1. Introduction
Hearing loss (HL) is the most prevalent sensorineural deficit, affecting about 20% of the global population (
https://www.who.int/health-topics/hearing-loss, accessed on 24 May 2022), with nearly half due to genetic causes [
1]. It is estimated that 1 in 500 newborns present with congenital HL, which profoundly affects language acquisition and consequently, social development [
2]. Current standards of care for HL include hearing aids for people with diminished auditory function or cochlear implants, indicated for severe to profound HL. Unfortunately, there are no biological treatments available for HL and no interventions that restore auditory capacity to normal levels. During the last decade, many studies have investigated gene therapy strategies in animal models as treatments for genetic HL [
3,
4]. However, none of these experimental therapies have advanced to the clinic for treatment of humans.
The auditory sensory epithelium is contained within the cochlea, the bony spiral structure of the inner ear, which can be accessed for therapeutic intervention via the round window membrane. Cochlear hair cells are organized in the organ of Corti within a tonotopic gradient: high frequencies are sensed at the base and low frequencies at the apex. The cochlea includes two types of hair cells: inner hair cells (IHCs), which convert mechanical signals into electrical signals and transmit them to contacting afferent neurons and on to the brain; and outer hair cells (OHCs), which serve as cochlear amplifiers, necessary for signal amplification and tuning.
To develop inner-ear gene therapy for genetic HL, our lab has focused on gene replacement for transmembrane channel-1 (
TMC1) [
5,
6,
7]. TMC1 forms the ion channel at the core of the mechanosensory transduction complex in auditory and vestibular hair cells [
8,
9]. Mutations in
TMC1 cause non-syndromic HL, which can be dominant (DFNA36) or recessive (DFNB7/11) [
10,
11]. Depending on the population, mutations in
TMC1 represent 3–8% of genetic HL [
12]. Mice that lack
Tmc1 (
Tmc1Δ/Δ) are profoundly deaf due to lack of sensory transduction and present with degeneration of cochlear hair cells beginning around four weeks of age [
13].
In prior work from our lab, we used an AAV2/1 viral capsid to deliver mouse
Tmc1 cDNA sequence into
Tmc1Δ/Δ mice and recovered their hearing capacity, albeit with high thresholds and only in the low frequency range [
5]. The limited recovery was the result of viral transduction in IHCs, but not OHCs [
5]. Even though the gene therapy strategy evoked a partial restoration of auditory thresholds in a limited hearing range, it demonstrated that hair cells can be targeted by AAV viruses and that the auditory function can be recovered in mice with profound genetic HL. In order to improve the extent of the recovery, non-conventional viral capsids were screened. Our lab discovered that the Anc80L065 capsid was able transduce both IHCs and OHCs in vivo [
14]. Delivery of
Tmc1 using this capsid extended the level of auditory recovery at low and middle frequencies [
6]. Recently, we showed that gene therapy for
Tmc1 using the synthetic AAV9-PHP.B capsid [
15] further improved the recovery of auditory thresholds in
Tmc1Δ/Δ mice [
7]. Importantly, the AAV9-PHP.B capsid transduced IHCs and OHCs efficiently along the entire tonotopic gradient [
16], which is required to enable broad-spectrum auditory recovery.
While the prior work focused on gene therapy recovery in
Tmc1Δ/Δ mice, there are no human
TMC1 mutations equivalent to that of
Tmc1Δ/Δ mice. Over 65 human
TMC1 mutations have been described that cause DFNB7/11. Some mutations are associated with severe-to-profound congenital deafness and result from premature stop mutations, frame shift mutations, and other functional null mutations. In contrast, several point mutations that yield single amino acid substitutions cause milder phenotypes with moderate-to-severe hearing loss during the first decade of life [
17]. For example, the human
