
Neuroprotective Effects of Chlorogenic Acid in a Mouse Model of Intracerebral Hemorrhage Associated with Reduced Extracellular Matrix Metalloproteinase Inducer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Intracerebral Hemorrhage Model of Mice
2.3. Experimental Design
2.3.1. Experiment 1
2.3.2. Experiment 2
2.3.3. Experiment 3
2.3.4. Experiment 4
2.3.5. Experiment 5
2.4. Neurological Function Tests
2.5. Analysis of Brain Cell Death
2.6. Measurement of Brain Water Content
2.7. Assessment of BBB Permeability
2.8. Immunofluorescence
2.9. Western Blot Analysis
2.10. Hematoma Volume Measurement
2.11. Hematoxylin and Eosin (H&E) Staining
2.12. Statistical Analysis
3. Results
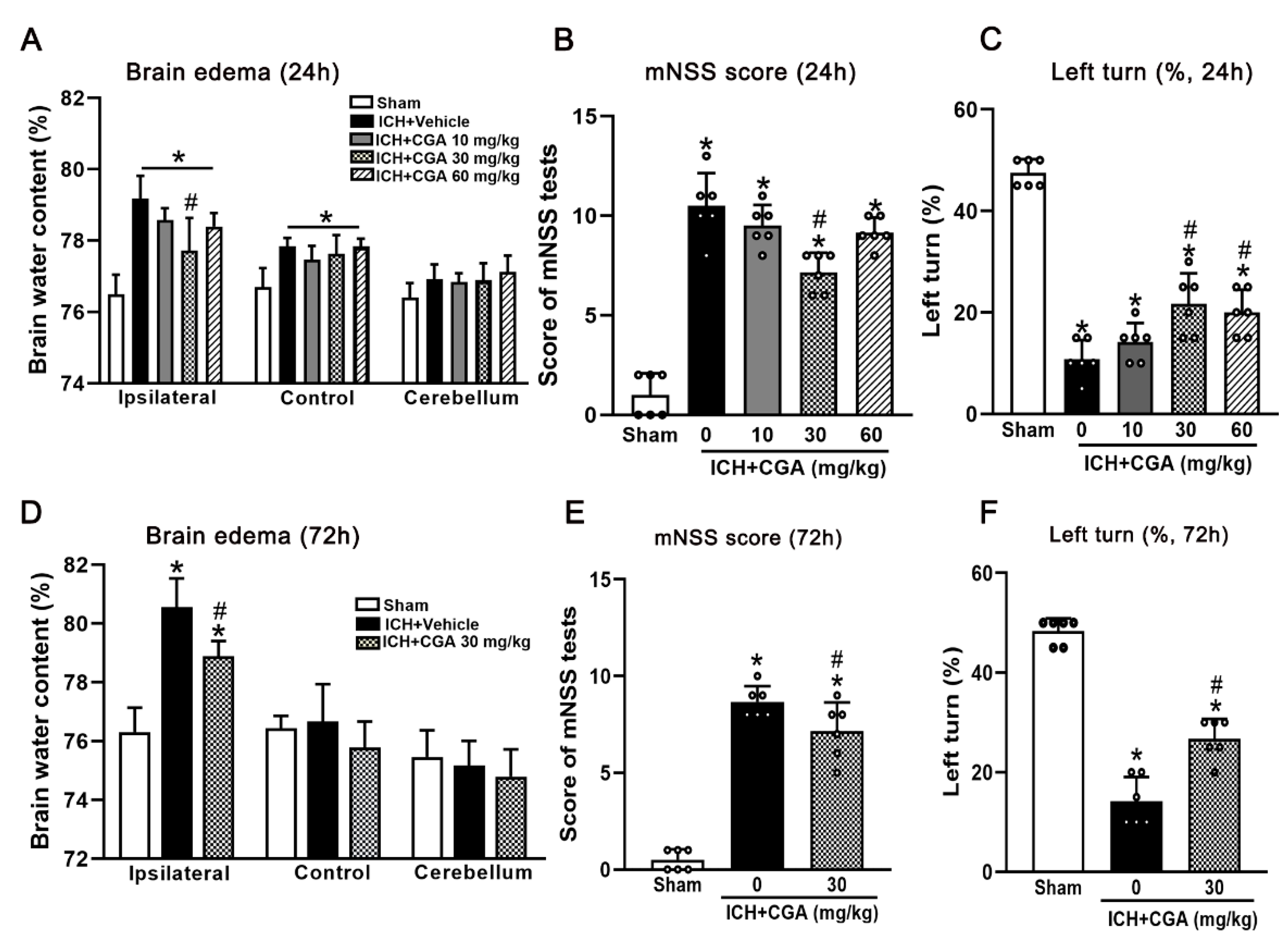
3.1. CGA Treatment Attenuated Neurological Impairments and Brain Edema at 24 h and 72 h after ICH
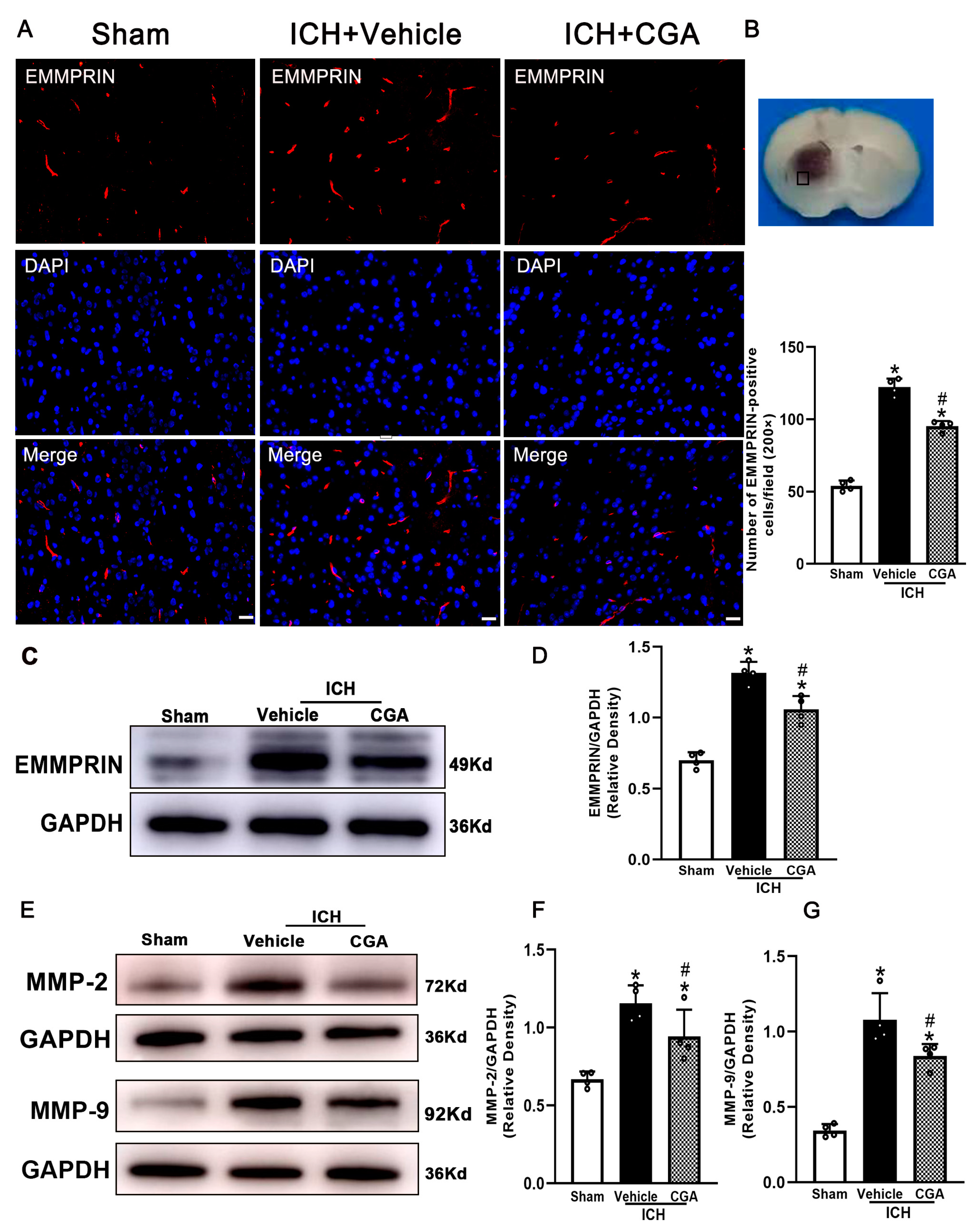
3.2. CGA Treatment Reduced the Expression of EMMPRIN and MMP-2/9
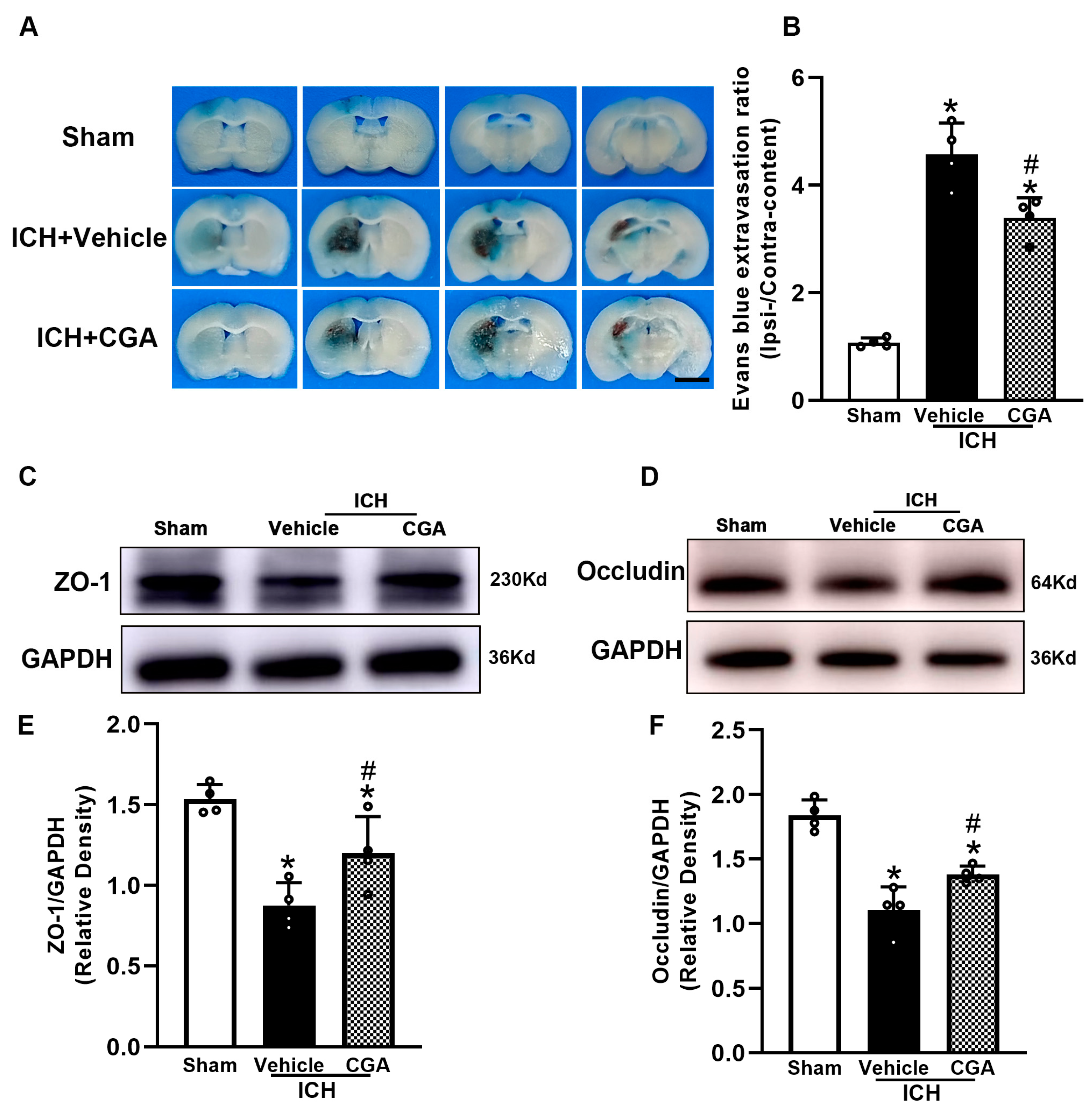
3.3. CGA Reduced BBB Permeability after ICH
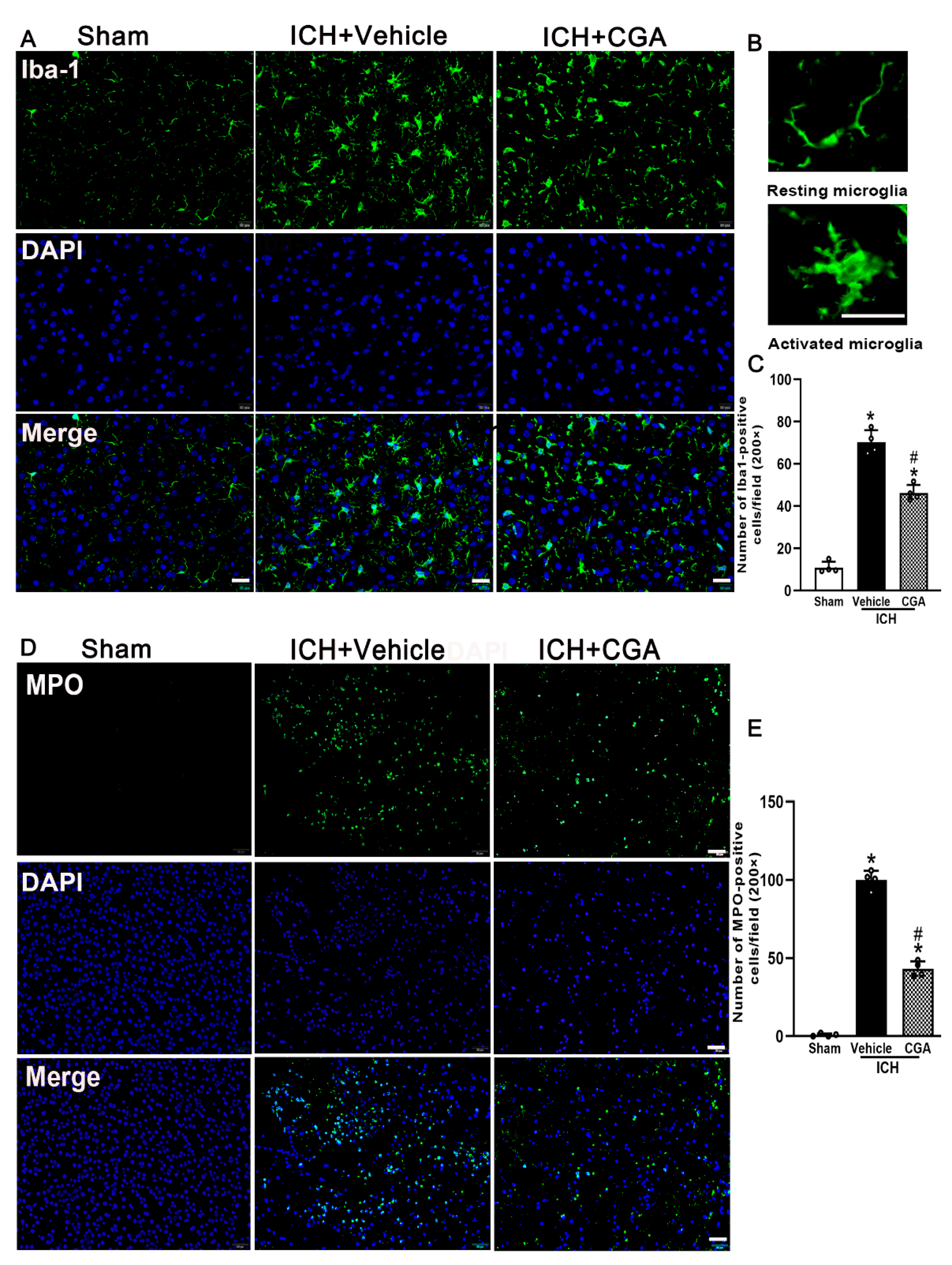
3.4. CGA Treatment Alleviated Microglia/Macrophage Activation and Neutrophil Infiltration after ICH
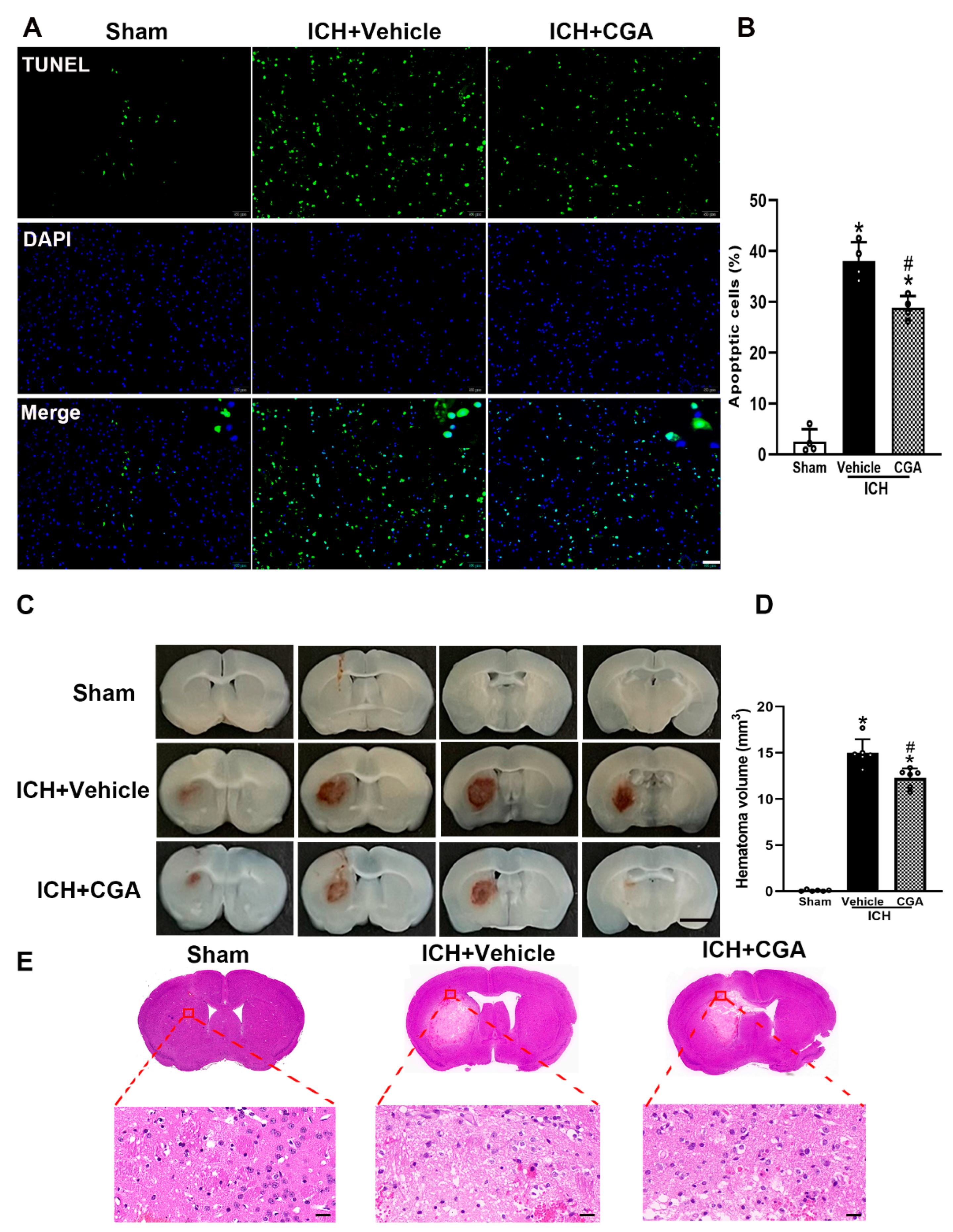
3.5. CGA Treatment Reduced Cell Death and Brain Injury after ICH
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Asch, C.J.; Luitse, M.J.; Rinkel, G.J.; van der Tweel, I.; Algra, A.; Klijn, C.J. Incidence, case fatality, and functional outcome of intracerebral haemorrhage over time, according to age, sex, and ethnic origin: A systematic review and meta-analysis. Lancet Neurol. 2010, 9, 167–176. [Google Scholar] [CrossRef]
