
The Dark Side of Sphingolipids: Searching for Potential Cardiovascular Biomarkers

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
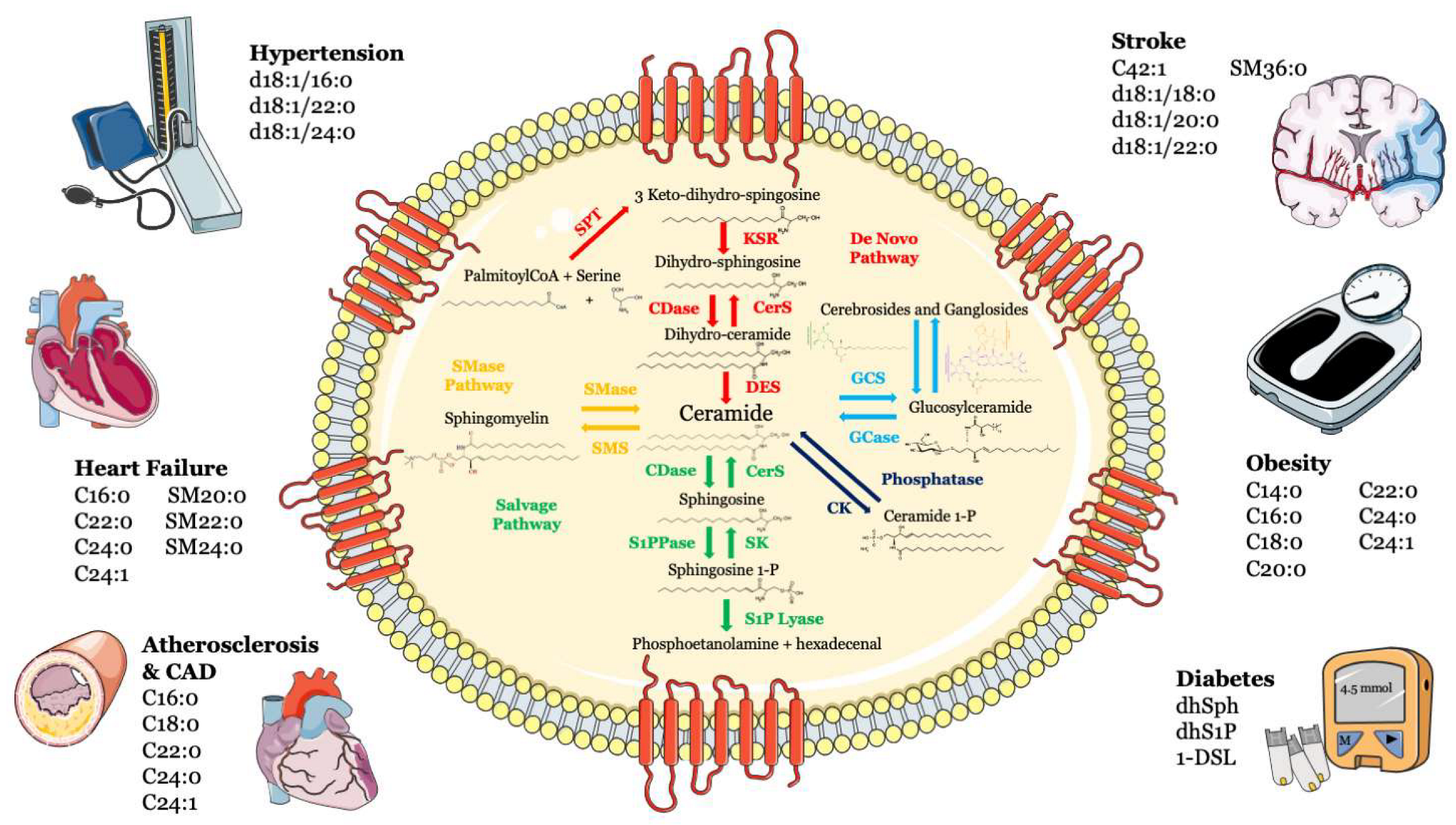
2. Atherosclerosis and Coronary Artery Disease
3. Heart Failure
4. Hypertension
5. Stroke
6. Vascular Dysfunction
7. Diabetes
8. Obesity
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenfeldt, H.M.; Hobson, J.P.; Maceyka, M.; Olivera, A.; Nava, V.E.; Milstien, S.; Spiegel, S. EDG-1 links the PDGF receptor to Src and focal adhesion kinase activation leading to lamellipodia formation and cell migration. FASEB J. 2001, 15, 2649–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; He, Q.; Sharma, R.P. Sphingosine kinase activity confers resistance to apoptosis by fumonisin B1 in human embryonic kidney (HEK-293) cells. Chem. Biol. Interact. 2004, 151, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Venable, M.E.; Webb-Froehlich, L.M.; Sloan, E.F.; Thomley, J.E. Shift in sphingolipid metabolism leads to an accumulation of ceramide in senescence. Mech. Ageing Dev. 2006, 127, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.A.; Zhang, Y.; Li, X.; Gulbins, E. Ceramide: A novel player in reactive oxygen species-induced signaling? Antioxid Redox Signal 2007, 9, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Becker, K.A.; Zhang, Y. Ceramide in redox signaling and cardiovascular diseases. Cell Physiol. Biochem. 2010, 26, 41–48. [Google Scholar] [CrossRef]
- Levade, T.; Auge, N.; Veldman, R.J.; Cuvillier, O.; Negre-Salvayre, A.; Salvayre, R. Sphingolipid mediators in cardiovascular cell biology and pathology. Circ. Res. 2001, 89, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Spijkers, L.J.; van den Akker, R.F.; Janssen, B.J.; Debets, J.J.; De Mey, J.G.; Stroes, E.S.; van den Born, B.J.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension is associated with marked alterations in sphingolipid biology: A potential role for ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef] [Green Version]
- Laaksonen, R.; Ekroos, K.; Sysi-Aho, M.; Hilvo, M.; Vihervaara, T.; Kauhanen, D.; Suoniemi, M.; Hurme, R.; Marz, W.; Scharnagl, H.; et al. Plasma ceramides predict cardiovascular death in patients with stable coronary artery disease and acute coronary syndromes beyond LDL-cholesterol. Eur. Heart J. 2016, 37, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and Lipoproteins in Health and Metabolic Disorders. Trends Endocrinol. Metab. 2017, 28, 506–518. [Google Scholar] [CrossRef]
- Holland, W.L.; Brozinick, J.T.; Wang, L.P.; Hawkins, E.D.; Sargent, K.M.; Liu, Y.; Narra, K.; Hoehn, K.L.; Knotts, T.A.; Siesky, A.; et al. Inhibition of ceramide synthesis ameliorates glucocorticoid-, saturated-fat-, and obesity-induced insulin resistance. Cell Metab. 2007, 5, 167–179. [Google Scholar] [CrossRef]
- Hojjati, M.R.; Li, Z.; Zhou, H.; Tang, S.; Huan, C.; Ooi, E.; Lu, S.; Jiang, X.C. Effect of myriocin on plasma sphingolipid metabolism and atherosclerosis in apoE-deficient mice. J. Biol. Chem. 2005, 280, 10284–10289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, R.; Akashi, H.; Drosatos, K.; Liao, X.; Jiang, H.; Kennel, P.J.; Brunjes, D.L.; Castillero, E.; Zhang, X.; Deng, L.Y.; et al. Increased de novo ceramide synthesis and accumulation in failing myocardium. JCI Insight 2017, 2, e82922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.J.; Holland, W.L.; Wilson, L.; Tanner, J.M.; Kearns, D.; Cahoon, J.M.; Pettey, D.; Losee, J.; Duncan, B.; Gale, D.; et al. Ceramide mediates vascular dysfunction in diet-induced obesity by PP2A-mediated dephosphorylation of the eNOS-Akt complex. Diabetes 2012, 61, 1848–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharath, L.P.; Ruan, T.; Li, Y.; Ravindran, A.; Wan, X.; Nhan, J.K.; Walker, M.L.; Deeter, L.; Goodrich, R.; Johnson, E.; et al. Ceramide-Initiated Protein Phosphatase 2A Activation Contributes to Arterial Dysfunction In Vivo. Diabetes 2015, 64, 3914–3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, R.N.; Yu, C.; Hoofnagle, A.; Hari, N.; Jensen, P.N.; Fretts, A.M.; Umans, J.G.; Howard, B.V.; Sitlani, C.M.; Siscovick, D.S.; et al. Circulating Sphingolipids, Insulin, HOMA-IR, and HOMA-B: The Strong Heart Family Study. Diabetes 2018, 67, 1663–1672. [Google Scholar] [CrossRef] [Green Version]
- Anroedh, S.; Hilvo, M.; Akkerhuis, K.M.; Kauhanen, D.; Koistinen, K.; Oemrawsingh, R.; Serruys, P.; van Geuns, R.J.; Boersma, E.; Laaksonen, R.; et al. Plasma concentrations of molecular lipid species predict long-term clinical outcome in coronary artery disease patients. J. Lipid Res. 2018, 59, 1729–1737. [Google Scholar] [CrossRef]
- Hilvo, M.; Vasile, V.C.; Donato, L.J.; Hurme, R.; Laaksonen, R. Ceramides and Ceramide Scores: Clinical Applications for Cardiometabolic Risk Stratification. Front. Endocrinol. 2020, 11, 570628. [Google Scholar] [CrossRef]
- Sattler, K.J.; Elbasan, S.; Keul, P.; Elter-Schulz, M.; Bode, C.; Graler, M.H.; Brocker-Preuss, M.; Budde, T.; Erbel, R.; Heusch, G.; et al. Sphingosine 1-phosphate levels in plasma and HDL are altered in coronary artery disease. Basic Res. Cardiol. 2010, 105, 821–832. [Google Scholar] [CrossRef]
- Symons, J.D.; Abel, E.D. Lipotoxicity contributes to endothelial dysfunction: A focus on the contribution from ceramide. Rev Endocr. Metab. Disord. 2013, 14, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Holland, W.L.; Summers, S.A. Sphingolipids, insulin resistance, and metabolic disease: New insights from in vivo manipulation of sphingolipid metabolism. Endocr. Rev. 2008, 29, 381–402. [Google Scholar] [CrossRef]
- Falk, E. Pathogenesis of atherosclerosis. J. Am. Coll Cardiol. 2006, 47, C7–C12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plutzky, J. Inflammatory pathways in atherosclerosis and acute coronary syndromes. Am. J. Cardiol. 2001, 88, 10K–15K. [Google Scholar] [CrossRef] [PubMed]
- Auge, N.; Negre-Salvayre, A.; Salvayre, R.; Levade, T. Sphingomyelin metabolites in vascular cell signaling and atherogenesis. Prog. Lipid Res. 2000, 39, 207–229. [Google Scholar] [CrossRef]
- Laulederkind, S.J.; Bielawska, A.; Raghow, R.; Hannun, Y.A.; Ballou, L.R. Ceramide induces interleukin 6 gene expression in human fibroblasts. J. Exp. Med. 1995, 182, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Blake, G.J.; Ridker, P.M. Novel clinical markers of vascular wall inflammation. Circ. Res. 2001, 89, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Auge, N.; Escargueil-Blanc, I.; Lajoie-Mazenc, I.; Suc, I.; Andrieu-Abadie, N.; Pieraggi, M.T.; Chatelut, M.; Thiers, J.C.; Jaffrezou, J.P.; Laurent, G.; et al. Potential role for ceramide in mitogen-activated protein kinase activation and proliferation of vascular smooth muscle cells induced by oxidized low density lipoprotein. J. Biol. Chem. 1998, 273, 12893–12900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornfeldt, K.E.; Graves, L.M.; Raines, E.W.; Igarashi, Y.; Wayman, G.; Yamamura, S.; Yatomi, Y.; Sidhu, J.S.; Krebs, E.G.; Hakomori, S.; et al. Sphingosine-1-phosphate inhibits PDGF-induced chemotaxis of human arterial smooth muscle cells: Spatial and temporal modulation of PDGF chemotactic signal transduction. J. Cell Biol. 1995, 130, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Pyne, S.; Chapman, J.; Steele, L.; Pyne, N.J. Sphingomyelin-derived lipids differentially regulate the extracellular signal-regulated kinase 2 (ERK-2) and c-Jun N-terminal kinase (JNK) signal cascades in airway smooth muscle. Eur. J. Biochem. 1996, 237, 819–826. [Google Scholar] [CrossRef]
