
Mitochondria Transfer from Adipose Stem Cells Improves the Developmental Potential of Cryopreserved Oocytes

 , ,
, , 
Abstract
:1. Introduction
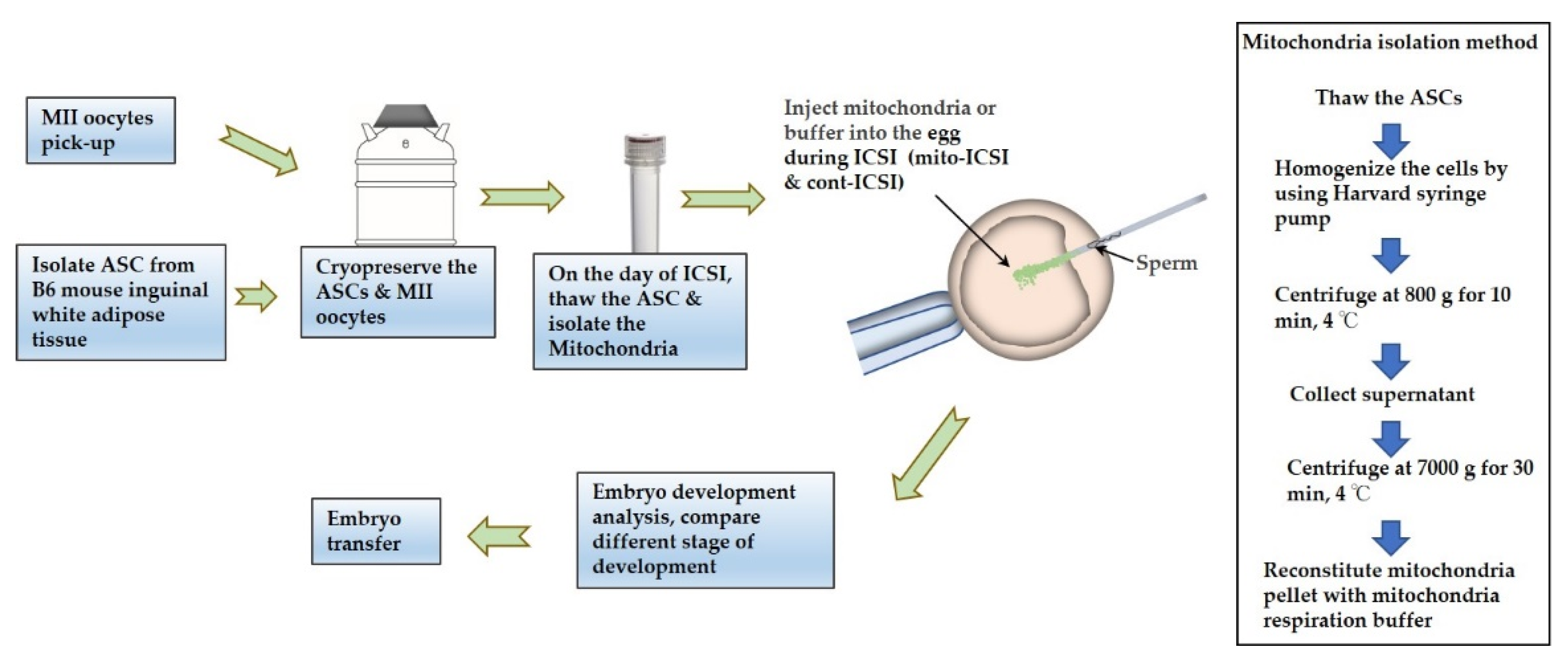
2. Materials and Methods
2.1. Animals
2.2. Ovarian Super-Stimulation, Oocyte Collection
2.3. Vitrification of Oocytes
2.4. Warming of Oocytes
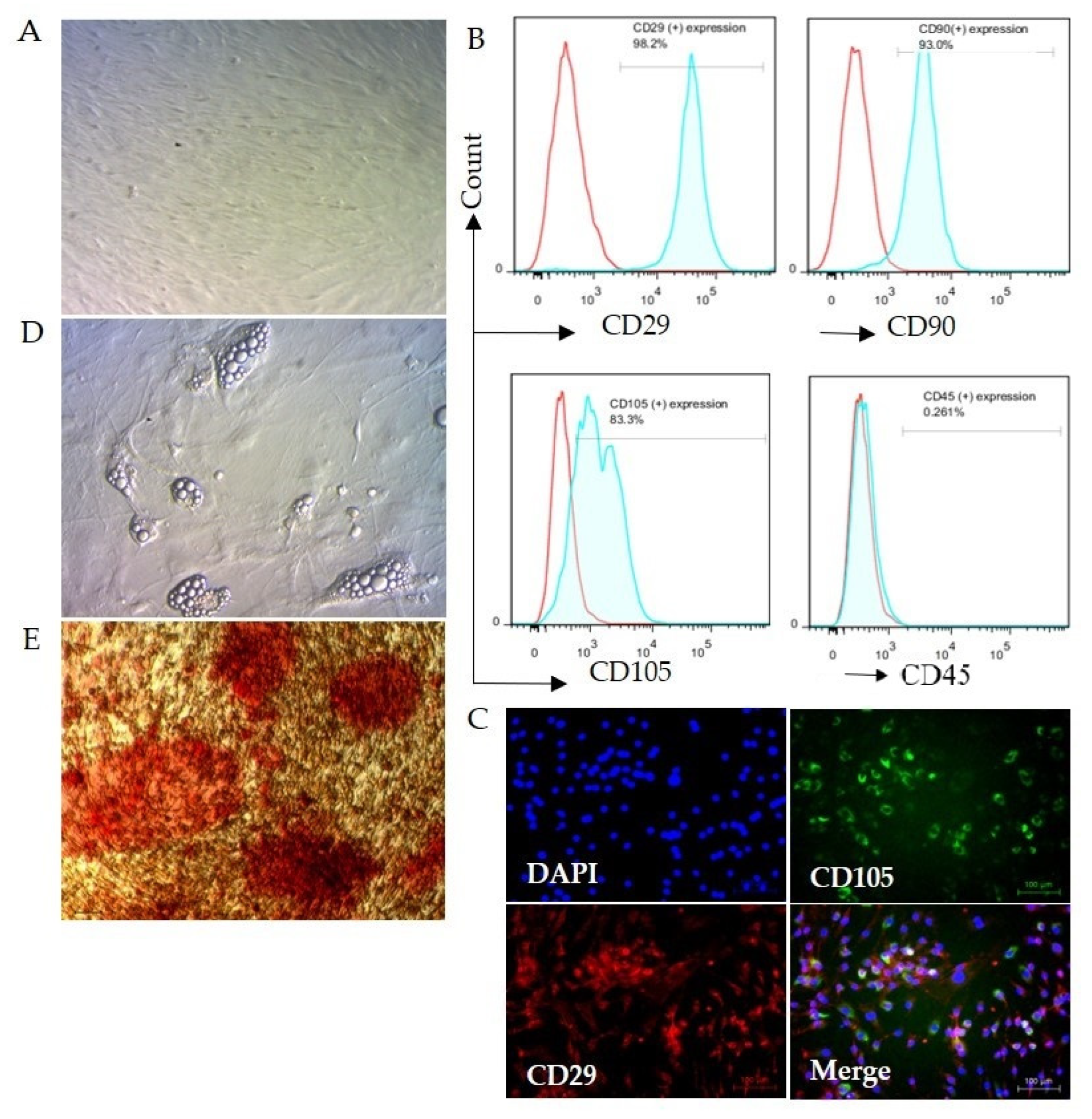
2.5. Cell Isolation, Culture, and Confirmation
2.6. Analysis of the Specificity of ASC
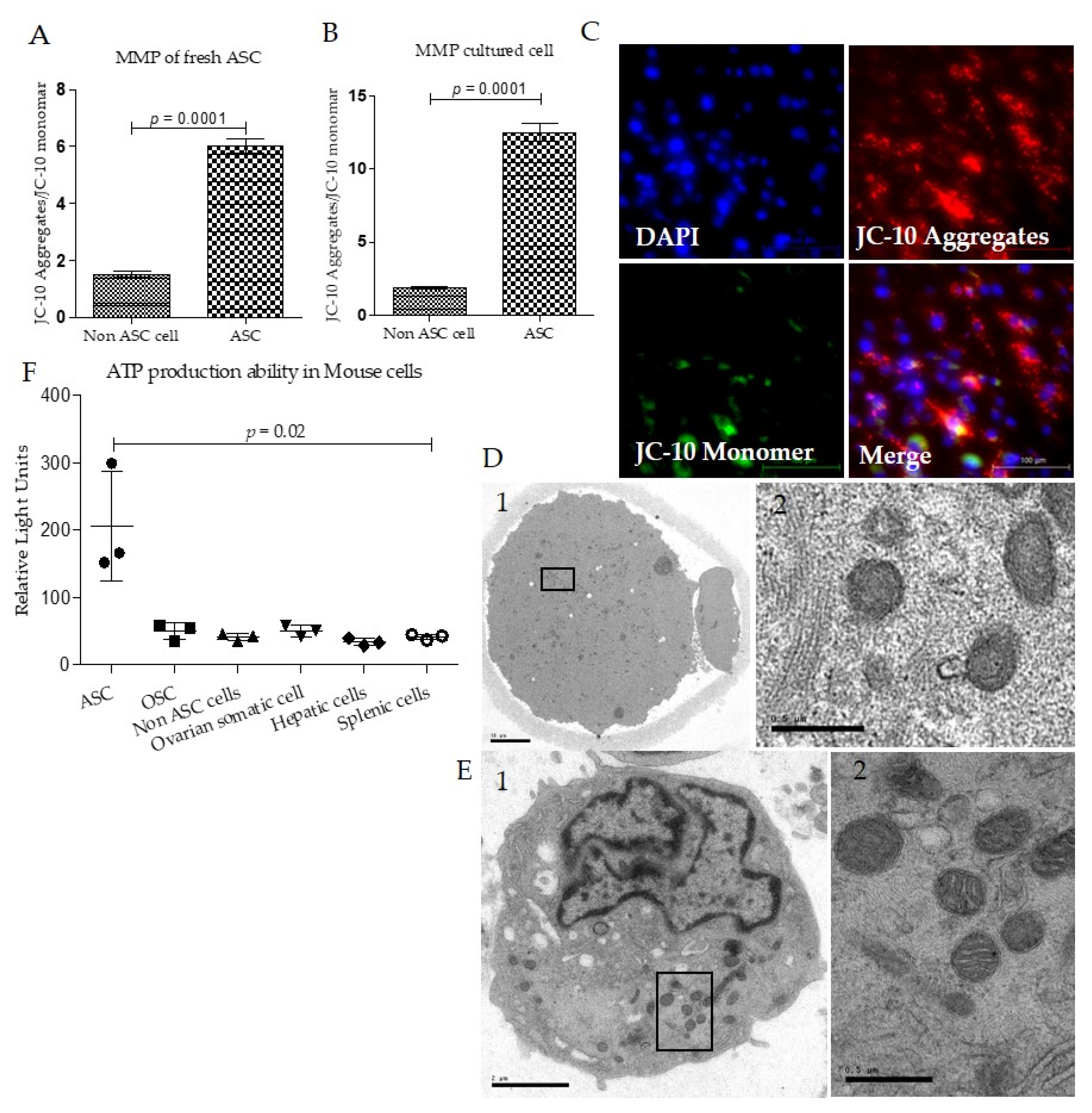
2.7. Mitochondria Membrane Potential Analysis
2.8. ATP Production Analysis
2.9. ROS Analysis
2.10. Transmission Electron Microscopy Analysis
2.11. Mitochondria Isolation and Supplementation
2.12. Embryo Development Analysis
2.13. Statistical Analysis
3. Results
3.1. Mouse Adipose Stem cell (ASC) Isolation and Confirmation
3.2. Mouse Adipose Stem Cell Mitochondria Have High Membrane Potentials and ATP Production Capacity
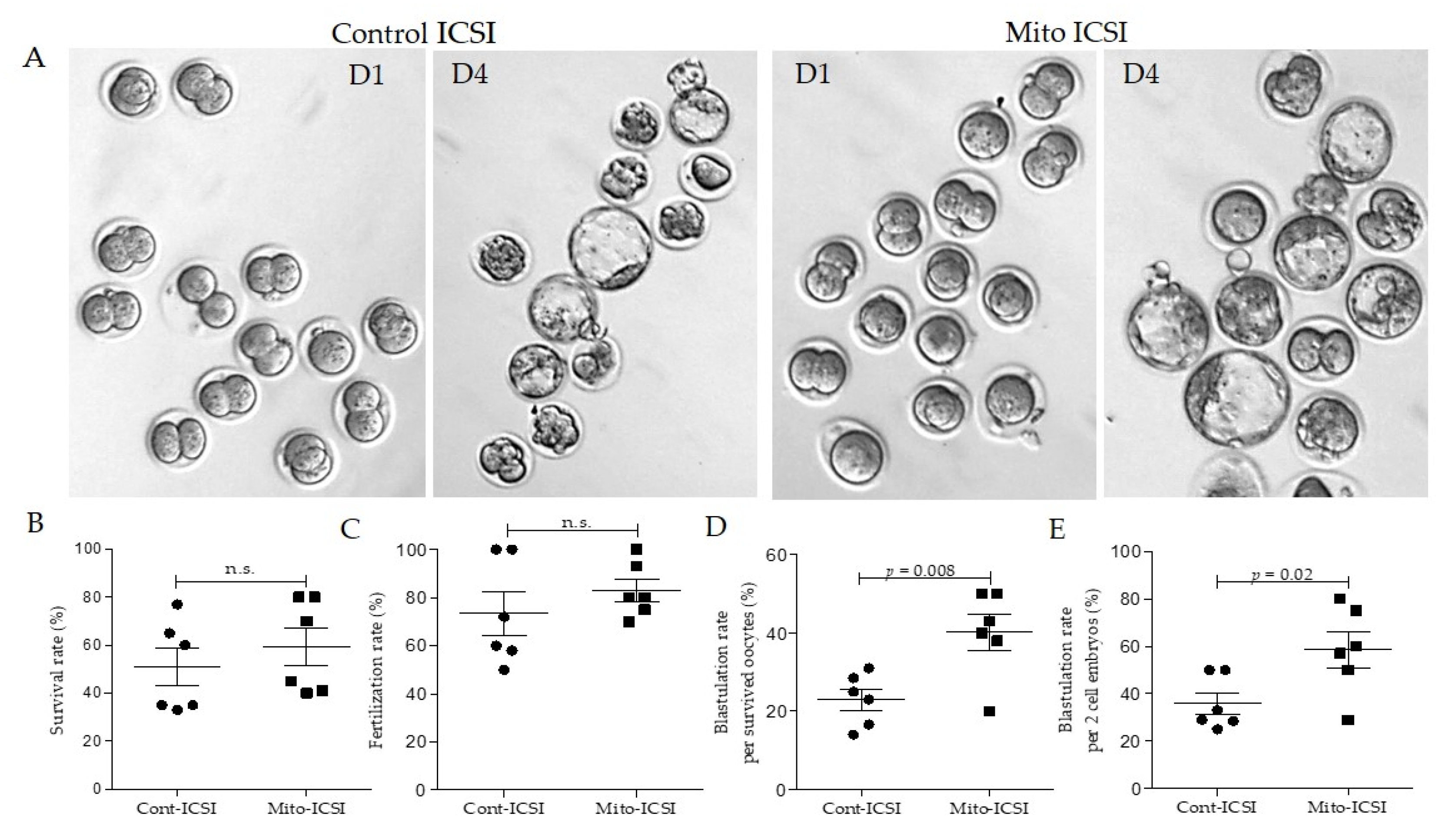
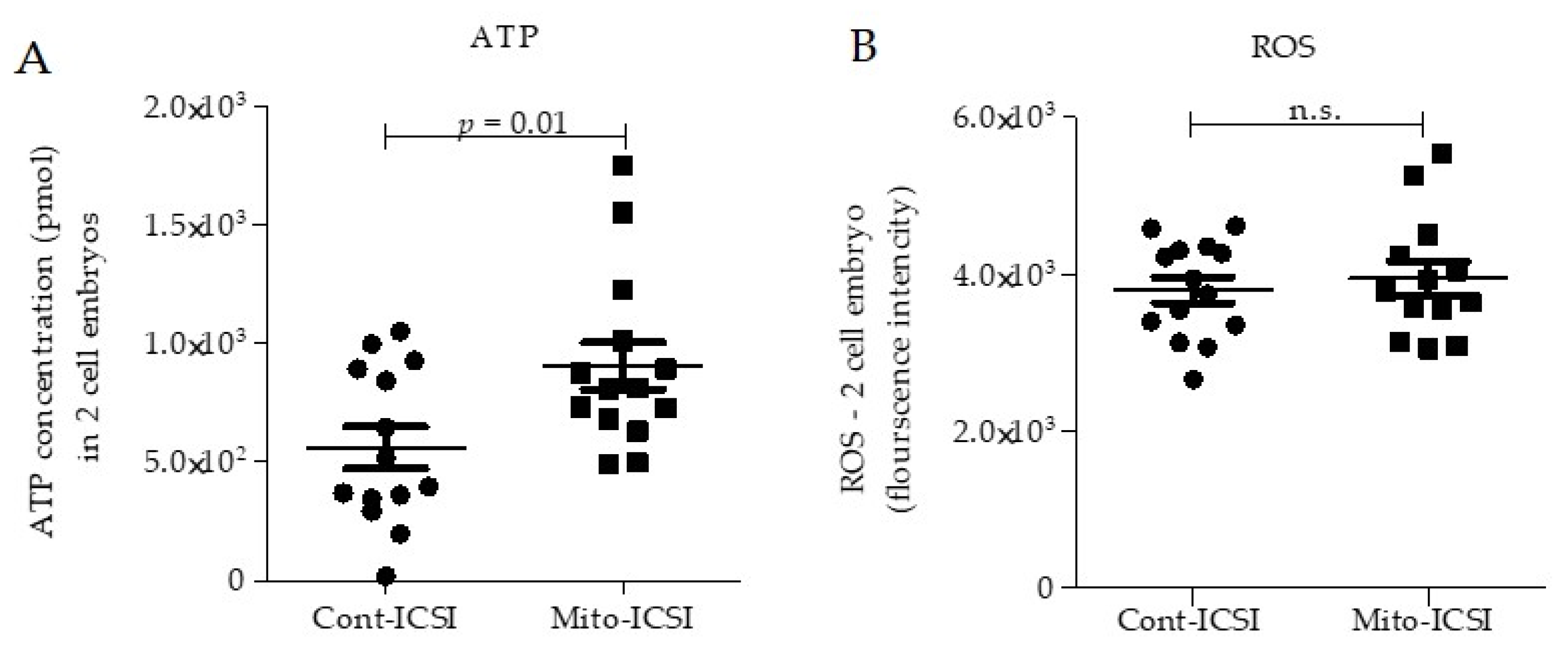
3.3. Adipose Stem Cell Mitochondria Supplementation during the ICSI Process Improves Embryo Development and Live Birth Capacity
3.4. The Mitochondria Supplemented Group Has Higher Active Mitochondria in Early Embryos
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iussig, B.; Maggiulli, R.; Fabozzi, G.; Bertelle, S.; Vaiarelli, A.; Cimadomo, D.; Ubaldi, F.M.; Rienzi, L. A Brief History of Oocyte Cryopreservation: Arguments and Facts. Acta Obstet. Gynecol. Scand. 2019, 98, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornet-Bartolomé, D.; Rodriguez, A.; García, D.; Barragán, M.; Vassena, R. Efficiency and Efficacy of Vitrification in 35 654 Sibling Oocytes from Donation Cycles. Hum. Reprod. 2020, 35, 2262–2271. [Google Scholar] [CrossRef] [PubMed]
- Ghetler, Y.; Yavin, S.; Shalgi, R.; Arav, A. The Effect of Chilling on Membrane Lipid Phase Transition in Human Oocytes and Zygotes. Hum. Reprod. 2005, 20, 3385–3389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghetler, Y.; Skutelsky, E.; Ben Nun, I.; Ben Dor, L.; Amihai, D.; Shalgi, R. Human Oocyte Cryopreservation and the Fate of Cortical Granules. Fertil. Steril. 2006, 86, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T.; Mohamed, M.S.; Aleem, A.A.N.; Sharma, R.K.; Worley, S.E.; Thornton, J.; Agarwal, A. Differential Growth of Human Embryos in Vitro: Role of Reactive Oxygen Species. Fertil. Steril. 2004, 82, 593–600. [Google Scholar] [CrossRef]
