
The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used


Abstract
:1. Introduction
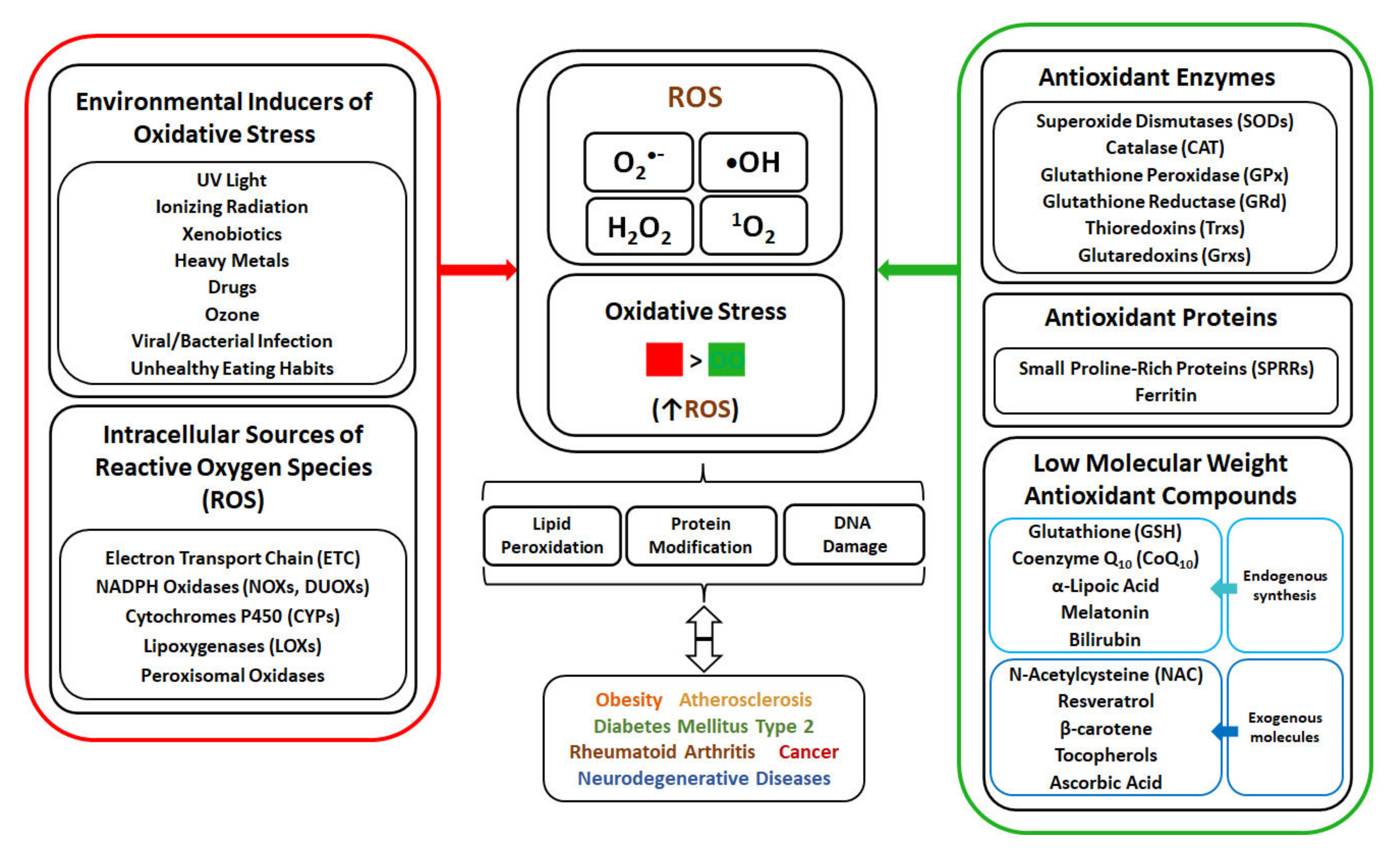
2. Environmental Factors as Inducers of Oxidative Stress
3. Metabolic Pathways as Sources of ROS Generation in Cells
4. Effect of ROS on Cellular Macromolecules
5. Oxidative Stress and Diseases Development
6. Anti-Oxidant Mechanisms as a Protection against Oxidative-Stress
6.1. Superoxide Dismutase (SOD) and Catalase (CAT)
6.2. Glutathione (GSH), Glutathione Peroxidase (GPx) and Glutathione Reductase (GRd)
6.3. Thioredoxins
6.4. Other Antioxidant Proteins
6.5. Antioxidant Low Molecular Weight Molecules
7. Vitamins as Antioxidants
7.1. Structural and Antioxidant Characteristics of β-Carotene, Tocopherols and Ascorbate
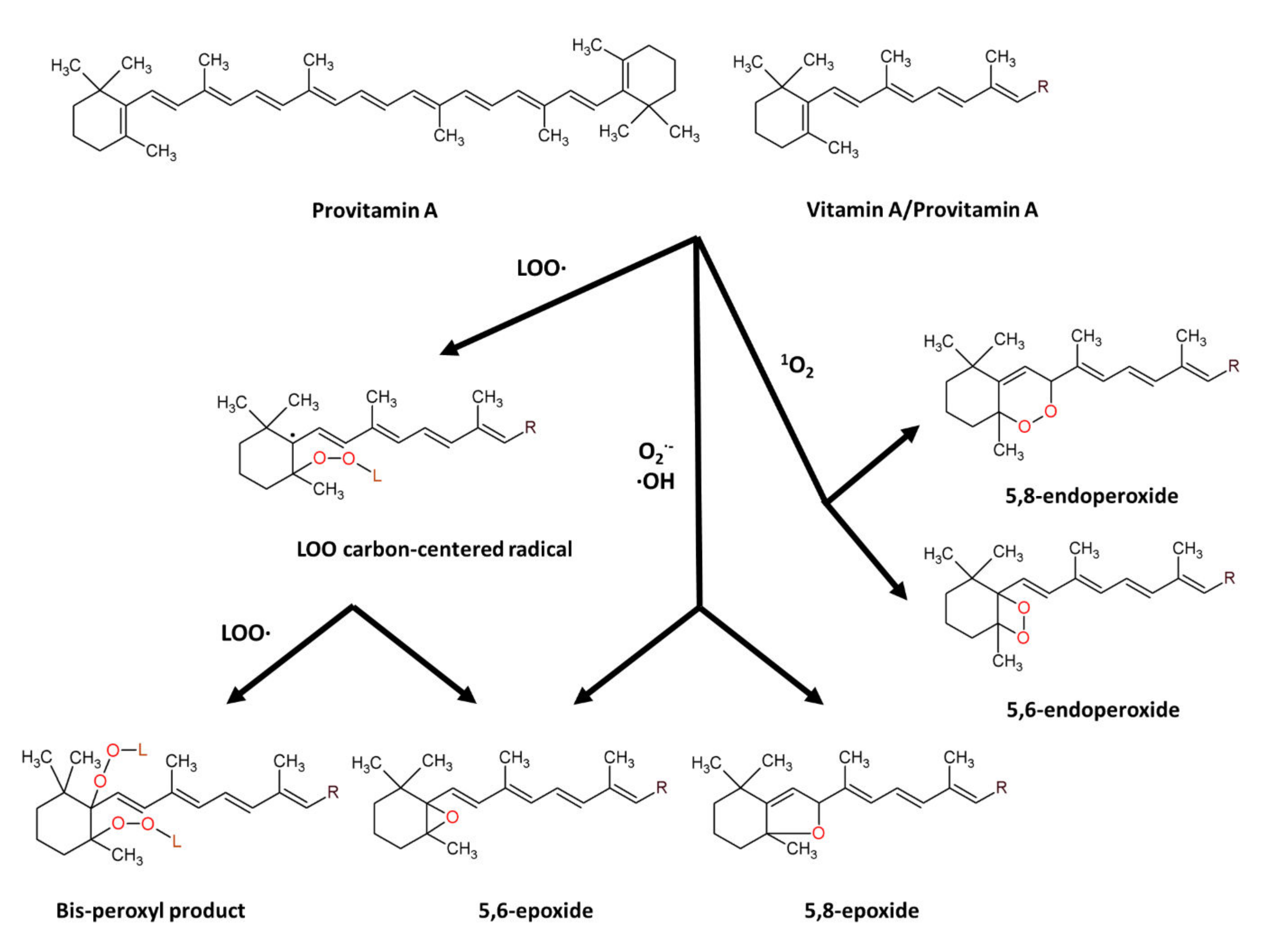
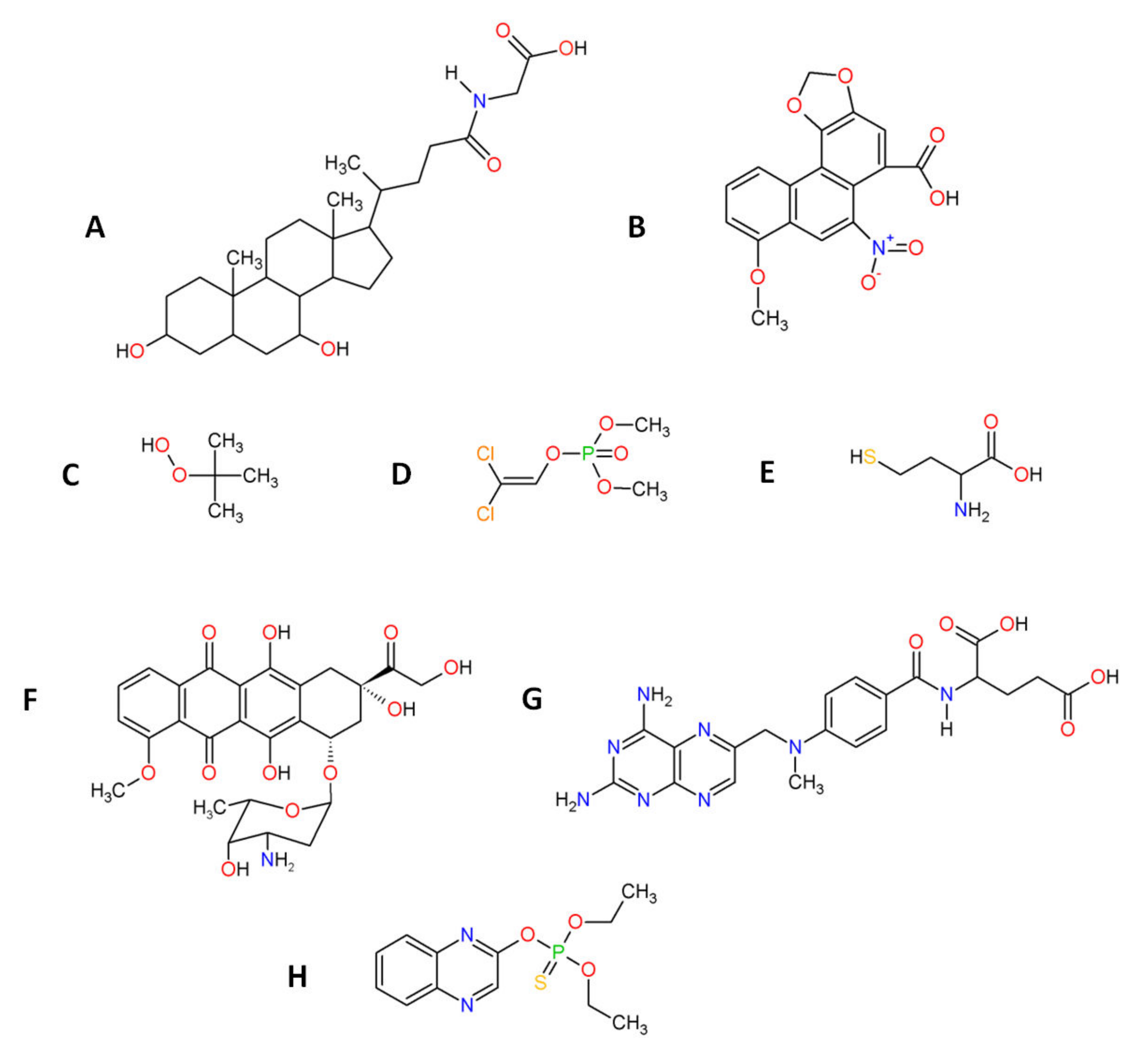
7.1.1. β-Carotene: Structure and Anti-Oxidant Property
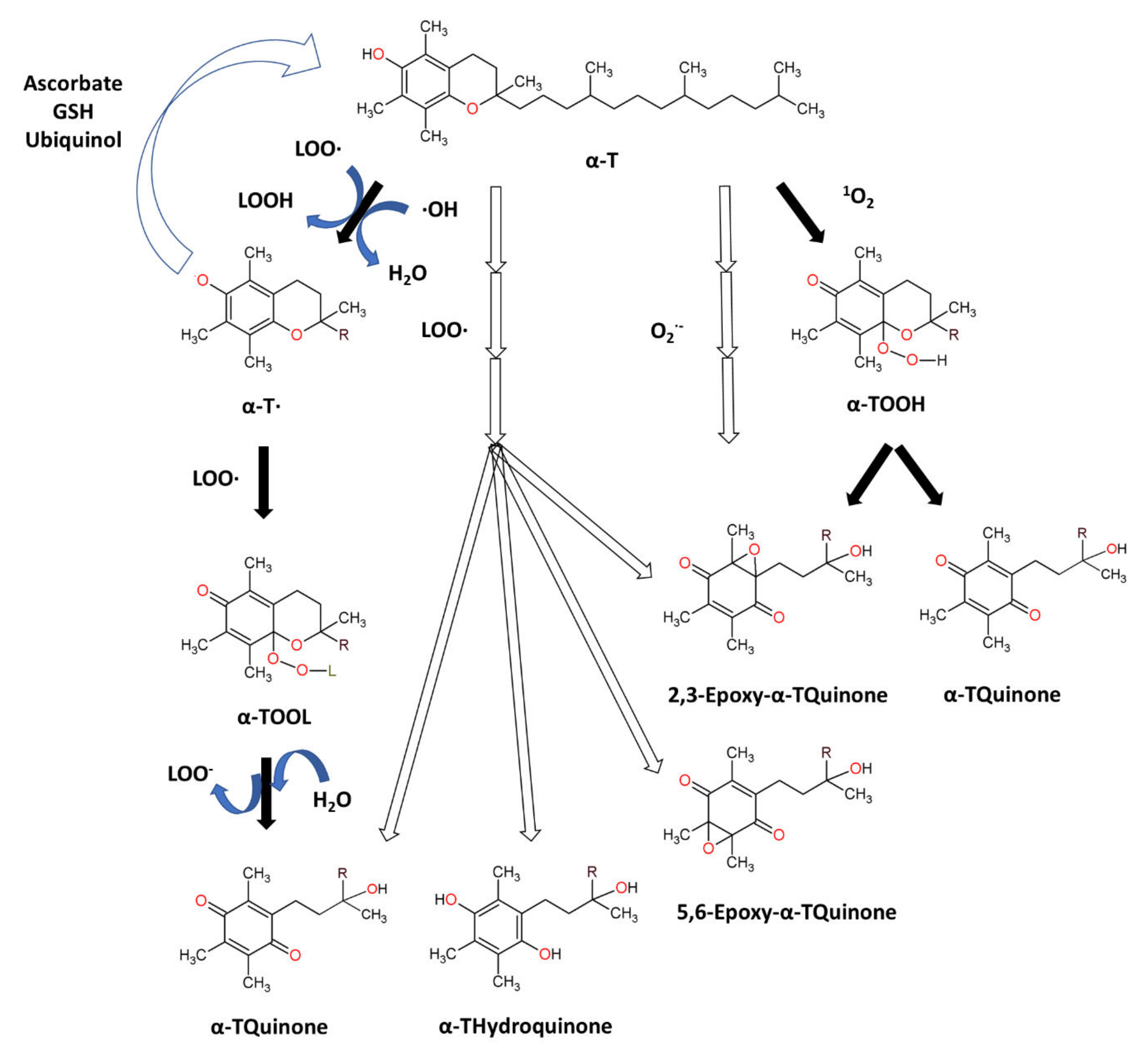
7.1.2. Tocopherols: Structure and Anti-Oxidant Property
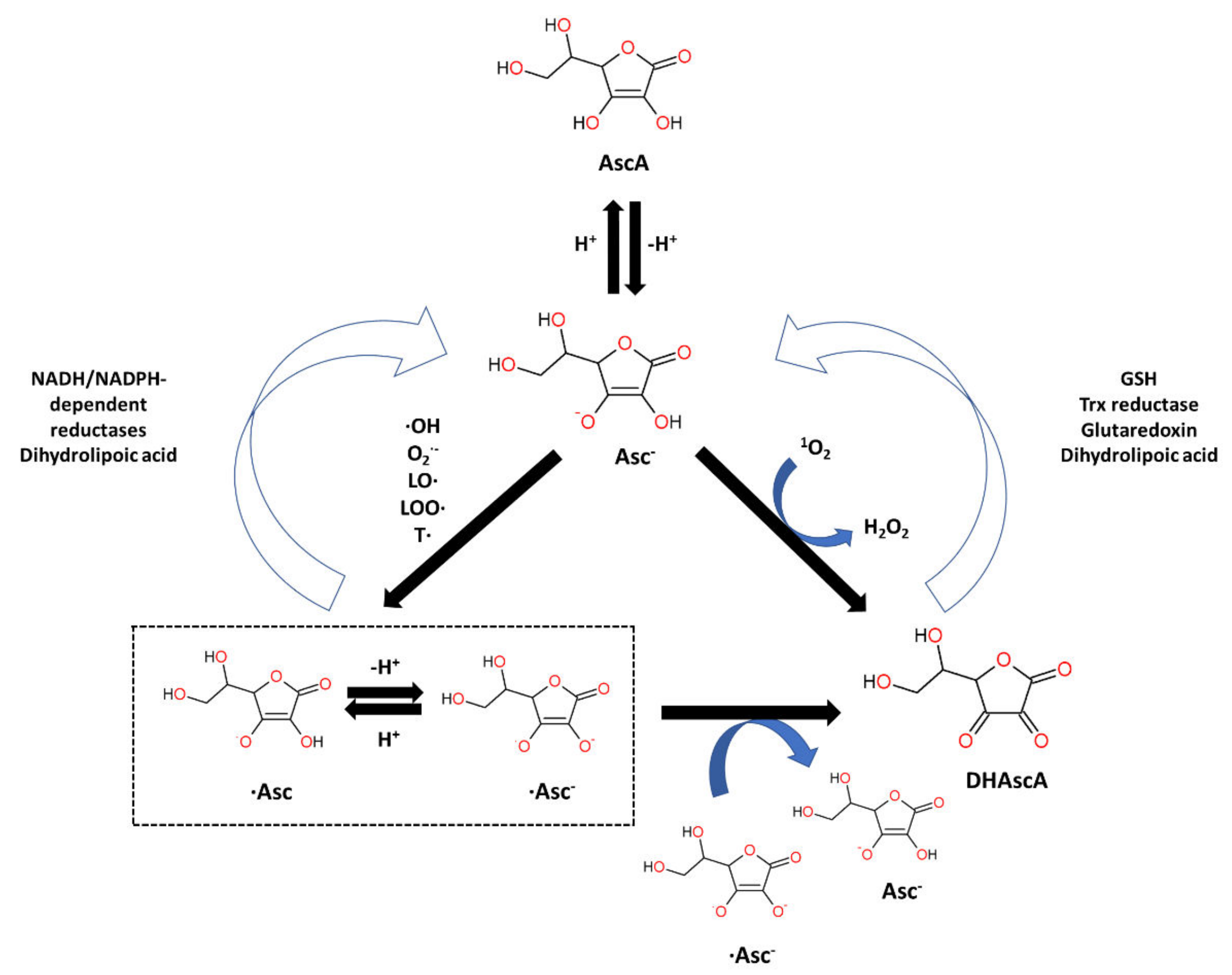
7.1.3. Ascorbic Acid: Structure and Anti-Oxidant Property
7.2. In Vivo and In Vitro Antioxidant Characteristics of β-Carotene, Tocopherols and Ascorbate
7.2.1. Oxidative Stress Inducers Used during In Vitro and In Vivo Studies
7.2.2. β-Carotene: Antioxidant Activity In Vitro
7.2.3. Tocopherols: Antioxidant Activity In Vitro
7.2.4. Ascorbic Acid: Antioxidant Activity In Vitro
7.2.5. β-Carotene: Antioxidant Activity In Vivo
7.2.6. Tocopherols: Antioxidant Activity In Vivo
7.2.7. Ascorbic Acid: Antioxidant Activity In Vivo
7.2.8. Antioxidant Characteristics of β-Carotene, Tocopherols and Ascorbate Based on Populational/Clinical Human Studies
7.2.9. Antioxidant Characteristics of β-Carotene, Tocopherols and Ascorbate: A Summary
7.3. Measurement of the Oxidative Stress in Cells and Tissues
7.3.1. Measurement of ROS Level: ESR Spectroscopy
7.3.2. Measurement of ROS Level: Fluorescent Method (DCFH-DA Probe Assay)
7.3.3. Measurement of ROS Level: Other Methods
7.3.4. Measurement of Lipid Peroxidation Products
Measurement of Lipid Peroxidation: MDA, 4-HNE
Measurement of Lipid Peroxidation: F2-Isoprostanes (F2-IsoPs)
Measurement of Lipid Peroxidation: Lipid Hydroperoxides
7.3.5. Measurement of Protein Oxidation: Protein Carbonyls
7.3.6. Measurement of DNA Oxidation: 8-OHdG
7.3.7. Analysis of Oxidative Stress and Vitamin Effect by Raman Spectroscopy and Imaging
7.3.8. Measurement of the Oxidative Stress: A Summary
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Pędziwiatr, P.; Mikołajczyk, F.; Zawadzki, D.; Mikołajczyk, K.; Bedka, A. Decomposition of Hydrogen Peroxide-Kinetics and Review of Chosen Catalysts. Acta Innov. 2018, 26, 45–52. [Google Scholar] [CrossRef]
- Haider, M.S.; Jaskani, M.J.; Fang, J. Overproduction of ROS: Underlying Molecular Mechanism of Scavenging and Redox Signaling. In Biocontrol Agents and Secondary Metabolites; Elsevier: Amsterdam, The Netherlands, 2021; pp. 347–382. ISBN 9780128229194. [Google Scholar]
- Kaur, H.; Hippargi, G.; Pophali, G.R.; Bansiwal, A.K. Treatment Methods for Removal of Pharmaceuticals and Personal Care Products from Domestic Wastewater. In Pharmaceuticals and Personal Care Products: Waste Management and Treatment Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 129–150. ISBN 9780128161890. [Google Scholar]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial Formation of Reactive Oxygen Species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, J.-W. Cellular Defense against Singlet Oxygen-Induced Oxidative Damage by Cytosolic NADP+-Dependent Isocitrate Dehydrogenase. Free Radic. Res. 2003, 37, 309–316. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- De Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet Light Induced Generation of Reactive Oxygen Species. Adv. Exp. Med. Biol. 2017, 996, 15–23. [Google Scholar] [CrossRef]
- Brand, R.M.; Wipf, P.; Durham, A.; Epperly, M.W.; Greenberger, J.S.; Falo, L.D., Jr. Targeting Mitochondrial Oxidative Stress to Mitigate UV-Induced Skin Damage. Front. Pharmacol. 2018, 9, 920. [Google Scholar] [CrossRef]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of Ionizing Radiation on Biological Molecules—Mechanisms of Damage and Emerging Methods of Detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef]
- Mu, H.; Sun, J.; Li, L.; Yin, J.; Hu, N.; Zhao, W.; Ding, D.; Yi, L. Ionizing Radiation Exposure: Hazards, Prevention, and Biomarker Screening. Environ. Sci. Pollut. Res. Int. 2018, 25, 15294–15306. [Google Scholar] [CrossRef]
- Nuszkiewicz, J.; Woźniak, A.; Szewczyk-Golec, K. Ionizing Radiation as a Source of Oxidative Stress—The Protective Role of Melatonin and Vitamin D. Int. J. Mol. Sci. 2020, 21, 5804. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing Radiation-Induced Metabolic Oxidative Stress and Prolonged Cell Injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Huang, C.; Lv, Y.-S.; Ma, S.-X.; Guo, Y.; Zeng, E.Y. Polycyclic Aromatic Hydrocarbon Exposure, Oxidative Potential in Dust, and Their Relationships to Oxidative Stress in Human Body: A Case Study in the Indoor Environment of Guangzhou, South China. Environ. Int. 2021, 149, 106405. [Google Scholar] [CrossRef]
- Henkler, F.; Brinkmann, J.; Luch, A. The Role of Oxidative Stress in Carcinogenesis Induced by Metals and Xenobiotics. Cancers 2010, 2, 376–396. [Google Scholar] [CrossRef]
- Klotz, L.-O.; Steinbrenner, H. Cellular Adaptation to Xenobiotics: Interplay between Xenosensors, Reactive Oxygen Species and FOXO Transcription Factors. Redox Biol. 2017, 13, 646–654. [Google Scholar] [CrossRef]
- Jabłońska-Trypuć, A.; Wołejko, E.; Wydro, U.; Butarewicz, A. The Impact of Pesticides on Oxidative Stress Level in Human Organism and Their Activity as an Endocrine Disruptor. J. Environ. Sci. Health B 2017, 52, 483–494. [Google Scholar] [CrossRef]
- Azeh Engwa, G.; Udoka Ferdinand, P.; Nweke Nwalo, F.; Unachukwu, M.N. Mechanism and Health Effects of Heavy Metal Toxicity in Humans. In Poisoning in the Modern World-New Tricks for an Old Dog? IntechOpen: London, UK, 2019; ISBN 9781838807856. [Google Scholar]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic Metals and Oxidative Stress Part I: Mechanisms Involved in Metal-Induced Oxidative Damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Jomova, K.; Jenisova, Z.; Feszterova, M.; Baros, S.; Liska, J.; Hudecova, D.; Rhodes, C.J.; Valko, M. Arsenic: Toxicity, Oxidative Stress and Human Disease: Toxicity of Arsenic. J. Appl. Toxicol. 2011, 31, 95–107. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Advances in Metal-Induced Oxidative Stress and Human Disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-Induced Oxidative Stress and Toxicity. J. Toxicol. 2012, 2012, 645460. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Mukherjea, D.; Dhukhwa, A.; Rybak, L.P. Oxidative Stress and Inflammation Caused by Cisplatin Ototoxicity. Antioxidants 2021, 10, 1919. [Google Scholar] [CrossRef] [PubMed]
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Ali, B.A.; Zulli, A.; Apostolopoulos, V. Mechanisms of Cisplatin-Induced Acute Kidney Injury: Pathological Mechanisms, Pharmacological Interventions, and Genetic Mitigations. Cancers 2021, 13, 1572. [Google Scholar] [CrossRef]
- Kamceva, G.; Arsova-Sarafinovska, Z.; Ruskovska, T.; Zdravkovska, M.; Kamceva-Panova, L.; Stikova, E. Cigarette Smoking and Oxidative Stress in Patients with Coronary Artery Disease. Open Access Maced. J. Med. Sci. 2016, 4, 636–640. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K. Tobacco Smoke: Involvement of Reactive Oxygen Species and Stable Free Radicals in Mechanisms of Oxidative Damage, Carcinogenesis and Synergistic Effects with Other Respirable Particles. Int. J. Environ. Res. Public Health 2009, 6, 445–462. [Google Scholar] [CrossRef]
- Faux, S.P.; Tai, T.; Thorne, D.; Xu, Y.; Breheny, D.; Gaca, M. The Role of Oxidative Stress in the Biological Responses of Lung Epithelial Cells to Cigarette Smoke. Biomarkers 2009, 14 (Suppl. 1), 90–96. [Google Scholar] [CrossRef]
- Isik, B.; Ceylan, A.; Isik, R. Oxidative Stress in Smokers and Non-Smokers. Inhal. Toxicol. 2007, 19, 767–769. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Oxidative Stress, the Paradigm of Ozone Toxicity in Plants and Animals. Water Air Soil Pollut. 2007, 187, 285–301. [Google Scholar] [CrossRef]
- Nery-Flores, S.D.; Mendoza-Magaña, M.L.; Ramírez-Herrera, M.A.; Ramírez-Vázquez, J.d.J.; Romero-Prado, M.M.d.J.; Cortez-Álvarez, C.R.; Ramírez-Mendoza, A.A. Curcumin Exerted Neuroprotection against Ozone-Induced Oxidative Damage and Decreased NF-ΚB Activation in Rat Hippocampus and Serum Levels of Inflammatory Cytokines. Oxid. Med. Cell. Longev. 2018, 2018, 9620684. [Google Scholar] [CrossRef]
- Tricarico, G.; Travagli, V. The Relationship between Ozone and Human Blood in the Course of a Well-Controlled, Mild, and Transitory Oxidative Eustress. Antioxidants 2021, 10, 1946. [Google Scholar] [CrossRef]
- Rivas-Arancibia, S.; Guevara-Guzmán, R.; López-Vidal, Y.; Rodríguez-Martínez, E.; Zanardo-Gomes, M.; Angoa-Pérez, M.; Raisman-Vozari, R. Oxidative Stress Caused by Ozone Exposure Induces Loss of Brain Repair in the Hippocampus of Adult Rats. Toxicol. Sci. 2010, 113, 187–197. [Google Scholar] [CrossRef]