- Li, W.; Yang, X.; Xing, S.; Bian, F.; Yao, W.; Bai, X.; Zheng, T.; Wu, G.; Jin, S. Endogenous ceramide contributes to the transcytosis of oxLDL across endothelial cells and promotes its subendothelial retention in vascular wall. Oxid. Med. Cell Longev. 2014, 2014, 823071. [Google Scholar] [CrossRef]
- Lallemand, T.; Rouahi, M.; Swiader, A.; Grazide, M.H.; Geoffre, N.; Alayrac, P.; Recazens, E.; Coste, A.; Salvayre, R.; Negre-Salvayre, A.; et al. nSMase2 (Type 2-Neutral Sphingomyelinase) Deficiency or Inhibition by GW4869 Reduces Inflammation and Atherosclerosis in Apoe(-/-) Mice. Arter. Thromb. Vasc. Biol. 2018, 38, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatomi, Y.; Ohmori, T.; Rile, G.; Kazama, F.; Okamoto, H.; Sano, T.; Satoh, K.; Kume, S.; Tigyi, G.; Igarashi, Y.; et al. Sphingosine 1-phosphate as a major bioactive lysophospholipid that is released from platelets and interacts with endothelial cells. Blood 2000, 96, 3431–3438. [Google Scholar] [CrossRef] [PubMed]
- Yatomi, Y.; Ruan, F.; Hakomori, S.; Igarashi, Y. Sphingosine-1-phosphate: A platelet-activating sphingolipid released from agonist-stimulated human platelets. Blood 1995, 86, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Soeda, S.; Honda, O.; Shimeno, H.; Nagamatsu, A. Sphingomyelinase and cell-permeable ceramide analogs increase the release of plasminogen activator inhibitor-1 from cultured endothelial cells. Thromb. Res. 1995, 80, 509–518. [Google Scholar] [CrossRef]
- Soeda, S.; Tsunoda, T.; Kurokawa, Y.; Shimeno, H. Tumor necrosis factor-alpha-induced release of plasminogen activator inhibitor-1 from human umbilical vein endothelial cells: Involvement of intracellular ceramide signaling event. Biochim. Biophys. Acta 1998, 1448, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Keul, P.; Peters, S.; von Wnuck Lipinski, K.; Schroder, N.H.; Nowak, M.K.; Duse, D.A.; Polzin, A.; Weske, S.; Graler, M.H.; Levkau, B. Sphingosine-1-Phosphate (S1P) Lyase Inhibition Aggravates Atherosclerosis and Induces Plaque Rupture in ApoE(-/-)Mice. Int. J. Mol. Sci. 2022, 23, 9606. [Google Scholar] [CrossRef] [PubMed]
- Dressler, K.A.; Mathias, S.; Kolesnick, R.N. Tumor necrosis factor-alpha activates the sphingomyelin signal transduction pathway in a cell-free system. Science 1992, 255, 1715–1718. [Google Scholar] [CrossRef]
- Modur, V.; Zimmerman, G.A.; Prescott, S.M.; McIntyre, T.M. Endothelial cell inflammatory responses to tumor necrosis factor alpha. Ceramide-dependent and -independent mitogen-activated protein kinase cascades. J. Biol. Chem. 1996, 271, 13094–13102. [Google Scholar] [CrossRef] [Green Version]
- McEver, R.P.; Moore, K.L.; Cummings, R.D. Leukocyte trafficking mediated by selectin-carbohydrate interactions. J. Biol. Chem. 1995, 270, 11025–11028. [Google Scholar] [CrossRef] [Green Version]
- Haimovitz-Friedman, A.; Cordon-Cardo, C.; Bayoumy, S.; Garzotto, M.; McLoughlin, M.; Gallily, R.; Edwards, C.K., 3rd; Schuchman, E.H.; Fuks, Z.; Kolesnick, R. Lipopolysaccharide induces disseminated endothelial apoptosis requiring ceramide generation. J. Exp. Med. 1997, 186, 1831–1841. [Google Scholar] [CrossRef]
- Escargueil-Blanc, I.; Andrieu-Abadie, N.; Caspar-Bauguil, S.; Brossmer, R.; Levade, T.; Negre-Salvayre, A.; Salvayre, R. Apoptosis and activation of the sphingomyelin-ceramide pathway induced by oxidized low density lipoproteins are not causally related in ECV-304 endothelial cells. J. Biol. Chem. 1998, 273, 27389–27395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada-Shiba, M.; Kinoshita, M.; Kamido, H.; Shimokado, K. Oxidized low density lipoprotein induces apoptosis in cultured human umbilical vein endothelial cells by common and unique mechanisms. J. Biol. Chem. 1998, 273, 9681–9687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holopainen, J.M.; Lehtonen, J.Y.; Kinnunen, P.K. Lipid microdomains in dimyristoylphosphatidylcholine-ceramide liposomes. Chem. Phys. Lipids 1997, 88, 1–13. [Google Scholar] [CrossRef]
- Holopainen, J.M.; Lemmich, J.; Richter, F.; Mouritsen, O.G.; Rapp, G.; Kinnunen, P.K. Dimyristoylphosphatidylcholine/C16:0-ceramide binary liposomes studied by differential scanning calorimetry and wide- and small-angle x-ray scattering. Biophys. J. 2000, 78, 2459–2469. [Google Scholar] [CrossRef] [Green Version]
- Schissel, S.L.; Tweedie-Hardman, J.; Rapp, J.H.; Graham, G.; Williams, K.J.; Tabas, I. Rabbit aorta and human atherosclerotic lesions hydrolyze the sphingomyelin of retained low-density lipoprotein. Proposed role for arterial-wall sphingomyelinase in subendothelial retention and aggregation of atherogenic lipoproteins. J. Clin. Invest. 1996, 98, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.B.; Dey, S.; Shi, W.Y.; Thomas, K.; Hutchins, G.M. Accumulation of glycosphingolipids in human atherosclerotic plaque and unaffected aorta tissues. Glycobiology 1997, 7, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Edsfeldt, A.; Duner, P.; Stahlman, M.; Mollet, I.G.; Asciutto, G.; Grufman, H.; Nitulescu, M.; Persson, A.F.; Fisher, R.M.; Melander, O.; et al. Sphingolipids Contribute to Human Atherosclerotic Plaque Inflammation. Arter. Thromb. Vasc. Biol. 2016, 36, 1132–1140. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.C.; Paultre, F.; Pearson, T.A.; Reed, R.G.; Francis, C.K.; Lin, M.; Berglund, L.; Tall, A.R. Plasma sphingomyelin level as a risk factor for coronary artery disease. Arter. Thromb. Vasc. Biol. 2000, 20, 2614–2618. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ke, C.; Liu, H.; Liu, W.; Li, K.; Yu, B.; Sun, M. Large-scale Metabolomic Analysis Reveals Potential Biomarkers for Early Stage Coronary Atherosclerosis. Sci. Rep. 2017, 7, 11817. [Google Scholar] [CrossRef] [Green Version]
- Hilvo, M.; Simolin, H.; Metso, J.; Ruuth, M.; Oorni, K.; Jauhiainen, M.; Laaksonen, R.; Baruch, A. PCSK9 inhibition alters the lipidome of plasma and lipoprotein fractions. Atherosclerosis 2018, 269, 159–165. [Google Scholar] [CrossRef]
- Croyal, M.; Kaabia, Z.; Leon, L.; Ramin-Mangata, S.; Baty, T.; Fall, F.; Billon-Crossouard, S.; Aguesse, A.; Hollstein, T.; Sullivan, D.R.; et al. Fenofibrate decreases plasma ceramide in type 2 diabetes patients: A novel marker of CVD? Diabetes Metab. 2018, 44, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Poss, A.M.; Maschek, J.A.; Cox, J.E.; Hauner, B.J.; Hopkins, P.N.; Hunt, S.C.; Holland, W.L.; Summers, S.A.; Playdon, M.C. Machine learning reveals serum sphingolipids as cholesterol-independent biomarkers of coronary artery disease. J. Clin. Invest. 2020, 130, 1363–1376. [Google Scholar] [CrossRef] [Green Version]
- Tu, C.; Xie, L.; Wang, Z.; Zhang, L.; Wu, H.; Ni, W.; Li, C.; Li, L.; Zeng, Y. Association between ceramides and coronary artery stenosis in patients with coronary artery disease. Lipids Health Dis. 2020, 19, 151. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bonapace, S.; Lunardi, G.; Canali, G.; Dugo, C.; Vinco, G.; Calabria, S.; Barbieri, E.; Laaksonen, R.; Bonnet, F.; et al. Associations between specific plasma ceramides and severity of coronary-artery stenosis assessed by coronary angiography. Diabetes Metab. 2020, 46, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Meikle, P.J.; Pedersen, E.R.; Tell, G.S.; Dhar, I.; Brenner, H.; Schottker, B.; Laaperi, M.; Kauhanen, D.; Koistinen, K.M.; et al. Development and validation of a ceramide- and phospholipid-based cardiovascular risk estimation score for coronary artery disease patients. Eur. Heart J. 2020, 41, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Toledo, E.; Hruby, A.; Rosner, B.A.; Willett, W.C.; Sun, Q.; Razquin, C.; Zheng, Y.; Ruiz-Canela, M.; Guasch-Ferre, M.; et al. Plasma Ceramides, Mediterranean Diet, and Incident Cardiovascular Disease in the PREDIMED Trial (Prevencion con Dieta Mediterranea). Circulation 2017, 135, 2028–2040. [Google Scholar] [CrossRef] [Green Version]