- du Plessis, S.S.; Makker, K.; Desai, N.R.; Agarwal, A. Impact of Oxidative Stress on IVF. Expert Rev. Obstet. Gynecol. 2008, 3, 539–554. [Google Scholar] [CrossRef]
- Nohales-Córcoles, M.; Sevillano-Almerich, G.; Di Emidio, G.; Tatone, C.; Cobo, A.C.; Dumollard, R.; De Los Santos Molina, M.J. Impact of Vitrification on the Mitochondrial Activity and Redox Homeostasis of Human Oocyte. Hum. Reprod. 2016, 31, 1850–1858. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.W.; Hwang, K.J.; Kwon, H.C.; Kim, H.S.; Choi, K.W.; Oh, K.S. Detection of Reactive Oxygen Species (ROS) and Apoptosis in Human Fragmented Embryos. Hum. Reprod. 1998, 13, 998–1002. [Google Scholar] [CrossRef]
- Attaran, M.; Pasqualotto, E.; Falcone, T.; Goldberg, J.M.; Miller, K.F.; Agarwal, A.; Sharma, R.K. The Effect of Follicular Fluid Reactive Oxygen Species on the Outcome of in Vitro Fertilization. Int. J. Fertil. Womens Med. 2000, 45, 314–320. [Google Scholar]
- Bittner, L.; Wyck, S.; Herrera, C.; Siuda, M.; Wrenzycki, C.; van Loon, B.; Bollwein, H. Negative Effects of Oxidative Stress in Bovine Spermatozoa on in Vitro Development and DNA Integrity of Embryos. Reprod. Fertil. Dev. 2018, 30, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Lan, K.-C.; Lin, Y.-C.; Chang, Y.-C.; Lin, H.-J.; Tsai, Y.-R.; Kang, H.-Y. Limited Relationships between Reactive Oxygen Species Levels in Culture Media and Zygote and Embryo Development. J. Assist. Reprod. Genet. 2019, 36, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohaya, N.; Fujiwara, K.; Ito, J.; Kashiwazaki, N. Generation of Live Offspring from Vitrified Mouse Oocytes of C57BL/6J Strain. PLoS ONE 2013, 8, e58063. [Google Scholar] [CrossRef] [Green Version]
- van Blerkom, J.; Davis, P.; Mathwig, V.; Alexander, S. Domains of High-Polarized and Low-Polarized Mitochondria May Occur in Mouse and Human Oocytes and Early Embryos. Hum. Reprod. 2002, 17, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.; van Blerkom, J.; Davis, P.; Toledo, A.A. Cryopreservation of Metaphase II Human Oocytes Effects Mitochondrial Membrane Potential: Implications for Developmental Competence. Hum. Reprod. 2004, 19, 1861–1866. [Google Scholar] [CrossRef] [Green Version]
- Gualtieri, R.; Iaccarino, M.; Mollo, V.; Prisco, M.; Iaccarino, S.; Talevi, R. Slow Cooling of Human Oocytes: Ultrastructural Injuries and Apoptotic Status. Fertil. Steril. 2009, 91, 1023–1034. [Google Scholar] [CrossRef]
