
Expression of Zyxin in Non-Small Cell Lung Cancer—A Preliminary Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Material
2.2. Preparation of Tissue Microarrays (TMAs)
2.3. Immunohistochemistry (IHC)
2.4. Assessment of IHC Reactions
2.5. Cell Lines
2.6. Immunocytochemistry (ICC)
2.7. Immunofluorescence (IF)
2.8. Western Blot
2.9. RNA Isolation, Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.10. Laser Capture Microdissection (LCM) and Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.11. Statistical Analysis
3. Results
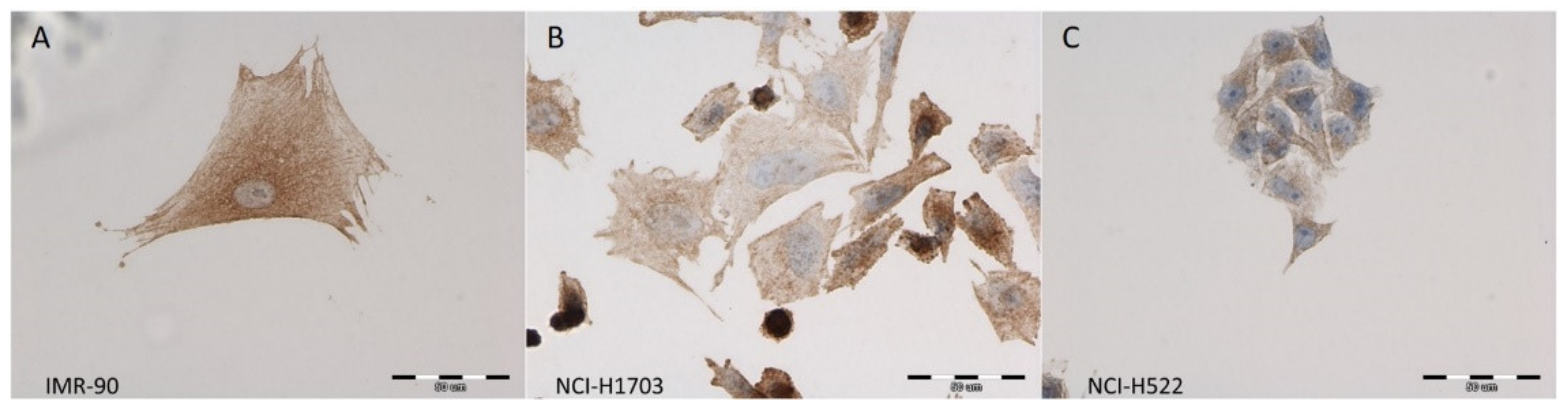
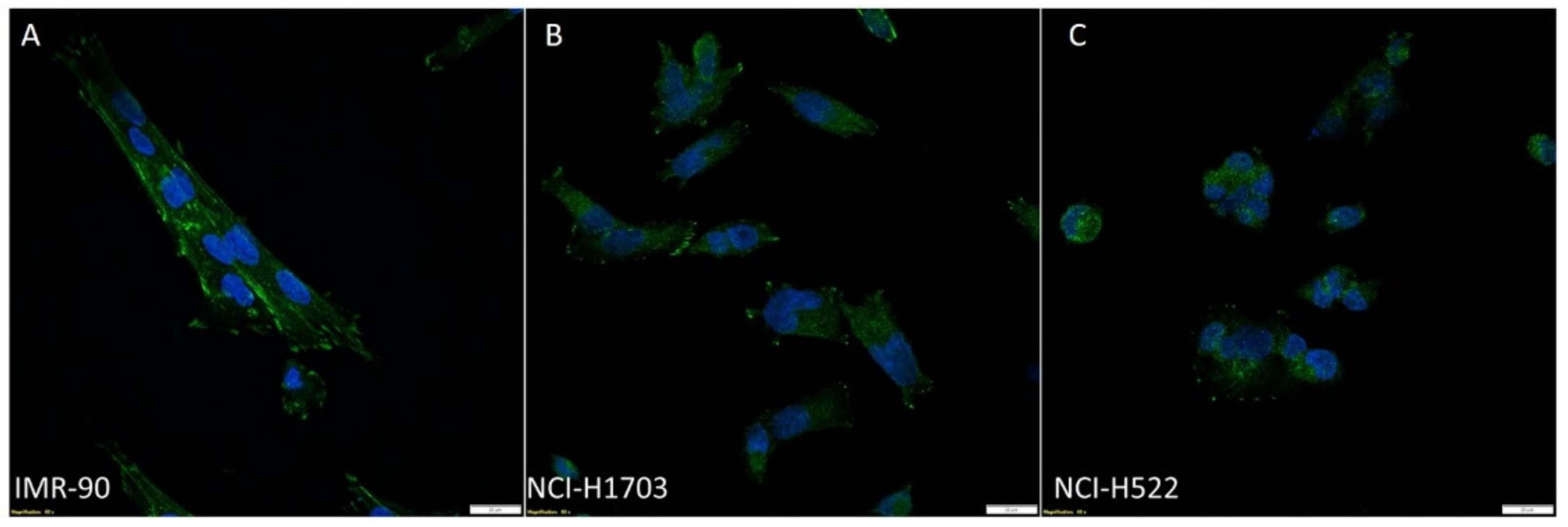
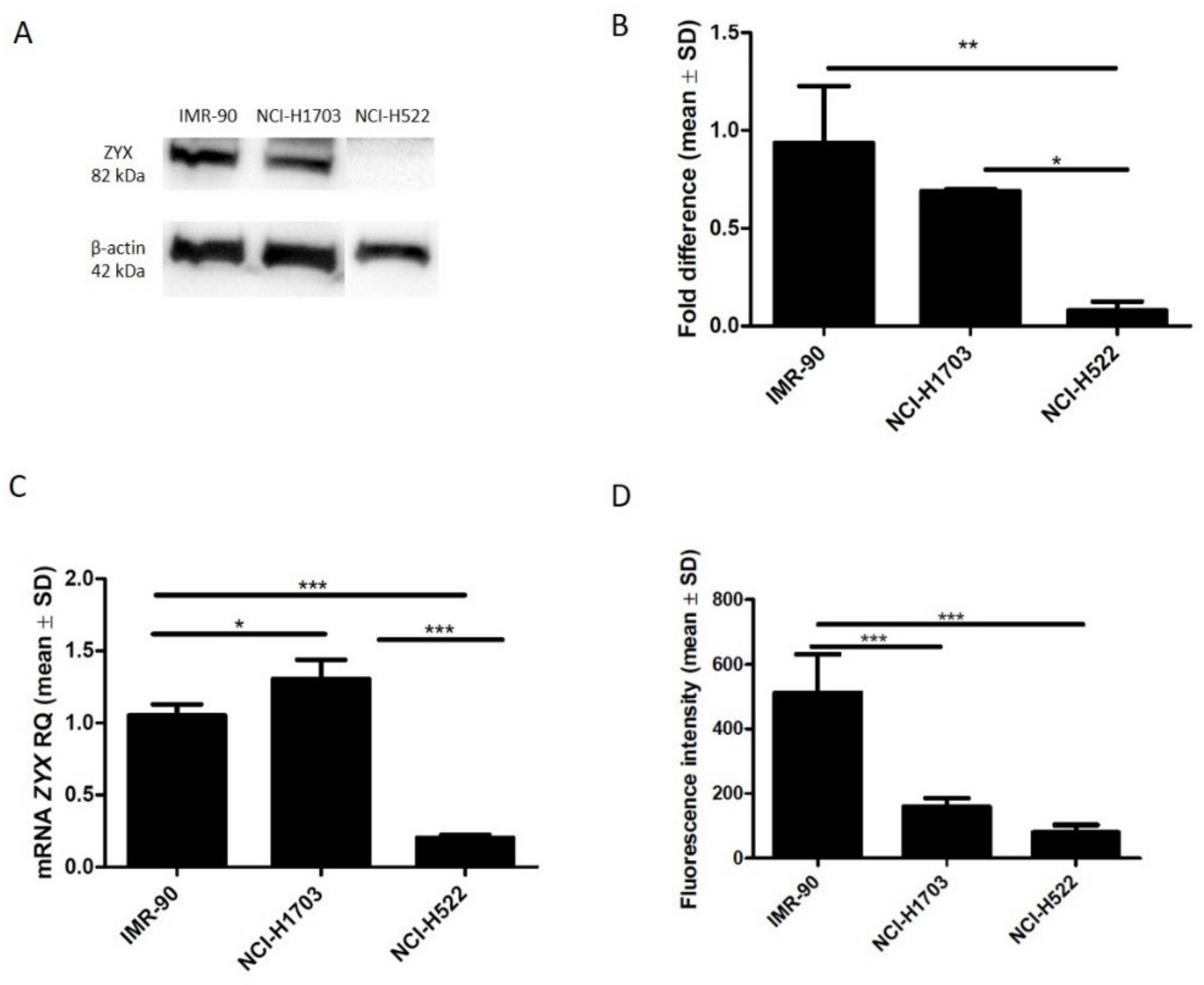
3.1. Expression of Zyxin in NSCLC Cell Lines and Normal Lung Fibroblasts
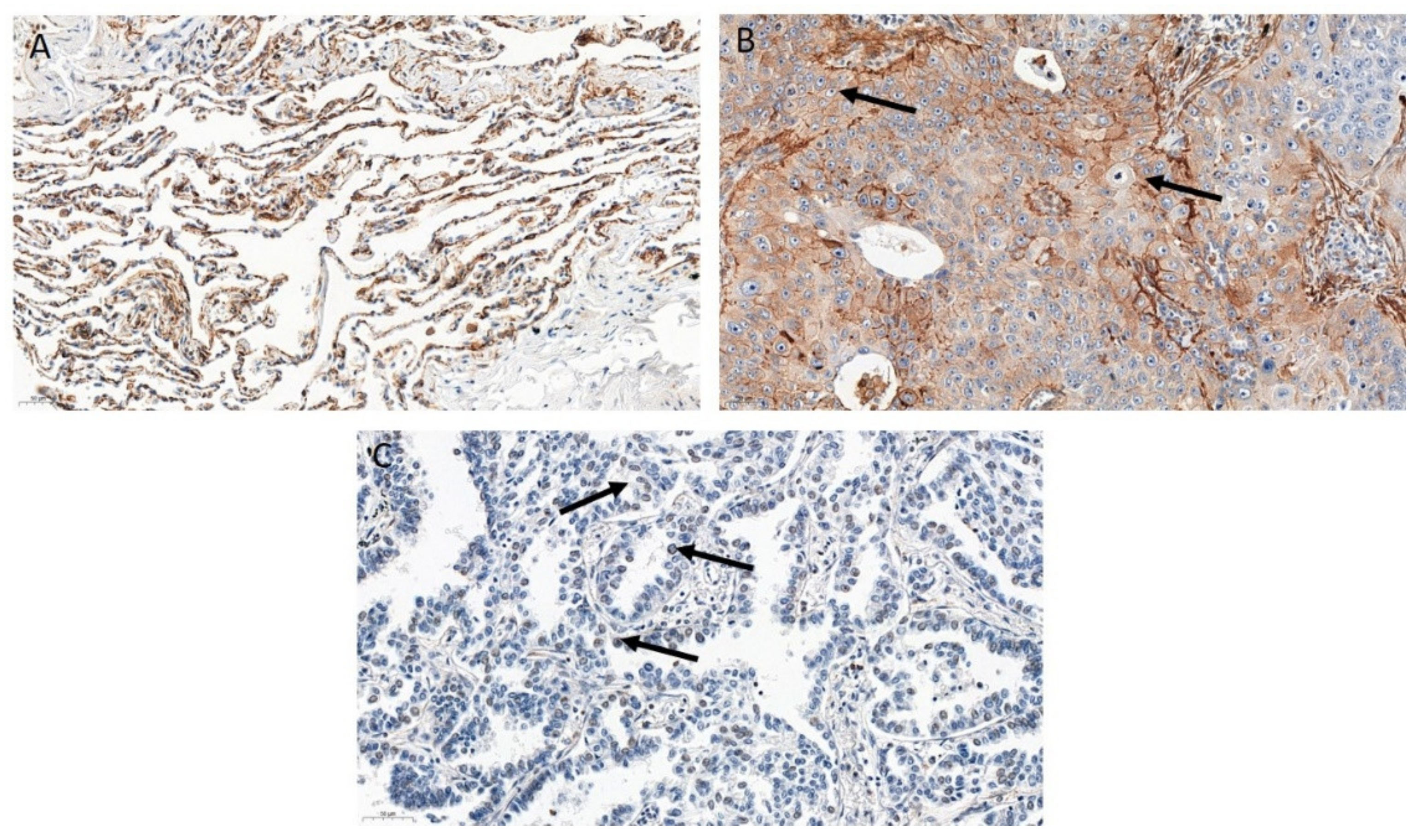
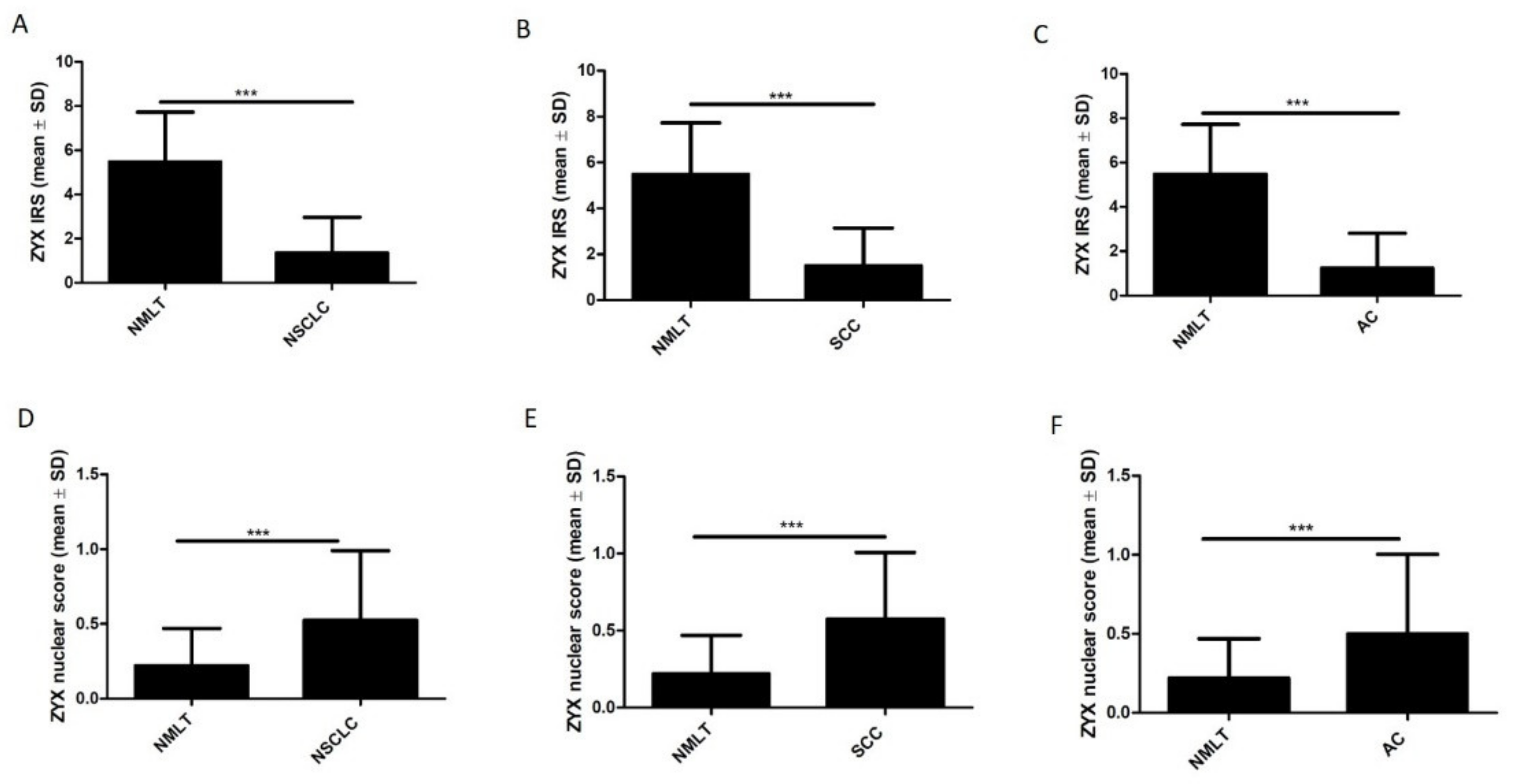
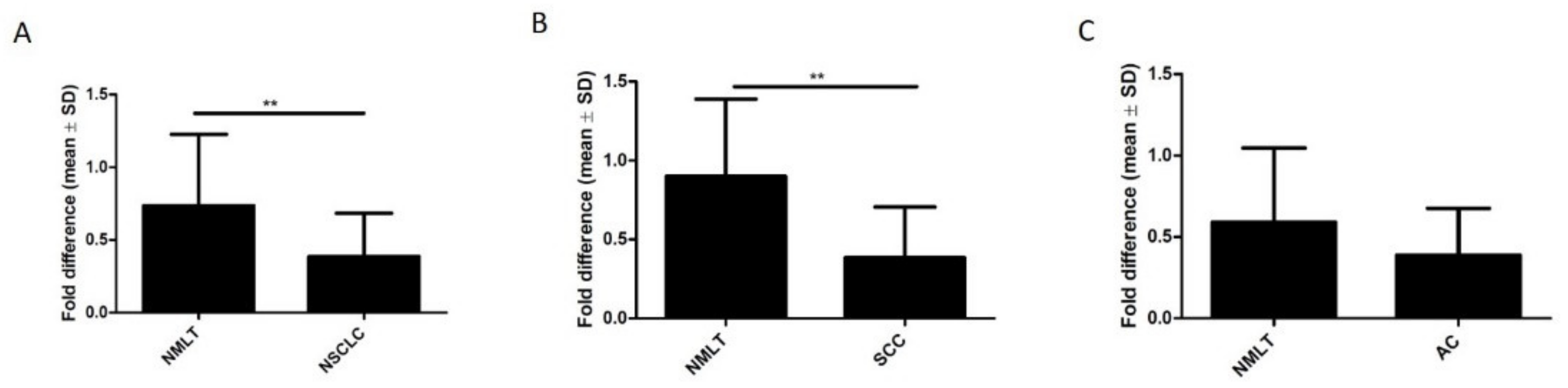
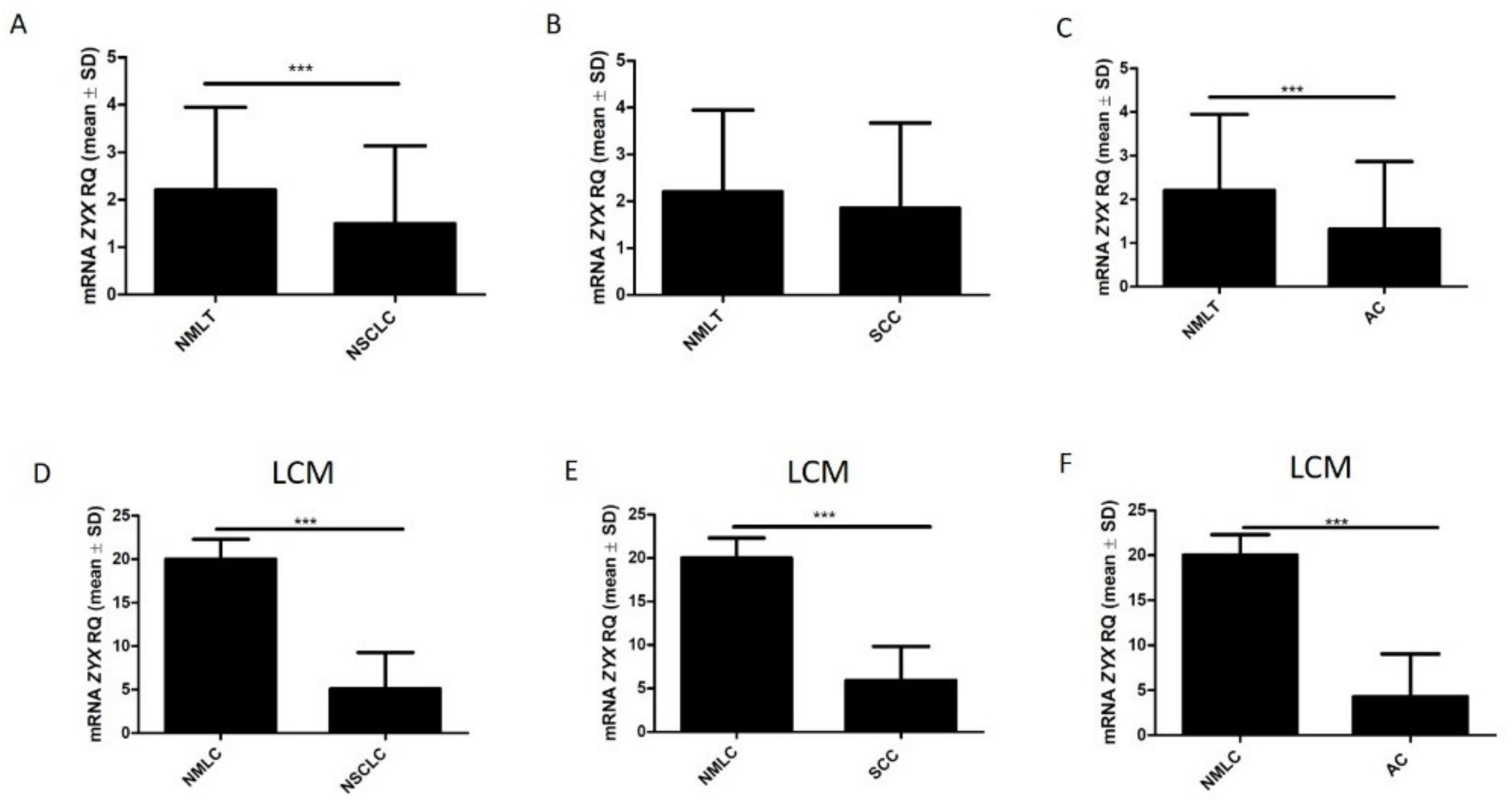
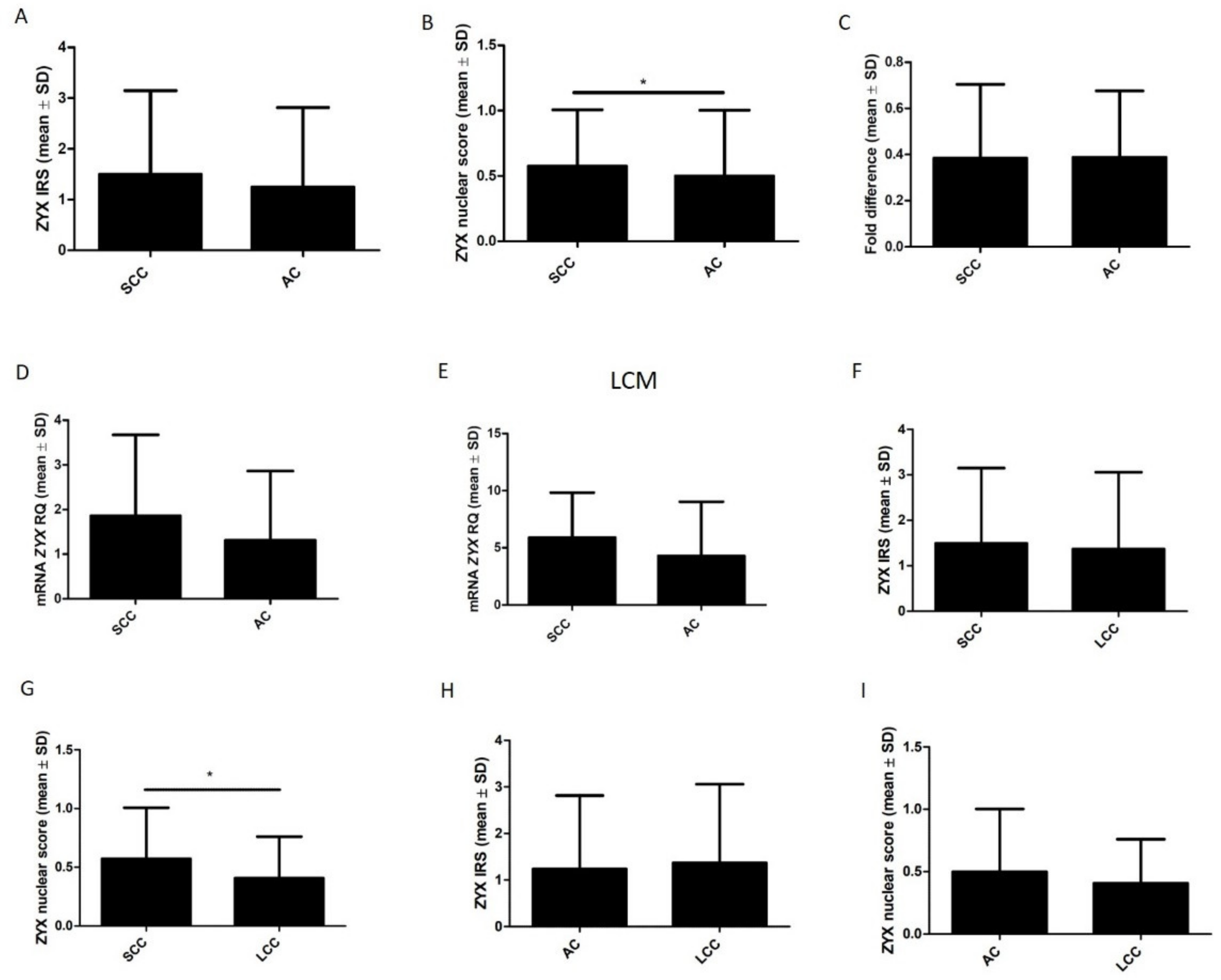
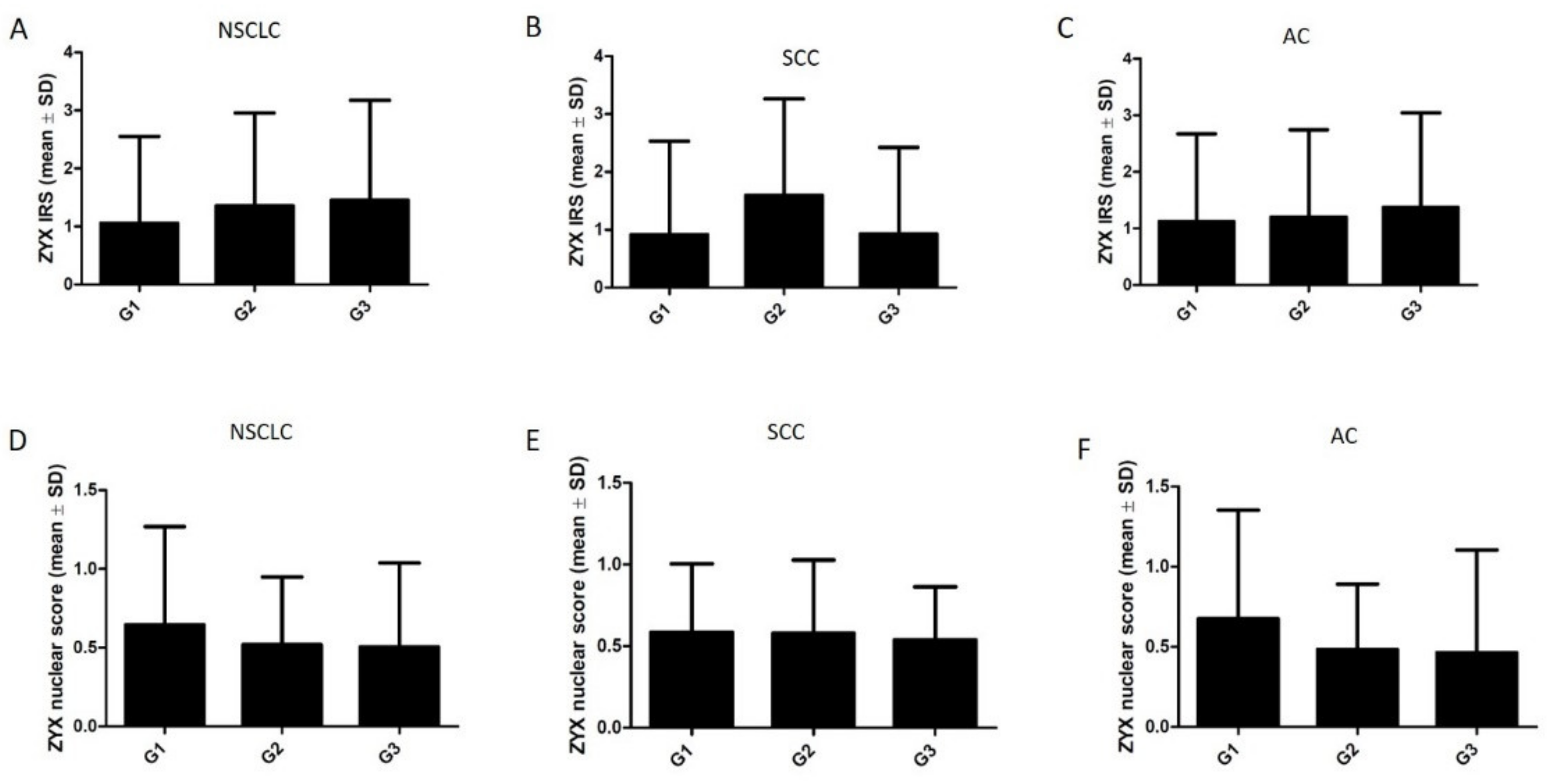
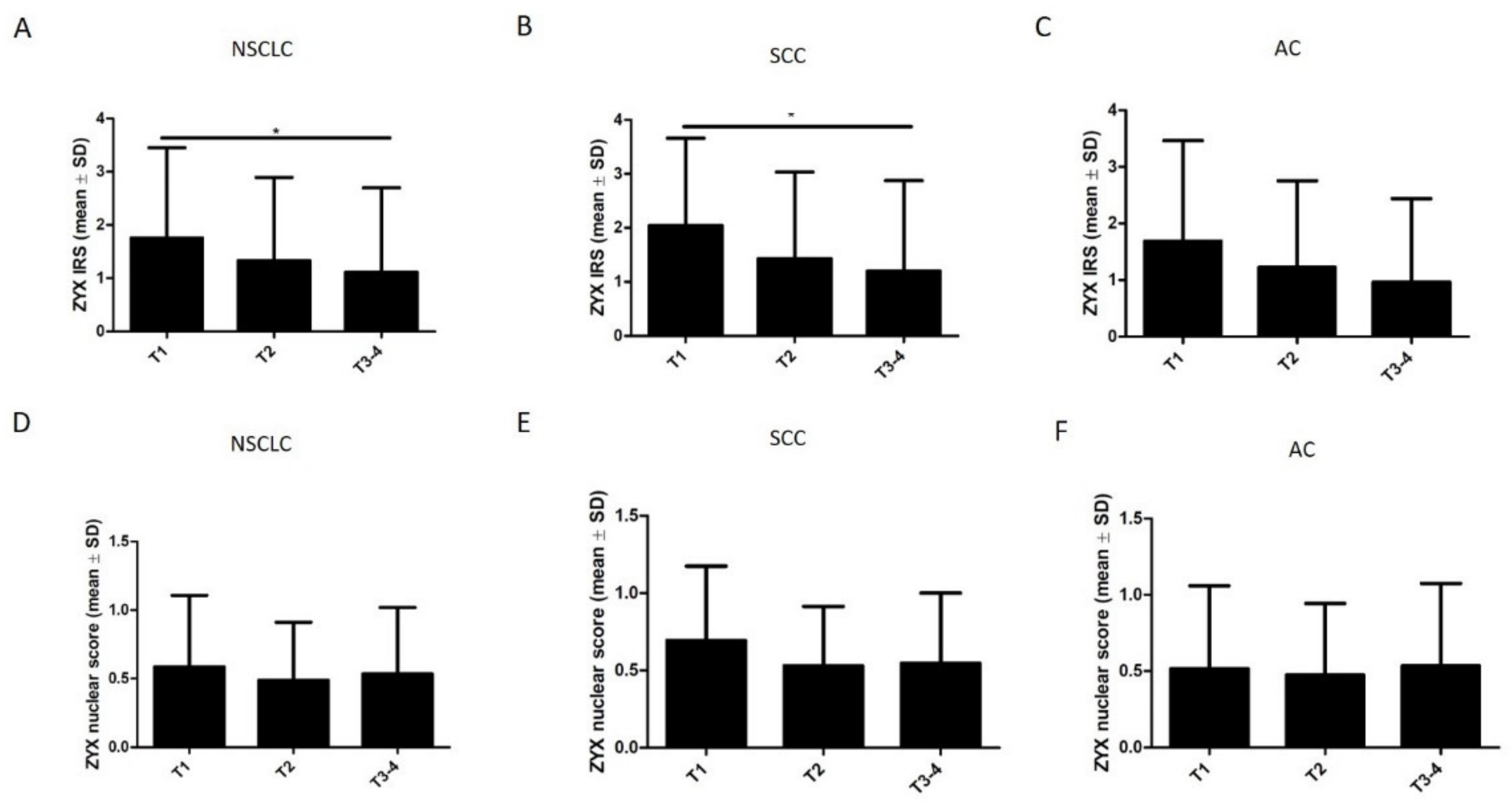
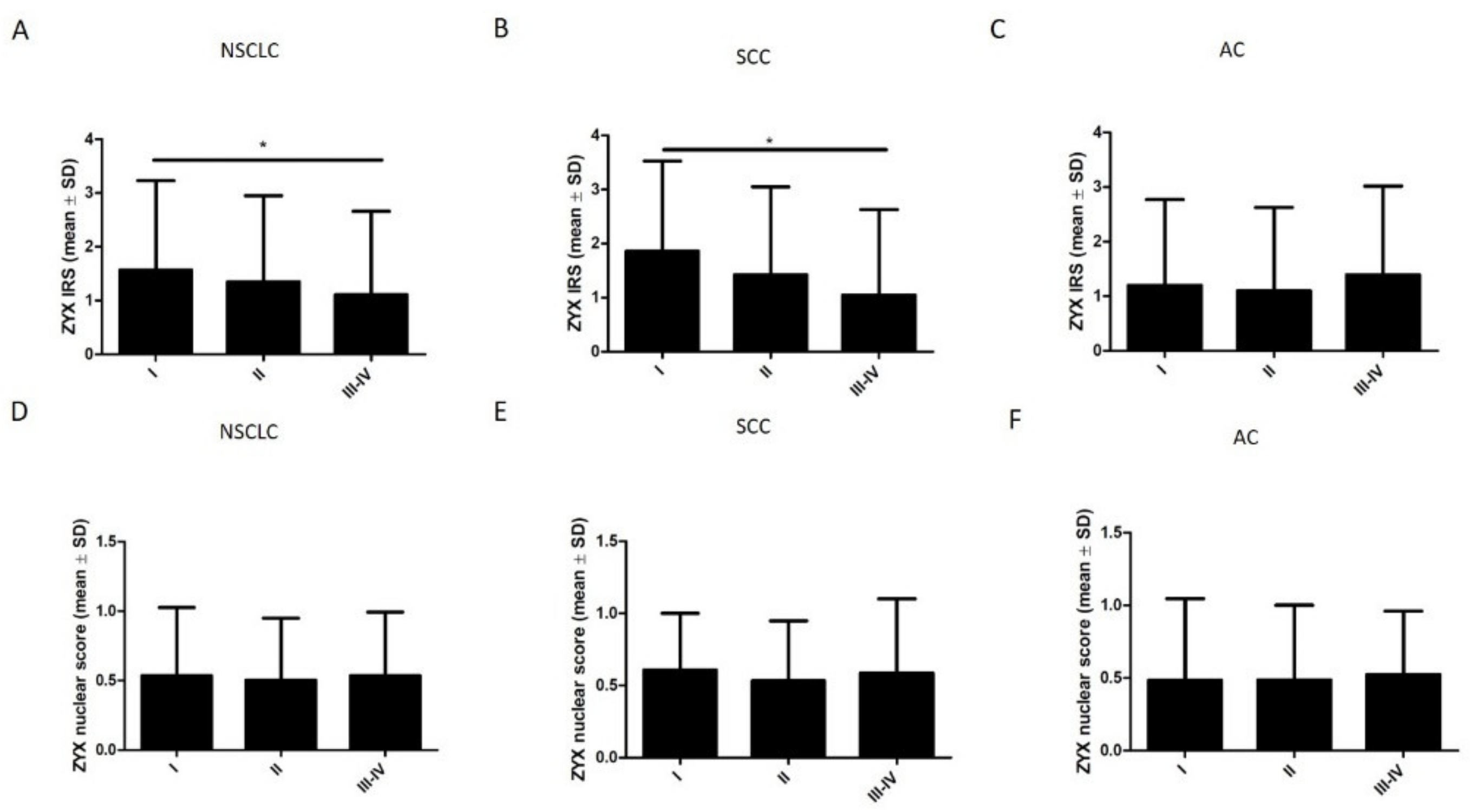
3.2. ZYX Expression in Patients with NSCLC
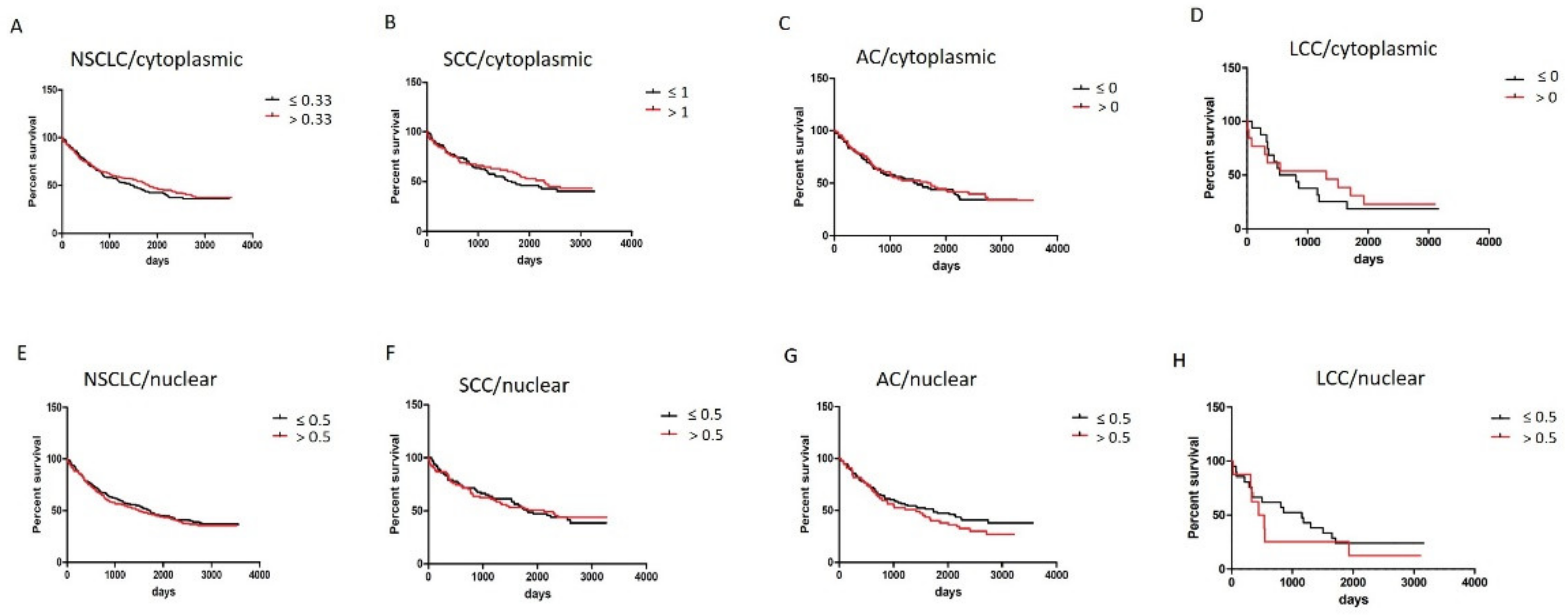
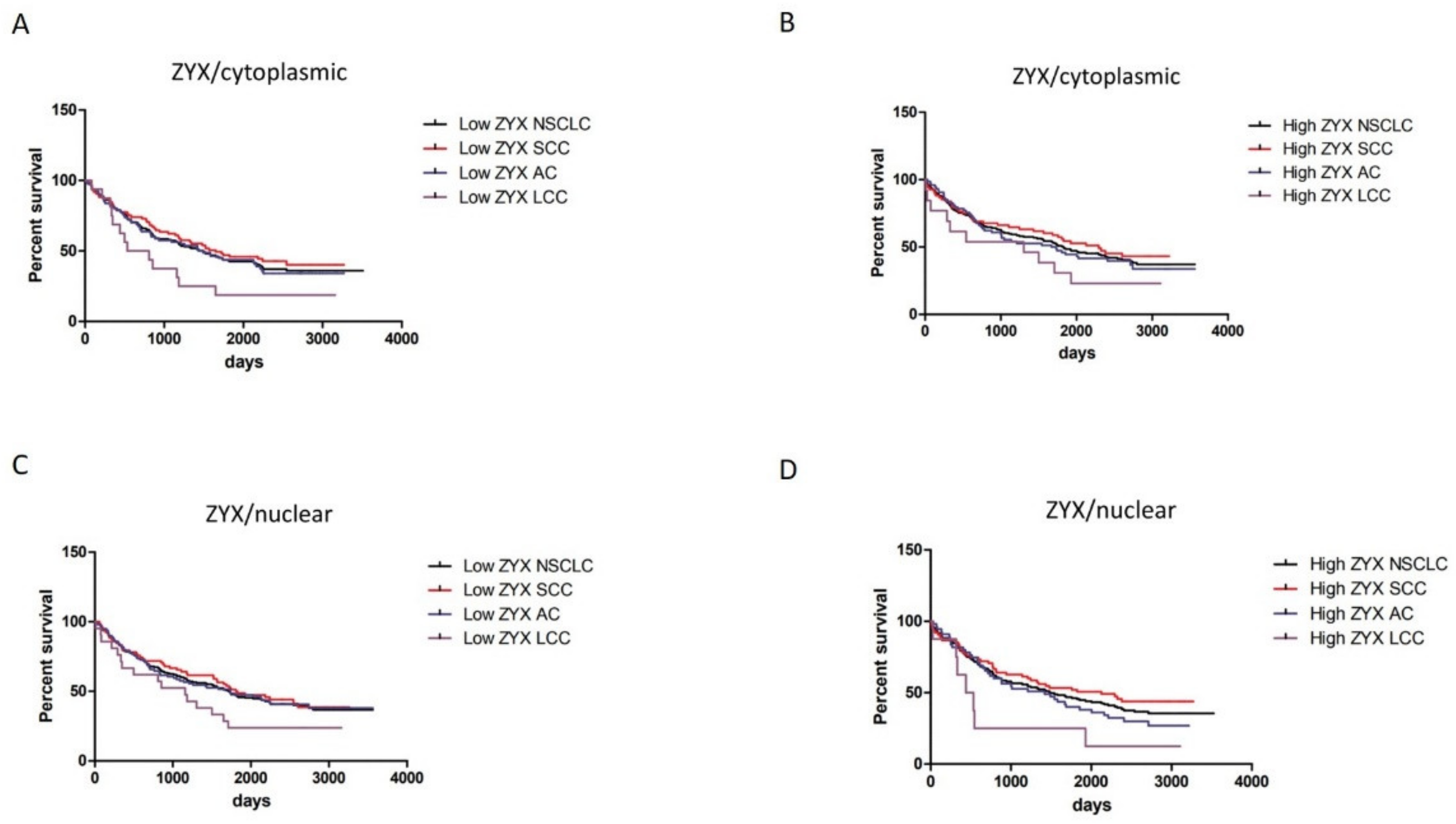
3.3. Survival Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oser, M.G.; Niederst, M.J.; Sequist, L.V.; Engelman, J.A. Transformation from Non-Small-Cell Lung Cancer to Small-Cell Lung Cancer: Molecular Drivers and Cells of Origin. Lancet Oncol. 2015, 16, e165–e172. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Fillmore, C.M.; Hammerman, P.S.; Kim, C.F.; Wong, K.-K. Non-Small-Cell Lung Cancers: A Heterogeneous Set of Diseases. Nat. Rev. Cancer 2014, 14, 535–546. [Google Scholar] [CrossRef]
- De Groot, P.M.; Wu, C.C.; Carter, B.W.; Munden, R.F. The Epidemiology of Lung Cancer. Transl. Lung Cancer Res. 2018, 7, 220–233. [Google Scholar] [CrossRef]
- Smith, M.A.; Hoffman, L.M.; Beckerle, M.C. LIM Proteins in Actin Cytoskeleton Mechanoresponse. Trends Cell Biol. 2014, 24, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Tatsumi, H.; Sokabe, M. Zyxin Emerges as a Key Player in the Mechanotransduction at Cell Adhesive Structures. Commun. Integr. Biol. 2008, 1, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Kollar, B.; Nahar, T.; Babu, S.S.; Wojtowicz, A.; Sticht, C.; Gretz, N.; Wagner, A.H.; Korff, T.; Hecker, M. Loss of the Mechanotransducer Zyxin Promotes a Synthetic Phenotype of Vascular Smooth Muscle Cells. J. Am. Heart Assoc. 