- Meeusen, J.W.; Donato, L.J.; Bryant, S.C.; Baudhuin, L.M.; Berger, P.B.; Jaffe, A.S. Plasma Ceramides. Arter. Thromb. Vasc. Biol. 2018, 38, 1933–1939. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.R.; Xanthakis, V.; Duncan, M.S.; Gross, S.; Friedrich, N.; Volzke, H.; Felix, S.B.; Jiang, H.; Sidhu, R.; Nauck, M.; et al. Ceramide Remodeling and Risk of Cardiovascular Events and Mortality. J. Am. Heart Assoc 2018, 7. [Google Scholar] [CrossRef]
- Pan, W.; Yu, J.; Shi, R.; Yan, L.; Yang, T.; Li, Y.; Zhang, Z.; Yu, G.; Bai, Y.; Schuchman, E.H.; et al. Elevation of ceramide and activation of secretory acid sphingomyelinase in patients with acute coronary syndromes. Coron Artery Dis. 2014, 25, 230–235. [Google Scholar] [CrossRef]
- Vasile, V.C.; Meeusen, J.W.; Medina Inojosa, J.R.; Donato, L.J.; Scott, C.G.; Hyun, M.S.; Vinciguerra, M.; Rodeheffer, R.R.; Lopez-Jimenez, F.; Jaffe, A.S. Ceramide Scores Predict Cardiovascular Risk in the Community. Arter. Thromb. Vasc. Biol. 2021, 41, 1558–1569. [Google Scholar] [CrossRef]
- Di Paola, M.; Cocco, T.; Lorusso, M. Ceramide interaction with the respiratory chain of heart mitochondria. Biochemistry 2000, 39, 6660–6668. [Google Scholar] [CrossRef] [PubMed]
- Obeid, L.M.; Linardic, C.M.; Karolak, L.A.; Hannun, Y.A. Programmed cell death induced by ceramide. Science 1993, 259, 1769–1771. [Google Scholar] [CrossRef] [PubMed]
- Bekhite, M.; Gonzalez-Delgado, A.; Hubner, S.; Haxhikadrija, P.; Kretzschmar, T.; Muller, T.; Wu, J.M.F.; Bekfani, T.; Franz, M.; Wartenberg, M.; et al. The role of ceramide accumulation in human induced pluripotent stem cell-derived cardiomyocytes on mitochondrial oxidative stress and mitophagy. Free Radic Biol. Med. 2021, 167, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Reforgiato, M.R.; Milano, G.; Fabrias, G.; Casas, J.; Gasco, P.; Paroni, R.; Samaja, M.; Ghidoni, R.; Caretti, A.; Signorelli, P. Inhibition of ceramide de novo synthesis as a postischemic strategy to reduce myocardial reperfusion injury. Basic Res. Cardiol. 2016, 111, 12. [Google Scholar] [CrossRef]
- Perez-Carrillo, L.; Gimenez-Escamilla, I.; Martinez-Dolz, L.; Sanchez-Lazaro, I.J.; Portoles, M.; Rosello-Lleti, E.; Tarazon, E. Implication of Sphingolipid Metabolism Gene Dysregulation and Cardiac Sphingosine-1-Phosphate Accumulation in Heart Failure. Biomedicines 2022, 10, 135. [Google Scholar] [CrossRef]
- Lemaitre, R.N.; Jensen, P.N.; Hoofnagle, A.; McKnight, B.; Fretts, A.M.; King, I.B.; Siscovick, D.S.; Psaty, B.M.; Heckbert, S.R.; Mozaffarian, D.; et al. Plasma Ceramides and Sphingomyelins in Relation to Heart Failure Risk. Circ. Heart Fail 2019, 12, e005708. [Google Scholar] [CrossRef]
- Forouzanfar, M.H.; Liu, P.; Roth, G.A.; Ng, M.; Biryukov, S.; Marczak, L.; Alexander, L.; Estep, K.; Hassen Abate, K.; Akinyemiju, T.F.; et al. Global Burden of Hypertension and Systolic Blood Pressure of at Least 110 to 115 mm Hg, 1990–2015. JAMA 2017, 317, 165–182. [Google Scholar] [CrossRef] [Green Version]
- Meissner, A.; Miro, F.; Jimenez-Altayo, F.; Jurado, A.; Vila, E.; Planas, A.M. Sphingosine-1-phosphate signalling-a key player in the pathogenesis of Angiotensin II-induced hypertension. Cardiovasc. Res. 2017, 113, 123–133. [Google Scholar] [CrossRef]
- Yogi, A.; Callera, G.E.; Aranha, A.B.; Antunes, T.T.; Graham, D.; McBride, M.; Dominiczak, A.; Touyz, R.M. Sphingosine-1-phosphate-induced inflammation involves receptor tyrosine kinase transactivation in vascular cells: Upregulation in hypertension. Hypertension 2011, 57, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Siedlinski, M.; Nosalski, R.; Szczepaniak, P.; Ludwig-Galezowska, A.H.; Mikolajczyk, T.; Filip, M.; Osmenda, G.; Wilk, G.; Nowak, M.; Wolkow, P.; et al. Vascular transcriptome profiling identifies Sphingosine kinase 1 as a modulator of angiotensin II-induced vascular dysfunction. Sci. Rep. 2017, 7, 44131. [Google Scholar] [CrossRef]
- Jujic, A.; Matthes, F.; Vanherle, L.; Petzka, H.; Orho-Melander, M.; Nilsson, P.M.; Magnusson, M.; Meissner, A. Plasma S1P (Sphingosine-1-Phosphate) Links to Hypertension and Biomarkers of Inflammation and Cardiovascular Disease: Findings From a Translational Investigation. Hypertension 2021, 78, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, P.; Carrizzo, A.; Sommella, E.; Oliveti, M.; Iacoviello, L.; Di Castelnuovo, A.; Acernese, F.; Damato, A.; De Lucia, M.; Merciai, F.; et al. Targeting the ASMase/S1P pathway protects from sortilin-evoked vascular damage in hypertension. J. Clin. Invest. 2022, 132, e146343. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, F.; Tan, X.; Wang, H.; Jiang, W.; Wang, X.; Li, S.; Zhang, Y.; Han, Q.; Wang, Y.; et al. Plasma Ceramides and Cardiovascular Events in Hypertensive Patients at High Cardiovascular Risk. Am. J. Hypertens. 2021, 34, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.S.; Yang, L.; Zhang, G.; Zhao, H.; Selim, M.; McCullough, L.D.; Kluk, M.J.; Sanchez, T. Critical role of sphingosine-1-phosphate receptor-2 in the disruption of cerebrovascular integrity in experimental stroke. Nat. Commun. 2015, 6, 7893. [Google Scholar] [CrossRef] [Green Version]
- Pepe, G.; Cotugno, M.; Marracino, F.; Giova, S.; Capocci, L.; Forte, M.; Stanzione, R.; Bianchi, F.; Marchitti, S.; Di Pardo, A.; et al. Differential Expression of Sphingolipid Metabolizing Enzymes in Spontaneously Hypertensive Rats: A Possible Substrate for Susceptibility to Brain and Kidney Damage. Int. J. Mol. Sci. 2021, 22, 3796. [Google Scholar] [CrossRef]
- Zheng, S.; Wei, S.; Wang, X.; Xu, Y.; Xiao, Y.; Liu, H.; Jia, J.; Cheng, J. Sphingosine kinase 1 mediates neuroinflammation following cerebral ischemia. Exp. Neurol. 2015, 272, 160–169. [Google Scholar] [CrossRef]
- Chao, H.C.; Lee, T.H.; Chiang, C.S.; Yang, S.Y.; Kuo, C.H.; Tang, S.C. Sphingolipidomics Investigation of the Temporal Dynamics after Ischemic Brain Injury. J. Proteome Res. 2019, 18, 3470–3478. [Google Scholar] [CrossRef]
- Sheth, S.A.; Iavarone, A.T.; Liebeskind, D.S.; Won, S.J.; Swanson, R.A. Targeted Lipid Profiling Discovers Plasma Biomarkers of Acute Brain Injury. PLoS ONE 2015, 10, e0129735. [Google Scholar] [CrossRef]
- Lee, T.H.; Cheng, C.N.; Chao, H.C.; Lee, C.H.; Kuo, C.H.; Tang, S.C.; Jeng, J.S. Plasma ceramides are associated with outcomes in acute ischemic stroke patients. J. Med. Assoc. 2022, 121, 43–50. [Google Scholar] [CrossRef]
- Lee, T.H.; Cheng, C.N.; Lee, C.W.; Kuo, C.H.; Tang, S.C.; Jeng, J.S. Investigating sphingolipids as biomarkers for the outcomes of acute ischemic stroke patients receiving endovascular treatment. J. Med. Assoc. 2022, 122, 19–28. [Google Scholar] [CrossRef]