- Al-Shahi Salman, R.; Labovitz, D.L.; Stapf, C. Spontaneous intracerebral haemorrhage. BMJ 2009, 339, b2586. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.W.; Wu, X.L.; Li, J.; Liu, Z.X.; Chen, F.H.; Zhang, L.B.; Zhang, H.F.; Wan, X.; Cheng, Q. Minimally Invasive Surgery is Superior to Conventional Craniotomy in Patients with Spontaneous Supratentorial Intracerebral Hemorrhage: A Systematic Review and Meta-Analysis. World Neurosurg. 2018, 115, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Hanley, D.F.; Thompson, R.E.; Rosenblum, M. Efficacy and safety of minimally invasive surgery with thrombolysis in intracerebral haemorrhage evacuation (MISTIE III): A randomised, controlled, open-label, blinded endpoint phase 3 trial. Lancet 2019, 393, 1596. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Khan, S.; Liu, Y.; Wu, G.; Yong, V.W.; Xue, M. Oxidative Stress Following Intracerebral Hemorrhage: From Molecular Mechanisms to Therapeutic Targets. Front. Immunol. 2022, 13, 847246. [Google Scholar] [CrossRef]
- Ford, G.A. Clinical pharmacological issues in the development of acute stroke therapies. Br. J. Pharmacol. 2008, 153 (Suppl. 1), S112–S119. [Google Scholar] [CrossRef]
- Morotti, A.; Brouwers, H.B.; Romero, J.M.; Jessel, M.J.; Vashkevich, A.; Schwab, K.; Afzal, M.R.; Cassarly, C.; Greenberg, S.M.; Martin, R.H.; et al. Intensive Blood Pressure Reduction and Spot Sign in Intracerebral Hemorrhage: A Secondary Analysis of a Randomized Clinical Trial. JAMA Neurol. 2017, 74, 950–960. [Google Scholar] [CrossRef]