2015, 4, e001712. [Google Scholar] [CrossRef] [Green Version]
- Hervy, M.; Hoffman, L.M.; Jensen, C.C.; Smith, M.; Beckerle, M.C. The LIM Protein Zyxin Binds CARP-1 and Promotes Apoptosis. Genes Cancer 2010, 1, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Youn, H.; Kim, E.-J.; Um, S.-J. Zyxin Cooperates with PTOV1 to Confer Retinoic Acid Resistance by Repressing RAR Activity. Cancer Lett. 2013, 331, 192–199. [Google Scholar] [CrossRef]
- Choi, Y.-H.; McNally, B.T.; Igarashi, P. Zyxin Regulates Migration of Renal Epithelial Cells through Activation of Hepatocyte Nuclear Factor-1β. Am. J. Physiol. Renal Physiol. 2013, 305, F100–F110. [Google Scholar] [CrossRef] [Green Version]
- Janssen, H.; Marynen, P. Interaction Partners for Human ZNF384/CIZ/NMP4—Zyxin as a Mediator for P130CAS Signaling? Exp. Cell Res. 2006, 312, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Li, P.; Yang, Z.; Huang, X.; Wei, G.; Sun, Y.; Kang, X.; Hu, X.; Deng, Q.; Chen, L.; et al. Zyxin Regulates Endothelial von Willebrand Factor Secretion by Reorganizing Actin Filaments around Exocytic Granules. Nat. Commun. 2017, 8, 14639. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Nakagami, H.; Koibuchi, N.; Miura, K.; Takami, Y.; Koriyama, H.; Hayashi, H.; Sabe, H.; Mochizuki, N.; Morishita, R.; et al. Zyxin Mediates Actin Fiber Reorganization in Epithelial-Mesenchymal Transition and Contributes to Endocardial Morphogenesis. Mol. Biol. Cell 2009, 20, 3115–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Cheng, H.; Gao, R.; Mu, C.; Chen, L.; Wu, S.; Chen, Q.; Zhu, Y. Zyxin-Siah2-Lats2 Axis Mediates Cooperation between Hippo and TGF-β Signalling Pathways. Nat. Commun. 2016, 7, 11123. [Google Scholar] [CrossRef] [Green Version]
- Zhong, C.; Yu, J.; Li, D.; Jiang, K.; Tang, Y.; Yang, M.; Shen, H.; Fang, X.; Ding, K.; Zheng, S.; et al. Zyxin as a Potential Cancer Prognostic Marker Promotes the Proliferation and Metastasis of Colorectal Cancer Cells. J. Cell. Physiol. 2019, 234, 15775–15789. [Google Scholar] [CrossRef]