TMC1 mutation p.N199I (c.596A > T) causes progressive moderate-to-severe hearing loss that develops during childhood [
17]. Thus, we wondered whether the milder hearing loss phenotype in patients with p.N199I, and other similar mutations, may provide an expanded capacity for therapeutic intervention using a gene replacement strategy.
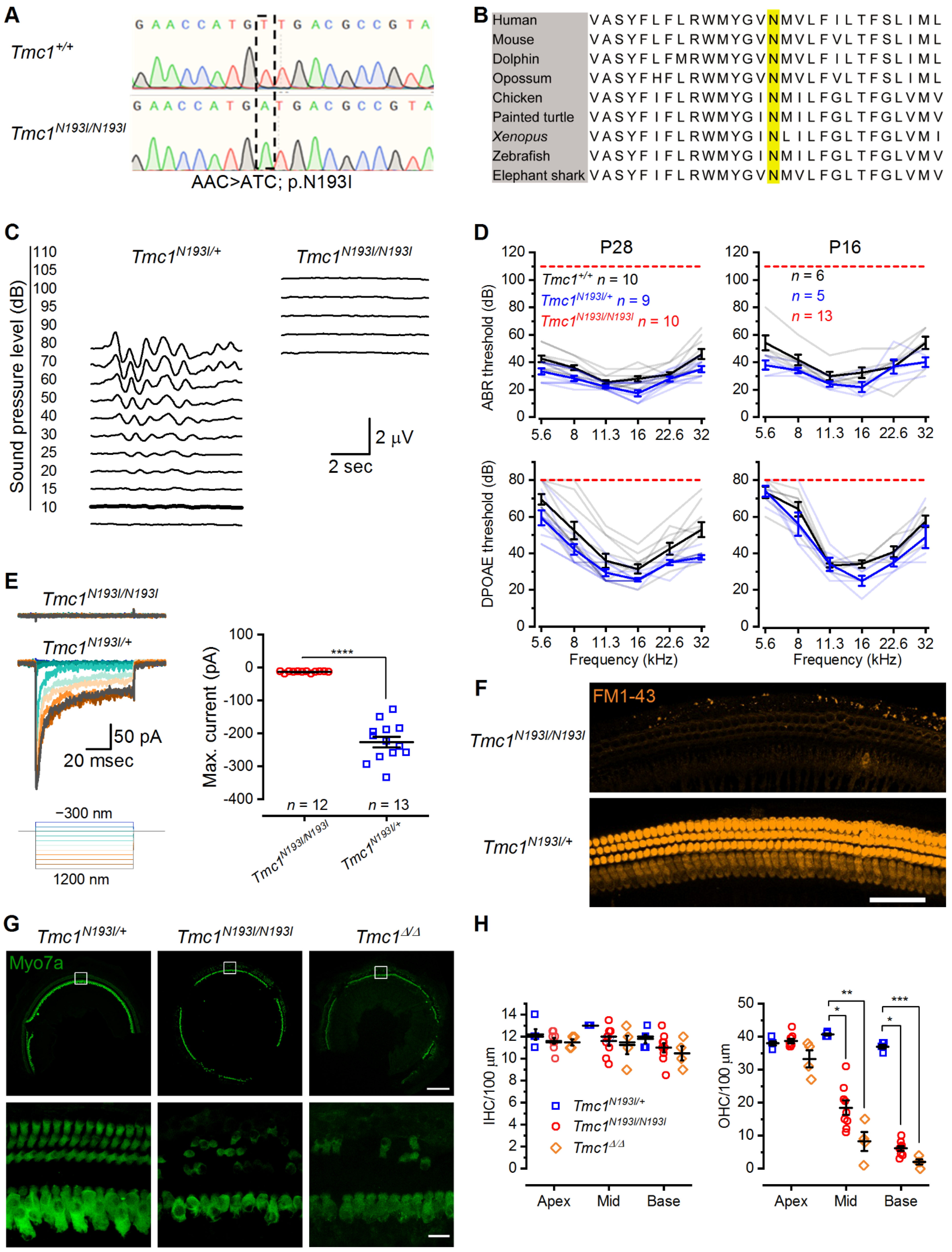
We sought to study the effect of the missense p.N199I mutation by generating a humanized murine model harboring the homologous mutation in Tmc1 (p.N193I) and developed a gene therapy strategy using gene replacement through AAV delivery. For this study, 143 mice were injected with one of three viral constructs at three different doses, and 117 mice were used as uninjected controls. We performed over 420 measurements of auditory function on the 260 mice in the study. We found that mice homozygous for the Tmc1 p.N193I mutation have profound HL due to loss of mechanosensory transduction in cochlear hair cells. Mutant mice injected with AAV9-PHP.B carrying a functional copy of human TMC1 (hTMC1) successfully recovered auditory capacity. Using a CB6 promoter to drive the expression of hTMC1 yielded auditory recovery that spanned the frequency spectrum. Additionally, the optimized gene therapy strategy efficiently recovered hearing capacity in Tmc1Δ/Δ mice, though to a lesser extent at high frequencies. We report unprecedented recovery of auditory thresholds using AAV9-PHP.B-optimized constructs encoding hTMC1 in a humanized mouse model for genetic HL. These results demonstrate that early and efficient gene therapy intervention can restore auditory function in mice, suggesting potential translation for treatment of DFNB7/11 patients.
2. Materials and Methods
2.1. Mice
All procedures met NIH guidelines for the care and use of laboratory animals and were approved by the Institutional Animal Care and Use Committee (protocols #18-01-3610R and #20-02-4149R) at Boston Children’s Hospital (BCH).
We used wild-type C57BL/6J mice (Jackson Laboratories, Bar Harbor, ME, USA) with corrected
Ahl allele,
Tmc1 mice with genotype
Tmc1Δ/Δ; Tmc2+/+ on a C57BL/6J background [
13], and knock-in mice with genotype
Tmc1N193I/N193I; Tmc2+/+ or
Tmc1N193I/N193I; Tmc2Δ/Δon a C57BL/6J background with the
Ahl allele corrected. Mouse line
Tmc1N193I/N193I; Tmc2+/+ was generated at the Mouse Gene Manipulation Core of the F.M. Kirby Neurobiology Center at BCH using the CRISPR/Cas9 system. The p.N193I (A
AC > A
TC) substitution in the
Tmc1 gene was introduced by disruption of the endogenous gene using SpCas9 (IDT, Coralville, Iowa, USA, Cat#181058) with a gRNA that specifically targeted the
Tmc1 sequence (5′-CAG GTGGATGTACGGCGTCAACATGG-3′) and homologous recombination using a template that spanned the whole exon (in upper case), part of the adjacent introns (in lower case) and contained the point mutation (in bold) (5′-ttggaagtcagggcttacCTCCGGTAACATGATGAGGCTGAAGGTCAACACAAAGAGAACCATGATGACGCCGTACATCCATCTGAGGAACAGGAAGTACGAGGCCACTGAGGAACCAAACTGACctgttacgaagaaaattaaaagactaaaggagagtcaaaacaacaacagcaccaatgaaaaac-3′).
The mutation was confirmed by Sanger sequencing analysis of a PCR product obtained from genomic DNA extracted from toe clips using lysis reagent (Viagen Biotech, Los Angeles, CA, USA) supplemented with Proteinase K (New England Biolabs, Ipswich, MA, USA) and genotyping primers (F: 5′-ATTGGGGTGGAAACCGATACGA-3′, R: 5′-TCTCCTGAGAGAAAGAAGAGCGA-3′). Amplification conditions were as follows: 98 °C for 30 s; 42 × (98 °C for 10 s; 66 °C for 30 s; 72 °C for 50 s); 72 °C for 2 min. PCR products were loaded into a 1.2% agarose gel stained with SYBR Safe (Invitrogen, Waltham, MA, USA) for electrophoresis. The amplicon band was identified in an iBright CL1500 Imaging System (Invitrogen) and separated from the rest of the gel for further purification using the Monarch DNA Gel Extraction Kit (New England Biolabs).
Both males and females were used for experiments in approximately equal proportion. All mice were kept in a 12 h light/12 h dark cycle with accessibility to food and water ad libitum.