- You, Q.; Peng, Q.; Yu, Z.; Jin, H.; Zhang, J.; Sun, W.; Huang, Y. Plasma lipidomic analysis of sphingolipids in patients with large artery atherosclerosis cerebrovascular disease and cerebral small vessel disease. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Bassi, S.; Albizzati, M.G.; Sbacchi, M.; Frattola, L.; Massarotti, M. Double-blind evaluation of monosialoganglioside (GM1) therapy in stroke. J. Neurosci. Res. 1984, 12, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Hoffbrand, B.I.; Bingley, P.J.; Oppenheimer, S.M.; Sheldon, C.D. Trial of ganglioside GM1 in acute stroke. J. Neurol. Neurosurg. Psychiatry 1988, 51, 1213–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argentino, C.; Sacchetti, M.L.; Toni, D.; Savoini, G.; D’Arcangelo, E.; Erminio, F.; Federico, F.; Milone, F.F.; Gallai, V.; Gambi, D.; et al. GM1 ganglioside therapy in acute ischemic stroke. Italian Acute Stroke Study--Hemodilution + Drug. Stroke 1989, 20, 1143–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldi, C.; Masi, M.C.; Manetti, M.; Carabelli, E.; Martini, A. A pilot study with monosialoganglioside GM1 on acute cerebral ischemia. Acta Neurol. 1990, 12, 214–221. [Google Scholar]
- Lenzi, G.L.; Grigoletto, F.; Gent, M.; Roberts, R.S.; Walker, M.D.; Easton, J.D.; Carolei, A.; Dorsey, F.C.; Rocca, W.A.; Bruno, R.; et al. Early treatment of stroke with monosialoganglioside GM-1. Efficacy and safety results of the Early Stroke Trial. Stroke 1994, 25, 1552–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocca, W.A.; Dorsey, F.C.; Grigoletto, F.; Gent, M.; Roberts, R.S.; Walker, M.D.; Easton, J.D.; Bruno, R.; Carolei, A.; Sancesario, G.; et al. Design and baseline results of the monosialoganglioside early stroke trial. The EST Study Group. Stroke 1992, 23, 519–526. [Google Scholar] [CrossRef] [Green Version]
- The SASS Trial. Ganglioside GM1 in acute ischemic stroke. Stroke 1994, 25, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Cantalupo, A.; Zhang, Y.; Kothiya, M.; Galvani, S.; Obinata, H.; Bucci, M.; Giordano, F.J.; Jiang, X.C.; Hla, T.; Di Lorenzo, A. Nogo-B regulates endothelial sphingolipid homeostasis to control vascular function and blood pressure. Nat. Med. 2015, 21, 1028–1037. [Google Scholar] [CrossRef] [Green Version]
- Theilmeier, G.; Schmidt, C.; Herrmann, J.; Keul, P.; Schafers, M.; Herrgott, I.; Mersmann, J.; Larmann, J.; Hermann, S.; Stypmann, J.; et al. High-density lipoproteins and their constituent, sphingosine-1-phosphate, directly protect the heart against ischemia/reperfusion injury in vivo via the S1P3 lysophospholipid receptor. Circulation 2006, 114, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Komarova, Y.A.; Mehta, D.; Malik, A.B. Dual regulation of endothelial junctional permeability. Sci. STKE 2007, 2007, re8. [Google Scholar] [CrossRef] [PubMed]
- Kerage, D.; Brindley, D.N.; Hemmings, D.G. Review: Novel insights into the regulation of vascular tone by sphingosine 1-phosphate. Placenta 2014, 35, S86–S92. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Chen, J.; Pang, L.; Chen, C.; Ye, J.; Liu, H.; Chen, H.; Zhang, S.; Liu, S.; Liu, B.; et al. The Acid Sphingomyelinase Inhibitor Amitriptyline Ameliorates TNF-alpha-Induced Endothelial Dysfunction. Cardiovasc. Drugs 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Zou, A.P.; Li, P.L. Ceramide reduces endothelium-dependent vasodilation by increasing superoxide production in small bovine coronary arteries. Circ. Res. 2001, 88, 824–831. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Li, W.; Wang, J.; Altura, B.T.; Altura, B.M. Sphingomyelinase and ceramide analogs induce contraction and rises in [Ca(2+)](i) in canine cerebral vascular muscle. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H1421–H1428. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Junk, P.; Huwiler, A.; Burkhardt, C.; Wallerath, T.; Pfeilschifter, J.; Forstermann, U. Dual effect of ceramide on human endothelial cells: Induction of oxidative stress and transcriptional upregulation of endothelial nitric oxide synthase. Circulation 2002, 106, 2250–2256. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, A.K.; Han, H.; Snowden, A.; Chatterjee, S. Redox-regulated signaling by lactosylceramide in the proliferation of human aortic smooth muscle cells. J. Biol. Chem. 1997, 272, 15642–15649. [Google Scholar] [CrossRef] [Green Version]
- Mollnau, H.; Wendt, M.; Szocs, K.; Lassegue, B.; Schulz, E.; Oelze, M.; Li, H.; Bodenschatz, M.; August, M.; Kleschyov, A.L.; et al. Effects of angiotensin II infusion on the expression and function of NAD(P)H oxidase and components of nitric oxide/cGMP signaling. Circ Res 2002, 90, E58–E65. [Google Scholar] [CrossRef] [Green Version]
- Bauersachs, J.; Schafer, A. Tetrahydrobiopterin and eNOS dimer/monomer ratio—A clue to eNOS uncoupling in diabetes? Cardiovasc. Res. 2005, 65, 768–769. [Google Scholar] [CrossRef]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Gray, C.W.; Coster, A.C.F. From insulin to Akt: Time delays and dominant processes. J. Biol. 2020, 507, 110454. [Google Scholar] [CrossRef] [PubMed]
- Rowland, A.F.; Fazakerley, D.J.; James, D.E. Mapping insulin/GLUT4 circuitry. Traffic 2011, 12, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.A.; Knotts, T.A.; Wang, L.P.; Li, G.; Dobrowsky, R.T.; Florant, G.L.; Summers, S.A. A role for ceramide, but not diacylglycerol, in the antagonism of insulin signal transduction by saturated fatty acids. J. Biol. Chem. 2003, 278, 10297–10303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, R.S.; Orena, S.J.; Rafidi, K.; Torchia, A.J.; Stock, J.L.; Hildebrandt, A.L.; Coskran, T.; Black, S.C.; Brees, D.J.; Wicks, J.R.; et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKB beta. J. Clin. Invest. 2003, 112, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Powell, D.J.; Hajduch, E.; Kular, G.; Hundal, H.S. Ceramide disables 3-phosphoinositide binding to the pleckstrin homology domain of protein kinase B (PKB)/Akt by a PKCzeta-dependent mechanism. Mol. Cell Biol. 2003, 23, 7794–7808. [Google Scholar] [CrossRef] [Green Version]
- Zinda, M.J.; Vlahos, C.J.; Lai, M.T. Ceramide induces the dephosphorylation and inhibition of constitutively activated Akt in PTEN negative U87 mg cells. Biochem. Biophys. Res. Commun. 2001, 280, 1107–1115. [Google Scholar] [CrossRef]
- Teruel, T.; Hernandez, R.; Lorenzo, M. Ceramide mediates insulin resistance by tumor necrosis factor-alpha in brown adipocytes by maintaining Akt in an inactive dephosphorylated state. Diabetes 2001, 50, 2563–2571. [Google Scholar] [CrossRef] [Green Version]
- Resjo, S.; Goransson, O.; Harndahl, L.; Zolnierowicz, S.; Manganiello, V.; Degerman, E. Protein phosphatase 2A is the main phosphatase involved in the regulation of protein kinase B in rat adipocytes. Cell Signal 2002, 14, 231–238. [Google Scholar] [CrossRef]
- Paz, K.; Hemi, R.; LeRoith, D.; Karasik, A.; Elhanany, E.; Kanety, H.; Zick, Y. A molecular basis for insulin resistance. Elevated serine/threonine phosphorylation of IRS-1 and IRS-2 inhibits their binding to the juxtamembrane region of the insulin receptor and impairs their ability to undergo insulin-induced tyrosine phosphorylation. J. Biol. Chem. 1997, 272, 29911–29918. [Google Scholar] [CrossRef] [Green Version]
- Peraldi, P.; Hotamisligil, G.S.; Buurman, W.A.; White, M.F.; Spiegelman, B.M. Tumor necrosis factor (TNF)-alpha inhibits insulin signaling through stimulation of the p55 TNF receptor and activation of sphingomyelinase. J. Biol. Chem. 1996, 271, 13018–13022. [Google Scholar] [CrossRef] [Green Version]