- Schwarz, K.B. Oxidative Stress during Viral Infection: A Review. Free Radic. Biol. Med. 1996, 21, 641–649. [Google Scholar] [CrossRef]
- Lee, C. Therapeutic Modulation of Virus-Induced Oxidative Stress via the Nrf2-Dependent Antioxidative Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 6208067. [Google Scholar] [CrossRef]
- Butcher, L.D.; den Hartog, G.; Ernst, P.B.; Crowe, S.E. Oxidative Stress Resulting from Helicobacter Pylori Infection Contributes to Gastric Carcinogenesis. Cell Mol. Gastroenterol. Hepatol. 2017, 3, 316–322. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P. Nutrients and Oxidative Stress: Friend or Foe? Oxid. Med. Cell. Longev. 2018, 2018, 9719584. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity: Implications for Metabolic Syndrome, Diabetes, Hypertension, Dyslipidemia, Atherosclerosis, and Cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef]
- Jiang, S.; Liu, H.; Li, C. Dietary Regulation of Oxidative Stress in Chronic Metabolic Diseases. Foods 2021, 10, 1854. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial Electron Transport Chain: Oxidative Phosphorylation, Oxidant Production, and Methods of Measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Zhao, R.-Z.; Jiang, S.; Zhang, L.; Yu, Z.-B. Mitochondrial Electron Transport Chain, ROS Generation and Uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef]
- Raimondi, V.; Ciccarese, F.; Ciminale, V. Oncogenic Pathways and the Electron Transport Chain: A DangeROS Liaison. Br. J. Cancer 2020, 122, 168–181. [Google Scholar] [CrossRef]
- Tarafdar, A.; Pula, G. The Role of NADPH Oxidases and Oxidative Stress in Neurodegenerative Disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef]
- Paravicini, T.M.; Touyz, R.M. NADPH Oxidases, Reactive Oxygen Species, and Hypertension: Clinical Implications and Therapeutic Possibilities. Diabetes Care 2008, 31, S170–S180. [Google Scholar] [CrossRef]
- Guo, S.; Chen, X. The Human Nox4: Gene, Structure, Physiological Function and Pathological Significance. J. Drug Target. 2015, 23, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox Family NADPH Oxidases: Molecular Mechanisms of Activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Anagnostopoulou, A.; Rios, F.; Montezano, A.C.; Camargo, L.L. NOX5: Molecular Biology and Pathophysiology. Exp. Physiol. 2019, 104, 605–616. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Pushkaran, S.; Konstantinidis, D.G.; Koochaki, S.; Malik, P.; Mohandas, N.; Zheng, Y.; Joiner, C.H.; Kalfa, T.A. Erythrocyte NADPH Oxidase Activity Modulated by Rac GTPases, PKC, and Plasma Cytokines Contributes to Oxidative Stress in Sickle Cell Disease. Blood 2013, 121, 2099–2107. [Google Scholar] [CrossRef]
- Violi, F.; Carnevale, R.; Loffredo, L.; Pignatelli, P.; Gallin, J.I. NADPH Oxidase-2 and Atherothrombosis: Insight from Chronic Granulomatous Disease. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 218–225. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Härtlein, I.; Smith, S.M.E.; Fieschi, F. NADPH Oxidases (NOX): An Overview from Discovery, Molecular Mechanisms to Physiology and Pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Magnani, F.; Mattevi, A. Structure and Mechanisms of ROS Generation by NADPH Oxidases. Curr. Opin. Struct. Biol. 2019, 59, 91–97. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Pircalabioru, G.G.; Bezirtzoglou, E. The Role of Cytochromes P450 in Infection. Front. Immunol. 2018, 9, 89. [Google Scholar] [CrossRef]
- Hrycay, E.G.; Bandiera, S.M. Involvement of Cytochrome P450 in Reactive Oxygen Species Formation and Cancer. Adv. Pharmacol. 2015, 74, 35–84. [Google Scholar] [CrossRef]
- Veith, A.; Moorthy, B. Role of Cytochrome P450s in the Generation and Metabolism of Reactive Oxygen Species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T. The Role of Lipoxygenases in Pathophysiology; New Insights and Future Perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef]
- Qian, S.; Guo, L.; Tang, Q. Lipids in the Transcriptional Regulation of Adipocyte Differentiation and Metabolism. In Lipid Signaling and Metabolism; Elsevier: Amsterdam, The Netherlands, 2020; pp. 81–98. ISBN 9780128194041. [Google Scholar]
- Cho, K.-J.; Seo, J.-M.; Kim, J.-H. Bioactive Lipoxygenase Metabolites Stimulation of NADPH Oxidases and Reactive Oxygen Species. Mol. Cells 2011, 32, 1–5. [Google Scholar] [CrossRef]
- Suzuki, H.; Kayama, Y.; Sakamoto, M.; Iuchi, H.; Shimizu, I.; Yoshino, T.; Katoh, D.; Nagoshi, T.; Tojo, K.; Minamino, T.; et al. Arachidonate 12/15-Lipoxygenase-Induced Inflammation and Oxidative Stress Are Involved in the Development of Diabetic Cardiomyopathy. Diabetes 2015, 64, 618–630. [Google Scholar] [CrossRef]
- Chornyi, S.; IJlst, L.; van Roermund, C.W.T.; Wanders, R.J.A.; Waterham, H.R. Peroxisomal Metabolite and Cofactor Transport in Humans. Front. Cell Dev. Biol. 2020, 8, 613892. [Google Scholar] [CrossRef]
- Wanders, R.J.A.; Vaz, F.M.; Ferdinandusse, S.; Kemp, S.; Ebberink, M.S.; Waterham, H.R. Laboratory Diagnosis of Peroxisomal Disorders in the -Omics Era and the Continued Importance of Biomarkers and Biochemical Studies. J. Inborn Errors Metab. Screen. 2018, 6, 232640981881028. [Google Scholar] [CrossRef]
- Schrader, M.; Fahimi, H.D. Peroxisomes and Oxidative Stress. Biochim. Biophys. Acta 2006, 1763, 1755–1766. [Google Scholar] [CrossRef]
- Aziz, N.; Jamil, R.T. Biochemistry, Xanthine Oxidase; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine Oxidoreductase: One Enzyme for Multiple Physiological Tasks. Redox Biol. 2021, 41, 101882. [Google Scholar] [CrossRef]
- Ferdinandusse, S.; Denis, S.; van Roermund, C.W.T.; Preece, M.A.; Koster, J.; Ebberink, M.S.; Waterham, H.R.; Wanders, R.J.A. A Novel Case of ACOX2 Deficiency Leads to Recognition of a Third Human Peroxisomal Acyl-CoA Oxidase. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Dansen, T.B.; Wirtz, K.W. The Peroxisome in Oxidative Stress. IUBMB Life 2001, 51, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Oaxaca-Castillo, D.; Andreoletti, P.; Vluggens, A.; Yu, S.; van Veldhoven, P.P.; Reddy, J.K.; Cherkaoui-Malki, M. Biochemical Characterization of Two Functional Human Liver Acyl-CoA Oxidase Isoforms 1a and 1b Encoded by a Single Gene. Biochem. Biophys. Res. Commun. 2007, 360, 314–319. [Google Scholar] [CrossRef]
- Grimsrud, P.A.; Xie, H.; Griffin, T.J.; Bernlohr, D.A. Oxidative Stress and Covalent Modification of Protein with Bioactive Aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef]
- Goto, S.; Radak, Z. Implications of Oxidative Damage to Proteins and DNA in Aging and Its Intervention by Caloric Restriction and Exercise. J. Sport Health Sci. 2013, 2, 75–80. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free Radicals, Natural Antioxidants, and Their Reaction Mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Aquila, G.; Marracino, L.; Martino, V.; Calabria, D.; Campo, G.; Caliceti, C.; Rizzo, P. The Use of Nutraceuticals to Counteract Atherosclerosis: The Role of the Notch Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 5470470. [Google Scholar] [CrossRef]
- Zmysłowski, A.; Szterk, A. Current Knowledge on the Mechanism of Atherosclerosis and Pro-Atherosclerotic Properties of Oxysterols. Lipids Health Dis. 2017, 16, 188. [Google Scholar] [CrossRef]
- Khatana, C.; Saini, N.K.; Chakrabarti, S.; Saini, V.; Sharma, A.; Saini, R.V.; Saini, A.K. Mechanistic Insights into the Oxidized Low-Density Lipoprotein-Induced Atherosclerosis. Oxid. Med. Cell Longev. 2020, 2020, 5245308. [Google Scholar] [CrossRef]
- Mathur, P.; Ding, Z.; Saldeen, T.; Mehta, J.L. Tocopherols in the Prevention and Treatment of Atherosclerosis and Related Cardiovascular Disease: Tocopherols in Atherosclerosis. Clin. Cardiol. 2015, 38, 570–576. [Google Scholar] [CrossRef]
- Babaniamansour, P.; Mohammadi, M.; Babaniamansour, S.; Aliniagerdroudbari, E. The Relation between Atherosclerosis Plaque Composition and Plaque Rupture. J. Med. Signals Sens. 2020, 10, 267–273. [Google Scholar] [CrossRef]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes Mellitus and Oxidative Stress—A Concise Review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Abad-Jiménez, Z.; Martínez de Marañón, A.; Iannantuoni, F.; Escribano-López, I.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef] [PubMed]
- Quiñonez-Flores, C.M.; González-Chávez, S.A.; Del Río Nájera, D.; Pacheco-Tena, C. Oxidative Stress Relevance in the Pathogenesis of the Rheumatoid Arthritis: A Systematic Review. BioMed. Res. Int. 2016, 2016, 6097417. [Google Scholar] [CrossRef]
- Cooper, G.M. The Development and Causes of Cancer. In The Cell: A Molecular Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000; ISBN 100878931066. [Google Scholar]
- Hassanpour, S.H.; Dehghani, M. Review of Cancer from Perspective of Molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Kamal, N.; Ilowefah, M.A.; Hilles, A.R.; Anua, N.A.; Awin, T.; Alshwyeh, H.A.; Aldosary, S.K.; Jambocus, N.G.S.; Alosaimi, A.A.; Rahman, A.; et al. Genesis and Mechanism of Some Cancer Types and an Overview on the Role of Diet and Nutrition in Cancer Prevention. Molecules 2022, 27, 1794. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Poetsch, A.R. The Genomics of Oxidative DNA Damage, Repair, and Resulting Mutagenesis. Comput. Struct. Biotechnol. J. 2020, 18, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative Stress and Oxidative Damage in Carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Bridge, G.; Rashid, S.; Martin, S.A. DNA Mismatch Repair and Oxidative DNA Damage: Implications for Cancer Biology and Treatment. Cancers 2014, 6, 1597–1614. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid. Med. Cell. Longev. 2017, 2017, 2525967. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative Stress in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef]
