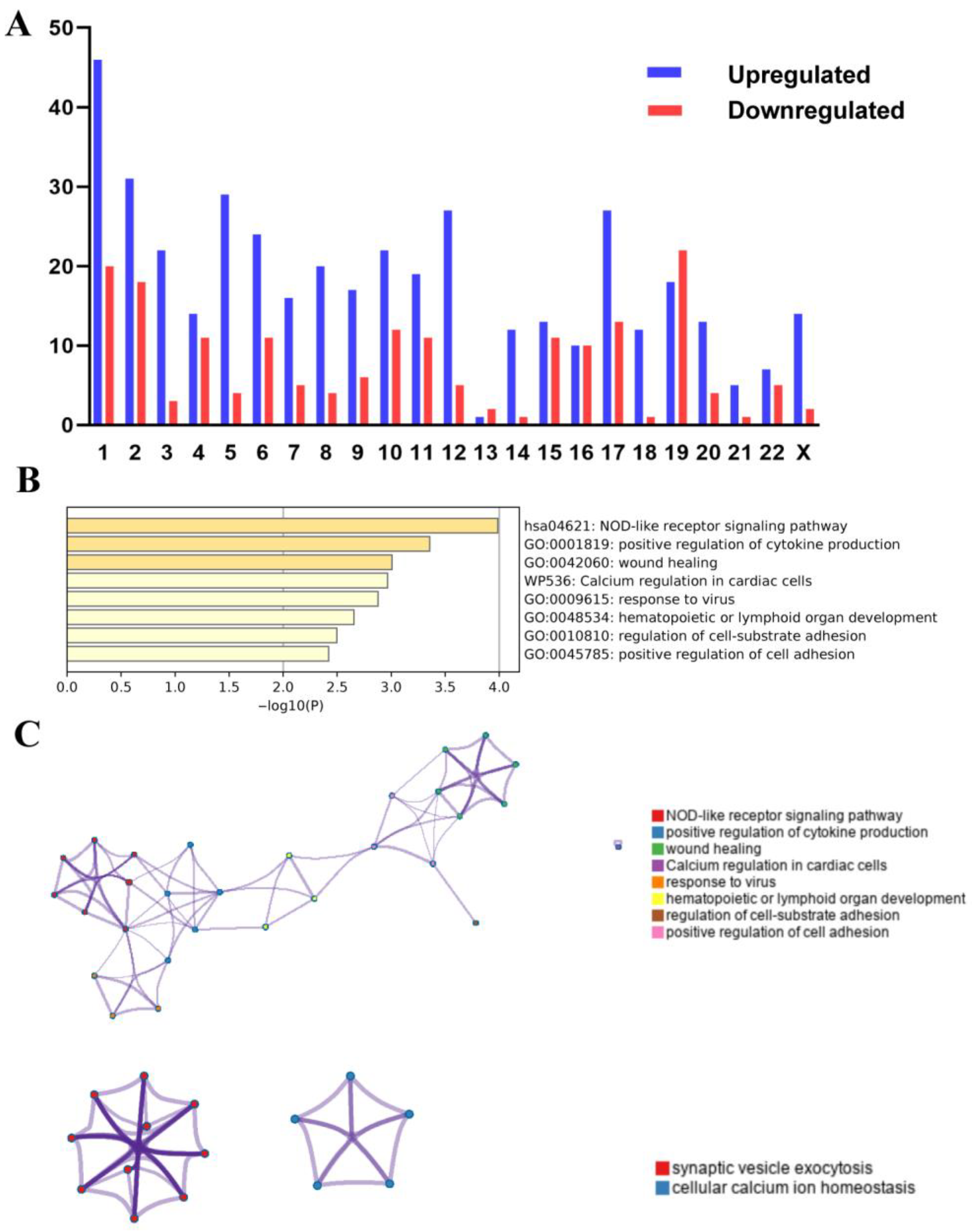
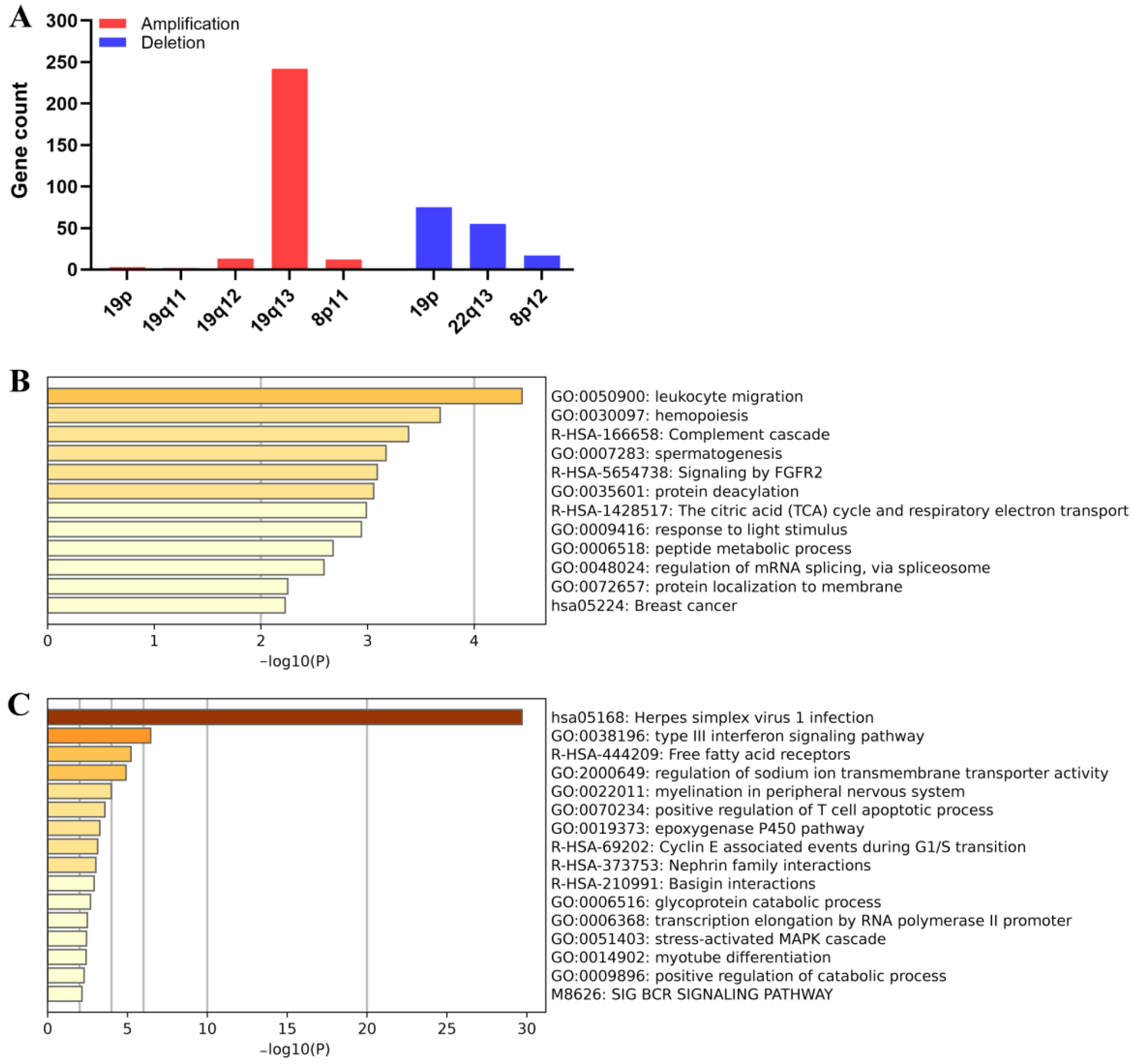
3.4. Effects of Chromosome Alterations on Gene Expression Regulation
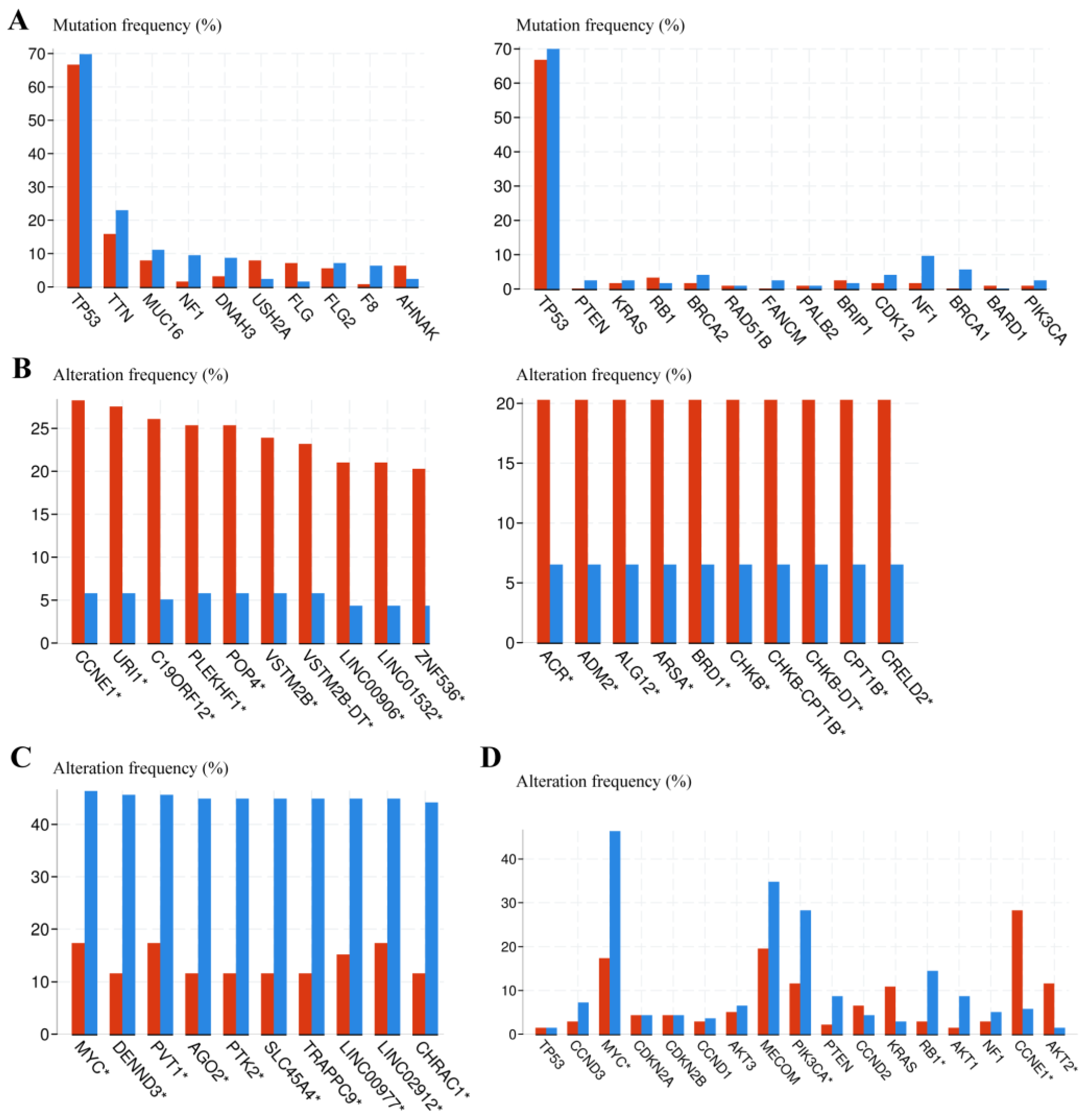
3.5. Genomic Alteration Analysis between Euploid and Aneuploid Ovarian Cancer Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Lheureux, S.; Msc, M.B.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baert, T.; Ferrero, A.; Sehouli, J.; O’Donnell, D.; González-Martín, A.; Joly, F.; van der Velden, J.; Blecharz, P.; Tan, D.; Querleu, D.; et al. The systemic treatment of recurrent ovarian cancer revisited. Ann. Oncol. 2021, 32, 710–725. [Google Scholar] [CrossRef] [PubMed]
- Corrado, G.; Salutari, V.; Palluzzi, E.; Distefano, M.G.; Scambia, G.; Ferrandina, G. Optimizing treatment in recurrent epithelial ovarian cancer. Expert Rev. Anticancer Ther. 2017, 17, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.A.; Cleveland, D.W. Does aneuploidy cause cancer? Curr. Opin. Cell Biol. 2006, 18, 658–667. [Google Scholar] [CrossRef]
- Watkins, T.B.K.; Lim, E.L.; Petkovic, M.; Elizalde, S.; Birkbak, N.J.; Wilson, G.A.; Moore, D.A.; Grönroos, E.; Rowan, A.; Dewhurst, S.M.; et al. Pervasive chromosomal instability and karyotype order in tumour evolution. Nature 2020, 587, 126–132. [Google Scholar] [CrossRef]
- Lukow, D.A.; Sausville, E.L.; Suri, P.; Chunduri, N.K.; Wieland, A.; Leu, J.; Smith, J.C.; Girish, V.; Kumar, A.A.; Kendall, J.; et al. Chromosomal instability accelerates the evolution of resistance to anti-cancer therapies. Dev. Cell 2021, 56, 2427–2439.e4. [Google Scholar] [CrossRef]
- Laughney, A.; Elizalde, S.; Genovese, G.; Bakhoum, S.F. Dynamics of Tumor Heterogeneity Derived from Clonal Karyotypic Evolution. Cell Rep. 2015, 12, 809–820. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, M.R.; Martis, V.; Martin, S.; Tijhuis, A.E.; Hong, C.; Wardenaar, R.; Dumont, M.; Zerbib, J.; Spierings, D.C.; Fachinetti, D.; et al. Gene copy-number changes and chromosomal instability induced by aneuploidy confer resistance to chemotherapy. Dev. Cell 2021, 56, 2440–2454.e6. [Google Scholar] [CrossRef]