- Sy, S.M.-H.; Lai, P.B.-S.; Pang, E.; Wong, N.L.-Y.; To, K.-F.; Johnson, P.J.; Wong, N. Novel Identification of Zyxin Upregulations in the Motile Phenotype of Hepatocellular Carcinoma. Mod. Pathol. 2006, 19, 1108–1116. [Google Scholar] [CrossRef]
- Kotb, A.; Hyndman, M.E.; Patel, T.R. The Role of Zyxin in Regulation of Malignancies. Heliyon 2018, 4, e00695. [Google Scholar] [CrossRef] [Green Version]
- Bernusso, V.A.; Machado-Neto, J.A.; Pericole, F.V.; Vieira, K.P.; Duarte, A.S.S.; Traina, F.; Hansen, M.D.; Saad, S.T.O.; Barcellos, K.S.A. Imatinib Restores VASP Activity and Its Interaction with Zyxin in BCR-ABL Leukemic Cells. Biochim. Biophys. Acta 2015, 1853, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ling, N.; Bai, Y.; Dong, W.; Hui, G.-Z.; Liu, D.; Zhao, J.; Hu, J. MiR-16-1 Plays a Role in Reducing Migration and Invasion of Glioma Cells. Anat. Rec. 2013, 296, 427–432. [Google Scholar] [CrossRef]
- Cadinu, D.; Hooda, J.; Alam, M.M.; Balamurugan, P.; Henke, R.M.; Zhang, L. Comparative Proteomic Analysis Reveals Characteristic Molecular Changes Accompanying the Transformation of Nonmalignant to Cancer Lung Cells. EuPA Open Proteom. 2014, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mise, N.; Savai, R.; Yu, H.; Schwarz, J.; Kaminski, N.; Eickelberg, O. Zyxin Is a Transforming Growth Factor-β (TGF-β)/Smad3 Target Gene That Regulates Lung Cancer Cell Motility via Integrin A5β1. J. Biol. Chem. 2012, 287, 31393–31405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Liu, S.; Qiao, Z.; Shang, Z.; Xia, Z.; Niu, X.; Qian, L.; Zhang, Y.; Fan, L.; Cao, C.-X.; et al. Systematic Comparison of Exosomal Proteomes from Human Saliva and Serum for the Detection of Lung Cancer. Anal. Chim. Acta 2017, 982, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Sertamo, K.; Pierrard, M.-A.; Mesmin, C.; Kim, S.Y.; Schlesser, M.; Berchem, G.; Domon, B. Verification of the Biomarker Candidates for Non-Small-Cell Lung Cancer Using a Targeted Proteomics Approach. J. Proteome Res. 2015, 14, 1412–1419. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.M.