- Rosa, A.C.; Corsi, D.; Cavi, N.; Bruni, N.; Dosio, F. Superoxide Dismutase Administration: A Review of Proposed Human Uses. Molecules 2021, 26, 1844. [Google Scholar] [CrossRef]
- Trist, B.G.; Hilton, J.B.; Hare, D.J.; Crouch, P.J.; Double, K.L. Superoxide Dismutase 1 in Health and Disease: How a Frontline Antioxidant Becomes Neurotoxic. Angew. Chem. Int. Ed. Engl. 2021, 60, 9215–9246. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef]
- Sarıkaya, E.; Doğan, S. Glutathione Peroxidase in Health and Diseases. In Glutathione System and Oxidative Stress in Health and Disease; IntechOpen: London, UK, 2020; ISBN 9781838801250. [Google Scholar]
- Pizzorno, J. Glutathione! Integ. Med. 2014, 13, 8–12. [Google Scholar]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione Peroxidase-1 in Health and Disease: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid. Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef]
- Marín-García, J. Oxidative Stress and Cell Death in Cardiovascular Disease. In Post-Genomic Cardiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 471–498. ISBN 9780124045996. [Google Scholar]
- Deponte, M. Glutathione Catalysis and the Reaction Mechanisms of Glutathione-Dependent Enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef]
- Ren, X.; Zou, L.; Zhang, X.; Branco, V.; Wang, J.; Carvalho, C.; Holmgren, A.; Lu, J. Redox Signaling Mediated by Thioredoxin and Glutathione Systems in the Central Nervous System. Antioxid. Redox Signal. 2017, 27, 989–1010. [Google Scholar] [CrossRef]
- Tabet, F.; Touyz, R.M. Reactive Oxygen Species, Oxidative Stress, and Vascular Biology in Hypertension. In Comprehensive Hypertension; Elsevier: Amsterdam, The Netherlands, 2007; pp. 337–347. ISBN 9780323039611. [Google Scholar]
- Becker, K.; Gromer, S.; Schirmer, R.H.; Müller, S. Thioredoxin Reductase as a Pathophysiological Factor and Drug Target: Thioredoxin Reductase in Medicine and Parasitology. Eur. J. Biochem. 2000, 267, 6118–6125. [Google Scholar] [CrossRef]
- Vlamis-Gardikas, A.; Holmgren, A. Thioredoxin and Glutaredoxin Isoforms. Methods Enzymol. 2002, 347, 286–296. [Google Scholar] [CrossRef]
- Lundberg, M.; Johansson, C.; Chandra, J.; Enoksson, M.; Jacobsson, G.; Ljung, J.; Johansson, M.; Holmgren, A. Cloning and Expression of a Novel Human Glutaredoxin (Grx2) with Mitochondrial and Nuclear Isoforms. J. Biol. Chem. 2001, 276, 26269–26275. [Google Scholar] [CrossRef]
- Évora, A.S.; Adams, M.J.; Johnson, S.A.; Zhang, Z. Corneocytes: Relationship between Structural and Biomechanical Properties. Ski. Pharmacol. Physiol. 2021, 34, 146–161. [Google Scholar] [CrossRef]
- Nozaki, I.; Lunz, J.G., 3rd; Specht, S.; Stolz, D.B.; Taguchi, K.; Subbotin, V.M.; Murase, N.; Demetris, A.J. Small Proline-Rich Proteins 2 Are Noncoordinately Upregulated by IL-6/STAT3 Signaling after Bile Duct Ligation. Lab. Investig. 2005, 85, 109–123. [Google Scholar] [CrossRef]
- Ishitsuka, Y.; Ogawa, T.; Roop, D. The KEAP1/NRF2 Signaling Pathway in Keratinization. Antioxidants 2020, 9, 751. [Google Scholar] [CrossRef]
- Vermeij, W.P.; Alia, A.; Backendorf, C. ROS Quenching Potential of the Epidermal Cornified Cell Envelope. J. Investig. Dermatol. 2011, 131, 1435–1441. [Google Scholar] [CrossRef]
- Schäfer, M.; Werner, S. The Cornified Envelope: A First Line of Defense against Reactive Oxygen Species. J. Investig. Dermatol. 2011, 131, 1409–1411. [Google Scholar] [CrossRef]
- MacKenzie, E.L.; Iwasaki, K.; Tsuji, Y. Intracellular Iron Transport and Storage: From Molecular Mechanisms to Health Implications. Antioxid. Redox Signal. 2008, 10, 997–1030. [Google Scholar] [CrossRef]
- Katsarou, M.-S.; Papasavva, M.; Latsi, R.; Drakoulis, N. Hemochromatosis: Hereditary Hemochromatosis and HFE Gene. Vitam. Horm. 2019, 110, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M. Antioxidant Properties of Wheat Bran against Oxidative Stress. In Wheat and Rice in Disease Prevention and Health; Elsevier: Amsterdam, The Netherlands, 2014; pp. 181–199. ISBN 9780124017160. [Google Scholar]
- Imam, M.U.; Zhang, S.; Ma, J.; Wang, H.; Wang, F. Antioxidants Mediate Both Iron Homeostasis and Oxidative Stress. Nutrients 2017, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Romeu, M.; Aranda, N.; Giralt, M.; Ribot, B.; Nogues, M.R.; Arija, V. Diet, Iron Biomarkers and Oxidative Stress in a Representative Sample of Mediterranean Population. Nutr. J. 2013, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Orino, K.; Lehman, L.; Tsuji, Y.; Ayaki, H.; Torti, S.V.; Torti, F.M. Ferritin and the Response to Oxidative Stress. Biochem. J. 2001, 357, 241. [Google Scholar] [CrossRef] [PubMed]
- Marappan, G. Coenzyme Q10: Regulators of Mitochondria and Beyond. In Apolipoproteins, Triglycerides and Cholesterol; IntechOpen: London, UK, 2020; ISBN 9781839625190. [Google Scholar]
- Acosta, M.J.; Vazquez Fonseca, L.; Desbats, M.A.; Cerqua, C.; Zordan, R.; Trevisson, E.; Salviati, L. Coenzyme Q Biosynthesis in Health and Disease. Biochim. Biophys. Acta 2016, 1857, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Yubero-Serrano, E.M.; Garcia-Rios, A.; Delgado-Lista, J.; Pérez-Martinez, P.; Camargo, A.; Perez-Jimenez, F.; Lopez-Miranda, J. Coenzyme Q10 as an Antioxidant in the Elderly. In Aging; Elsevier: Amsterdam, The Netherlands, 2014; pp. 109–117. ISBN 9780124059337. [Google Scholar]
- Sangsefidi, Z.S.; Yaghoubi, F.; Hajiahmadi, S.; Hosseinzadeh, M. The Effect of Coenzyme Q10 Supplementation on Oxidative Stress: A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials: The Effect of Coenzyme Q10 Supplementation on Oxidative Stress: A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials. Food Sci. Nutr. 2020, 8, 1766–1776. [Google Scholar] [CrossRef]
- Melhuish Beaupre, L.M.; Brown, G.M.; Gonçalves, V.F.; Kennedy, J.L. Melatonin’s Neuroprotective Role in Mitochondria and Its Potential as a Biomarker in Aging, Cognition and Psychiatric Disorders. Transl. Psychiatry 2021, 11, 339. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Bernatoniene, J. Molecular Mechanisms of Melatonin-Mediated Cell Protection and Signaling in Health and Disease. Pharmaceutics 2021, 13, 129. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Mayo, J.C.; Sainz, R.M.; Leon, J.; Czarnocki, Z. Melatonin as an Antioxidant: Biochemical Mechanisms and Pathophysiological Implications in Humans. Acta Biochim. Pol. 2003, 50, 1129–1146. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Rosales-Corral, S.; Manucha, W.; Chuffa, L.G.d.A.; Zuccari, D.A.P.d.C. Melatonin and Pathological Cell Interactions: Mitochondrial Glucose Processing in Cancer Cells. Int. J. Mol. Sci. 2021, 22, 12494. [Google Scholar] [CrossRef]
- Malhotra, S.; Sawhney, G.; Pandhi, P. The Therapeutic Potential of Melatonin: A Review of the Science. MedGenMed 2004, 6, 46. [Google Scholar]
- Ziberna, L.; Martelanc, M.; Franko, M.; Passamonti, S. Bilirubin Is an Endogenous Antioxidant in Human Vascular Endothelial Cells. Sci. Rep. 2016, 6, 29240. [Google Scholar] [CrossRef]
- Adin, C.A. Bilirubin as a Therapeutic Molecule: Challenges and Opportunities. Antioxidants 2021, 10, 1536. [Google Scholar] [CrossRef]
- Inoguchi, T.; Sonoda, N.; Maeda, Y. Bilirubin as an Important Physiological Modulator of Oxidative Stress and Chronic Inflammation in Metabolic Syndrome and Diabetes: A New Aspect on Old Molecule. Diabetol. Int. 2016, 7, 338–341. [Google Scholar] [CrossRef]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and Glutathione Have Complementary Antioxidant and Cytoprotective Roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef]
- Otero Regino, W.; Velasco, H.; Sandoval, H. The protective role of bilirubin in human beings. Rev. Colomb. Gastroenterol. 2009, 24, 293–301. [Google Scholar]
- DiNicolantonio, J.J.; McCarty, M.F.; O’Keefe, J.H. Antioxidant Bilirubin Works in Multiple Ways to Reduce Risk for Obesity and Its Health Complications. Open Heart 2018, 5, e000914. [Google Scholar] [CrossRef]
- Anthony, R.M.; MacLeay, J.M.; Jewell, D.E.; Brejda, J.J.; Gross, K.L. Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs. Animals 2021, 11, 274. [Google Scholar] [CrossRef]
- Golbidi, S.; Badran, M.; Laher, I. Diabetes and Alpha Lipoic Acid. Front. Pharmacol. 2011, 2, 69. [Google Scholar] [CrossRef]
- Ghibu, S.; Craciun, C.E.; Rusu, R.; Morgovan, C.; Mogosan, C.; Rochette, L.; Gal, A.F.; Dronca, M. Impact of Alpha-Lipoic Acid Chronic Discontinuous Treatment in Cardiometabolic Disorders and Oxidative Stress Induced by Fructose Intake in Rats. Antioxidants 2019, 8, 636. [Google Scholar] [CrossRef]