- Amoushahi, M.; Salehnia, M.; Mowla, S.J. Vitrification of Mouse MII Oocyte Decreases the Mitochondrial DNA Copy Number, TFAM Gene Expression and Mitochondrial Enzyme Activity. J. Reprod. Infertil. 2017, 18, 343–351. [Google Scholar]
- Gualtieri, R.; Mollo, V.; Barbato, V.; Fiorentino, I.; Iaccarino, M.; Talevi, R. Ultrastructure and Intracellular Calcium Response during Activation in Vitrified and Slow-Frozen Human Oocytes. Hum. Reprod. 2011, 26, 2452–2460. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Qazi, I.H.; Guo, S.; Yang, J.; Qin, J.; Lv, T.; Zang, S.; Zhang, Y.; Zeng, C.; Meng, Q.; et al. Melatonin Improves the First Cleavage of Parthenogenetic Embryos from Vitrified-Warmed Mouse Oocytes Potentially by Promoting Cell Cycle Progression. J. Anim. Sci. Biotechnol. 2021, 12, 84. [Google Scholar] [CrossRef]
- Harada, Y.; Kinutani, M.; Horiuchi, T. Improved Developmental Potential of Mouse Vitrified-Warmed Oocytes Achieved by Culturing in Recovery Medium with Glutathione Ethyl Ester (GSH-OEt). Reprod. Med. Biol. 2021, 20, 505–512. [Google Scholar] [CrossRef]
- Wang, N.; Hao, H.-S.; Li, C.-Y.; Zhao, Y.-H.; Wang, H.-Y.; Yan, C.-L.; Du, W.-H.; Wang, D.; Liu, Y.; Pang, Y.-W.; et al. Calcium Ion Regulation by BAPTA-AM and Ruthenium Red Improved the Fertilisation Capacity and Developmental Ability of Vitrified Bovine Oocytes. Sci. Rep. 2017, 7, 10652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wai, T.; Ao, A.; Zhang, X.; Cyr, D.; Dufort, D.; Shoubridge, E.A. The Role of Mitochondrial DNA Copy Number in Mammalian Fertility1. Biol. Reprod. 2010, 83, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, K.; Iwase, A.; Mawatari, M.; Wang, J.; Yamashita, M.; Kikkawa, F. Mitochondrial Membrane Potential in 2-Cell Stage Embryos Correlates with the Success of Preimplantation Development. Reproduction 2014, 147, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Labarta, E.; de Los Santos, M.J.; Escribá, M.J.; Pellicer, A.; Herraiz, S. Mitochondria as a Tool for Oocyte Rejuvenation. Fertil. Steril. 2019, 111, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Varela, C.; Labarta, E. Clinical Application of Antioxidants to Improve Human Oocyte Mitochondrial Function: A Review. Antioxidants 2020, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, S.; Darbandi, M.; Khorram Khorshid, H.R.; Sadeghi, M.R.; Agarwal, A.; Sengupta, P.; Al-Hasani, S.; Akhondi, M.M. Ooplasmic Transfer in Human Oocytes: Efficacy and Concerns in Assisted Reproduction. Reprod. Biol. Endocrinol. 2017, 15, 77. [Google Scholar] [CrossRef] [Green Version]