2.2. Plasmids and AAV Production
Human TMC1 (NM_138691.3) codon-optimized sequence (hTMC1) was cloned into AAV2 plasmids downstream of a cytomegalovirus (CMV) or chicken beta-actin-derived (CB6) promoter and followed by a woodchuck hepatitis post-transcriptional regulatory element (WPRE), unless otherwise indicated. Vector was produced by triple transfection of suspension HEK293 cells grown in Expi293 media (Gibco, Waltham, MA, USA). Briefly, cells at ~2 × 106 cells/mL were transfected with pALDX-80 helper plasmid, AAV2/AAV9PHP.B rep/cap plasmid, and hTMC1 plasmid using PEIpro (Polyplus, Illkirch-Graffenstaden, France) formulated in OptiPRO SFM (Gibco). Seventy-two hours after transfection, cells were harvested by addition of Benzonase and 10× lysis buffer (500 mM Tris, 20 mM Magnesium Chloride, 10% Polysorbate 20, pH 8.0) followed by addition of 5 M Sodium Chloride to a final concentration of 500 mM. Lysates were clarified by centrifugation and stored at −80 °C until purification.
Immediately prior to chromatography, lysates were filtered across a 0.45/0.22 µm polish filter (Millipore, St. Louis, MO, USA) and then passed over POROS CaptureSelect AAV9 (ThermoFisher, Waltham, MA, USA) affinity resin using an AKTA Pure instrument (Cytiva Life Sciences, Marlborough, MA, USA). Elution fractions with A280 values > 100 mAU were pooled and immediately neutralized with 10% volume of neutralization buffer (500 mM Bis-Tris Propane, 200 mM NaCl, 1%
w/
v Sucrose, 0.001% Poloxamer 188, pH 9.0). Following purification, the pooled affinity neutralized eluate was buffer-exchanged into final formulation buffer (dPBS, 5%
w/
v Sorbitol, 0.001%
w/
v Poloxamer 188, pH 7.0). Titer was determined using ddPCR as previously described [
18]. The following titers of genome-containing particles were used:
AAV2/9-PHP.B-CMV-hTMC1-WPRE: 3.26 × 10
13 gc/mL or 6.3 × 10
13 gc/mL;
AAV2/9-PHP.B-CB6-hTMC1 No WPRE: 3.14 × 10
12 gc/mL;
AAV2/9-PHP.B-CB6-hTMC1-WPRE: 3.14 × 10
12 gc/mL, 1.57 × 10
13 gc/mL, 3.14–4.30 10
13 gc/mL or 7.16 10
13 gc/mL. Virus aliquots were stored at −80 °C.
2.3. Animal Surgery
All viral inner-ear injections were performed according the Institutional Animal Care and Use Committee at BCH protocols (#00001240 and #20-02-4149R) and were done via utricle injection as previously described [
16]. In brief, P1-2 mice were anesthetized with hypothermia through 3 min of exposure to ice water. During the surgery (10–15 min), mice were kept on an ice pad. Using a stereo microscope (Stemi 2000, Zeiss, Oberkochen, Germany) for visualization, a small postauricular incision was made to expose the cochlea bulla and semicircular canals surrounding the utricle. After puncturing the temporal bone, a glass micropipette was inserted into the puncture to manually inject 1–1.2 µL of AAV at a constant rate. Only left ears were used for injections. Following the procedure, mice were placed on a heating pad until fully recovered, and standard postoperative care was applied
2.4. Electrophysiology
Temporal bones were collected from P6–7 mice in Minimum Essential Media (Gibco). Overlying bone was removed with forceps and a needle until the cochlea could be accessed at the modiolus and separated from the vestibular portion of the inner ear. The organ of Corti was isolated from the modiolus and spiral ligament and anchored to a glass slide with a stainless-steel pin (Fine Science Tools, Foster City, CA, USA). After 1–2 days of incubation at 37 °C in minimum essential media supplemented with 10% fetal bovine serum (Life Technologies, Waltham, MA, USA) and 100 µg/mL ampicillin (Sigma-Aldrich, St. Louis, MO), the tectorial membrane was removed, and the tissue was placed under an Axioskop FS (Carl Zeiss) upright microscope and visualized using a 63× water-immersion objective. Patch clamp recordings in whole-cell configuration of inner hair cells from the basal portion of the cochlea were performed as previously described in Pan et al. (2013, 2018). Briefly, tissues were kept at room temperature and submerged in an artificial perilymph solution containing (in mM): 137 NaCl, 10 HEPES, 5.8 KCl, 5.6 d-glucose, 1.3 CaCl2, 0.9 MgCl2, 0.7 NaH2PO4, and vitamins (1:100) and amino acids (1:50) (ThermoFisher) at pH 7.4 and 311 mOsmol/kg. Recording electrodes (3–5 MΩ) were pulled from R6 capillary glass (King Precision Glass, Claremont, CA, USA) and filled with an internal solution containing (in mM): 140 CsCl, 5 HEPES, 5 EGTA-KOH, 3.5 MgCl2, 2.5 Na2ATP, and 0.1 CaCl2 at pH 7.4 and 284 mOsm/kg. Sensory transduction currents were recorded at a holding potential of −84 mV using an Axopatch 200B (Molecular Devices, San Jose, CA, USA). Currents were low-pass filtered at 2–5 kHz with a Bessel filter, sampled at 20 kHz with a 12-bit acquisition board (Digidata 1322A), recorded with pCLAMP software (Molecular Devices), and corrected for a 4 mV liquid junction potential. Hair cell bundles were deflected using a 4–5 µm tip diameter stiff glass probe mounted on a PICMA chip piezo actuator (Physik Instruments, Karlsruhe, Germany). The actuator was driven by an LPZT amplifier (Physik Instruments) and filtered with an 8-pole Bessel filter at 40 kHz to eliminate residual pipette resonance. Data were analyzed with Clampfit and Origin (OriginLab, Northampton, MA, USA).
2.5. FM1-43 Uptake
All experiments with membrane dye FM1-43 (Invitrogen) were performed at room temperature. FM1-43 stock solution (10 mM) was diluted to a working concentration (5 µM) in minimum essential media (Gibco). Mouse temporal bones were harvested at age P6, followed by microdissection of the organ of Corti. Freshly prepared FM1-43 was added to the culture for 10 s and then washed twice (5 min each wash) with minimum essential media. Live tissues were imaged with a 40× water immersion objective in an LSM 800 (Carl Zeiss) microscope.