- Tripathi, R.; Modur, V.; Senovilla, L.; Kroemer, G.; Komurov, K. Suppression of tumor antigen presentation during aneuploid tumor evolution contributes to immune evasion. OncoImmunology 2019, 8, 1657374. [Google Scholar] [CrossRef]
- Ming, D.; Zhang, S.; Liu, X.; Xu, C.; Zhang, X. Nondiploid cancer cells: Stress, tolerance and therapeutic inspirations. Biochim. Biophys Acta Rev. Cancer 2022, 1877, 188794. [Google Scholar] [CrossRef]
- Kildal, W.; Risberg, B.; Abeler, V.M.; Kristensen, G.B.; Sudbø, J.; Nesland, J.M.; Danielsen, H.E. β-catenin expression, DNA ploidy and clinicopathological features in ovarian cancer: A study in 253 patients. Eur. J. Cancer 2005, 41, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Gallerani, G.; Tesei, A.; Scarpi, E.; Verdecchia, G.M.; Virzì, S.; Fabbri, F.; Arienti, C. DNA ploidy and S-phase fraction analysis in peritoneal carcinomatosis from ovarian cancer: Correlation with clinical pathological factors and response to chemotherapy. OncoTargets Ther. 2017, 10, 4657–4664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tropé, C.; Kaern, J.; Hogberg, T.; Abeler, V.; Hagen, B.; Kristensen, G.; Onsrud, M.; Pettersen, E.; Rosenberg, P.; Sandvei, R.; et al. Randomized study on adjuvant chemotherapy in stage I high-risk ovarian cancer with evaluation of DNA-ploidy as prognostic instrument. Ann. Oncol. 2000, 11, 281–288. [Google Scholar] [CrossRef]
- Penner-Goeke, S.; Lichtensztejn, Z.; Neufeld, M.; Ali, J.L.; Altman, A.D.; Nachtigal, M.W.; McManus, K.J. The temporal dynamics of chromosome instability in ovarian cancer cell lines and primary patient samples. PLoS Genet. 2017, 13, e1006707. [Google Scholar] [CrossRef] [Green Version]
- Morden, C.R.; Farrell, A.C.; Sliwowski, M.; Lichtensztejn, Z.; Altman, A.D.; Nachtigal, M.W.; McManus, K.J. Chromosome instability is prevalent and dynamic in high-grade serous ovarian cancer patient samples. Gynecol. Oncol. 2021, 161, 769–778. [Google Scholar] [CrossRef]