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 World Health Organization Classification of Lung Tumors: Impact of Genetic, Clinical and Radiologic Advances Since the 2004 Classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detterbeck, F.C.; Boffa, D.J.; Kim, A.W.; Tanoue, L.T. The Eighth Edition Lung Cancer Stage Classification. Chest 2017, 151, 193–203. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Recommendation for Uniform Definition of an Immunoreactive Score (IRS) for Immunohistochemical Estrogen Receptor Detection (ER-ICA) in Breast Cancer Tissue. Pathologe 1987, 8, 138–140. [Google Scholar]
- Dziegiel, P.; Salwa-Zurawska, W.; Zurawski, J.; Wojnar, A.; Zabel, M. Prognostic Significance of Augmented Metallothionein (MT) Expression Correlated with Ki-67 Antigen Expression in Selected Soft Tissue Sarcomas. Histol. Histopathol. 2005, 20, 83–89. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Partynska, A.; Gomulkiewicz, A.; Dziegiel, P.; Podhorska-Okolow, M. The Role of Zyxin in Carcinogenesis. Anticancer Res. 2020, 40, 5981–5988. [Google Scholar] [CrossRef]
- Yu, Y.P.; Luo, J.-H. Myopodin-Mediated Suppression of Prostate Cancer Cell Migration Involves Interaction with Zyxin. Cancer Res. 2006, 66, 7414–7419. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Carbayo, M.; Socci, N.D.; Charytonowicz, E.; Lu, M.; Prystowsky, M.; Childs, G.; Cordon-Cardo, C. Molecular Profiling of Bladder Cancer Using CDNA Microarrays: Defining Histogenesis and Biological Phenotypes. Cancer Res. 2002, 62, 6973–6980. [Google Scholar] [PubMed]
- Shiina, H.; Igawa, M.; Urakami, S.; Shigeno, K.; Yoneda, T.; Terashima, M.; Deguchi, M.; Ribeiro-Filho, L.; Dahiya, R. Alterations of β- and γ-Catenin in N-Butyl-N-(-4-Hydroxybutyl)Nitrosamine-Induced Murine Bladder Cancer. Cancer Res. 2001, 61, 7101–7109. [Google Scholar] [PubMed]
- Degenhardt, Y.Y.; Silverstein, S. Interaction of Zyxin, a Focal Adhesion Protein, with the E6 Protein from Human Papillomavirus Type 6 Results in Its Nuclear Translocation. J. Virol. 2001, 75, 11791–11802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, H.-S.; Even-Ram, S.; Kleinman, H.K.; Cha, H.-J. Zyxin Is Upregulated in the Nucleus by Thymosin β4 in SiHa Cells. Exp. Cell Res. 2006, 312, 3425–3431. [Google Scholar] [CrossRef]