2.6. Tissue Preparation and Confocal Immunofluorescence
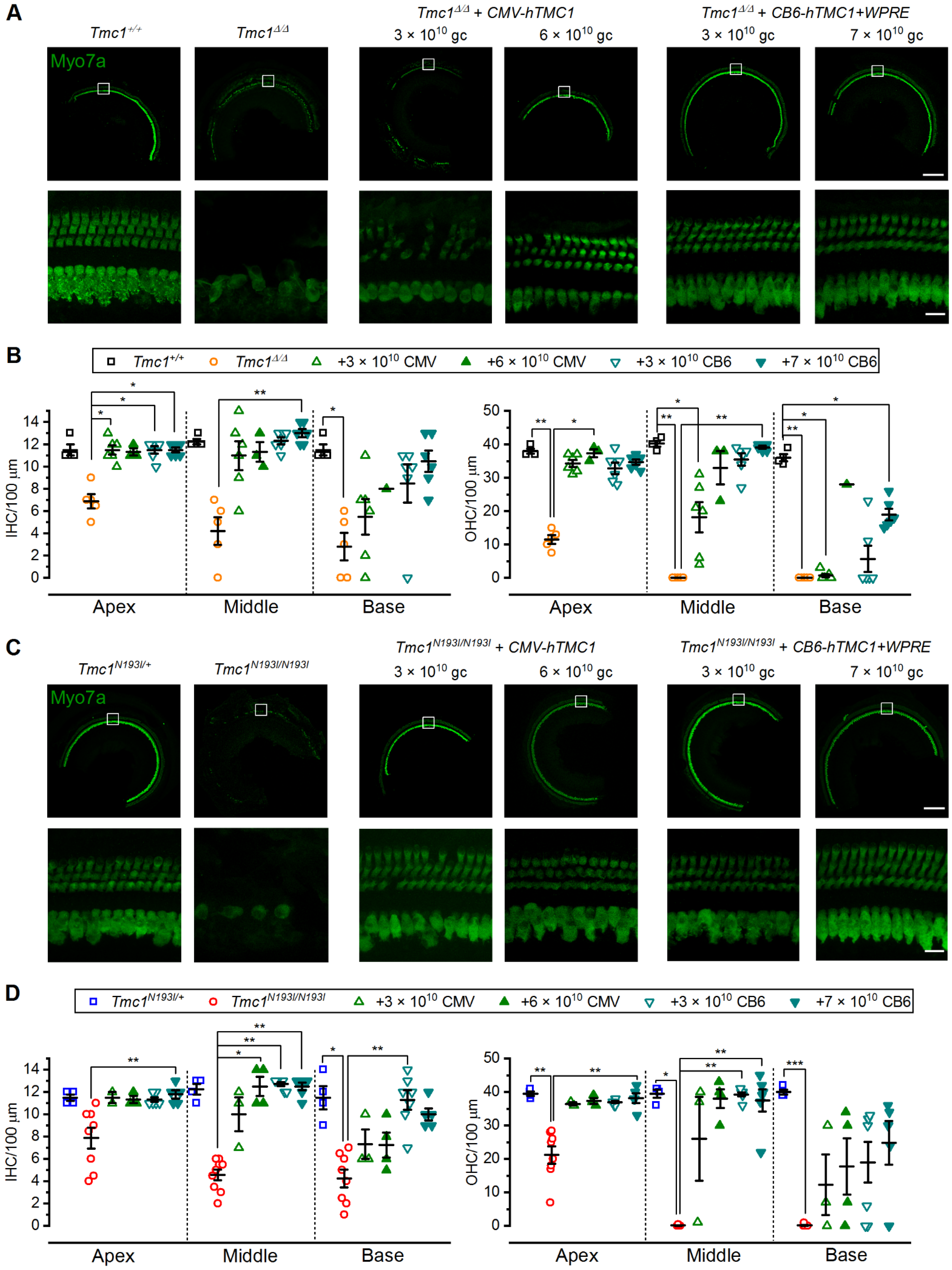
Euthanization of 4- or 12-week-old mice was conducted via CO2 inhalation. Temporal bones were harvested, punctured at the round and oval windows and helicotrema, and fixed in 4% paraformaldehyde for 1 h at room temperature. Tissues were then decalcified in 120 mM EDTA for 16–24 h. Cochleas were sectioned into apical, middle, and basal portions. The organ of Corti was isolated and prepared for whole-mount processing by removal of the lateral wall, spiral limbus, and tectorial membrane. Tissues were permeabilized for 1 h in 0.25% Triton X-100, blocked for 1 h in 2.5% normal donkey serum, and stained at 4 °C overnight with rabbit anti-myosin 7a primary antibody (1:500 Proteus Biosciences, Ramona, CA, USA #25-6790). After washing with PBS, samples were incubated for 3–4 h with fluorophore-conjugated donkey anti-rabbit secondary antibody (1:400 Alexa Fluor 647: Thermo Fisher #A31573) and fluorophore-conjugated phalloidin (1:400 Alexa Fluor Plus 405: Thermo Fisher #A30104). Tissues were then mounted on a glass coverslip with Vectashield mounting medium (Vector Laboratories, Burlingame, CA, USA). Confocal imaging was performed using 10× air and 63× oil-immersion objectives with an LSM 800 (Carl Zeiss) microscope. Maximum intensity projection images were generated in ImageJ. Hair cell count analysis was performed manually.
2.7. Auditory Brainstem Response (ABR) Measurement
P16, 4-, 8-, or 12-week-old mice were anesthetized through intraperitoneal injection of a ketamine (100 mg/kg)/xylazine (15 mg/kg) cocktail. Mice were placed on a heating pad in a sound-proof chamber. The external auditory meatus was exposed through resection of the overlying skin and cartilage, and a custom speaker/microphone apparatus (EPL PXI Systems) was positioned directly above the entrance of the ear canal to deliver and record acoustic signals. Sound pressure at this location was calibrated for all stimulus frequencies before each test. Acoustic stimuli were delivered as 5 ms tone bursts at half-octave steps from 5.6 to 32 kHz and at 5 to 10 dB steps from 10 to 110 dB sound-pressure level (SPL) or until threshold was identified by the presence of a reproducible ABR waveform with a detectable peak 1. At each SPL, 512 responses to an alternating polarity stimulus were averaged. Electrical activity was recorded through subcutaneous needle electrodes behind the pinna (active electrode), at the vertex (reference), and at the hind leg (ground). ABR responses were amplified (10,000×), filtered (0.3 to 10 kHz), and digitized with LabVIEW software (Eaton-Peabody Laboratories). Electrode voltages and acoustic stimuli were sampled every 40 µs using a digital input-output board (National Instruments, Austin, TX, USA). Data were then plotted in Origin (OriginLab).