- Quinton, R.J.; DiDomizio, A.; Vittoria, M.A.; Kotýnková, K.; Ticas, C.J.; Patel, S.; Koga, Y.; Vakhshoorzadeh, J.; Hermance, N.; Kuroda, T.S.; et al. Whole-genome doubling confers unique genetic vulnerabilities on tumour cells. Nature 2021, 590, 492–497. [Google Scholar] [CrossRef]
- Zack, T.I.; Schumacher, S.E.; Carter, S.L.; Cherniack, A.D.; Saksena, G.; Tabak, B.; Lawrence, M.S.; Zhang, C.Z.; Wala, J.; Mermel, C.H.; et al. Pan-cancer patterns of somatic copy number alteration. Nat. Genet. 2013, 45, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Carter, S.L.; Cibulskis, K.; Helman, E.; McKenna, A.; Shen, H.; Zack, T.; Laird, P.W.; Onofrio, R.C.; Winckler, W.; Weir, B.A.; et al. Absolute quantification of somatic DNA alterations in human cancer. Nat. Biotechnol. 2012, 30, 413–421. [Google Scholar] [CrossRef]
- Cheng, Z.; Mirza, H.; Ennis, D.P.; Smith, P.; Gavarró, L.M.; Sokota, C.; Giannone, G.; Goranova, T.; Bradley, T.; Piskorz, A.; et al. The Genomic Landscape of Early-Stage Ovarian High-Grade Serous Carcinoma. Clin. Cancer Res. 2022, 28, 2911–2922. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, J.W.; Kim, Y.T.; Kim, S.; Yoon, B.S.; Nam, E.J.; Kim, H.Y. Analysis of chromosomal changes in serous ovarian carcinoma using high-resolution array comparative genomic hybridization: Potential predictive markers of chemoresistant disease. Genes Chromosom. Cancer 2006, 46, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Etemadmoghadam, D.; Defazio, A.; Beroukhim, R.; Mermel, C.; George, J.; Getz, G.; Tothill, R.; Okamoto, A.; Raeder, M.B.; Harnett, P.; et al. Integrated Genome-Wide DNA Copy Number and Expression Analysis Identifies Distinct Mechanisms of Primary Chemoresistance in Ovarian Carcinomas. Clin. Cancer Res. 2009, 15, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malek, J.A.; Méry, E.; Mahmoud, Y.A.; Al-Azwani, E.K.; Roger, L.; Huang, R.Y.-J.; Jouve, E.; Lis, R.; Thiery, J.