- Baharoglu, M.I.; Cordonnier, C.; Salman, R.A.S.; de Gans, K.; Koopman, M.M.; Brand, A.; Majoie, C.B.; Beenen, L.F.; Marquering, H.A.; Vermeulen, M.; et al. Investigators, P. Platelet transfusion versus standard care after acute stroke due to spontaneous cerebral haemorrhage associated with antiplatelet therapy (PATCH): A randomised, open-label, phase 3 trial. Lancet 2016, 387, 2605–2613. [Google Scholar] [CrossRef]
- Zhang, R.; Xue, M.; Yong, V.W. Central Nervous System Tissue Regeneration after Intracerebral Hemorrhage: The Next Frontier. Cells 2021, 10, 2513. [Google Scholar] [CrossRef]
- Lan, X.; Han, X.; Liu, X.; Wang, J. Inflammatory responses after intracerebral hemorrhage: From cellular function to therapeutic targets. J. Cereb. Blood Flow Metab. 2019, 39, 184–186. [Google Scholar] [CrossRef]
- Xue, M.; Yong, V.W. Neuroinflammation in intracerebral haemorrhage: Immunotherapies with potential for translation. Lancet Neurol. 2020, 19, 1023–1032. [Google Scholar] [CrossRef]
- Bai, Q.; Xue, M.Z.; Yong, V.W. Microglia and macrophage phenotypes in intracerebral haemorrhage injury: Therapeutic opportunities. Brain 2020, 143, 1297–1314. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.M.; Yong, V.W. The many faces of EMMPRIN—Roles in neuroinflammation. Biochim. Biophys. Acta 2011, 1812, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morancho, A.; Rosell, A.; Garcia-Bonilla, L.; Montaner, J. Metalloproteinase and stroke infarct size: Role for anti-inflammatory treatment? Ann. N. Y. Acad. Sci. 2010, 1207, 123–133. [Google Scholar] [CrossRef]
- Xue, M.; Yong, V.W. Matrix metalloproteinases in intracerebral hemorrhage. Neurol. Res. 2008, 30, 775–782. [Google Scholar] [CrossRef]