- Elshazly, S.M.; El-Moselhy, M.A.; Barakat, W. Insights in the Mechanism Underlying the Protective Effect of α-Lipoic Acid against Acetaminophen-Hepatotoxicity. Eur. J. Pharmacol. 2014, 726, 116–123. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-Lipoic Acid as a Dietary Supplement: Molecular Mechanisms and Therapeutic Potential. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 1149–1160. [Google Scholar] [CrossRef]
- Tenório, M.C.d.S.; Graciliano, N.G.; Moura, F.A.; Oliveira, A.C.M.d.; Goulart, M.O.F. N-Acetylcysteine (NAC): Impacts on Human Health. Antioxidants 2021, 10, 967. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an Antioxidant and Disulphide Breaking Agent: The Reasons Why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Magdalena Radomska-Leśniewska, D.; Skopiński, P. N-acetylcysteine as an Anti-oxidant and Anti-inflammatory Drug and its Some Clinical Applications. Centr. Eur. J. Immunol. 2012, 37, 57–66. [Google Scholar]
- Tian, B.; Liu, J. Resveratrol: A Review of Plant Sources, Synthesis, Stability, Modification and Food Application. J. Sci. Food Agric. 2020, 100, 1392–1404. [Google Scholar] [CrossRef]
- Ramírez-Garza, S.L.; Laveriano-Santos, E.P.; Marhuenda-Muñoz, M.; Storniolo, C.E.; Tresserra-Rimbau, A.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M. Health Effects of Resveratrol: Results from Human Intervention Trials. Nutrients 2018, 10, 1892. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, İ. Antioxidant Properties of Resveratrol: A Structure–Activity Insight. Innov. Food Sci. Emerg. Technol. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant Polyphenols as Antioxidant and Antibacterial Agents for Shelf-Life Extension of Meat and Meat Products: Classification, Structures, Sources, and Action Mechanisms: Polyphenols Extending Meat Shelf-Life…. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Daiber, A.; Förstermann, U.; Li, H. Antioxidant Effects of Resveratrol in the Cardiovascular System: Antioxidant Effects of Resveratrol. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and Protective Effects of Natural Carotenoids. Biochim. Biophys. Acta 2005, 1740, 101–107. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Fiedor, J.; Fiedor, L.; Haessner, R.; Scheer, H. Cyclic Endoperoxides of Beta-Carotene, Potential pro-Oxidants, as Products of Chemical Quenching of Singlet Oxygen. Biochim. Biophys. Acta 2005, 1709, 1–4. [Google Scholar] [CrossRef]
- Terao, J.; Minami, Y.; Bando, N. Singlet Molecular Oxygen-Quenching Activity of Carotenoids: Relevance to Protection of the Skin from Photoaging. J. Clin. Biochem. Nutr. 2011, 48, 57–62. [Google Scholar] [CrossRef]
- Kiokias, S.; Gordon, M.H. Antioxidant Properties of Carotenoids in Vitro and in Vivo. Food Rev. Int. 2004, 20, 99–121. [Google Scholar] [CrossRef]
- Moussa, Z.; Judeh, Z.M.A.; Ahmed, S.A. Nonenzymatic Exogenous and Endogenous Antioxidants. In Free Radical Medicine and Biology; IntechOpen: London, UK, 2020; ISBN 9781789851434. [Google Scholar]
- Chang, C.-S.; Chang, C.-L.; Lai, G.-H. Reactive Oxygen Species Scavenging Activities in a Chemiluminescence Model and Neuroprotection in Rat Pheochromocytoma Cells by Astaxanthin, Beta-Carotene, and Canthaxanthin. Kaohsiung J. Med. Sci. 2013, 29, 412–421. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction and Scavenging Mechanism of β-Carotene and Zeaxanthin with Reactive Oxygen Species. J. Oleo Sci. 2017, 66, 77–84. [Google Scholar] [CrossRef]
- Yamauchi, R. Vitamin E: Mechanism of Its Antioxidant Activity. Food Sci. Technol. Int. Tokyo 1997, 3, 301–309. [Google Scholar] [CrossRef]
- Tucker, J.M.; Townsend, D.M. Alpha-Tocopherol: Roles in Prevention and Therapy of Human Disease. Biomed. Pharmacother. 2005, 59, 380–387. [Google Scholar] [CrossRef]
- Ebhohimen, I.E.; Okanlawon, T.S.; Osagie, A.O.; Izevbigie, O.N. Vitamin E in Human Health and Oxidative Stress Related Diseases. In Vitamin E in Health and Disease—Interactions, Diseases and Health Aspects; Erkekoglu, P., Santos, J.S., Eds.; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/77766 (accessed on 7 June 2022). [CrossRef]
- Ha, Y.L.; Csallany, A.S. Alpha-Tocopherol Oxidation Mediated by Superoxide Anion (O2-). II. Identification of the Stable Alpha-Tocopherol Oxidation Products. Lipids 1992, 27, 201–205. [Google Scholar] [CrossRef]
- Yamauchi, R.; Matsushita, S. Products Formed by Photosensitized Oxidation of Tocopherols. Agric. Biol. Chem. 1979, 43, 2151–2156. [Google Scholar] [CrossRef]
- Niki, E.; Noguchi, N. Antioxidant Action of Vitamin E in Vivo as Assessed from Its Reaction Products with Multiple Biological Oxidants. Free Radic. Res. 2021, 55, 352–363. [Google Scholar] [CrossRef]
- Liebler, D.C.; Burr, J.A.; Philips, L.; Ham, A.J. Gas Chromatography-Mass Spectrometry Analysis of Vitamin E and Its Oxidation Products. Anal. Biochem. 1996, 236, 27–34. [Google Scholar] [CrossRef]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic Acid: Chemistry, Biology and the Treatment of Cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef]
- Wilson, J.X. Regulation of Vitamin C Transport. Annu. Rev. Nutr. 2005, 25, 105–125. [Google Scholar] [CrossRef]
- Zhitkovich, A. Nuclear and Cytoplasmic Functions of Vitamin C. Chem. Res. Toxicol. 2020, 33, 2515–2526. [Google Scholar] [CrossRef]
- Carr, A.C.; Zhu, B.-Z.; Frei, B. Potential Antiatherogenic Mechanisms of Ascorbate (Vitamin C) and α-Tocopherol (Vitamin E). Circ. Res. 2000, 87, 349–354. [Google Scholar] [CrossRef]
- Figueroa-Méndez, R.; Rivas-Arancibia, S. Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain. Front. Physiol. 2015, 6, 397. [Google Scholar] [CrossRef]
- Sriset, Y.; Chatuphonprasert, W.; Jarukamjorn, K. Optimized Models of Xenobiotic-Induced Oxidative Stress in HepG2 Cells. Trop. J. Pharm. Res. 2021, 18, 1001–1007. [Google Scholar] [CrossRef]
- Chen, Y.; Qing, W.; Sun, M.; Lv, L.; Guo, D.; Jiang, Y. Melatonin Protects Hepatocytes against Bile Acid-Induced Mitochondrial Oxidative Stress via the AMPK-SIRT3-SOD2 Pathway. Free Radic. Res. 2015, 49, 1275–1284. [Google Scholar] [CrossRef]
- Yu, F.-Y.; Chu, T.-Y.; Lian, J.-D.; Wu, S.-W.; Hung, T.-W.; Chang, H.-R. Induction of Oxidative Stress and DNA Damage in Human Renal Proximal Tubular Cells by Aristolochic Acid. J. Food Drug Anal. 2011, 19, 2. [Google Scholar] [CrossRef]
- Bist, R.; Chaudhary, B.; Bhatt, D.K. Defensive Proclivity of Bacoside A and Bromelain against Oxidative Stress and AChE Gene Expression Induced by Dichlorvos in the Brain of Mus Musculus. Sci. Rep. 2021, 11, 3668. [Google Scholar] [CrossRef]
- Aykutoglu, G.; Tartik, M.; Darendelioglu, E.; Ayna, A.; Baydas, G. Melatonin and Vitamin E Alleviate Homocysteine-Induced Oxidative Injury and Apoptosis in Endothelial Cells. Mol. Biol. Rep. 2020, 47, 5285–5293. [Google Scholar] [CrossRef]
- Pazdro, R.; Burgess, J.R. Differential Effects of α-Tocopherol and N-Acetyl-Cysteine on Advanced Glycation End Product-Induced Oxidative Damage and Neurite Degeneration in SH-SY5Y Cells. Biochim. Biophys. Acta 2012, 1822, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qu, W.; Kadiiska, M.B. Role of Oxidative Stress in Cadmium Toxicity and Carcinogenesis. Toxicol. Appl. Pharmacol. 2009, 238, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Chang, P.; Yang, L.; Wang, Y.; Zhu, S.; Shan, H.; Zhang, M.; Tao, L. Neuroprotective Effects of Hydrogen Sulfide on Sodium Azide-Induced Oxidative Stress in PC12 Cells. Int. J. Mol. Med. 2018, 41, 242–250. [Google Scholar] [CrossRef]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative Stress and Cellular Response to Doxorubicin: A Common Factor in the Complex Milieu of Anthracycline Cardiotoxicity. Oxid. Med. Cell. Longev. 2017, 2017, 1521020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Chiang, E.-P.I. Low-Dose Methotrexate Inhibits Methionine S-Adenosyltransferase in Vitro and in Vivo. Mol. Med. 2012, 18, 423–432. [Google Scholar] [CrossRef]
- Omolara, O.O. Induction of Oxidative Stress by Methotrexate in the Testicular Tissue of Swiss Albino Mice. Ann. Clin. Toxicol. 2019, 2, 1022. [Google Scholar]
- Oktem, F.; Yilmaz, H.R.; Ozguner, F.; Olgar, S.; Ayata, A.; Uzare, E.; Uz, E. Methotrexate-Induced Renal Oxidative Stress in Rats: The Role of a Novel Antioxidant Caffeic Acid Phenethyl Ester. Toxicol. Ind. Health 2006, 22, 241–247. [Google Scholar] [CrossRef]
- Cortés-Iza, S.C.; Rodríguez, A.I. Oxidative Stress and Pesticide Disease: A Challenge for Toxicology. Rev. Fac. Med. Univ. Nac. Colomb. 2018, 66, 261–267. [Google Scholar] [CrossRef]
- Eraslan, G.; Saygi, S.; Essiz, D.; Aksoy, A.; Gul, H.; Macit, E. Evaluation of Aspect of Some Oxidative Stress Parameters Using Vitamin E, Proanthocyanidin and N-Acetylcysteine against Exposure to Cyfluthrin in Mice. Pestic. Biochem. Physiol. 2007, 88, 43–49. [Google Scholar] [CrossRef]
- Sadowska-Woda, I.; Wójcik, N.; Karowicz-Bilińska, A.; Bieszczad-Bedrejczuk, E. Effect of Selected Antioxidants in Beta-Cyfluthrin-Induced Oxidative Stress in Human Erythrocytes in Vitro. Toxicol. In Vitro 2010, 24, 879–884. [Google Scholar] [CrossRef]
- Gumpricht, E.; Dahl, R.; Devereaux, M.W.; Sokol, R.J. Β-Carotene Prevents Bile Acid-Induced Cytotoxicity in the Rat Hepatocyte: Evidence for an Antioxidant and Anti-Apoptotic Role of β-Carotene in Vitro. Pediatr. Res. 2004, 55, 814–821. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, Y.; Feng, Y.; Zhang, H.; Li, L.; Shen, W.; Zhao, M.; Min, L. Β-Carotene Improves Oocyte Development and Maturation under Oxidative Stress in Vitro. In Vitro Cell Dev. Biol. Anim. 2019, 55, 548–558. [Google Scholar] [CrossRef]