-P.; Querleu, D.; et al. Copy Number Variation Analysis of Matched Ovarian Primary Tumors and Peritoneal Metastasis. PLoS ONE 2011, 6, e28561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paracchini, L.; Beltrame, L.; Grassi, T.; Inglesi, A.; Fruscio, R.; Landoni, F.; Ippolito, D.; Marchette, M.D.; Paderno, M.; Adorni, M.; et al. Genome-wide Copy-number Alterations in Circulating Tumor DNA as a Novel Biomarker for Patients with High-grade Serous Ovarian Cancer. Clin. Cancer Res. 2020, 27, 2549–2559. [Google Scholar] [CrossRef]
- Braicu, E.I.; du Bois, A.; Sehouli, J.; Beck, J.; Prader, S.; Kulbe, H.; Eiben, B.; Harter, P.; Traut, A.; Pietzner, K.; et al. Cell-Free-DNA-Based Copy Number Index Score in Epithelial Ovarian Cancer—Impact for Diagnosis and Treatment Monitoring. Cancers 2021, 14, 168. [Google Scholar] [CrossRef]
- Boudhraa, Z.; Zaoui, K.; Fleury, H.; Cahuzac, M.; Gilbert, S.; Tchakarska, G.; Kendall-Dupont, J.; Carmona, E.; Provencher, D.; Mes-Masson, A.-M. NR1D1 regulation by Ran GTPase via miR4472 identifies an essential vulnerability linked to aneuploidy in ovarian cancer. Oncogene 2021, 41, 309–320. [Google Scholar] [CrossRef]
- Chui, M.H.; Doodnauth, S.A.; Erdmann, N.; Tiedemann, R.E.; Sircoulomb, F.; Drapkin, R.; Shaw, P.; Rottapel, R. Chromosomal Instability and mTORC1 Activation through PTEN Loss Contribute to Proteotoxic Stress in Ovarian Carcinoma. Cancer Res. 2019, 79, 5536–5549. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.-C.; Williams, B.R.; Siegel, J.J.; Amon, A. Identification of Aneuploidy-Selective Antiproliferation Compounds. Cell 2011, 144, 499–512. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Jilderda, L.J.; Foijer, F. Exploiting aneuploidy-imposed stresses and coping mechanisms to battle cancer. Open Biol. 2020, 10, 200148. [Google Scholar] [CrossRef]
- Oromendia, A.B.; Dodgson, S.E.; Amon, A. Aneuploidy causes proteotoxic stress in yeast. Genes Dev. 2012, 26, 2696–2708. [Google Scholar] [CrossRef]