2.8. Distortion Product Otoacoustic Emission (DPOAE) Measurement
DPOAE and ABR data were collected consecutively under identical conditions. DPOAE at 2f1-f2 were measured at half-octave f2 steps from 5.6 to 45.2 kHz and at 5 dB f2 steps from 10 to 80 dB SPL (f2/f1 = 1.2, L1–L2 = 10 dB). Threshold was determined after spectral averaging as the lowest L2 value to produce a DPOAE distinguishable from the surrounding noise floor and maintained at higher dB SPLs. Data were plotted in Origin (OriginLab).
2.9. RNA and DNA Isolation
Twelve-week-old animals were euthanized by cervical dislocation. The following organs were harvested, flash frozen immediately in liquid nitrogen and stored at −80 °C: injected (left) inner ear, contralateral (right) inner ear, brain and liver. RNA and DNA were isolated from frozen mouse cochlea using TRI-Reagent (Zymo Research, Irvine, CA, USA) and Direct-zol DNA/RNA Miniprep kits (Zymo Research). Briefly, frozen mouse cochleas were placed in a tube with a stainless-steel ball and TRI-Reagent. Samples were subjected to bead beating twice using a TissueLyser II (Qiagen, Hilden, Germany). Chloroform was added to samples and centrifuged to separate RNA, DNA, and protein. The top aqueous layer of RNA was carefully removed, followed by the interphase layer of DNA. Each layer was transferred to their respective tubes and diluted with 100% ethanol 1:1. RNA and DNA were subsequently purified through individual columns following the DNA/RNA Miniprep purification protocols. RNA and DNA from mouse brain and liver were purified using the All-Prep DNA/RNA kit (Qiagen) according to the manufacturer’s protocol. Peripheral tissues were lysed similar to cochleas using the kits lysis buffer instead of TRI-Reagent.
2.10. Quantitative PCR
RNA and DNA were quantified according to the manufacturer’s protocols using the Quant-iT RNA Assay Kit (ThermoFisher) and the Quant-iT dsDNA Assay Kit (ThermoFisher) respectively; 100 ng of RNA underwent cDNA synthesis using the RT2 Easy First Strand Kit (Qiagen) according to the manufacturer’s protocol.
To assess vector genomes and transgene expression, qPCR was performed on gDNA and cDNA using PrimeTime Gene Expression 2× Master Mix (IDT), a fluorescently labeled 5′ nuclease probe (IDT), and hTMC1 opt primers with the following sequences: forward primer, 5′-GATCAGGATGGTCACGTATG-3′ and reverse primer 5′-GCTGGG AGACAATGGTAG-3′ and CFX-96 (BioRad, Hercules, CA, USA) instruments. A standard curve was prepared using a gBlock standard established over a six-fold dilution series from 5E1–5E6 copies per reaction. Samples were analyzed by the absolute quantification method against the standard curve using CFX Maestro software (BioRad). Transgene expression was normalized and represented as copies/1 µg RNA. Vector genome copies were normalized to amount and reported as copies/1 µg DNA.
2.11. Statistical Analyses
Normality tests were performed on the data prior to running statistical analysis. All figures present datasets that do not show normal distribution, and therefore, non-parametric tests were performed. For comparing two different groups (i.e., electrophysiology recording), Mann–Whitney test was used. For comparing datasets with more than two groups (i.e., hair cell counts), Kruskal-Wallis followed by Dunn test for multiple comparisons was applied. A
p value equal to or less than 0.05 was considered significant. Statistical analysis was performed using Origin (OriginLab). Individual traces and mean values ± S.E.M. are presented in all figures. All values and statistics are presented in
Supplemental Tables S1–S5.
4. Discussion
In the recent years, significant improvements in gene therapy as a treatment for HL have been made in animal models [
3,
4,
23]. The discovery of viral capsids that target hair cells with higher efficiency has enabled specific delivery of genes involved in sound perception. However, further optimization of viral constructs in animal models of genetic HL is needed prior to translation to the clinic.
Due to a lack of appropriate models, gene therapy for recessive DFNB7/11 HL has not been tested in a mouse model that bears a human
TMC1 mutation. Prior work in
Tmc1Δ/Δ mice provided important proof-of-concept data [
5,
6,
7], but as there are no equivalent mutations in human
TMC1, questions remain regarding relevance to the human condition. As such, we sought to develop a mouse model bearing a recessive human
TMC1 mutation and focused on the p.N199I mutation [
17]. We reasoned that since patients carrying this mutation have a milder form of HL, there may be an expanded capacity for clinical intervention. Patients homozygous for the p.N199I mutation present with moderate HL at 3–5 years of age, which progresses over the next decade to more severe HL. Given the phenotype and the role of TMC1 as the hair cell sensory transduction channel, we hypothesized that the p.N199I mutation may yield a hypofunctional ion channel which may be sufficient to allow for some auditory function and survival of auditory hair cells during the first decade of life. However, to our surprise, when we generated and validated the mouse model bearing the equivalent mutation, p.N193I, we discovered that the homozygous mice were profoundly deaf at the earliest time point tested, P16. Furthermore, the mice lacked sensory transduction entirely, suggesting that, contrary to our hypothesis, the p.N193I version of TMC1 was not hypofunctional; rather, the single amino acid substitution caused a complete loss of function. These unanticipated results raise several important and interesting questions.