- Benedit, P.; Paciucci, R.; Thomson, T.M.; Valeri, M.; Nadal, M.; Càceres, C.; de Torres, I.; Estivill, X.; Lozano, J.J.; Morote, J.; et al. PTOV1, a Novel Protein Overexpressed in Prostate Cancer Containing a New Class of Protein Homology Blocks. Oncogene 2001, 20, 1455–1464. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, T.G.P.; Kammerer, U.; Winkler, C.; Schindler, D.; Sickmann, A.; Honig, A.; Butt, E. Overexpression of LASP-1 Mediates Migration and Proliferation of Human Ovarian Cancer Cells and Influences Zyxin Localisation. Br. J. Cancer 2007, 96, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Grunewald, T.G.P.; Kammerer, U.; Schulze, E.; Schindler, D.; Honig, A.; Zimmer, M.; Butt, E. Silencing of LASP-1 Influences Zyxin Localization, Inhibits Proliferation and Reduces Migration in Breast Cancer Cells. Exp. Cell Res. 2006, 312, 974–982. [Google Scholar] [CrossRef]
- Butt, E.; Raman, D. New Frontiers for the Cytoskeletal Protein LASP1. Front. Oncol. 2018, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Xiaochuan, L.; Jiangyong, Y.; Ping, Z.; Xiaonan, W.; Lin, L. Clinical Characteristics and Prognosis of Pulmonary Large Cell Carcinoma: A Population-Based Retrospective Study Using SEER Data. Thorac. Cancer 2020, 11, 1522–1532. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Feature | NSCLC | SCC | AC | LCC | ||||
|---|---|---|---|---|---|---|---|---|
| n = 399 | % | n = 169 | % | n = 168 | % | n = 31 | % | |
| Age | ||||||||
| ≤62 | 212 | 53.13% | ≤64 89 | 52.66% | ≤61 90 | 53.57% | ≤62 16 | 51.61% |
| >62 | 187 | 46.87% | >64 80 | 47.34% | >61 78 | 46.43% | >62 15 | 48.39% |
| Sex | ||||||||
| Female | 116 | 29.07% | 32 | 18.93% | 65 | 38.69% | 6 | 19.35% |
| Male | 283 | 70.93% | 137 | 81.07% | 103 | 61.31% | 25 | 80.65% |
| Histological grade | ||||||||
| G1 | 24 | 6.02% | 4 | 2.37% | 19 | 11.31% | 1 | 3.23% |
| G2 | 293 | 73.43% | 143 | 84.62% | 109 | 64.88% | 24 | 77.42% |
| G3 | 66 | 16.54% | 22 | 13.02% | 38 | 22.62% | 6 | 19.35% |
| No data | 16 | 4.01% | 0 | 0.00% | 2 | 1.19% | 0 | 0.00% |