- Kanety, H.; Hemi, R.; Papa, M.Z.; Karasik, A. Sphingomyelinase and ceramide suppress insulin-induced tyrosine phosphorylation of the insulin receptor substrate-1. J. Biol. Chem. 1996, 271, 9895–9897. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.A. Ceramides in insulin resistance and lipotoxicity. Prog. Lipid Res. 2006, 45, 42–72. [Google Scholar] [CrossRef] [PubMed]
- Denhez, B.; Rousseau, M.; Spino, C.; Dancosst, D.A.; Dumas, M.E.; Guay, A.; Lizotte, F.; Geraldes, P. Saturated fatty acids induce insulin resistance in podocytes through inhibition of IRS1 via activation of both IKKbeta and mTORC1. Sci. Rep. 2020, 10, 21628. [Google Scholar] [CrossRef]
- Ruvolo, P.P. Intracellular signal transduction pathways activated by ceramide and its metabolites. Pharm. Res. 2003, 47, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Gual, P.; Le Marchand-Brustel, Y.; Tanti, J.F. Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie 2005, 87, 99–109. [Google Scholar] [CrossRef]
- Aguirre, V.; Werner, E.D.; Giraud, J.; Lee, Y.H.; Shoelson, S.E.; White, M.F. Phosphorylation of Ser307 in insulin receptor substrate-1 blocks interactions with the insulin receptor and inhibits insulin action. J. Biol. Chem. 2002, 277, 1531–1537. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Wang, W.; Li, N.; Yan, S.; Rong, K.; Lan, T.; Xia, P. Sphingosine kinase 2 promotes lipotoxicity in pancreatic beta-cells and the progression of diabetes. FASEB J. 2019, 33, 3636–3646. [Google Scholar] [CrossRef]
- Aji, G.; Huang, Y.; Ng, M.L.; Wang, W.; Lan, T.; Li, M.; Li, Y.; Chen, Q.; Li, R.; Yan, S.; et al. Regulation of hepatic insulin signaling and glucose homeostasis by sphingosine kinase 2. Proc. Natl. Acad. Sci. USA 2020, 117, 24434–24442. [Google Scholar] [CrossRef]
- Poitout, V.; Robertson, R.P. Glucolipotoxicity: Fuel excess and beta-cell dysfunction. Endocr. Rev. 2008, 29, 351–366. [Google Scholar] [CrossRef]
- van Herpen, N.A.; Schrauwen-Hinderling, V.B. Lipid accumulation in non-adipose tissue and lipotoxicity. Physiol. Behav. 2008, 94, 231–241. [Google Scholar] [CrossRef]
- Veret, J.; Bellini, L.; Giussani, P.; Ng, C.; Magnan, C.; Le Stunff, H. Roles of Sphingolipid Metabolism in Pancreatic beta Cell Dysfunction Induced by Lipotoxicity. J. Clin. Med. 2014, 3, 646–662. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Qian, Y.; Xi, X.; Hu, X.; Zhu, J.; Han, X. Blockage of ceramide metabolism exacerbates palmitate inhibition of pro-insulin gene expression in pancreatic beta-cells. Mol. Cell Biochem. 2010, 338, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Fontes, G.; Semache, M.; Hagman, D.K.; Tremblay, C.; Shah, R.; Rhodes, C.J.; Rutter, J.; Poitout, V. Involvement of Per-Arnt-Sim Kinase and extracellular-regulated kinases-1/2 in palmitate inhibition of insulin gene expression in pancreatic beta-cells. Diabetes 2009, 58, 2048–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boslem, E.; Weir, J.M.; MacIntosh, G.; Sue, N.; Cantley, J.; Meikle, P.J.; Biden, T.J. Alteration of endoplasmic reticulum lipid rafts contributes to lipotoxicity in pancreatic beta-cells. J. Biol. Chem. 2013, 288, 26569–26582. [Google Scholar] [CrossRef] [Green Version]
- Veluthakal, R.; Palanivel, R.; Zhao, Y.; McDonald, P.; Gruber, S.; Kowluru, A. Ceramide induces mitochondrial abnormalities in insulin-secreting INS-1 cells: Potential mechanisms underlying ceramide-mediated metabolic dysfunction of the beta cell. Apoptosis 2005, 10, 841–850. [Google Scholar] [CrossRef]
- Syed, I.; Szulc, Z.M.; Ogretmen, B.; Kowluru, A. L- threo -C6-pyridinium-ceramide bromide, a novel cationic ceramide, induces NADPH oxidase activation, mitochondrial dysfunction and loss in cell viability in INS 832/13 beta-cells. Cell Physiol. Biochem. 2012, 30, 1051–1058. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [Green Version]
- Verheij, M.; Bose, R.; Lin, X.H.; Yao, B.; Jarvis, W.D.; Grant, S.; Birrer, M.J.; Szabo, E.; Zon, L.I.; Kyriakis, J.M.; et al. Requirement for ceramide-initiated SAPK/JNK signalling in stress-induced apoptosis. Nature 1996, 380, 75–79. [Google Scholar] [CrossRef]
- Lang, F.; Ullrich, S.; Gulbins, E. Ceramide formation as a target in beta-cell survival and function. Expert Opin. Targets 2011, 15, 1061–1071. [Google Scholar] [CrossRef]
- Mandal, N.; Grambergs, R.; Mondal, K.; Basu, S.K.; Tahia, F.; Dagogo-Jack, S. Role of ceramides in the pathogenesis of diabetes mellitus and its complications. J. Diabetes Complicat. 2021, 35, 107734. [Google Scholar] [CrossRef]
- Cnop, M.; Welsh, N.; Jonas, J.C.; Jorns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54 (Suppl. 2), S97–S107. [Google Scholar] [CrossRef] [PubMed]
- Alshehry, Z.H.; Mundra, P.A.; Barlow, C.K.; Mellett, N.A.; Wong, G.; McConville, M.J.; Simes, J.; Tonkin, A.M.; Sullivan, D.R.; Barnes, E.H.; et al. Plasma Lipidomic Profiles Improve on Traditional Risk Factors for the Prediction of Cardiovascular Events in Type 2 Diabetes Mellitus. Circulation 2016, 134, 1637–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, H.; Sun, L.; Wu, Q.; Zong, G.; Qi, Q.; Li, H.; Zheng, H.; Zeng, R.; Liang, L.; Lin, X. Associations among circulating sphingolipids, beta-cell function, and risk of developing type 2 diabetes: A population-based cohort study in China. PLoS Med. 2020, 17, e1003451. [Google Scholar] [CrossRef] [PubMed]
- Wigger, L.; Cruciani-Guglielmacci, C.; Nicolas, A.; Denom, J.; Fernandez, N.; Fumeron, F.; Marques-Vidal, P.; Ktorza, A.; Kramer, W.; Schulte, A.; et al. Plasma Dihydroceramides Are Diabetes Susceptibility Biomarker Candidates in Mice and Humans. Cell Rep. 2017, 18, 2269–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, W.S.; Torta, F.; Ji, S.; Choi, H.; Begum, H.; Sim, X.; Khoo, C.M.; Khoo, E.Y.H.; Ong, W.Y.; Van Dam, R.M.; et al. Large-scale lipidomics identifies associations between plasma sphingolipids and T2DM incidence. JCI Insight 2019, 5, e126925. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, W.; Xia, M.F.; Lu, Y.L.; Bian, H.; Yu, C.; Li, X.Y.; Vadas, M.A.; Gao, X.; Lin, H.D.; et al. Identification of circulating sphingosine kinase-related metabolites for prediction of type 2 diabetes. J. Transl. Med. 2021, 19, 393. [Google Scholar] [CrossRef] [PubMed]
- Fridman, V.; Zarini, S.; Sillau, S.; Harrison, K.; Bergman, B.C.; Feldman, E.L.; Reusch, J.E.B.; Callaghan, B.C. Altered plasma serine and 1-deoxydihydroceramide profiles are associated with diabetic neuropathy in type 2 diabetes and obesity. J. Diabetes Complicat. 2021, 35, 107852. [Google Scholar] [CrossRef]
- Mwinyi, J.; Bostrom, A.; Fehrer, I.; Othman, A.; Waeber, G.; Marti-Soler, H.; Vollenweider, P.; Marques-Vidal, P.; Schioth, H.B.; von Eckardstein, A.; et al. Plasma 1-deoxysphingolipids are early predictors of incident type 2 diabetes mellitus. PLoS ONE 2017, 12, e0175776. [Google Scholar] [CrossRef]
- Othman, A.; Saely, C.H.; Muendlein, A.; Vonbank, A.; Drexel, H.; von Eckardstein, A.; Hornemann, T. Plasma 1-deoxysphingolipids are predictive biomarkers for type 2 diabetes mellitus. BMJ Open Diabetes Res. Care 2015, 3, e000073. [Google Scholar] [CrossRef] [Green Version]
- Penno, A.; Reilly, M.M.; Houlden, H.; Laura, M.; Rentsch, K.; Niederkofler, V.; Stoeckli, E.T.; Nicholson, G.; Eichler, F.; Brown, R.H., Jr.; et al. Hereditary sensory neuropathy type 1 is caused by the accumulation of two neurotoxic sphingolipids. J. Biol. Chem. 2010, 285, 11178–11187. [Google Scholar] [CrossRef] [Green Version]