Given that TMC1 has the same function in mice and humans and both sequences are highly conserved [
27], we expect that the effect of the mutation on protein function is the same for both species. Therefore, if the p.N199I mutation causes profound loss of TMC1 function, how is it that humans bearing this mutation retain some residual auditory function? We reasoned that a compensatory mechanism that overcomes the loss of TMC1 function must exist in humans during the first decade of life. While the identity of this compensatory mechanism remains unknown, we suspect TMC2 may be playing an expanded or compensatory role in human cochlear hair cells. In mouse cochlear hair cells, TMC2 expression rises during the first postnatal week and is sufficient to support sensory transduction in inner and outer hair cells [
8,
13]. However, TMC2 expression begins to fall at the end of the first postnatal week while TMC1 expression begins to rise [
13]. While the temporal expression patterns of TMC1 and TMC2 in human cochlear hair cells is unknown, perhaps heterochronic differences between mouse and human cochlear development may result in a longer or compensatory expression profile for human TMC2. If so, the residual auditory function in humans with p.N199I mutations may be a consequence of TMC2 function.
Regardless of the underlying cause of the residual hearing function in patients with p.N199I mutations, the presence of any auditory function implies there must be some surviving and functional hair cells during the first decade of life. As such, we suspect that these hair cells may be viable targets for gene therapy intervention and may provide an expanded capacity for auditory recovery. To investigate this hypothesis, we tested three viral constructs in
Tmc1N193I/N193I mice and in the
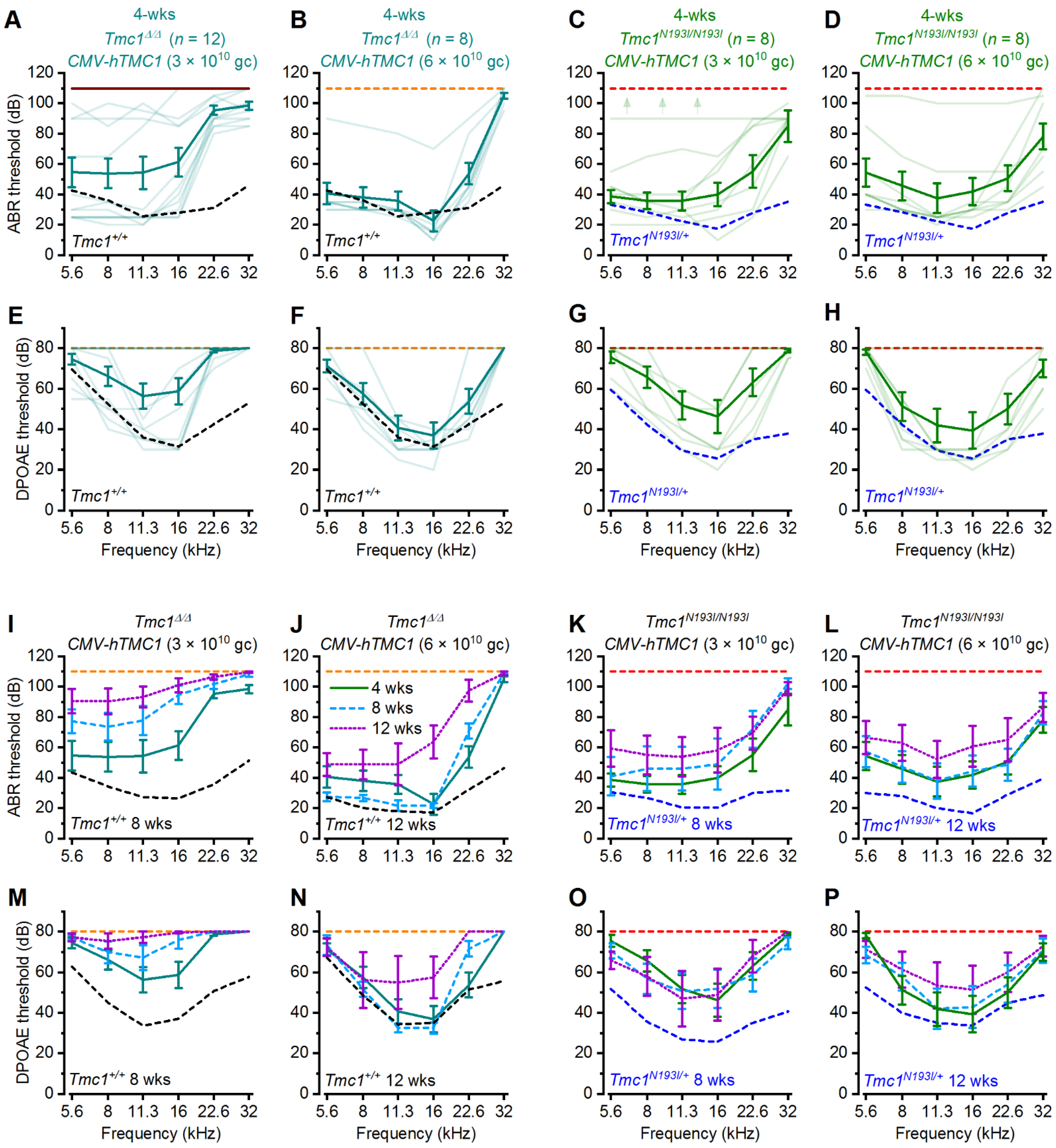
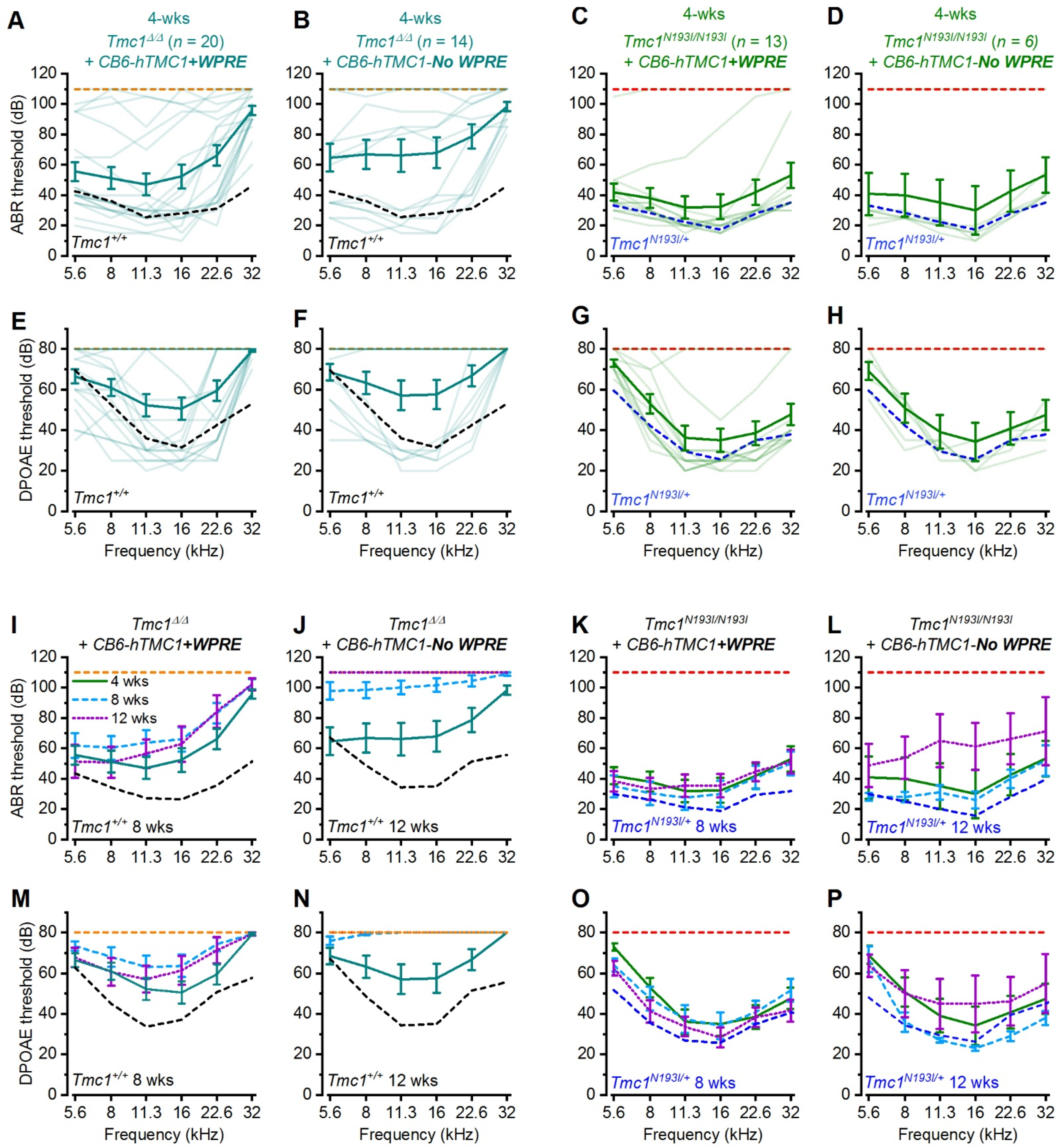
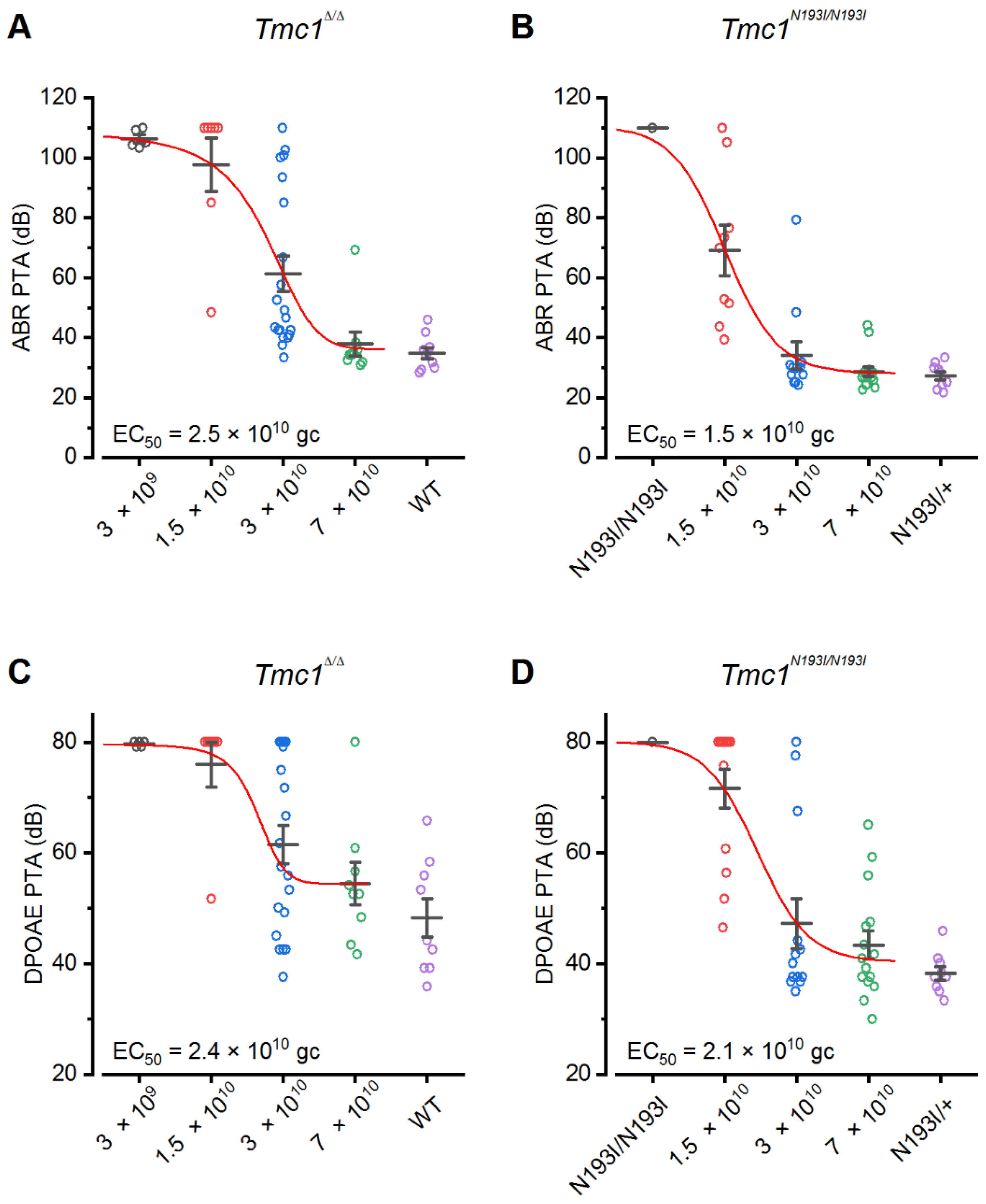
Tmc1Δ/Δ model. First, we were interested to determine if a codon optimized version of human TMC1 was capable of restoring auditory function in mice. Indeed, in both lines, we observed substantial recovery of ABR and DPOAE thresholds and enhanced survival of auditory hair cells, confirming that the codon-optimized human coding sequence could substitute for the mouse sequence. Next, we investigated a modified chicken beta-actin-promoter,
CB6, which is known to drive stable expression in a variety of cell types [
24]. In addition, we investigated the necessity of the post-transcriptional regulatory element,
WPRE, for the durability of auditory recovery. We found that together, the
CB6 promoter and the