- Tejima, E.; Zhao, B.Q.; Tsuji, K.; Rosell, A.; van Leyen, K.; Gonzalez, R.G.; Montaner, J.; Wang, X.Y.; Lo, E.H. Astrocytic induction of matrix metalloproteinase-9 and edema in brain hemorrhage. J. Cereb. Blood Flow Metab. 2007, 27, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Lattanzi, S.; Di Napoli, M.; Ricci, S.; Divani, A.A. Matrix Metalloproteinases in Acute Intracerebral Hemorrhage. Neurotherapeutics 2020, 17, 484–496. [Google Scholar] [CrossRef]
- Patrizz, A.; Doran, S.J.; Han, A.C.; Ahnstedt, H.; Roy-O’Reilly, M.; Lai, Y.J.; Weston, G.; Tarabishy, S.; Patel, A.R.; Verma, R.; et al. EMMPRIN/CD147 plays a detrimental role in clinical and experimental ischemic stroke. Aging 2020, 12, 5139–5152. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Khan, S.; Zhang, R.Y.; Wei, R.X.; Zhang, Y.; Xue, M.Z.; Yong, V.W. Neuroprotection of minocycline by inhibition of extracellular matrix metalloproteinase inducer expression following intracerebral hemorrhage in mice. Neurosci. Lett. 2021, 764, 136297. [Google Scholar] [CrossRef]
- Zhang, R.; Yong, V.W.; Xue, M. Revisiting Minocycline in Intracerebral Hemorrhage: Mechanisms and Clinical Translation. Front. Immunol. 2022, 13, 844163. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, Q.; Yong, V.W.; Xue, M. EMMPRIN Promotes the Expression of MMP-9 and Exacerbates Neurological Dysfunction in a Mouse Model of Intracerebral Hemorrhage. Neurochem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Yamazawa, T. Chlorogenic acid, a polyphenol in coffee, protects neurons against glutamate neurotoxicity. Life Sci. 2015, 139, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.; Rao, L.J.M. An Outlook on Chlorogenic Acids-Occurrence, Chemistry, Technology, and Biological Activities. Crit. Rev. Food Sci. 2013, 53, 968–984. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients 2016, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana-Galvez, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. Chlorogenic Acid: Recent Advances on Its Dual Role as a Food Additive and a Nutraceutical against Metabolic Syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, H.; Sun, X.L.; Watanabe, M.; Okamoto, M.; Hatano, T. Chlorogenic acid and its metabolite m-coumaric acid evoke neurite outgrowth in hippocampal neuronal cells. Biosci. Biotechnol. Biochem. 2008, 72, 885–888. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Mukherjee, S.; Paliwal, P.; Singh, S.S.; Birla, H.; Singh, S.P.; Krishnamurthy, S.; Patnaik, R. Neuroprotective effect of chlorogenic acid in global cerebral ischemia-reperfusion rat model. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1293–1309. [Google Scholar] [CrossRef]
- Lee, K.; Lee, J.S.; Jang, H.J.; Kim, S.M.; Chang, M.S.; Park, S.H.; Kim, K.S.; Bae, J.; Park, J.W.; Lee, B.; et al. Chlorogenic acid ameliorates brain damage and edema by inhibiting matrix metalloproteinase-2 and 9 in a rat model of focal cerebral ischemia. Eur. J. Pharmacol. 2012, 689, 89–95. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Mun-Bryce, S.; Wesley, M.; Kornfeld, M. Collagenase-induced intracerebral hemorrhage in rats. Stroke 1990, 21, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Manaenko, A.; Ou, Y.B.; Shao, A.W.; Yang, S.X.; Zhang, J.H. Bosutinib Attenuates Inflammation via Inhibiting Salt-Inducible Kinases in Experimental Model of Intracerebral Hemorrhage on Mice. Stroke 2017, 48, 3108–3116. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Schallert, T.; Keep, R.F.; Wu, J.M.; Hoff, J.T.; Xi, G.H. Behavioral tests after intracerebral hemorrhage in the rat. Stroke 2002, 33, 2478–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Han, B.; He, Y.; Li, D.; Ma, X.; Liu, Q.; Hao, J. MicroRNA-132 attenuates neurobehavioral and neuropathological changes associated with intracerebral hemorrhage in mice. Neurochem. Int. 2017, 107, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Q.; Wang, L.; Sun, X.G.; Tang, J.P. Haematoma scavenging in intracerebral haemorrhage: From mechanisms to the clinic. J. Cell Mol. Med. 2018, 22, 768–777. [Google Scholar] [CrossRef]
- Ahn, E.H.; Kim, D.W.; Shin, M.J.; Kwon, S.W.; Kim, Y.N.; Kim, D.S.; Lim, S.S.; Kim, J.; Park, J.; Eum, W.S.; et al. Chlorogenic Acid Improves Neuroprotective Effect of PEP-1-Ribosomal Protein S3 Against Ischemic Insult. Exp. Neurobiol. 2011, 20, 169–175. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Tejada, S.; Setzer, W.N.; Gortzi, O.; Sureda, A.; Braidy, N.; Daglia, M.; Manayi, A.; Nabavi, S.M. Chlorogenic Acid and Mental Diseases: From Chemistry to Medicine. Curr. Neuropharmacol. 2017, 15, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef]
- Jin, R.; Zhong, W.; Liu, S.; Li, G. CD147 as a key mediator of the spleen inflammatory response in mice after focal cerebral ischemia. J. Neuroinflamm. 2019, 16, 198. [Google Scholar] [CrossRef]