| Tumor size | ||||||||
| T1 | 88 | 22.06% | 39 | 23.08% | 33 | 19.64% | 8 | 25.81% |
| T2 | 186 | 46.62% | 76 | 44.97% | 86 | 51.19% | 13 | 41.94% |
| T3 | 79 | 19.80% | 37 | 21.89% | 31 | 18.45% | 6 | 19.35% |
| T4 | 46 | 11.53% | 17 | 10.06% | 18 | 10.71% | 4 | 12.90% |
| Lymph node metastases | ||||||||
| N0 | 260 | 65.16% | 110 | 65.09% | 108 | 64.29% | 19 | 61.29% |
| N1 | 70 | 17.54% | 40 | 23.67% | 22 | 13.10% | 4 | 12.90% |
| N2 | 69 | 17.29% | 19 | 11.24% | 38 | 22.62% | 8 | 25.81% |
| Distant metastases | ||||||||
| M0 | 395 | 99.00% | 169 | 100.00% | 165 | 98.21% | 30 | 96.77% |
| M1 | 4 | 1.00% | 0 | 0.00% | 3 | 1.79% | 1 | 3.23% |
| Clinical stage | ||||||||
| I | 145 | 36.34% | 64 | 37.87% | 62 | 36.90% | 7 | 22.58% |
| II | 130 | 32.58% | 64 | 37.87% | 46 | 27.38% | 12 | 38.71% |
| III | 120 | 30.08% | 41 | 24.26% | 57 | 33.93% | 11 | 35.48% |
| IV | 4 | 1.00% | 0 | 0.00% | 3 | 1.79% | 1 | 3.23% |
| Smoking | ||||||||
| Smokers | 336 | 84.21% | 157 | 92.90% | 128 | 76.19% | 29 | 93.55% |
| Non-smokers | 63 | 15.79% | 12 | 7.10% | 40 | 23.81% | 2 | 6.45% |
| Living in urban areas | ||||||||
| Yes | 31 | 7.77% | 16 | 9.47% | 10 | 5.95% | 1 | 3.23% |
| No | 368 | 92.23% | 153 | 90.53% | 158 | 94.05% | 30 | 96.77% |
| Death | ||||||||
| Yes | 244 | 61.15% | 92 | 54.44% | 108 | 64.29% | 24 | 77.42% |
| No | 147 | 36.84% | 74 | 43.79% | 59 | 35.12% | 6 | 19.35% |
| No data | 8 | 2.01% | 3 | 1.78% | 1 | 0.60% | 1 | 3.23% |
| Points | Percentage of Cells with Positive Nuclear Reaction |
|---|---|
| 0 | 0% |
| 1 | ≤10% |
| 2 | 11–25% |
| 3 | 26–50% |
| 4 | >50% |
| A | ||||||
|---|---|---|---|---|---|---|
| Overall Survival | ||||||
| NSCLC | ||||||
| Univariate | Multivariate | |||||
| Clinical Feature | p Value | HR | Confidence Interval 95% (HR) | p Value | HR | Confidence Interval 95% (HR) |
| Age | ||||||
| ≤62 vs. >62 | 0.005911 | 1.459308 | 1.114989–1.909957 | 0.001879 | 1.540898 | 1.173239–2.023770 |
| Sex | ||||||
| Female vs. male | 0.000262 | 1.820726 | 1.319828–2.511725 | 0.000222 | 1.845819 | 1.333147–2.555644 |
| Smoking | ||||||
| No vs. Yes | 0.164272 | 1.325817 | 0.891007–1.972812 | |||
| Living in | ||||||