- Fakih, M.H. The AUGMENTSM Treatment: Physician Reported Outcomes of the Initial Global Patient Experience. J. Fertil. IVF-Worldw. Reprod. Med. Genet. Stem Cell Biol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Oktay, K.; Baltaci, V.; Sonmezer, M.; Turan, V.; Unsal, E.; Baltaci, A.; Aktuna, S.; Moy, F. Oogonial Precursor Cell-Derived Autologous Mitochondria Injection to Improve Outcomes in Women with Multiple IVF Failures due to Low Oocyte Quality. Reprod. Sci. 2015, 22, 1612–1617. [Google Scholar] [CrossRef]
- Wang, Z.B.; Hao, J.X.; Meng, T.G.; Guo, L.; Dong, M.Z.; Fan, L.H.; Ouyang, Y.C.; Wang, G.; Sun, Q.Y.; Ou, X.H.; et al. Transfer of Autologous Mitochondria from Adipose Tissue-Derived Stem Cells Rescues Oocyte Quality and Infertility in Aged Mice. Aging 2017, 9, 2480–2488. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Tao, L.; Yue, Y.; Ren, L.; Zhang, Z.; Wang, X.; Tian, J.; An, L. Mitochondrial Transfer from Induced Pluripotent Stem Cells Rescues Developmental Potential of in Vitro Fertilized Embryos from Aging Females. Biol. Reprod. 2021, 104, 1114–1125. [Google Scholar] [CrossRef]
- Igarashi, H.; Takahashi, T.; Abe, H.; Nakano, H.; Nakajima, O.; Nagase, S. Poor Embryo Development in Post-Ovulatory in Vivo-Aged Mouse Oocytes Is Associated with Mitochondrial Dysfunction, but Mitochondrial Transfer from Somatic Cells Is not Sufficient for Rejuvenation. Hum. Reprod. 2016, 31, 2331–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Liu, Z.; Li, H.; Zhu, L.; Xing, F. Pregnancy in a 46-Year-Old Woman after Autologous Granular Cell Mitochondria Transfer. Di Yi Jun Yi Da Xue Xue Bao 2003, 23, 743–747. [Google Scholar] [PubMed]
- White, Y.A.R.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte Formation by Mitotically Active Germ Cells Purified from Ovaries of Reproductive-Age Women. Nat. Med. 2012, 18, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.; Minami, N.; Yamada, M.; Imai, H. Excessive Concentration of Glucose during in Vitro Maturation Impairs the Developmental Competence of Bovine Oocytes after in Vitro Fertilization: Relevance to Intracellular Reactive Oxygen Species and Glutathione Contents. Mol. Reprod. Dev. 2000, 56, 520–526. [Google Scholar] [CrossRef]