WPRE element enhanced the auditory recovery and promoted the durability of recovery in both animal models. Importantly, there was a remarkable level of auditory recovery in the
Tmc1N193I/N193I mice, with ABR and DPOAE thresholds that were indistinguishable from wild-type thresholds across the entire auditory spectrum for 10 of 13 mice injected with the AAV9-PHP.B
-CB6-hTMC1-WPRE construct. Furthermore, the recovery was durable up to 12 weeks post-injection. Injected
Tmc1Δ/Δ mice also had remarkable and durable recovery with the same viral construct at a higher dose, but the recovery was reduced at the high frequency end of the spectrum. We did notice a decline in auditory function in both p.N193I mice and
Tmc1Δ/Δ mice injected with the construct lacking the
WPRE element, underscoring the necessity of the
WPRE sequence for durable recovery of function. When the same viral construct, AAV9-PHP.B
-CB6-hTMC1-WPRE, was injected into wild-type mice, we found no loss of auditory function in eight of nine mice. In the one mouse with elevated thresholds, we suspect the injection procedure itself may have damaged the middle ear structures, as histological analysis revealed a full complement of surviving hair cells in the injected ear.
Due to age-related decline in viral tropism for mouse outer hair cells (Lee et al., 2020), we did not explore viral injection beyond the first postnatal week. However, we note that previous work in adult non-human primates showed that the AAV9-PHP.B viral capsid, with a CBA promoter and a WPRE sequence, was capable of transfecting and driving the expression of a reporter transgene in IHCs and OHCs [
28]. Interestingly, the viral titers that were effective in targeting hair cells along the different tonotopic regions of the primate cochlea were similar to those found to be effective for mice in this study (3.5 and 7 × 10
11 dose, injecting 10 µL) [
28]. Recently, van Beelen et al. [
29] showed that AAV9-PHP.B viral capsids carrying a CMV promoter and the GFP coding sequence can transduce and drive expression of GFP in fetal and adult human hair cells in
ex vivo explants of biopsied inner-ear tissue. These results suggest that the AAV9-PHP.B-
CB6-hTMC1 + WPRE could efficiently target hair cells and drive expression of
hTMC1 in non-human primate hair cells, and perhaps ultimately in humans as well.
In our mouse studies, biodistribution analysis revealed viral DNA and RNA in the contralateral ear, brain and liver, suggesting that in neonatal mice, some vector does escape the injected ear. However, as no overt consequences were noted in these animals, we conclude that systemic biodistribution of the viral construct does not cause noticeable toxicity in mice.