- Micci, F.; Skotheim, R.I.; Haugom, L.; Weimer, J.; Eibak, A.M.E.; Abeler, V.M.; Trope, C.G.; Arnold, N.; Lothe, R.A.; Heim, S. Array-CGH analysis of microdissected chromosome 19 markers in ovarian carcinoma identifies candidate target genes. Genes, Chromosom. Cancer 2010, 49, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Amfo, K.; Neyns, B.; Teugels, E.; Lissens, W.; Bourgain, C.; De Sutter, P.; Vandamme, B.; Vamos, E.; De Greve, J. Frequent deletion of chromosome 19 and a rare rearrangement of 19p13.3 involving the insulin receptor gene in human ovarian cancer. Oncogene 1995, 11, 351–358. [Google Scholar] [PubMed]
- van der Riet-Fox, M.F.; Retief, A.E.; van Niekerk, W.A. Chromosome changes in 17 human neoplasms studied with banding. Cancer 1979, 44, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Gallion, H.; Powell, D.; Smith, L.; Morrow, J.; Martin, A.; van Nagell, J.; Donaldson, E. Chromosome abnormalities in human epithelial ovarian malignancies. Gynecol. Oncol. 1990, 38, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689.e3. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Sharir, Y.; McFarland, J.M.; Abdusamad, M.; Marquis, C.; Bernhard, S.V.; Kazachkova, M.; Tang, H.; Ippolito, M.R.; Laue, K.; Zerbib, J.; et al. Aneuploidy renders cancer cells vulnerable to mitotic checkpoint inhibition. Nature 2021, 590, 486–491. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Ghandi, M.; Huang, F.W.; Jané-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., III; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Peng, J.; Xu, H.; Cai, J. Establishment and characterization of a new gastric cancer cell line, XGC-1. Cancer Cell Int. 2020, 20, 437. [Google Scholar] [CrossRef]
- Oh, S.; Min, K.; Kim, M.; Lee, S.K. Sex Chromosomes Are Severely Disrupted in Gastric Cancer Cell Lines. Int. J. Mol. Sci. 2020, 21, 4598. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav Brain Res 2001, 125, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschke, A.V.; Tonon, G.; Gehlhaus, K.S.; Mctyre, N.; Bussey, K.J.; Lababidi, S.; Scudiero, D.A.; Weinstein, J.N.; Kirsch, I.R. Karyotypic complexity of the NCI-60 drug-screening panel. Cancer Res. 2003, 63, 8634–8647. [Google Scholar]