| urban areas | ||||||
| No vs. Yes | 0.053255 | 1.575759 | 0.993642–2.498906 | |||
| Clinical stage | ||||||
| I–II vs. III–IV | 0.000000 | 2.351361 | 1.782315–3.102088 | 0.009199 | 1.702469 | 1.140750–2.540783 |
| Histological grade | ||||||
| G1–G2 vs. G3 | 0.035460 | 1.441683 | 1.025203–2.027353 | 0.011715 | 1.564191 | 1.104626–2.214952 |
| pT | ||||||
| pT1-pT2 vs. | ||||||
| pT3-pT4 | 0.000000 | 2.161526 | 1.639607–2.849582 | 0.008051 | 1.559392 | 1.122639–2.166059 |
| pN | ||||||
| N0 vs. N1–N2 | 0.000564 | 1.621062 | 1.231833–2.133276 | 0.163298 | 1.285130 | 0.903181–1.828603 |
| p63 | ||||||
| ≤25 vs. >25% | 0.097096 | 0.796109 | 0.608116–1.042217 | |||
| TTF-1 | ||||||
| ≤25 vs. >25% | 0.936561 | 0.989066 | 0.754490–1.296575 | |||
| Ki-67 | ||||||
| ≤25 vs. >25% | 0.968136 | 0.994309 | 0.751467–1.315628 | |||
| Cytoplasmic | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| High vs. Low | 0.506478 | 1.095422 | 0.837153–1.433368 | |||
| Nuclear | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| Low vs. High | 0.649726 | 1.064851 | 0.811928–1.396562 | |||
| B | ||||||
| Overall Survival | ||||||
| SCC | ||||||
| Univariate | Multivariate | |||||
| Clinical Feature | p Value | HR | Confidence Interval 95% (HR) | p Value | HR | Confidence Interval 95% (HR) |
| Age | ||||||
| ≤64 vs. >64 | 0.083694 | 1.456415 | 0.951168–2.230043 | |||
| Sex | ||||||
| Female vs. male | 0.121189 | 1.595476 | 0.883699–2.880554 | |||
| Smoking | ||||||
| No vs. Yes | 0.748266 | 1.159491 | 0.469650–2.862597 | |||
| Living in | ||||||
| urban areas | ||||||
| No vs. Yes | 0.502217 | 1.266994 | 0.634712–2.529139 | |||
| Clinical stage | ||||||
| I–II vs. III–IV | 0.001125 | 2.128965 | 1.351164–3.354511 | 0.247973 | 1.390731 | 0.794747–2.433645 |
| Histological grade | ||||||
| G1–G2 vs. G3 | 0.000091 | 2.997641 | 1.729638–5.195220 | 0.002834 | 2.414826 | 1.353633–4.307953 |
| pT | ||||||
| pT1-pT2 vs. | ||||||
| pT3-pT4 | 0.000534 | 2.148663 | 1.393743–3.312486 | 0.049517 | 1.682302 | 1.001099–2.827031 |
| pN | ||||||
| N0 vs. N1–N2 | 0.908460 | 1.026365 | 0.658646–1.599380 | |||
| p63 | ||||||
| ≤25 vs. >25% | 0.365148 | 0.767183 | 0.432303–1.361474 | |||