The difference in the response to viral gene therapy between the
Tmc1N193I/N193I mice and
Tmc1Δ/Δ models raises important questions. Why did the
Tmc1N193I/N193I mice respond with complete recovery of auditory function, whereas the knock-out model did not? Since the knock-out model lacks expression of the full-length TMC1 protein, we suspect the consequences of this mutation may be more severe, leading to collapse of the hair cell transduction apparatus, more rapid hair cell degeneration, and somewhat diminished ability to respond to gene therapy intervention. On the other hand, the
Tmc1N193I/N193I mice generate a full-length, albeit non-functional, TMC1 protein. The presence of the protein, even if non-functional, may provide structural support for the transduction complex and help stabilize the apparatus. Alternatively, the mutation may indirectly disrupt the ion channel pore but may leave other TMC1 functions intact. Since the location of N193 is in the first transmembrane domain, the Alpha-fold structure predicts that this residue is within range to interact with Q581 in the seventh transmembrane domain, which lines the channel pore [
9,
19]. Since the polar side chains are facing each other at 2.8–3.4 Å, mutation at the N193 site may destabilize the pore region but leave other regions of TMC1 unaffected.
Although the Tmc1N193I/N193I mice did not mimic the milder and progressive auditory phenotype seen in humans with p.N199I mutations, the remarkable gene therapy recovery in the p.N193I model may suggest that the recovery expected in the presence of the milder human phenotype may be equal to or greater than the recovery documented for mice. Furthermore, we wonder whether this level of recovery may be representative not only of the response expected for p.N199I patients, but perhaps more broadly for all recessive TMC1 point mutations that yield a full-length but non-functional TMC1 protein. More than half of recessive mutations in human TMC1 (40/70) are single amino acid substitutions, raising the possibility that patients harboring these mutations may have an expanded capacity to benefit from TMC1 gene therapy.
The Tmc1Δ/Δ mice also had remarkable auditory recovery in the low-frequency range. We suggest these mice may be a better model of functional null mutations in human TMC1 such as frame shifts, premature stop codons, and so forth. In conclusion, we suggest that the AAV9-PHP.B-CB6-hTMC1-WPRE vector may be well-suited as a gene therapy reagent for treatment of patients with all forms of recessive TMC1 hearing loss.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