- Revin, V.V.; Gromova, N.V.; Revina, E.S.; Samonova, A.Y.; Tychkov, A.Y.; Bochkareva, S.S.; Moskovkin, A.A.; Kuzmenko, T.P. The Influence of Oxidative Stress and Natural Antioxidants on Morphometric Parameters of Red Blood Cells, the Hemoglobin Oxygen Binding Capacity, and the Activity of Antioxidant Enzymes. BioMed. Res. Int. 2019, 2019, 2109269. [Google Scholar] [CrossRef]
- Luqman, S.; Kaushik, S.; Srivastava, S.; Kumar, R.; Bawankule, D.U.; Pal, A.; Darokar, M.P.; Khanuja, S.P.S. Protective Effect of Medicinal Plant Extracts on Biomarkers of Oxidative Stress in Erythrocytes. Pharm. Biol. 2009, 47, 483–490. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, X.; Wang, H.; Chen, Z. Beta Carotene Protects H9c2 Cardiomyocytes from Advanced Glycation End Product-Induced Endoplasmic Reticulum Stress, Apoptosis, and Autophagy via the PI3K/Akt/MTOR Signaling Pathway. Ann. Transl. Med. 2020, 8, 647. [Google Scholar] [CrossRef]
- Palozza, P.; Luberto, C.; Ricci, P.; Sgarlata, E.; Calviello, G.; Bartoli, G.M. Effect of Beta-Carotene and Canthaxanthin on Tert-Butyl Hydroperoxide-Induced Lipid Peroxidation in Murine Normal and Tumor Thymocytes. Arch. Biochem. Biophys. 1996, 325, 145–151. [Google Scholar] [CrossRef]
- Wu, C.-M.; Cheng, Y.-L.; Dai, Y.-H.; Chen, M.-F.; Wang, C.-C. α-Tocopherol Protects Keratinocytes against Ultraviolet A Irradiation by Suppressing Glutathione Depletion, Lipid Peroxidation and Reactive Oxygen Species Generation. Biomed. Rep. 2014, 2, 419–423. [Google Scholar] [CrossRef]
- Eroğlu, S.; Pandir, D.; Uzun, F.G.; Bas, H. Protective Role of Vitamins C and E in Dichlorvos-Induced Oxidative Stress in Human Erythrocytes in Vitro. Biol. Res. 2013, 46, 33–38. [Google Scholar] [CrossRef]
- Zappe, K.; Pointner, A.; Switzeny, O.J.; Magnet, U.; Tomeva, E.; Heller, J.; Mare, G.; Wagner, K.-H.; Knasmueller, S.; Haslberger, A.G. Counteraction of Oxidative Stress by Vitamin E Affects Epigenetic Regulation by Increasing Global Methylation and Gene Expression of MLH1 and DNMT1 Dose Dependently in Caco-2 Cells. Oxid. Med. Cell. Longev. 2018, 2018, 3734250. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Pan, Y.; Wang, H.; Wei, C.; Yu, Y. Vitamin E (A-tocopherol) Ameliorates Aristolochic Acid-induced Renal Tubular Epithelial Cell Death by Attenuating Oxidative Stress and Caspase-3 Activation. Mol. Med. Rep. 2018, 17, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, X.; Du, M.; Li, F.; Xiao, M.; Zhang, W. Synergistic Antioxidant Effects of Araloside A and L-Ascorbic Acid on H2O2-Induced HEK293 Cells: Regulation of Cellular Antioxidant Status. Oxid. Med. Cell. Longev. 2021, 2021, 9996040. [Google Scholar] [CrossRef] [PubMed]
- Tram, N.K.; McLean, R.M.; Swindle-Reilly, K.E. Glutathione Improves the Antioxidant Activity of Vitamin C in Human Lens and Retinal Epithelial Cells: Implications for Vitreous Substitutes. Curr. Eye Res. 2021, 46, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Kim, Y.J.; Lee, E.K.; Park, S.W.; Yu, H.G. Antioxidative Effects of Ascorbic Acid and Astaxanthin on ARPE-19 Cells in an Oxidative Stress Model. Antioxidants 2020, 9, 833. [Google Scholar] [CrossRef]
- Wu, T.-K.; Wei, C.-W.; Pan, Y.-R.; Cherng, S.-H.; Chang, W.-J.; Wang, H.-F.; Yu, Y.-L. Vitamin C Attenuates the Toxic Effect of Aristolochic Acid on Renal Tubular Cells via Decreasing Oxidative Stress-mediated Cell Death Pathways. Mol. Med. Rep. 2015, 12, 6086–6092. [Google Scholar] [CrossRef]
- Gęgotek, A.; Bielawska, K.; Biernacki, M.; Zaręba, I.; Surażyński, A.; Skrzydlewska, E. Comparison of Protective Effect of Ascorbic Acid on Redox and Endocannabinoid Systems Interactions in in Vitro Cultured Human Skin Fibroblasts Exposed to UV Radiation and Hydrogen Peroxide. Arch. Derm. Res. 2017, 309, 285–303. [Google Scholar] [CrossRef]
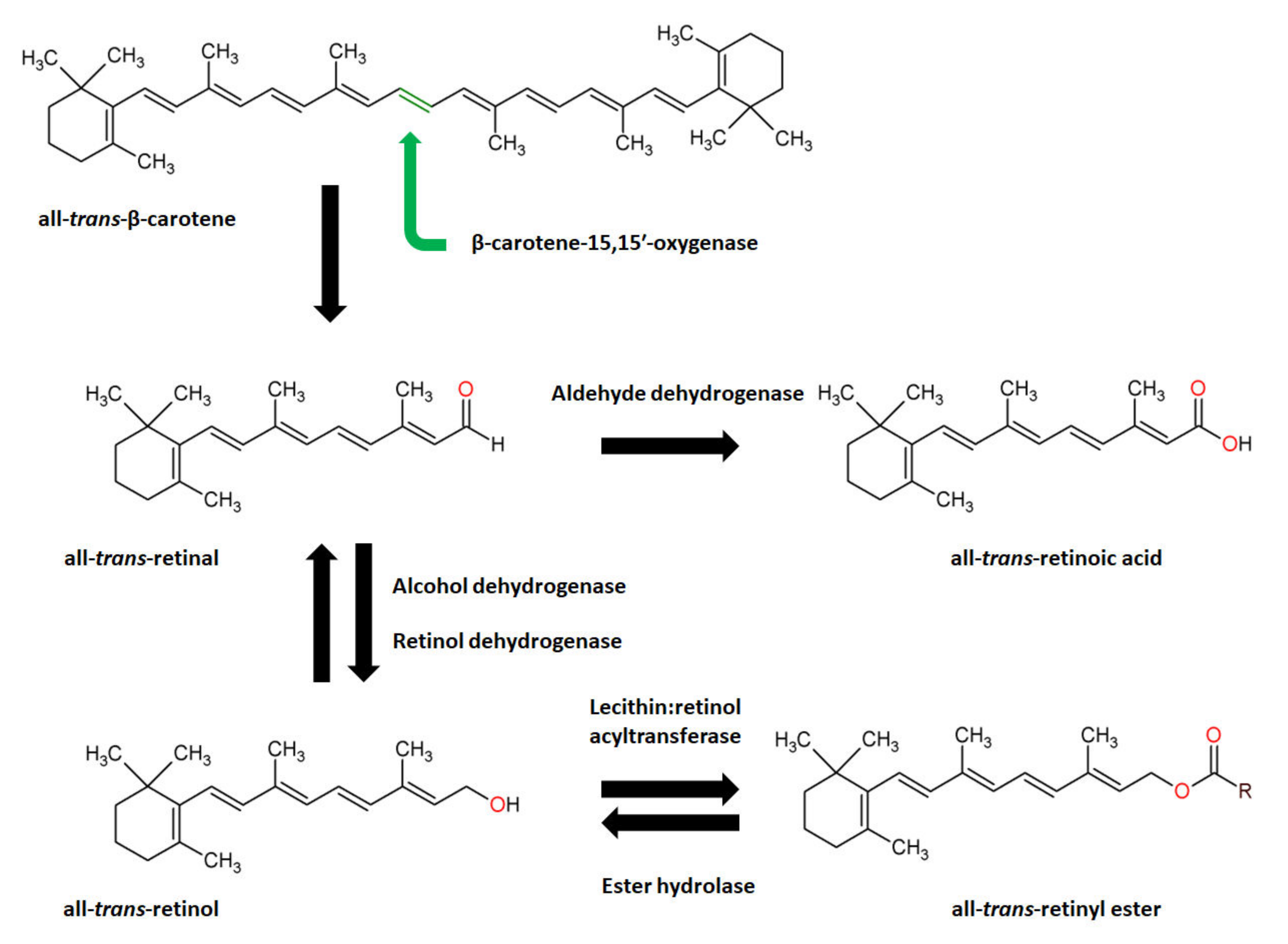
- Shete, V.; Quadro, L. Mammalian Metabolism of β-Carotene: Gaps in Knowledge. Nutrients 2013, 5, 4849–4868. [Google Scholar] [CrossRef]
- Haeseleer, F.; Huang, J.; Lebioda, L.; Saari, J.C.; Palczewski, K. Molecular Characterization of a Novel Short-Chain Dehydrogenase/Reductase That Reduces All-Trans-Retinal. J. Biol. Chem. 1998, 273, 21790–21799. [Google Scholar] [CrossRef]
- Getrude, N.; Albert, N.; Gabriel, G.; Finbarrs-Bello, E.; Elizabeth, C.; Austin, O. Beta-Carotene: Positive Effect on Oxidative Stress, Lipid Peroxidation, Insulin and Leptin Resistance Induced by Dietary Fat Consumption. J. Adv. Med. Med. Res. 2018, 27, 1–7. [Google Scholar] [CrossRef]
- Vardi, N.; Parlakpinar, H.; Cetin, A.; Erdogan, A.; Cetin Ozturk, I. Protective Effect of Beta-Carotene on Methotrexate-Induced Oxidative Liver Damage. Toxicol. Pathol. 2010, 38, 592–597. [Google Scholar] [CrossRef]
- Zhou, L.; Ouyang, L.; Lin, S.; Chen, S.; Liu, Y.; Zhou, W.; Wang, X. Protective Role of β-Carotene against Oxidative Stress and Neuroinflammation in a Rat Model of Spinal Cord Injury. Int. Immunopharmacol. 2018, 61, 92–99. [Google Scholar] [CrossRef]
- Chen, P.; Li, L.; Gao, Y.; Xie, Z.; Zhang, Y.; Pan, Z.; Tu, Y.; Wang, H.; Han, Q.; Hu, X.; et al. β-Carotene Provides Neuro Protection after Experimental Traumatic Brain Injury via the Nrf2-ARE Pathway. J. Integr. Neurosci. 2019, 18, 153–161. [Google Scholar] [CrossRef]
- Nagpal, I.; Abraham, S.K. Protective Effects of Tea Polyphenols and β-Carotene against γ-Radiation Induced Mutation and Oxidative Stress in Drosophila Melanogaster. Genes Environ. 2017, 39, 24. [Google Scholar] [CrossRef]
- Smolarek, A.K.; Suh, N. Chemopreventive Activity of Vitamin E in Breast Cancer: A Focus on γ- and δ-Tocopherol. Nutrients 2011, 3, 962–986. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, K.; Chojnacka, A.; Górnicka, M. Tocopherols and Tocotrienols—Bioactive Dietary Compounds; What Is Certain, What Is Doubt? Int. J. Mol. Sci. 2021, 22, 6222. [Google Scholar] [CrossRef] [PubMed]
- Schmölz, L.; Birringer, M.; Lorkowski, S.; Wallert, M. Complexity of Vitamin E Metabolism. World J. Biol. Chem. 2016, 7, 14–43. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E. Vitamin E Intestinal Absorption: Regulation of Membrane Transport across the Enterocyte: Vitamin E Membrane Transport in the Enterocyte. IUBMB Life 2019, 71, 416–423. [Google Scholar] [CrossRef]
- Rigotti, A. Absorption, Transport, and Tissue Delivery of Vitamin E. Mol. Asp. Med. 2007, 28, 423–436. [Google Scholar] [CrossRef]
- Novelli Filho, J.L.V.B.; Novelli, E.L.B.; Manzano, M.A.; Lopes, A.M.; Cataneo, A.C.; Barbosa, L.L.; Ribas, B.O. Effect of Alpha-Tocopherol on Superoxide Radical and Toxicity of Cadmium Exposure. Int. J. Environ. Health Res. 2000, 10, 125–134. [Google Scholar] [CrossRef]
- Donia, T.; Eldaly, S.; Ali, E.M.M. Ameliorating Oxidative Stress and Inflammation by Hesperidin and Vitamin E in Doxorubicin Induced Cardiomyopathy. Turk. J. Biochem. 2019, 44, 207–217. [Google Scholar] [CrossRef]
- Wallert, M.; Ziegler, M.; Wang, X.; Maluenda, A.; Xu, X.; Yap, M.L.; Witt, R.; Giles, C.; Kluge, S.; Hortmann, M.; et al. α-Tocopherol Preserves Cardiac Function by Reducing Oxidative Stress and Inflammation in Ischemia/Reperfusion Injury. Redox Biol. 2019, 26, 101292. [Google Scholar] [CrossRef]
- Wang, X.; Dong, W.; Yuan, B.; Yang, Y.; Yang, D.; Lin, X.; Chen, C.; Zhang, W. Vitamin E Confers Cytoprotective Effects on Cardiomyocytes under Conditions of Heat Stress by Increasing the Expression of Metallothionein. Int. J. Mol. Med. 2016, 37, 1429–1436. [Google Scholar] [CrossRef]
- Shen, X.; Tang, Q.; Wu, J.; Feng, Y.; Huang, J.; Cai, W. Effect of Vitamin E Supplementation on Oxidative Stress in a Rat Model of Diet-Induced Obesity. Int. J. Vitam. Nutr. Res. 2009, 79, 255–263. [Google Scholar] [CrossRef]
- Leskovec, J.; Levart, A.; Nemec Svete, A.; Perić, L.; Đukić Stojčić, M.; Žikić, D.; Salobir, J.; Rezar, V. Effects of Supplementation with α-Tocopherol, Ascorbic Acid, Selenium, or Their Combination in Linseed Oil-Enriched Diets on the Oxidative Status in Broilers. Poult. Sci. 2018, 97, 1641–1650. [Google Scholar] [CrossRef]
- Carr, A.C.; Vissers, M.C.M. Synthetic or Food-Derived Vitamin C—Are They Equally Bioavailable? Nutrients 2013, 5, 4284–4304. [Google Scholar] [CrossRef]
- Chambial, S.; Dwivedi, S.; Shukla, K.K.; John, P.J.; Sharma, P. Vitamin C in Disease Prevention and Cure: An Overview. Indian J. Clin. Biochem. 2013, 28, 314–328. [Google Scholar] [CrossRef]
- Pappenberger, G.; Hohmann, H.-P. Industrial Production of L-Ascorbic Acid (Vitamin C) and D-Isoascorbic Acid. Adv. Biochem. Eng. Biotechnol. 2014, 143, 143–188. [Google Scholar] [CrossRef]
- Ranjan, R.; Ranjan, A.; Dhaliwal, G.S.; Patra, R.C. L-Ascorbic Acid (Vitamin C) Supplementation to Optimize Health and Reproduction in Cattle. Vet. Q. 2012, 32, 145–150. [Google Scholar] [CrossRef]
- Naidu, K.A. Vitamin C in Human Health and Disease Is Still a Mystery? An Overview. Nutr. J. 2003, 2, 7. [Google Scholar] [CrossRef]
- Lindblad, M.; Tveden-Nyborg, P.; Lykkesfeldt, J. Regulation of Vitamin C Homeostasis during Deficiency. Nutrients 2013, 5, 2860–2879. [Google Scholar] [CrossRef]