- Gantner, M.L.; Eade, K.; Wallace, M.; Handzlik, M.K.; Fallon, R.; Trombley, J.; Bonelli, R.; Giles, S.; Harkins-Perry, S.; Heeren, T.F.C.; et al. Serine and Lipid Metabolism in Macular Disease and Peripheral Neuropathy. N. Engl. J. Med. 2019, 381, 1422–1433. [Google Scholar] [CrossRef] [PubMed]
- Guntert, T.; Hanggi, P.; Othman, A.; Suriyanarayanan, S.; Sonda, S.; Zuellig, R.A.; Hornemann, T.; Ogunshola, O.O. 1-Deoxysphingolipid-induced neurotoxicity involves N-methyl-d-aspartate receptor signaling. Neuropharmacology 2016, 110, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.R.; Kugathasan, U.; Abramov, A.Y.; Clark, A.J.; Bennett, D.L.H.; Reilly, M.M.; Greensmith, L.; Kalmar, B. Hereditary sensory neuropathy type 1-associated deoxysphingolipids cause neurotoxicity, acute calcium handling abnormalities and mitochondrial dysfunction in vitro. Neurobiol. Dis. 2018, 117, 1–14. [Google Scholar] [CrossRef]
- Zuellig, R.A.; Hornemann, T.; Othman, A.; Hehl, A.B.; Bode, H.; Guntert, T.; Ogunshola, O.O.; Saponara, E.; Grabliauskaite, K.; Jang, J.H.; et al. Deoxysphingolipids, novel biomarkers for type 2 diabetes, are cytotoxic for insulin-producing cells. Diabetes 2014, 63, 1326–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, D.; Myers, S.; McGowan, C.; Henstridge, D.; Eri, R.; Sonda, S.; Caruso, V. 1-Deoxysphingolipids, Early Predictors of Type 2 Diabetes, Compromise the Functionality of Skeletal Myoblasts. Front. Endocrinol. 2021, 12, 772925. [Google Scholar] [CrossRef]
- Hammerschmidt, P.; Bruning, J.C. Contribution of specific ceramides to obesity-associated metabolic diseases. Cell Mol. Life Sci. 2022, 79, 395. [Google Scholar] [CrossRef]
- Turpin-Nolan, S.M.; Bruning, J.C. The role of ceramides in metabolic disorders: When size and localization matters. Nat. Rev. Endocrinol. 2020, 16, 224–233. [Google Scholar] [CrossRef]
- Raichur, S.; Brunner, B.; Bielohuby, M.; Hansen, G.; Pfenninger, A.; Wang, B.; Bruning, J.C.; Larsen, P.J.; Tennagels, N. The role of C16:0 ceramide in the development of obesity and type 2 diabetes: CerS6 inhibition as a novel therapeutic approach. Mol. Metab. 2019, 21, 36–50. [Google Scholar] [CrossRef]
- Turpin, S.M.; Nicholls, H.T.; Willmes, D.M.; Mourier, A.; Brodesser, S.; Wunderlich, C.M.; Mauer, J.; Xu, E.; Hammerschmidt, P.; Bronneke, H.S.; et al. Obesity-induced CerS6-dependent C16:0 ceramide production promotes weight gain and glucose intolerance. Cell Metab. 2014, 20, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Turpin-Nolan, S.M.; Hammerschmidt, P.; Chen, W.; Jais, A.; Timper, K.; Awazawa, M.; Brodesser, S.; Bruning, J.C. CerS1-Derived C18:0 Ceramide in Skeletal Muscle Promotes Obesity-Induced Insulin Resistance. Cell Rep. 2019, 26, 1–10e17. [Google Scholar] [CrossRef] [Green Version]
- Blachnio-Zabielska, A.U.; Roszczyc-Owsiejczuk, K.; Imierska, M.; Pogodzinska, K.; Rogalski, P.; Daniluk, J.; Zabielski, P. CerS1 but Not CerS5 Gene Silencing, Improves Insulin Sensitivity and Glucose Uptake in Skeletal Muscle. Cells 2022, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.Z.; Garske, K.M.; Alvarez, M.; Bhagat, Y.V.; Boocock, J.; Nikkola, E.; Miao, Z.; Raulerson, C.K.; Cantor, R.M.; Civelek, M.; et al. Integration of human adipocyte chromosomal interactions with adipose gene expression prioritizes obesity-related genes from GWAS. Nat. Commun. 2018, 9, 1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Zan, W.; Qin, L.; Han, S.; Ye, L.; Wang, M.; Jiang, B.; Fang, P.; Liu, Q.; Shao, C.; et al. Ablation of ORMDL3 impairs adipose tissue thermogenesis and insulin sensitivity by increasing ceramide generation. Mol. Metab. 2022, 56, 101423. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Bui, H.H.; Sanders, P.; Siddall, P.; Wei, T.; Thomas, M.K.; et al. Muscle sphingolipids during rest and exercise: A C18:0 signature for insulin resistance in humans. Diabetologia 2016, 59, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Bui, H.H.; Sanders, P.; Siddall, P.; Kuo, M.S.; Perreault, L. Serum sphingolipids: Relationships to insulin sensitivity and changes with exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E398–E408 . [Google Scholar] [CrossRef] [Green Version]
- Perreault, L.; Newsom, S.A.; Strauss, A.; Kerege, A.; Kahn, D.E.; Harrison, K.A.; Snell-Bergeon, J.K.; Nemkov, T.; D’Alessandro, A.; Jackman, M.R.; et al. Intracellular localization of diacylglycerols and sphingolipids influences insulin sensitivity and mitochondrial function in human skeletal muscle. JCI Insight 2018, 3, e96805. [Google Scholar] [CrossRef] [Green Version]
- Kasumov, T.; Solomon, T.P.; Hwang, C.; Huang, H.; Haus, J.M.; Zhang, R.; Kirwan, J.P. Improved insulin sensitivity after exercise training is linked to reduced plasma C14:0 ceramide in obesity and type 2 diabetes. Obesity 2015, 23, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Warshauer, J.T.; Lopez, X.; Gordillo, R.; Hicks, J.; Holland, W.L.; Anuwe, E.; Blankfard, M.B.; Scherer, P.E.; Lingvay, I. Effect of pioglitazone on plasma ceramides in adults with metabolic syndrome. Diabetes Metab. Res. Rev. 2015, 31, 734–744. [Google Scholar] [CrossRef]
- Hanamatsu, H.; Ohnishi, S.; Sakai, S.; Yuyama, K.; Mitsutake, S.; Takeda, H.; Hashino, S.; Igarashi, Y. Altered levels of serum sphingomyelin and ceramide containing distinct acyl chains in young obese adults. Nutr. Diabetes 2014, 4, e141. [Google Scholar] [CrossRef] [Green Version]
- Mamtani, M.; Meikle, P.J.; Kulkarni, H.; Weir, J.M.; Barlow, C.K.; Jowett, J.B.; Bellis, C.; Dyer, T.D.; Almasy, L.; Mahaney, M.C.; et al. Plasma dihydroceramide species associate with waist circumference in Mexican American families. Obesity 2014, 22, 950–956. [Google Scholar] [CrossRef]


{kind=link}
| Clinical Condition | Study | Model | Major Findings |
|---|---|---|---|
| Atherosclerosis and CAD | Jiang et al. [48] | Plasma from 279 biethnic patients with coronary artery disease (CAD) was related with 277 controls. | Higher plasma sphingomyelin (SM) level in CAD patients than in control subjects; higher ratio of SM to SM + phosphatidylcholine (PC) in patients than in controls. |
| Gao et al. [49] | Plasma from 100 patients with Coronary atherosclerosis and 100 controls was compared. | Increased levels of 24 metabolites and decreased of 18 metabolites in early CAS patients compared with the controls; nine metabolites resulted useful as biomarkers to distinguish early-stage CAS patients from controls. | |
| Hilvo et al. [50] | Plasma of non-diabetic patients with established coronary heart disease (CHD), stratified in 25 patients receiving RG7652 (PCSK9 inhibitor) and 15 placebo, was analyzed. | PCSK9 inhibition decrease plasma levels of several lipid classes, including sphingolipids (dihydroceramides, glucosylceramides, sphingomyelins, ceramides), cholesteryl esters and free cholesterol. | |
| Croyal et al. [51] | Plasma from 102 patients with T2DM before and after fenofibrate treatment (200 mg/day) was correlated. | Ceramide levels decreased in 73.5% of patients; reduction of plasma apoC-II, apoC-III, apoB100, SMase, with an increase in apoA-II and adiponectin levels. | |
| Poss et al. [52] | Correlation between serum of 462 individuals with familial coronary artery disease (CAD) and 212 population-based controls. | 30 sphingolipids were found elevated in serum of patients with CAD compared with healthy controls. | |