- Jin, R.; Xiao, A.Y.; Chen, R.; Granger, D.N.; Li, G. Inhibition of CD147 (Cluster of Differentiation 147) Ameliorates Acute Ischemic Stroke in Mice by Reducing Thromboinflammation. Stroke 2017, 48, 3356–3365. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Verny, M.A.; Besson, C.; Remesy, C.; Scalbert, A. Chlorogenic acid bioavailability largely depends on its metabolism by the gut microflora in rats. J. Nutr. 2003, 133, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Qiao, M.; Xu, H.; Wang, L.; Li, F. Determination of chlorogenic acid in rat plasma by high performance chromatography after peritoneal administration of compound Daqingye injection. Yakugaku Zasshi 2006, 126, 811–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermawati, E.; Arfian, N.; Mustofa, M.; Partadiredja, G. Chlorogenic acid ameliorates memory loss and hippocampal cell death after transient global ischemia. Eur. J. Neurosci. 2020, 51, 651–669. [Google Scholar] [CrossRef]
- Hill, J.W.; Poddar, R.; Thompson, J.F.; Rosenberg, G.A.; Yang, Y. Intranuclear matrix metalloproteinases promote DNA damage and apoptosis induced by oxygen-glucose deprivation in neurons. Neuroscience 2012, 220, 277–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Khan, S.; Liu, Y.; Siddique, R.; Zhang, R.; Yong, V.W.; Xue, M. Gap Junctions and Hemichannels Composed of Connexins and Pannexins Mediate the Secondary Brain Injury Following Intracerebral Hemorrhage. Biology 2021, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, M.; Kaczmarek, L. MMP-9 Inhibition: A Therapeutic Strategy in Ischemic Stroke. Mol. Neurobiol. 2014, 49, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Lee, B.J.; Bu, Y. Protective Effects of Dihydrocaffeic Acid, a Coffee Component Metabolite, on a Focal Cerebral Ischemia Rat Model. Molecules 2015, 20, 11930–11940. [Google Scholar] [CrossRef] [Green Version]