- Akinrinde, A.S.; Fapuro, J.; Soetan, K.O. Zinc and Ascorbic Acid Treatment Alleviates Systemic Inflammation and Gastrointestinal and Renal Oxidative Stress Induced by Sodium Azide in Rats. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 17. [Google Scholar] [CrossRef]
- Akolkar, G.; da Silva Dias, D.; Ayyappan, P.; Bagchi, A.K.; Jassal, D.S.; Salemi, V.M.C.; Irigoyen, M.C.; De Angelis, K.; Singal, P.K. Vitamin C Mitigates Oxidative/Nitrosative Stress and Inflammation in Doxorubicin-Induced Cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H795–H809. [Google Scholar] [CrossRef]
- Shati, A.A.; Zaki, M.S.A.; Alqahtani, Y.A.; Haidara, M.A.; Al-Shraim, M.; Dawood, A.F.; Eid, R.A. Potential Protective Effect of Vitamin C on Qunalphos-Induced Cardiac Toxicity: Histological and Tissue Biomarker Assay. Biomedicines 2022, 10, 39. [Google Scholar] [CrossRef]
- Kasperczyk, S.; Dobrakowski, M.; Kasperczyk, J.; Ostałowska, A.; Zalejska-Fiolka, J.; Birkner, E. Beta-Carotene Reduces Oxidative Stress, Improves Glutathione Metabolism and Modifies Antioxidant Defense Systems in Lead-Exposed Workers. Toxicol. Appl. Pharmacol. 2014, 280, 36–41. [Google Scholar] [CrossRef]
- Levy, Y.; Zaltsberg, H.; Ben-Amotz, A.; Kanter, Y.; Aviram, M. Dietary Supplementation of a Natural Isomer Mixture of Beta-Carotene Inhibits Oxidation of LDL Derived from Patients with Diabetes Mellitus. Ann. Nutr. Metab. 2000, 44, 54–60. [Google Scholar] [CrossRef]
- Roberts, L.J., 2nd; Oates, J.A.; Linton, M.F.; Fazio, S.; Meador, B.P.; Gross, M.D.; Shyr, Y.; Morrow, J.D. The Relationship between Dose of Vitamin E and Suppression of Oxidative Stress in Humans. Free Radic. Biol. Med. 2007, 43, 1388–1393. [Google Scholar] [CrossRef]
- McArdle, F.; Rhodes, L.E.; Parslew, R.A.G.; Close, G.L.; Jack, C.I.A.; Friedmann, P.S.; Jackson, M.J. Effects of Oral Vitamin E and Beta-Carotene Supplementation on Ultraviolet Radiation-Induced Oxidative Stress in Human Skin. Am. J. Clin. Nutr. 2004, 80, 1270–1275. [Google Scholar] [CrossRef]
- Helmersson, J.; Arnlöv, J.; Larsson, A.; Basu, S. Low Dietary Intake of Beta-Carotene, Alpha-Tocopherol and Ascorbic Acid Is Associated with Increased Inflammatory and Oxidative Stress Status in a Swedish Cohort. Br. J. Nutr. 2009, 101, 1775–1782. [Google Scholar] [CrossRef]
- Ble-Castillo, J.L.; Carmona-Díaz, E.; Méndez, J.D.; Larios-Medina, F.J.; Medina-Santillán, R.; Cleva-Villanueva, G.; Díaz-Zagoya, J.C. Effect of Alpha-Tocopherol on the Metabolic Control and Oxidative Stress in Female Type 2 Diabetics. Biomed. Pharmacother. 2005, 59, 290–295. [Google Scholar] [CrossRef]
- Ruknarong, L.; Boonthongkaew, C.; Chuangchot, N.; Jumnainsong, A.; Leelayuwat, N.; Jusakul, A.; Gaudieri, S.; Leelayuwat, C. Vitamin C Supplementation Reduces Expression of Circulating MiR-451a in Subjects with Poorly Controlled Type 2 Diabetes Mellitus and High Oxidative Stress. PeerJ 2021, 9, e10776. [Google Scholar] [CrossRef]
- Shinde, S.N.; Dhadke, V.N.; Suryakar, A.N. Evaluation of Oxidative Stress in Type 2 Diabetes Mellitus and Follow-up along with Vitamin E Supplementation. Indian J. Clin. Biochem. 2011, 26, 74–77. [Google Scholar] [CrossRef]
- Tantavisut, S.; Tanavalee, A.; Honsawek, S.; Suantawee, T.; Ngarmukos, S.; Adisakwatana, S.; Callaghan, J.J. Effect of Vitamin E on Oxidative Stress Level in Blood, Synovial Fluid, and Synovial Tissue in Severe Knee Osteoarthritis: A Randomized Controlled Study. BMC Musculoskelet. Disord. 2017, 18, 281. [Google Scholar] [CrossRef]
- Kuhn, M.J.; Sordillo, L.M. Vitamin E Analogs Limit in Vitro Oxidant Damage to Bovine Mammary Endothelial Cells. J. Dairy Sci. 2021, 104, 7154–7167. [Google Scholar] [CrossRef]
- Bohn, T. Carotenoids and Markers of Oxidative Stress in Human Observational Studies and Intervention Trials: Implications for Chronic Diseases. Antioxidants 2019, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Appel, L.J.; Croft, K.D.; Miller, E.R., 3rd; Mori, T.A.; Puddey, I.B. Effects of Vitamin C and Vitamin E on in Vivo Lipid Peroxidation: Results of a Randomized Controlled Trial. Am. J. Clin. Nutr. 2002, 76, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Ullegaddi, R.; Powers, H.J.; Gariballa, S.E. Antioxidant Supplementation Enhances Antioxidant Capacity and Mitigates Oxidative Damage Following Acute Ischaemic Stroke. Eur. J. Clin. Nutr. 2005, 59, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Berretta, M.; Quagliariello, V.; Maurea, N.; Di Francia, R.; Sharifi, S.; Facchini, G.; Rinaldi, L.; Piezzo, M.; Manuela, C.; Nunnari, G.; et al. Multiple Effects of Ascorbic Acid against Chronic Diseases: Updated Evidence from Preclinical and Clinical Studies. Antioxidants 2020, 9, 1182. [Google Scholar] [CrossRef]
- Lin, J.; Cook, N.R.; Albert, C.; Zaharris, E.; Gaziano, J.M.; Van Denburgh, M.; Buring, J.E.; Manson, J.E. Vitamins C and E and Beta Carotene Supplementation and Cancer Risk: A Randomized Controlled Trial. J. Natl. Cancer Inst. 2009, 101, 14–23. [Google Scholar] [CrossRef]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids that Exhibit both Anti- and Pro-Oxidative Mechanisms—A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef]
- Lohan, S.B.; Lauer, A.-C.; Arndt, S.; Friedrich, A.; Tscherch, K.; Haag, S.F.; Darvin, M.E.; Vollert, H.; Kleemann, A.; Gersonde, I.; et al. Determination of the Antioxidant Status of the Skin by In Vivo-Electron Paramagnetic Resonance (EPR) Spectroscopy. Cosmetics 2015, 2, 286–301. [Google Scholar] [CrossRef]
- Kohno, M. Applications of Electron Spin Resonance Spectrometry for Reactive Oxygen Species and Reactive Nitrogen Species Research. J. Clin. Biochem. Nutr. 2010, 47, 1–11. [Google Scholar] [CrossRef]
- Togashi, H.; Shinzawa, H.; Matsuo, T.; Takeda, Y.; Takahashi, T.; Aoyama, M.; Oikawa, K.; Kamada, H. Analysis of Hepatic Oxidative Stress Status by Electron Spin Resonance Spectroscopy and Imaging. Free Radic. Biol. Med. 2000, 28, 846–853. [Google Scholar] [CrossRef]
- Gotham, J.P.; Li, R.; Tipple, T.E.; Lancaster, J.R., Jr.; Liu, T.; Li, Q. Quantitation of Spin Probe-Detectable Oxidants in Cells Using Electron Paramagnetic Resonance Spectroscopy: To Probe or to Trap? Free Radic. Biol. Med. 2020, 154, 84–94. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Lima, J.L.F.C. Fluorescence Probes Used for Detection of Reactive Oxygen Species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar] [CrossRef]
- Kazemi, S.Y.; Abedirad, S.M. Effect of FsDNA on Chemiluminescence Characteristics of Luminol–H2O2 System Catalyzed by Mn(III)–Tetrakis (4-Sulfonatophenyl)-Porphyrin. J. Iran. Chem. Soc. 2013, 10, 251–256. [Google Scholar] [CrossRef]
- Chi, Q.; Chen, W.; He, Z. Mechanism of Alcohol-Enhanced Lucigenin Chemiluminescence in Alkaline Solution: Alcohol-Enhanced Lucigenin Chemiluminescence. Luminescence 2015, 30, 990–995. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Weitner, T.; Inić, S.; Jablan, J.; Gabričević, M.; Domijan, A.-M. Spectrophotometric Determination of Malondialdehyde in Urine Suitable for Epidemiological Studies. Croat. Chem. Acta 2016, 89, 133–139. [Google Scholar] [CrossRef]
- Ito, F.; Sono, Y.; Ito, T. Measurement and Clinical Significance of Lipid Peroxidation as a Biomarker of Oxidative Stress: Oxidative Stress in Diabetes, Atherosclerosis, and Chronic Inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef]
- Russ, M.; Weinheimer, L.; Jauk, S.; Andrä, M.; Wintersteiger, R.; Ortner, A. Characterization of Oxidative Stress in Chicken Heart Tissue by High-Performance Liquid Chromatography with Ultraviolet-visible (HPLC–UV–Vis) Detection. Anal. Lett. 2019, 52, 892–901. [Google Scholar] [CrossRef]
- Freeman, L.M.; Rush, J.E.; Milbury, P.E.; Blumberg, J.B. Antioxidant Status and Biomarkers of Oxidative Stress in Dogs with Congestive Heart Failure. J. Vet. Intern. Med. 2005, 19, 537. [Google Scholar] [CrossRef]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of Malondialdehyde (MDA) by High-Performance Liquid Chromatography in Serum and Liver as a Biomarker for Oxidative Stress. Application to a Rat Model for Hypercholesterolemia and Evaluation of the Effect of Diets Rich in Phenolic Antioxidants from Fruits. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 827, 76–82. [Google Scholar] [CrossRef]
- Vanova, N.; Muckova, L.; Schmidt, M.; Herman, D.; Dlabkova, A.; Pejchal, J.; Jun, D. Simultaneous Determination of Malondialdehyde and 3-Nitrotyrosine in Cultured Human Hepatoma Cells by Liquid Chromatography-Mass Spectrometry. Biomed. Chromatogr. 2018, 32, e4349. [Google Scholar] [CrossRef]
- Tsikas, D.; Rothmann, S.; Schneider, J.Y.; Gutzki, F.-M.; Beckmann, B.; Frölich, J.C. Simultaneous GC-MS/MS Measurement of Malondialdehyde and 4-Hydroxy-2-Nonenal in Human Plasma: Effects of Long-Term L-Arginine Administration. Anal. Biochem. 2017, 524, 31–44. [Google Scholar] [CrossRef]
- Bevan, R.J.; Durand, M.F.; Hickenbotham, P.T.; Kitas, G.D.; Patel, P.R.; Podmore, I.D.; Griffiths, H.R.; Waller, H.L.; Lunec, J. Validation of a Novel ELISA for Measurement of MDA-LDL in Human Plasma. Free Radic. Biol. Med. 2003, 35, 517–527. [Google Scholar] [CrossRef]
- Mehta, K.; Patel, V.B. Measurement of 4-Hydroxynonenal (4-HNE) Protein Adducts by ELISA. Methods Mol. Biol. 2019, 1990, 43–52. [Google Scholar] [CrossRef]
- Coolen, S.A.J.; van Buuren, B.; Duchateau, G.; Upritchard, J.; Verhagen, H. Kinetics of Biomarkers: Biological and Technical Validity of Isoprostanes in Plasma. Amino Acids 2005, 29, 429–436. [Google Scholar] [CrossRef]