| Tu et al. [53] | Plasma from 553 patients with definite or suspected CAD. | High ratio of Cer (d18:1/24:1) to Cer (d18:1/24:0), female gender, HbA1c%, unstable angina (UAP) and acute myocardial infarction (AMI) diagnosis during hospitalization were related with severe coronary artery stenosis. | |
| Mantovani et al. [54] | Plasma from 167 patients with established or suspected CAD, who underwent urgent or elective coronary angiography. | Higher levels of plasma Cer(d18:1/20:0), Cer(d18:1/22:0) and Cer(d18:1/24:0) were associated with the presence of LAD stenosis ≥ 50%. | |
| Hilvo et al. [55] | Plasma from 3789 individuals of the WECAC (The Western Norway Coronary Angiography Cohort), 5991 individuals of the LIPID (Long-Term Intervention with Pravastatin in Ischaemic Disease) trial, and 1023 individuals of the KAROLA (Langzeiterfolge der KARdiOLogischen Anschlussheilbehandlung) | Higher ceramides levels were found in subjects with CAD. | |
| Laaksonen et al. [8] | Plasma from 160 individuals of the Corogene cohort study including stable CAD patients; 1637 individuals of the prospective Special Program University Medicine—Inflammation in Acute Coronary Syndromes (SPUM-ACS) cohort. | Ceramides were associated with CV death in all studies, independent of other lipid markers and C-reactive protein. | |
| Wang et al. [56] | Plasma from 980 participants from the PREDIMED trial (Prevención con Dieta Mediterránea); 230 incident cases of CVD and 787 randomly selected participants at baseline (including 37 overlapping cases). | The ceramide score, calculated as a weighted sum of concentrations of four ceramides, was correlated with a 2.18-fold higher risk of CVD. | |
| Meeusen et al. [57] | Plasma from 495 participants before nonurgent coronary angiography. | Coronary artery disease was identified in 265 (54%) cases; the hazard ratios were 1.50 (1.16–1.93) for Cer(16:0), 1.42 (1.11–1.83) for Cer(18:0), 1.43 (1.08–1.89) for Cer(24:1), and 1.58 (1.22–2.04) for the ceramide risk score. | |
| Peterson et al. [58] | Plasma from 2642 participants of the Framingham Heart Study (FHS) and 3134 participants of the Study of Health in Pomerania (SHIP). | During a mean follow-up of 6 years in FHS, there were 88 coronary heart disease (CHD) and 90 heart failure (HF) events and 239 deaths. During a median follow-up time of 5.75 years for CHD and HF and 8.24 years for mortality, in SHIP there were 209 CHD and 146 HF events and 377 deaths. C24:0/C16:0 ceramide ratios were inversely associated with incident CHD and inversely associated with incident HF; C24:0/C16:0 and C22:0/C16:0 ceramide ratios were inversely associated with all-cause mortality. | |
| Pan et al. [59] | Plasma from 304 patients and 52 healthy individuals divided in four groups: 52 control group, 98b stable angina pectoris (SAP) group, 92 unstable angina pectoris (UAP) group, 114 and acute myocardial infarction (AMI) group. | Higher levels of sphingomyelin (SPM) in patients with UAP and AMI compared with the controls and SAP participants. Higher ceramide levels and S-SMase activity in patients with UAP and AMI than controls and SAP participants. | |
| Heart failure | Ji et al. [12] | Serum and myocardial tissue from 65 patients with advanced heart failure (HF). | Increase of long–chain ceramides in myocardium and serum of patients with advanced HF. Blocking the serine palmitoyl transferase (SPT), the enzyme of the pathway of ceramide synthesis, higher ceramides were found. |
| Anroedh et al. [16] | Blood from 581 patients who underwent diagnostic coronary angiography or percutaneous coronary intervention for stable angina pectoris (SAP) or acute coronary syndrome (ACS). | During a median follow-up of 4.7 years, 155 patients (27%) had MACEs. Cer(d18:1/16:0) concentration was related with MACEs; concentrations of Cer(d18:1/16:0), Cer(d18:1/20:0), Cer(d18:1/24:1), and their ratios to Cer(d18:1/24:0) were associated with the composite endpoint death or nonfatal ACS. | |
| Perez-Carrillo et al. [65] | Myocardial tissue samples from the left ventricle of 52 subjects: 42 patients with HF (non-ischemic dilated and ischemic cardiomyopathy patients) undergoing cardiac transplantation and 10 samples from non-diseased donor hearts. | Sphingolipid metabolism gene dysregulation was found in HF human cardiac tissue, with the major changes occurring in the expression of genes involved in the de novo and salvage pathways; S1P is enhanced in HF cardiac tissue. | |
| Lemaitre et al. [66] | Plasma from 4249 patients. | Identified 1179 cases of incident heart failure among 4249 study participants. With Cox regression, higher levels of Cer-16 and SM-16 were correlated with higher risk of incident heart failure. In contrast, higher levels of Cer-22, SM-20, SM-22 and SM-24 were correlated with lower risk of heart failure. | |
| Hypertension | Spijkers et al. [7] | Plasma from 12 patients with stage 1 hypertension; 19 patients with stage 2 and 3 hypertension and 18 normotensive controls. | Higher ceramide levels in patients with stage 2 or 3 hypertension compared to healthy normotensive controls. |
| Jujic et al. [71] | Plasma from 1046 individuals of the Malmö Offspring Study (MOS). | Individuals with systolic BP ≥140 mm Hg had higher S1P plasma concentrations compared with subjects with BP <120 mm Hg independent of age and sex. | |
| Di Pietro et al. [72] | Plasma from the Campania Salute Network Registry divided in 71 patients with hypertension, 36 healthy donor control subjects and 36 hypertensive patients with no diagnosis of PAD. Plasma from 270 individuals of the Moli-Sani study stratified into normotensive subjects (n = 89) and controlled (n = 91) or uncontrolled hypertensive subjects (n = 90). | Increased plasma ASMase activity and levels of S1P, in hypertensive subjects; the increase was more pronounced in hypertensive subjects with uncontrolled blood pressure. | |
| Yin et al. [73] | Plasma from 920 patients with hypertension. | During mean 2.3-year follow-up, 71 patients had MACE. Cer(d18:1/16:0), Cer(d18:1/22:0), and Cer(d18:1/24:0) were highly significant in predicting MACE. | |
| Stroke | Sheth et al. [78] | Plasma from 14 patients with acute stroke symptoms. | Higher sphingolipid scores in patients with true stroke compared to patients who hsd non-stroke causes of their symptoms. |
| Lee et al. [79] | Plasma from 87 patients with acute ischemic stroke (AIS) and 30 nonstroke controls. | Decreased S1P and very-long-chain ceramides in AIS patients compared to non-stroke controls; increased long-chain ceramides in AIS patients. | |
| Lee et al. [80] | Plasma from 75 AIS patients who underwent endovascular thrombectomy before (T1), immediately after (T2), and 24 h after (T3) the procedures and 19 controls. | Higher plasma levels of long-chain ceramides Cer (d18:1/16:0) at all three time points, Cer (d18:1/18:0) at T1 and T3, and Cer (d18:1/20:0) at T1 and very-long-chain ceramide Cer (d18:1/24:1) at T1 in AIS patients compared to controls. Lower plasma levels of sphingosine-1-phosphate in AIS patients compared to controls at all three time points. | |