- Woods, D.C.; Tilly, J.L. Isolation, Characterization and Propagation of Mitotically Active Germ Cells from Adult Mouse and Human Ovaries. Nat. Protoc. 2013, 8, 966–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagnone, G.L.M.; Tsai, T.-S.; Makanji, Y.; Matthews, P.; Gould, J.; Bonkowski, M.S.; Elgass, K.D.; Wong, A.S.A.; Wu, L.E.; McKenzie, M.; et al. Restoration of Normal Embryogenesis by Mitochondrial Supplementation in Pig Oocytes Exhibiting Mitochondrial DNA Deficiency. Sci. Rep. 2016, 6, 23229. [Google Scholar] [CrossRef] [PubMed]
- Kilroy, G.; Dietrich, M.; Wu, X.; Gimble, J.M.; Floyd, Z.E. Isolation of Murine Adipose-Derived Stromal/Stem Cells for Adipogenic Differentiation or Flow Cytometry-Based Analysis. Methods Mol. Biol. 2018, 1773, 137–146. [Google Scholar] [CrossRef]
- Yoshimura, K.; Shigeura, T.; Matsumoto, D.; Sato, T.; Takaki, Y.; Aiba-Kojima, E.; Sato, K.; Inoue, K.; Nagase, T.; Koshima, I.; et al. Characterization of Freshly Isolated and Cultured Cells Derived from the Fatty and Fluid Portions of Liposuction Aspirates. J. Cell Physiol. 2006, 208, 64–76. [Google Scholar] [CrossRef]
- Kladnická, I.; Čedíková, M.; Kripnerová, M.; Dvořáková, J.; Kohoutová, M.; Tůma, Z.; Müllerová, D.; Kuncová, J. Mitochondrial Respiration of Adipocytes Differentiating from Human Mesenchymal Stem Cells Derived from Adipose Tissue. Physiol. Res. 2019, 68, S287–S296. [Google Scholar] [CrossRef]
- Revel, A.; Moshe, N.; Helman, A.; Safran, A.; Simon, A.; Koler, M. Mouse Embryos Generated from Frozen-Thawed Oocytes Can Successfully Survive a Second Cryopreservation. Hum. Reprod. 2004, 19, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Chamayou, S.; Alecci, C.; Ragolia, C.; Storaci, G.; Maglia, E.; Russo, E.; Guglielmino, A. Comparison of In-Vitro Outcomes from Cryopreserved Oocytes and Sibling Fresh Oocytes. Reprod. Biomed. Online 2006, 12, 730–736. [Google Scholar] [CrossRef]
- Endoh, K.; Mochida, K.; Ogonuki, N.; Ohkawa, M.; Shinmen, A.; Ito, M.; Kashiwazaki, N.; Ogura, A. The Developmental Ability of Vitrified Oocytes from Different Mouse Strains Assessed by Parthenogenetic Activation and Intracytoplasmic Sperm Injection. J. Reprod. Dev. 2007, 53, 1199–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.; Morimoto, N.; Yamanaka, M.; Matsumoto, H.; Yamochi, T.; Goto, H.; Inoue, M.; Nakaoka, Y.; Shibahara, H.; Morimoto, Y. Quantitative and Qualitative Changes of Mitochondria in Human Preimplantation Embryos. J. Assist. Reprod. Genet. 