- Liu, W.; Morrow, J.D.; Yin, H. Quantification of F2-Isoprostanes as a Reliable Index of Oxidative Stress in Vivo Using Gas Chromatography–Mass Spectrometry (GC-MS) Method. Free Radic. Biol. Med. 2009, 47, 1101–1107. [Google Scholar] [CrossRef]
- Södergren, E.; Cederberg, J.; Basu, S.; Vessby, B. Vitamin E Supplementation Decreases Basal Levels of F(2)-Isoprostanes and Prostaglandin f(2alpha) in Rats. J. Nutr. 2000, 130, 10–14. [Google Scholar] [CrossRef]
- Basu, S. ReviewIsoprostanes: Novel Bioactive Products of Lipid Peroxidation. Free Radic. Res. 2004, 38, 105–122. [Google Scholar] [CrossRef]
- Södergren, E.; Nourooz-Zadeh, J.; Berglund, L.; Vessby, B. Re-Evaluation of the Ferrous Oxidation in Xylenol Orange Assay for the Measurement of Plasma Lipid Hydroperoxides. J. Biochem. Biophys. Methods 1998, 37, 137–146. [Google Scholar] [CrossRef]
- Nourooz-Zadeh, J. Ferrous Ion Oxidation in Presence of Xylenol Orange for Detection of Lipid Hydroperoxides in Plasma. Methods Enzymol. 1999, 300, 58–62. [Google Scholar] [CrossRef]
- Castegna, A.; Drake, J.; Pocernich, C.; Butterfield, D.A. Protein Carbonyl Levels—An Assessment of Protein Oxidation. In Methods in Biological Oxidative Stress; Humana Press: Totowa, NJ, USA, 2003; pp. 161–168. ISBN 9781592594245. [Google Scholar]
- Weber, D.; Davies, M.J.; Grune, T. Determination of Protein Carbonyls in Plasma, Cell Extracts, Tissue Homogenates, Isolated Proteins: Focus on Sample Preparation and Derivatization Conditions. Redox Biol. 2015, 5, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Nestorova, G.; Rissman, R.A.; Feng, J. Detection and Quantification of 8-Hydroxy-2′-Deoxyguanosine in Alzheimer’s Transgenic Mouse Urine Using Capillary Electrophoresis: CE and CEC. Electrophoresis 2013, 34, 2268–2274. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′ -Deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Chepelev, N.L.; Kennedy, D.A.; Gagné, R.; White, T.; Long, A.S.; Yauk, C.L.; White, P.A. HPLC Measurement of the DNA Oxidation Biomarker, 8-Oxo-7,8-Dihydro-2′-Deoxyguanosine, in Cultured Cells and Animal Tissues. J. Vis. Exp. 2015, 102, e52697. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Jaruga, P.; Rodriguez, H. Measurement of 8-Hydroxy-2′-Deoxyguanosine in DNA by High-Performance Liquid Chromatography-Mass Spectrometry: Comparison with Measurement by Gas Chromatography-Mass Spectrometry. Nucleic Acids Res. 2001, 29, E12. [Google Scholar] [CrossRef]
- Devpura, S.; Barton, K.N.; Brown, S.L.; Palyvoda, O.; Kalkanis, S.; Naik, V.M.; Siddiqui, F.; Naik, R.; Chetty, I.J. Vision 20/20: The Role of Raman Spectroscopy in Early Stage Cancer Detection and Feasibility for Application in Radiation Therapy Response Assessment: Raman Spectroscopy for Cancer Detection/Radiation Therapy Response Assessment. Med. Phys. 2014, 41, 050901. [Google Scholar] [CrossRef]
- Pence, I.; Mahadevan-Jansen, A. Clinical Instrumentation and Applications of Raman Spectroscopy. Chem. Soc. Rev. 2016, 45, 1958–1979. [Google Scholar] [CrossRef]
- Imiela, A.; Surmacki, J.; Abramczyk, H. Novel Strategies of Raman Imaging for Monitoring the Therapeutic Benefit of Temozolomide in Glioblastoma. J. Mol. Struct. 2020, 1217, 128381. [Google Scholar] [CrossRef]
- Abramczyk, H.; Surmacki, J.; Kopeć, M.; Olejnik, A.K.; Kaufman-Szymczyk, A.; Fabianowska-Majewska, K. Epigenetic Changes in Cancer by Raman Imaging, Fluorescence Imaging, AFM and Scanning near-Field Optical Microscopy (SNOM). Acetylation in Normal and Human Cancer Breast Cells MCF10A, MCF7 and MDA-MB-231. Analyst 2016, 141, 5646–5658. [Google Scholar] [CrossRef]
- Brozek-Pluska, B. Statistics Assisted Analysis of Raman Spectra and Imaging of Human Colon Cell Lines–Label Free, Spectroscopic Diagnostics of Colorectal Cancer. J. Mol. Struct. 2020, 1218, 128524. [Google Scholar] [CrossRef]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2007, 42, 493–541. [Google Scholar] [CrossRef]
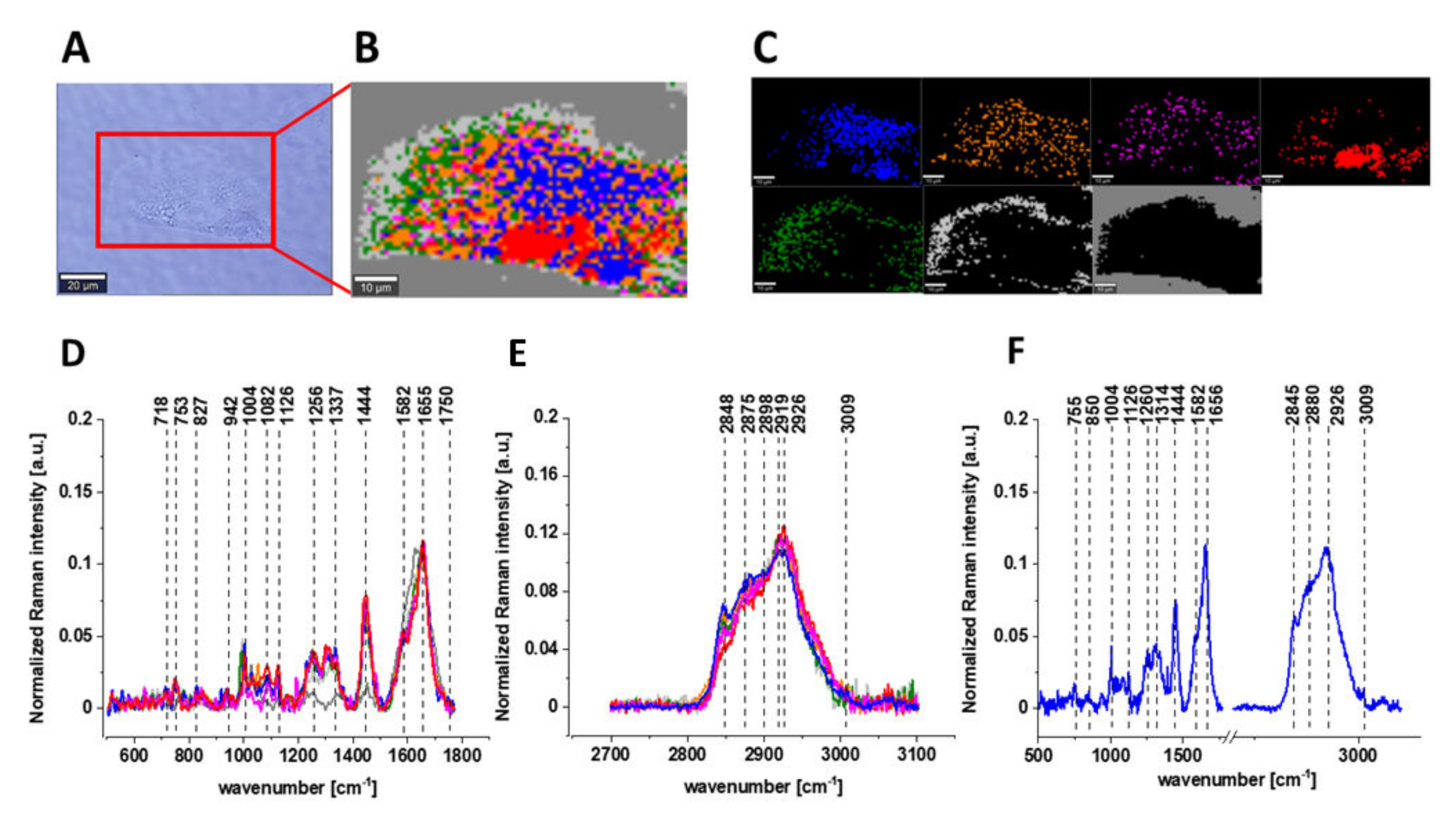
- Beton, K.; Brozek-Pluska, B. Vitamin C—Protective Role in Oxidative Stress Conditions Induced in Human Normal Colon Cells by Label-Free Raman Spectroscopy and Imaging. Int. J. Mol. Sci. 2021, 22, 6928. [Google Scholar] [CrossRef]
- Brozek-Pluska, B.; Beton, K. Oxidative Stress Induced by TBHP in Human Normal Colon Cells by Label Free Raman Spectroscopy and Imaging. The Protective Role of Natural Antioxidants in the Form of β-Carotene. RSC Adv. 2021, 11, 16419–16434. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exemplary Structures of Oxidation Products | ||
|---|---|---|
| Name | Structure Description | Mechanism of Formation |
| Malondialdehyde (MDA) |  | PUFA peroxyl radical undergoes intramolecular cyclization to endoperoxide with further breakdown to MDA |
| 4-hydroxy-2-nonenal (4-HNE; HNE) |  | Peroxidation of n-6 PUFAs and the generation of α,β unsaturated aldehydes |
| Dityrosine |  | Generation of a tyrosyl radical, radical isomerization, diradical reaction, and enolization |
| HNE-Lys adduct (protein carbonyl product) |  | Michael addition of the HNE double bond to NH2-group of lysine (Lys) |
| HNE-Cys adduct (protein carbonyl product) |  | Michael addition of the HNE double bond to SH-group of cysteine (Cys) |
| HNE-His adduct (protein carbonyl product) |  | Michael addition of the HNE double bond to NH in an imidazole of histidine (His) |
| 8-oxo-2′-deoxyguanosine nucleotide |  | Reaction between C-8 of guanine (G) and hydroxyl radical (•OH) |
| 8-oxo-2′-deoxyadenosine nucleotide |  | Reaction between C-8 of adenine (A) and hydroxyl radical (•OH) |
| 5,6-dihydroxy-5,6-dihydrothymidine nucleotide |  | Reaction of hydroxyl radical (•OH) with C-5 and C-6 of thymine (T) |
| 5,6-dihydroxy-5,6-dihydrocytidine nucleotide |  | Reaction of hydroxyl radical (•OH) with C-5 and C-6 of cytosine (C) |
| Antioxidant Effect of β-Carotene Towards OS-Induced Cells | ||||
|---|---|---|---|---|
| Cell Type | Oxidative Stress Inducer | Exposure Mode | Intracellular Effect | Ref. |
| Rat pheochromocytoma (PC12) cells (undifferentiated) | H2O2 | Incubation: 12 h without β-C and then 12 h with 40 µM H2O2 | ↑1 ROS | [153] |
| Incubation: 12 h with 0.5–10 µM β-C and then 12 h with 40 µM H2O2 | ↓2 ROS | |||
| Rat hepatocytes | Glycocheno-deoxycholic acid (GCDC) | Incubation: 0.5 h without β-C and then 4 h with 100 µM GCDC | ↑1 ROS | [183] |
| Incubation: 0.5 h with 50 µM β-C and then 4 h with 100 µM GCDC | ↓2 ROS | |||
| Oocytes | Rosup | Incubation: 4 h with 33.3 µg/mL Rosup | ↑1 ROS | [184] |
| Incubation: 4 h with 10 µM β-C and 33.3 µg/mL Rosup | ↓2 ROS | |||
| Human erythrocytes | H2O2 | Incubation: 0.5 h with 20 µM H2O2 | ↑1 lipid peroxidation, ↓1 SOD, ↓1 CAT | [185] |
| Incubation: 0.5 h with 3 µM β-C and 20 µM H2O2 | ↓2 lipid peroxidation, ↑2 SOD, ↑2 CAT | |||
| Mice erythrocytes | H2O2 | Incubation: 2 mM H2O2 | ↑1 MDA | [186] |
| Incubation: 10 µg/mL β-C and 2 mM H2O2 | ↓2 MDA | |||
| Cardiomyocyte cells (H9c2) | AGEs | Incubation: 24 h with 200 µg/mL AGEs | ↑1 ROS, ↑1 MDA, ↓1 GPx, ↓1 SOD | [187] |
| Incubation: 24 h with 200 µg/mL AGEs + 24 h with 40 µM β-C | ↓2 ROS, ↓2 MDA, ↑2 GPx, ↑2 SOD | |||
| Normal and tumor thymocytes | t-BuOOH | Incubation: 2 h with 0.5 mM t-BuOOH | ↑1 MDA | [188] |
| Incubation: 0.5 h with β-C (2.8 nmol/mg dry wt) and 2 h with 0.5 mM t-BuOOH | ↓2 MDA | |||
| Antioxidant Effect of Vitamin E towards OS-Induced Cells | ||||
|---|---|---|---|---|
| Cell Type | Oxidative Stress Inducer | Exposure Mode | Exposure Effect | Ref. |
| Human keratinocyte cells | UVA | Irradiation (UVA, 8 J/cm2) + incubation for 24 h | ↑1 ROS, ↑1 MDA | [189] |
| Incubation for 24 h with α-T (2.9–14.7 IU/mL), irradiation (UVA, 8 J/cm2) + incubation for 24 h | ↓2 ROS, ↓2 MDA | |||
| Human neuroblastoma (SH-SY5Y) cells | AGEs | Incubation for 24 h without α-T, then incubation with 1.5 mg/mL AGEs for 72 h | ↑1 lipid peroxidation, ↑1 protein carbonyls | [173] |
| Incubation for 24 h with α-T (200 µM), then incubation with 1.5 mg/mL AGEs for 72 h | ↓2 lipid peroxidation, ↓2 protein carbonyls | |||
| Human umbilical vein endothelial cells | Hcy | Incubation with Hcy (1 mM) | ↑1 ROS, ↑1 MDA | [172] |
| Incubation with Hcy (1 mM) and VitE (50 µM) | ↓2 ROS, ↓2 MDA | |||
| Human erythrocytes | Dichlorvos (DDVP) | Incubation with DDVP (10 µM) | ↑1 MDA, ↓1 SOD, ↓1 CAT, ↓1 GPx | [190] |
| Incubation with DDVP (10 µM) and Vit E (30 µM) | ↓2 MDA, ↑2 SOD, ↑2 CAT, ↑2 GPx | |||
| Human colorectal adenocarcinoma cell line (Caco-2) | H2O2 | Incubation for 24 h, incubation for 48 h with H2O2 (250 µM) | ↑1 MDA | [191] |
| Incubation for 24 h, incubation for 48 h with H2O2 (250 µM) and VitE (10 µM) | ↓2 MDA | |||
| Rat hepatocytes | Glycocheno-deoxycholic acid | Incubation: 0.5 h without α-T and then 4 h with 100 µM GCDC | ↑1 ROS | [183] |
| Incubation: 0.5 h with 100 µM α-T and then 4 h with 100 µM GCDC | ↓2 ROS | |||
| Rat renal tubular epithelial cells (NRK-52E) | Aristolochic acid (AA) | Incubation: AA (10 µM) | ↑1 H2O2 | [192] |
| Incubation: AA (10 µM) + α-T (5–100 µM) | ↓2 H2O2 | |||
| Antioxidant Effect of Ascorbic Acid towards OS-Induced Cells | ||||
|---|---|---|---|---|
| Cell Type | Oxidative Stress Inducer | Exposure Mode | Exposure Effect | Ref. |