| You et al. [81] | Plasma from 20 patients with large artery atherosclerosis (LAA), 20 patients with age-related cerebral small vessel disease (CSVD), 10 patients with Fabry disease and 14 controls. | Cer (d36:3), Cer (d34:2), Cer (d38:6), Cer (d36:4) and Cer (d16:0/18:1) were increased in LAA; SM (d34:1), Cer (d34:2), Cer (d36:4), Cer (d16:0/18:1), Cer (d38:6), Cer (d36:3) and Cer (d32:0) were increased in age-related CSVD. Cer (d36:4) and SM (d34:1) were increased in age-related CSVD compared with LAA. Total trihexosyl ceramides were higher in Fabry group compared with control; SM (d34:1) was increased in Fabry group. | |
| Vascular dysfunction | Siedlinski et al. [70] | Serum from 82 patients. | Higher S1P serum levels were correlated with impaired endothelial function as well as with increased vessels contractility. |
| Diabetes | Alshehry et al. [132] | Plasma from 3779 patients with T2DM. | Plasma lipids species sphingolipids, phospholipids (including lyso- and ether- species), cholesteryl esters, and glycerolipids were related to future cardiovascular events and cardiovascular death. |
| Yun et al. [133] | Plasma from 1974 individuals followed-up for 6 years T2DM. | 11 novel and 3 reported sphingolipids, namely ceramides (d18:1/18:1, d18:1/20:0, d18:1/20:1, d18:1/22:1), saturated sphingomyelins (C34:0, C36:0, C38:0, C40:0), unsaturated sphingomyelins (C34:1, C36:1, C42:3), hydroxyl-sphingomyelins (C34:1, C38:3), and a hexosylceramide (d18:1/20:1), were associated with incident T2DM. | |
| Wigger et al. [134] | Plasma from 250 individuals of two different prospective cohorts who developed T2DM. | Specific long-chain fatty-acid-containing dihydroceramides were higher in the plasma of individuals up to 9 years before disease onset. | |
| Chew et al. [135] | Plasma from plasma of 2302 ethnically-Chinese Singaporeans. | Two distinct sphingomyelins, d16:1/C18:0 and d18:1/C18:0 were related to a higher risk of T2DM. | |
| Chen et al. [136] | Serum from a total of 2486 non-diabetic adults at baseline, 100 subjects who developed T2DM after a mean follow-up of 4.2-years and 100 control subjects matched strictly with age, sex, BMI and fasting glucose. | Compared to the control group, medians of serum dhS1P and dhS1P/dhSph ratio at baseline were elevated prior to the onset of T2DM. Each SD increment of dhS1P and dhS1P/dhSph ratio was related to 53.5% and 54.1% higher risk of incident diabetes, respectively. | |
| Fridman et al. [137] | Plasma from 75 individuals stratified in: 19 lean controls (LC), 19 with obesity, 18 with obesity and T2DM without diabetic neuropathy (DN) (ob/T2DM), and 19 with obesity, T2DM and DN (Ob/T2DM/DN). | Increased 1-deoxydihydroceramides across these four groups. 1-deoxydihydroceramide species were higher in ob/T2DM/DN versus LC group. No significant differences in 1-deoxydihydroceramides were found between the ob/T2DM and ob/T2DM/DN groups. | |
| Mwinyi et al. [138] | Plasma from 605 non-diabetic individuals. | 1-deoxysphingolipids were higher in individuals who developed T2DM during 5-year follow-up; 1-deoxy-sphinganine and 1-deoxy-sphingosine were predictive for T2DM. | |
| Othman et al. [139] | Plasma from 339 individuals. | Increased 1-deoxysphingolipids levels in patients with metabolic syndrome, impaired fasting glucose, and T2DM. Patients who developed T2DM during 8-year follow-up period showed an increase in 1-deoxysphingolipids levels at baseline compared with those who did not develop T2DM until the end of the study. | |
| Obesity | Turpin et al. [149] | White adipose tissue (WAT) from 439 obese subjects. | CERS6 mRNA expression and C16:0 ceramides were elevated in adipose tissue of obese humans, and increased CERS6 expression correlates with insulin resistance. |
| Pan et al. [152] | Subcutaneous abdominal adipose needle biopsy from 10,197 individuals. | Identified the SPT suppressor ORMDL3 as an obesity-related gene, and its expression in human subcutaneous WAT was inversely correlated with BMI. | |
| Bergman et al. [154] | Serum and percutaneous needle biopsy between the greater trochanter of the femur and the patella was taken from 14 obese sedentary individuals, 15 type 2 diabetic patients and 15 endurance trained athletes. | Muscle C18:0 ceramide, dihydroceramide and glucosylceramide species were inversely associated with insulin sensitivity without differences in total ceramide, dihydroceramide, and glucosylceramide concentration. Muscle C18:0 dihydroceramide was correlated to markers of muscle inflammation. | |
| Bergman et al. [155] | Serum from 14 obese sedentary controls, 15 individuals with T2DM, and 15 endurance-trained athletes. | Basal C18:0, C20:0, and C24:1 ceramide and C18:0 and total dihydroceramide were higher in T2DM and, along with C16:0 ceramide and C18:0 sphingomyelin, correlated positively with insulin resistance. | |
| Perreault et al. [156] | Muscle biopsies from 15 lean individuals, 16 endurance-trained athletes, and 12 obese men and women with T2DM and 15 without T2DM. | Sarcolemmal sphingomyelins were inversely correlated to insulin sensitivity, with the strongest relationships found for the C18:1, C18:0, and C18:2 species. | |
| Kasumov et al. [157] | Plasma from 24 adults with obesity and normal glucose tolerance (NGT, n = 14) or diabetes (n = 10). | Plasma ceramides were similar for the subjects with obesity and NGT and the subjects with diabetes. Exercise reduced body weight and adiposity and increased peripheral insulin sensitivity in both groups. Reduced plasma levels of C14:0, C16:0, C18:1, and C24:0 ceramide in all subjects following the intervention. | |
| Warshauer et al. [158] | Plasma from 37 subjects with metabolic syndrome receiving pioglitazone or placebo. | Decreased plasma levels of C18:0, C20:0, C24:1, dihydroceramide C18:0, dihydroceramide C24:1, lactosylceramide C16:0 and the hexosylceramides C16:0, C18:0, C22:0 and C24:1 in patients in treatment with pioglitazone compared to placebo. | |
| Hanamatsu et al. [159] | Serum from 12 obese participants and 11 controls. | C18:0 and C24:0 levels were significantly higher in the obese individuals. C20:0 and C22:0 levels tended to be higher in the obese group than in the control group. SM C18:0, C20:0, C22:0 and C24:0 correlated with the parameters for obesity, insulin resistance, liver function and lipid metabolism, respectively. | |
| Mamtani et al. [160] | Plasma of 1208 Mexican Americans from 42 extended families. | Dihydroceramide species 18:0, 20:0, 22:0, and 24:1 were genetically associated with Waist circumference (WC). Two sphingomyelin species (31:1 and 41:1) were also related with WC. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Pietro, P.; Izzo, C.; Abate, A.C.; Iesu, P.; Rusciano, M.R.; Venturini, E.; Visco, V.; Sommella, E.; Ciccarelli, M.; Carrizzo, A.; et al. The Dark Side of Sphingolipids: Searching for Potential Cardiovascular Biomarkers. Biomolecules 2023, 13, 168. https://doi.org/10.3390/biom13010168
Di Pietro P, Izzo C, Abate AC, Iesu P, Rusciano MR, Venturini E, Visco V, Sommella E, Ciccarelli M, Carrizzo A, et al. The Dark Side of Sphingolipids: Searching for Potential Cardiovascular Biomarkers. Biomolecules. 2023; 13(1):168. https://doi.org/10.3390/biom13010168
Chicago/Turabian StyleDi Pietro, Paola, Carmine Izzo, Angela Carmelita Abate, Paola Iesu, Maria Rosaria Rusciano, Eleonora Venturini, Valeria Visco, Eduardo Sommella, Michele Ciccarelli, Albino Carrizzo, and et al. 2023. "The Dark Side of Sphingolipids: Searching for Potential Cardiovascular Biomarkers" Biomolecules 13, no. 1: 168. https://doi.org/10.3390/biom13010168