- Castedo, M.; Galluzzi, L.; Vitale, I.; Senovilla, L.; Métivier, D.; Jèmaà, M.; Rello-Varona, S.; Kroemer, G. Cytofluorometric Purification of Diploid and Tetraploid Cancer Cells. 2011, 761, 47–63. In Cell Cycle Synchronization; Humana Press: Totowa, NJ, USA, 2011; Volume 761, pp. 47–63. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.L.; Compton, D.A. Examining the link between chromosomal instability and aneuploidy in human cells. J. Cell Biol. 2008, 180, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Niu, N.; Mercado-Uribe, I.; Liu, J. Dedifferentiation into blastomere-like cancer stem cells via formation of polyploid giant cancer cells. Oncogene 2017, 36, 4887–4900. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Tsai, H.-J.; Gordon, M.R.; Li, R. Cellular Stress Associated with Aneuploidy. Dev. Cell 2018, 44, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Vitale, I.; Manic, G.; Senovilla, L.; Kroemer, G.; Galluzzi, L. Karyotypic Aberrations in Oncogenesis and Cancer Therapy. Trends Cancer 2015, 1, 124–135. [Google Scholar] [CrossRef]
- Auslander, N.; Wolf, Y.I.; Koonin, E.V. Interplay between DNA damage repair and apoptosis shapes cancer evolution through aneuploidy and microsatellite instability. Nat. Commun. 2020, 11, 1234. [Google Scholar] [CrossRef]
- Choudhary, A.; Zachek, B.; Lera, R.F.; Zasadil, L.M.; Lasek, A.; Denu, R.A.; Kim, H.; Kanugh, C.; Laffin, J.J.; Harter, J.M.; et al. Identification of Selective Lead Compounds for Treatment of High-Ploidy Breast Cancer. Mol. Cancer Ther. 2016, 15, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Replogle, J.M.; Zhou, W.; Amaro, A.E.; McFarland, J.M.; Villalobos-Ortiz, M.; Ryan, J.; Letai, A.; Yilmaz, O.; Sheltzer, J.; Lippard, S.J.; et al. Aneuploidy increases resistance to chemotherapeutics by antagonizing cell division. Proc. Natl. Acad. Sci. USA 2020, 117, 30566–30576. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.Y.; Wang, D.D.; Li, L.; Lin, P.P. Identification and Comprehensive Co-Detection of Necrotic and Viable Aneuploid Cancer Cells in Peripheral Blood. Cancers 2021, 13, 5108. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Qi, C.; Zhang, C.; Liu, D.; Deng, Y.; Fu, Y.; Khadka, V.S.; Wang, D.D.; Tan, S.; et al. Dysregulated KRAS gene-signaling axis and abnormal chromatin remodeling drive therapeutic resistance in heterogeneous-sized circulating tumor cells in gastric cancer patients. Cancer Lett. 2021, 517, 78–87. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Liu, D.; Gong, J.; Wang, D.D.; Li, S.; Peng, Z.; Li, Y.; Wang, X.; Lin, P.P.; et al. Evolutionary Expression of HER2 Conferred by Chromosome Aneuploidy on Circulating Gastric Cancer Cells Contributes to Developing Targeted and Chemotherapeutic Resistance. Clin. Cancer Res. 2018, 24, 5261–5271. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Paek, A.L.; Hammer, M.; Rangarajan, S.; Ruijtenbeek, R.; Cooke, L.; Weterings, E.; Mahadevan, D. Drug-induced aneuploidy and polyploidy is a mechanism of disease relapse in MYC/BCL2-addicted diffuse large B-cell lymphoma. Oncotarget 2018, 9, 35875–35890. [Google Scholar] [CrossRef] [Green Version]
- Davoli, T.; Uno, H.; Wooten, E.C.; Elledge, S.J. Tumor aneuploidy correlates with markers of immune evasion and with reduced response to immunotherapy. Science 2017, 355, eaaf8399. [Google Scholar] [CrossRef] [Green Version]
- Boilève, A.; Senovilla, L.; Vitale, I.; Lissa, D.; Martins, I.; Métivier, D.; Brink, S.V.D.; Clevers, H.; Galluzzi, L.; Castedo, M.; et al. Immunosurveillance against tetraploidization-induced colon tumorigenesis. Cell Cycle 2013, 12, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.W.; Viganò, S.; Ben-David, U.; Amon, A.; Santaguida, S. Aneuploid senescent cells activate NF-κB to promote their immune clearance by NK cells. EMBO Rep. 2021, 22, e52032. [Google Scholar] [CrossRef]