| Human embryonic kidney (HEK293) cells | H2O2 | Incubation: 4 h with H2O2 (400 µM) | ↑1 ROS, ↑1 MDA, ↓1 CAT, ↓1 SOD, ↓1 GPx, ↑1 protein carbonyl, ↑1 8-OHdG | [193] |
| Incubation: 24 h with Vit C (1–20 µM), and then 4 h with H2O2 (400 µM) | ↓2 ROS, ↓2 MDA, ↑2 CAT, ↑2 SOD, ↑2 GPx, ↓2 protein carbonyl, ↓2 8-OHdG | |||
| Human lens epithelial cells (LEC) | H2O2 | Incubation: 24 h without VitC, and 0.5 h with H2O2 (0.2 mM) | ↑1 ROS | [194] |
| Incubation: 24 h with VitC (1 mM), and 0.5 h with H2O2 (0.2 mM) | ↓2 ROS | |||
| Human retinal pigment epithelial (ARPE-19) cells | H2O2 or UVB | Incubation: UVB irradiation (20–100 mJ/cm2) Incubation: 24 h with H2O2 (0.2 mM) | ↑1 ROS | [195] |
| Incubation: 6 h with VitC (500 µM), then UVB irradiation (100 mJ/cm2) Incubation: 6 h with VitC (20 µM), then 24 h with H2O2 (0.2 mM) | ↓2 ROS | |||
| Human erythrocytes | Dichlorvos | Incubation with DDVP (10 µM) | ↑1 MDA, ↓1 SOD, ↓1 CAT, ↓1 GPx | [190] |
| Incubation with DDVP (10 µM) and Vit C (10 µM) | ↓2 MDA, ↑2 SOD, ↑2 CAT, ↑2 GPx | |||
| Human hepatoma (HepG2) cells | Ethanol, sodium selenite or t-BuOOH | Incubation with ethanol (10–500 µM) or sodium selenite (1–10 µM) or t-BuOOH (20–200 µM) for 24 h | ↑1 MDA ↓1 SOD, ↓1 CAT, ↓1 GSH | [168] |
| Cotreatment with Vit C (25–100 µM) and one of OS inducer for 24 h | ↓2 MDA, ↑2 SOD, ↑2 CAT, ↑2 GSH | |||
| Human skin fibroblasts (CCD 1112Sk) | UVA, UVB or H2O2 | For irradiation treatment: 20 J/cm2 (UVA) or 200 mJ/cm2 (UVB) + 24 h incubation; For H2O2 treatment: incubation with 200 µM H2O2 for 24 h | ↑1 ROS, ↑1 MDA, ↑1 4-HNE, ↑1 Carbonyl groups (for all stress inducers); ↑1 Isoprostanes (for UVA, H2O2) | [197] |
| Stress induction treatment + 24 h incubation with 100 µM ascorbic acid | ↓2 ROS, ↓2 MDA, ↓2 4-HNE, ↓2 Carbonyl groups (for all stress inducers); ↓2 Isoprostanes (for UVA, H2O2) | |||
| Rat renal tubular epithelial cells (NRK-52E) | Aristolochic acid (AA) | Incubation: AA (10 µM) | ↑1 H2O2 | [196] |
| Incubation: AA (10 µM) + Vit C (5 µM) | ↓2 H2O2 | |||
| Antioxidant Effect of β-Carotene, Vitamin E and/or Vitamin C In Vivo | ||||
|---|---|---|---|---|
| Species/Tissue | Oxidative Stress Inducer | Exposure Mode | Exposure Effect | Ref. |
| Wistar rats n = 24 (total)/Liver tissue | Methotrexate (MTX) | Single MTX dose (20 mg/kg) on day 21 of experiment (24 days) | ↑1 MDA, ↓1 SOD, ↓1 CAT, ↓1 GPx | [201] |
| β-C dose (10 mg/kg/day) for 24 days + MTX dose (20 mg/kg) on day 21 | ↓2 MDA, ↑2 SOD, ↑2 CAT, ↑2 GPx | |||
| Wistar rats n = 30 (total)/Blood | High-fat diet (HFD) (Mixing of cow fat (60%) with normal rat chow (40%)) | HFD for 14 weeks + 24 h starving | ↑1 MDA | [200] |
| β-C administration (300 mg/kg body weight) for 2 weeks before or after 12-week HFD + 24 h starving | ↓2 MDA | |||
| Male Sprague–Dawley rats n = 299 (total)/Spinal Cord tissue | Spinal Cord Injury (SCI) | SCI surgery + 72 h | ↑1 MDA, ↓1 SOD | [202] |
| SCI surgery + β-C (20–80 mg/kg) administered intraperitoneally once immediately after the surgery + 72 h | ↓2 MDA, ↑2 SOD | |||
| Male C57BL/6 mice n = 108 (total)/Brain tissue | Traumatic Brain Injury (TBI) | TBI surgery + 7 days | ↑1 MDA, ↓1 SOD | [203] |
| TBI surgery + β-C (30 mg/kg) administered 3 h after the surgery and then every day during 7 days | ↓2 MDA, ↑2 SOD | |||
| Drosophila melanogaster larvae | Gamma irradiation | Exposure to 10 Gy γ-irradiation | ↑1 Lipid peroxidation | [204] |
| Larvae feeding on β-C before exposure to 10 Gy γ-irradiation | ↓2 Lipid peroxidation | |||
| Wistar male rats n = 60 (total)/Serum | Cd2+ (CdCl2·H2O) | Single intraperitoneal injection of Cd2+ ions (2 mg Kg−1) | ↑1 Lipoperoxide, ↓1 SOD | [210] |
| Single intraperitoneal injection of Cd2+ ions (2 mg/Kg) + administration of drink aqueous solutions of α-T (40 mg/L) for 15 days | ↓2 Lipoperoxide, ↑2 SOD | |||
| Male Sprague–Dawley rats n = 70 (total)/Heart tissue | Doxorubicin (DOX) | Intraperitoneal injection (4 mg DOX/ kg body weight) three times per week for 2 weeks | ↑1 MDA, ↑1 CAT | [211] |
| Intra-gastric administration (100 mg VitE/kg body weight), two times per week for 3 weeks, started 1 week before DOX injection | ↓2 MDA, ↓2 CAT | |||
| Female white mice n = 160 (total)/Blood | Cyfluthrin | Oral administration of cyfluthrin (a single dose of 100 mg/kg/body weight) | ↑1 MDA, ↓1 CAT | [181] |
| Oral administration of cyfluthrin (a dose of 100 mg/kg/body weight) followed by intramuscular injection of VitE (a dose of 100 mg/kg/body weight, for 7 days) | ↓2 MDA, ↑2 CAT | |||
| C57BL/6 mice/Infarcted tissue | Ischemia/Reperfusion (I/R) injury | I/R injury + 3 days | ↑1 Oxidized lipids | [212] |
| Intraperitoneal injection of α-TOH (2.5 mg/kg BW) 2 h before surgery, immediately after reperfusion and twice per day for 3 days | ↓2 Oxidized lipids | |||
| BALB/c mice n = 40 (total)/Heart tissue | Heat stress (HS) | HS conditions (temperature: 40 °C; humidity: 60%) for 4 h per day during a 4-week period | ↑1 MDA, ↓1 SOD | [213] |
| Oral administration of VitE (500 mg/kg) 2 h before the initiation of HS | ↓2 MDA, ↑2 SOD | |||
| Sprague-Dawley male rats n = 30 (total)/Blood | High-fat diet | A 10-week feeding on high-fat diet | ↑1 Lipid peroxidation, ↓1 SOD, ↓1 GPx | [214] |
| A 10-week feeding on high-fat diet supplemented with VitE (350 mg/kg diet) | ↓2 Lipid peroxidation, ↑2 SOD, ↑2 GPx | |||
| Ross 308 male broilers (21-day old) n = 400 (total)/Blood, Breast muscle | High n-3 dietary PUFAs intake | Chickens fed with commercial starter diet (1–12 days), commercial grower diet (13–20 days), finisher diet enriched with 5% cold-pressed linseed oil and supplemented with VitE (200 IU/kg) (21–40 days) | ↓3 MDA | [215] |
| Wistar rats n = 28 (total)/Stomach, Colon, Kidney tissue | Sodium azide (NaN3) | Oral administration of NaN3 (20 mg/kg BW) for 9 days | ↑1 MDA, ↑1 Protein Carbonyls | [222] |
| Oral administration of NaN3 and VitC (200 mg/kg BW) for 9 days | ↓2 MDA, ↓2 Protein Carbonyls | |||
| Wistar male rats n = 46/Heart tissue | Doxorubicin (DOX) | Six intraperitoneal DOX injections (2.5 mg/kg body wt) over 3 weeks | ↑1 Superoxide anion, ↑1 Lipid peroxidation, ↑1 Protein Carbonyls | [223] |
| Oral daily administration of VitC (50 mg/kg) started 1 week before the start of DOX administration and continued for 2 weeks after the last DOX injection | ↓2 Superoxide anion, ↓2 Lipid peroxidation, ↓2 Protein Carbonyls | |||
| Sprague–Dawley male rats n = 18 (total)/Heart tissue | Quinalphos (QP) | Oral dose of QP (14 mg/kg), daily for 10 days | ↑1 MDA, ↓1 CAT, ↓1 GPx | [224] |
| Oral administration of VitC (20 mg/kg) daily, 4 h after QP administration, for 10 days | ↓2 MDA, ↑2 CAT, ↑2 GPx | |||
| Antioxidant Effect of β-Carotene, Vitamin E and/or Vitamin C in Human Studies | ||||
|---|---|---|---|---|
| Patients/Participants | Origin of Oxidative Stress | Treatment Mode | Treatment Effect | Ref. |
| Male workers exposed to lead, n = 85 (total)/Blood | Pb | No administration of antioxidants for 12 weeks | - | [225] |
| Oral administration of β-carotene (10 mg/day) for 12 weeks | ↓1,2 MDA, ↓1,2 LHP | |||
| Patients with NIDDM, n = 20 (total)/Blood | NIDDM | Supplementation with β-carotene (60 mg/day) for 3 weeks | ↓C1 MDA | [226] |
| Participants with polygenic hypercholesterolemia, n = 35/Blood | Enhanced oxidative stress | Supplementation with placebo for 16 weeks | - | [227] |
| Supplementation with VitE (1600–3200 I.U.) for 16 weeks | ↓2 F2-isoprostane | |||
| White volunteers, n = 8 (total)/Skin | UVR | Supplementation with α-tocopherol (400 IU/day) for 8 weeks, skin exposure to UVR (120 mJ/cm2), skin biopsy 6 h after UVR exposure | ↓2 MDA | [228] |
| Participants at age of 77, n = 704/Urine | n.d. | Dietary intake of ascorbic acid and β-carotene | ↓I F2-isoprostane | [229] |
| Female participants with type 2 diabetes, n = 34 (total)/Blood | Type 2 diabetes | Oral administration of placebo (800 mg/day) for 6 weeks | - | [230] |
| Oral administration of α-tocopherol (800 IU/day) for 6 weeks | ↓1,2 MDA | |||
| Participants with DMT2, n = 5 or 8 (total)/Blood | DMT2 | Supplementation with VitC (1000 mg/day) for 6 weeks | ↓1 MDA ↓1 F2-isoprostane | [231] |
| Participants with DMT2 and administered with hypoglycemic drug, n = 80 (total)/Blood | DMT2 | No supplementation of VitE within 3 months | ↑1,C2 MDA | [232] |
| Supplementation of VitE (400 mg/day) for 3 months | ↓1,2 MDA | |||
| Patients with late-stage knee osteoarthritis, n = 72 (total)/Blood, synovial fluid | Enhanced oxidative stress | Oral administration of placebo once daily for 2 months | - | [233] |
| Oral administration of VitE (400 IU/day) for 2 months | ↓2 MDA | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miazek, K.; Beton, K.; Śliwińska, A.; Brożek-Płuska, B. The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used. Biomolecules 2022, 12, 1087. https://doi.org/10.3390/biom12081087
Miazek K, Beton K, Śliwińska A, Brożek-Płuska B. The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used. Biomolecules. 2022; 12(8):1087. https://doi.org/10.3390/biom12081087
Chicago/Turabian StyleMiazek, Krystian, Karolina Beton, Agnieszka Śliwińska, and Beata Brożek-Płuska. 2022. "The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used" Biomolecules 12, no. 8: 1087. https://doi.org/10.3390/biom12081087