2017, 34, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Ishibashi, S.; Iglesias-Gonzalez, J.; Chen, Y.; Love, N.R.; Amaya, E. Ca2+-Induced Mitochondrial ROS Regulate the Early Embryonic Cell Cycle. Cell Rep 2018, 22, 218–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, M.; Richings, N.M.; Leung, A.; Sakkas, D.; Catt, S. A Major Increase in Oocyte Cryopreservation Cycles in the USA, Australia and New Zealand since 2010 Is Highlighted by Younger Women but a Need for Standardized Data Collection. Hum. Reprod. 2021, 36, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Niederberger, C.; Pellicer, A.; Cohen, J.; Gardner, D.K.; Palermo, G.D.; O’Neill, C.L.; Chow, S.; Rosenwaks, Z.; Cobo, A.; Swain, J.E.; et al. Forty Years of IVF. Fertil. Steril. 2018, 110, 185–324.e5. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, J.K.; Yoon, S.Y.; Park, E.A.; Jun, J.H.; Lim, H.J.; Kim, J.; Song, H. Advanced Maternal Age Deteriorates the Developmental Competence of Vitrified Oocytes in Mice. Cells 2021, 10, 1563. [Google Scholar] [CrossRef]
- Palmerini, M.G.; Antinori, M.; Maione, M.; Cerusico, F.; Versaci, C.; Nottola, S.A.; Macchiarelli, G.; Khalili, M.A.; Antinori, S. Ultrastructure of Immature and Mature Human Oocytes after Cryotop Vitrification. J. Reprod. Dev. 2014, 60, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Wu, C.; Muneri, C.W.; Niu, Y.; Zhang, S.; Rui, R.; Zhang, D. Changes in Mitochondrial Function in Porcine Vitrified MII-Stage Oocytes and Their Impacts on Apoptosis and Developmental Ability. Cryobiology 2015, 71, 291–298. [Google Scholar] [CrossRef]
- Belli, M.; Palmerini, M.G.; Bianchi, S.; Bernardi, S.; Khalili, M.A.; Nottola, S.A.; Macchiarelli, G. Ultrastructure of Mitochondria of Human Oocytes in Different Clinical Conditions during Assisted Reproduction. Arch. Biochem. Biophys. 2021, 703, 108854. [Google Scholar] [CrossRef]
- Pikó, L.; Taylor, K.D. Amounts of Mitochondrial DNA and Abundance of Some Mitochondrial Gene Transcripts in Early Mouse Embryos. Dev. Biol. 1987, 123, 364–374. [Google Scholar] [CrossRef]
- May-Panloup, P.; Vignon, X.; Chrétien, M.-F.; Heyman, Y.; Tamassia, M.; Malthièry, Y.; Reynier, P. Increase of Mitochondrial DNA Content and Transcripts in Early Bovine Embryogenesis Associated with Upregulation of MtTFA and NRF1 Transcription Factors. Reprod. Biol. Endocrinol. 2005, 3, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spikings, E.C.; Alderson, J.; John, J.C. Regulated Mitochondrial DNA Replication During Oocyte Maturation Is Essential for Successful Porcine Embryonic Development. Biol. Reprod. 2007, 76, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K.; Wale, P.L. Analysis of Metabolism to Select Viable Human Embryos for Transfer. Fertil. Steril. 2013, 99, 1062–1072. [Google Scholar] [CrossRef]