- Bakker, B.; Van Den Bos, H.; Lansdorp, P.M.; Foijer, F. How to count chromosomes in a cell: An overview of current and novel technologies. BioEssays 2015, 37, 570–577. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Type | Name | Karyotype Analysis | ||
|---|---|---|---|---|
| Modal Number | Ploidy | Numerical Range 1 | ||
| Adenocarcinoma | ||||
| Serous | NIHOVCAR3 | 53 | hyperdiploid | 49–57 |
| OVCAR5 | 54 | hyperdiploid | 47–57 | |
| OVCAR8 | 54 | hyperdiploid | 50–59 | |
| Endometrial | TOV112D | 52 | hyperdiploid | 47–55 |
| A2780 | 46 | diploid | 46 | |
| A2780CP | 63 | hypotriploid | 56–67 | |
| Clear cell | OVISE | 57 | hypotriploid | 54–58 |
| TOV21G | 46 | diploid | 45–47 | |
| Mixed | IGROV1 | 86 | hypotetraploid | 84–89 |
| NS 2 | HO8910 | 55 | hyperdiploid | 51–55 3 |
| HEY | 82 | hypotetraploid | 72–83 | |
| Cystadenocarcinoma | SKOV3 | 83 | hypotetraploid | 79–83 |
| Granulosa cell tumor | COV434 | 46 | diploid | 44–46 |
| Name | Ploidy Status | |||
|---|---|---|---|---|
| Modal Number | Ploidy Estimation | Aneuploid Score | Ploidy Value | |
| IGROV1 | 86 | hypotetraploid | 0 | 2.00 |
| SKOV3 | 81 | hypotetraploid | 5 | 1.90 |
| OVISE | 57 | hypotriploid | 23 | 2.88 |
| OVCAR8 | 54 | hyperdiploid | 28 | 2.56 |
| NIHOVCAR3 | 53 | hyperdiploid | 26 | 3.52 |
| TOV112D | 52 | hyperdiploid | 6 | 1.07 |
| A2780 | 46 | diploid | 2 | 2.01 |
| TOV21G | 46 | diploid | 2 | 2.05 |
| COV434 | 46 | diploid | 0 | 2.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, M.; Zhang, S.; Liu, X.; Xu, C.; Zhang, X. Ploidy Status of Ovarian Cancer Cell Lines and Their Association with Gene Expression Profiles. Biomolecules 2023, 13, 92. https://doi.org/10.3390/biom13010092
Du M, Zhang S, Liu X, Xu C, Zhang X. Ploidy Status of Ovarian Cancer Cell Lines and Their Association with Gene Expression Profiles. Biomolecules. 2023; 13(1):92. https://doi.org/10.3390/biom13010092
Chicago/Turabian StyleDu, Ming, Shuo Zhang, Xiaoxia Liu, Congjian Xu, and Xiaoyan Zhang. 2023. "Ploidy Status of Ovarian Cancer Cell Lines and Their Association with Gene Expression Profiles" Biomolecules 13, no. 1: 92. https://doi.org/10.3390/biom13010092