- Biswas, M.H.U.; Almeida, S.; Lopez-Gonzalez, R.; Mao, W.; Zhang, Z.; Karydas, A.; Geschwind, M.D.; Biernat, J.; Mandelkow, E.M.; Futai, K.; et al. MMP-9 and MMP-2 Contribute to Neuronal Cell Death in iPSC Models of Frontotemporal Dementia with MAPT Mutations. Stem Cell Rep. 2016, 7, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Giancotti, F.G.; Ruoslahti, E. Transduction—Integrin signaling. Science 1999, 285, 1028–1032. [Google Scholar] [CrossRef]
- Frisch, S.M.; Screaton, R.A. Anoikis mechanisms. Curr. Opin. Cell Biol. 2001, 13, 555–562. [Google Scholar] [CrossRef]
- Zhang, Y.; Khan, S.; Liu, Y.; Zhang, R.; Li, H.; Wu, G.; Tang, Z.; Xue, M.; Yong, V.W. Modes of Brain Cell Death Following Intracerebral Hemorrhage. Front Cell Neurosci 2022, 16, 799753. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, F.; Li, Z.; Mu, Y.; Yong, V.W.; Xue, M. Neuroprotective Effects of Chlorogenic Acid in a Mouse Model of Intracerebral Hemorrhage Associated with Reduced Extracellular Matrix Metalloproteinase Inducer. Biomolecules 2022, 12, 1020. https://doi.org/10.3390/biom12081020
Liu Y, Wang F, Li Z, Mu Y, Yong VW, Xue M. Neuroprotective Effects of Chlorogenic Acid in a Mouse Model of Intracerebral Hemorrhage Associated with Reduced Extracellular Matrix Metalloproteinase Inducer. Biomolecules. 2022; 12(8):1020. https://doi.org/10.3390/biom12081020
Chicago/Turabian StyleLiu, Yang, Fei Wang, Zhe Li, Yanling Mu, Voon Wee Yong, and Mengzhou Xue. 2022. "Neuroprotective Effects of Chlorogenic Acid in a Mouse Model of Intracerebral Hemorrhage Associated with Reduced Extracellular Matrix Metalloproteinase Inducer" Biomolecules 12, no. 8: 1020. https://doi.org/10.3390/biom12081020