- Bradley, J.; Swann, K. Mitochondria and Lipid Metabolism in Mammalian Oocytes and Early Embryos. Int. J. Dev. Biol. 2019, 63, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Rugarli, E.I.; Langer, T. Mitochondrial Quality Control: A Matter of Life and Death for Neurons. EMBO J. 2012, 31, 1336–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, R.P.S.; de Boer, K. The Bottleneck: Mitochondrial Imperatives in Oogenesis and Ovarian Follicular Fate. Mol. Cell. Endocrinol. 1998, 145, 81–88. [Google Scholar] [CrossRef]
- Wilding, M.; Dale, B.; Marino, M.; di Matteo, L.; Alviggi, C.; Pisaturo, M.L.; Lombardi, L.; de Placido, G. Mitochondrial Aggregation Patterns and Activity in Human Oocytes and Preimplantation Embryos. Hum. Reprod. 2001, 16, 909–917. [Google Scholar] [CrossRef]
- Harvey, A.J. Mitochondria in Early Development: Linking the Microenvironment, Metabolism and the Epigenome. Reproduction 2019, 157, R159–R179. [Google Scholar] [CrossRef] [Green Version]
- Chappel, S. The Role of Mitochondria from Mature Oocyte to Viable Blastocyst. Obstet. Gynecol. Int. 2013, 2013, 183024. [Google Scholar] [CrossRef] [Green Version]
- Konari, N.; Nagaishi, K.; Kikuchi, S.; Fujimiya, M. Mitochondria Transfer from Mesenchymal Stem Cells Structurally and Functionally Repairs Renal Proximal Tubular Epithelial Cells in Diabetic Nephropathy In Vivo. Sci. Rep. 2019, 9, 5184. [Google Scholar] [CrossRef] [Green Version]
- Mohammadalipour, A.; Dumbali, S.P.; Wenzel, P.L. Mitochondrial Transfer and Regulators of Mesenchymal Stromal Cell Function and Therapeutic Efficacy. Front. Cell Dev. Biol 2020, 8, 603292. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, S.; Chaudhuri, R.; Agrawal, A.; Mohanty, S. Regenerative Abilities of Mesenchymal Stem Cells through Mitochondrial Transfer. J. Biomed. Sci. 2018, 25, 31. [Google Scholar] [CrossRef] [PubMed]
- Motta, P.M.; Nottola, S.A.; Makabe, S.; Heyn, R. Mitochondrial Morphology in Human Fetal and Adult Female Germ Cells. Hum. Reprod. 2000, 15, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef]
- May-Panloup, P.; Chrétien, M.F.; Jacques, C.; Vasseur, C.; Malthièry, Y.; Reynier, P. Low Oocyte Mitochondrial DNA Content in Ovarian Insufficiency. Hum. Reprod. 2005, 20, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Nagano, M.; Katagiri, S.; Takahashi, Y. ATP Content and Maturational/Developmental Ability of Bovine Oocytes with Various Cytoplasmic Morphologies. Zygote 2006, 14, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Zander-Fox, D.L.; Fullston, T.; McPherson, N.O.; Sandeman, L.; Kang, W.X.; Good, S.B.; Spillane, M.; Lane, M. Reduction of Mitochondrial Function by FCCP During Mouse Cleavage Stage Embryo Culture Reduces Birth Weight and Impairs the Metabolic Health of Offspring. Biol. Reprod. 2015, 92, 124. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y. Adenosine Triphosphate Content in Human Unfertilized Oocytes, Undivided Zygotes and Embryos Unsuitable for Transfer or Cryopreservation. J. Int. Med. Res. 2012, 40, 734–739. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cont-ICSI | Mito-ICSI | p-Value | |
|---|---|---|---|
| Total number of oocytes injected | 105 | 104 | |
| The survival rate after ICSI (%) | 50.4 (53/105) | 58.6 (61/104) | 0.46 |
| Fertilization rate per survived oocytes (%) | 75.4 (40/53) | 86.8 (53/61) | 0.36 |
| 2 cell rate per survived oocytes (%) | 59 (34/53) | 64.3 (44/61) | 0.71 |
| Blastulation rate per survived oocytes (%) | 24.5 (13/53) | 40.9 (25/61) | 0.008 |
| Blastulation rate per 2 cell embryos (%) | 38.2 (13/34) | 56.8 (25/44) | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kankanam Gamage, U.S.; Hashimoto, S.; Miyamoto, Y.; Nakano, T.; Yamanaka, M.; Koike, A.; Satoh, M.; Morimoto, Y. Mitochondria Transfer from Adipose Stem Cells Improves the Developmental Potential of Cryopreserved Oocytes. Biomolecules 2022, 12, 1008. https://doi.org/10.3390/biom12071008
Kankanam Gamage US, Hashimoto S, Miyamoto Y, Nakano T, Yamanaka M, Koike A, Satoh M, Morimoto Y. Mitochondria Transfer from Adipose Stem Cells Improves the Developmental Potential of Cryopreserved Oocytes. Biomolecules. 2022; 12(7):1008. https://doi.org/10.3390/biom12071008
Chicago/Turabian StyleKankanam Gamage, Udayanga Sanath, Shu Hashimoto, Yuki Miyamoto, Tatsuya Nakano, Masaya Yamanaka, Akiko Koike, Manabu Satoh, and Yoshiharu Morimoto. 2022. "Mitochondria Transfer from Adipose Stem Cells Improves the Developmental Potential of Cryopreserved Oocytes" Biomolecules 12, no. 7: 1008. https://doi.org/10.3390/biom12071008