| TTF-1 | ||||||
| ≤25 vs. >25% | 0.546339 | 1.199180 | 0.664646–2.163608 | |||
| Ki-67 | ||||||
| ≤25 vs. >25% | 0.836690 | 0.956480 | 0.626516–1.460224 | |||
| Cytoplasmic | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| High vs. Low | 0.603775 | 1.119886 | 0.730239–1.717444 | |||
| Nuclear | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| Low vs. High | 0.823766 | 0.953000 | 0.623872–1.455763 | |||
| C | ||||||
| Overall Survival | ||||||
| AC | ||||||
| Univariate | Multivariate | |||||
| Clinical Feature | p Value | HR | Confidence Interval 95% (HR) | p Value | HR | Confidence Interval 95% (HR) |
| Age | ||||||
| ≤61 vs. >61 | 0.439639 | 1.170082 | 0.785611–1.742711 | |||
| Sex | ||||||
| Female vs. male | 0.000411 | 2.195089 | 1.419128–3.395335 | 0.000119 | 2.371420 | 1.527428–3.681766 |
| Smoking | ||||||
| No vs. Yes | 0.331944 | 1.270446 | 0.783337–2.060456 | |||
| Living in | ||||||
| urban areas | ||||||
| No vs. Yes | 0.029915 | 2.244604 | 1.081842–4.657101 | 0.029913 | 2.314151 | 1.085072–4.935430 |
| Clinical stage | ||||||
| I–II vs. III–IV | 0.000000 | 2.922240 | 1.945881–4.388492 | 0.364952 | 1.311295 | 0.729573–2.356852 |
| Histological grade | ||||||
| G1–G2 vs. G3 | 0.880118 | 1.037945 | 0.639707–1.684099 | |||
| pT | ||||||
| pT1-pT2 vs. | ||||||
| pT3-pT4 | 0.000001 | 2.846085 | 1.873553–4.323444 | 0.002122 | 2.191669 | 1.328609–3.615369 |
| pN | ||||||
| N0 vs. N1–N2 | 0.000000 | 2.883628 | 1.922023–4.326332 | 0.001392 | 2.317220 | 1.384130–3.879337 |
| p63 | ||||||
| ≤25 vs. >25% | 0.907950 | 1.032147 | 0.603679–1.764726 | |||
| TTF-1 | ||||||
| ≤25 vs. >25% | 0.133316 | 0.720412 | 0.469498–1.105423 | |||
| Ki-67 | ||||||
| ≤25 vs. >25% | 0.894629 | 1.033806 | 0.632080–1.690855 | |||
| Cytoplasmic | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| High vs. Low | 0.758237 | 1.064633 | 0.714493–1.586360 | |||
| Nuclear | ||||||
| zyxin levels | ||||||
| in cancer cells | ||||||
| Low vs. High | 0.246701 | 1.271330 | 0.846950–1.908353 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Partynska, A.; Gomulkiewicz, A.; Piotrowska, A.; Grzegrzolka, J.; Rzechonek, A.; Ratajczak-Wielgomas, K.; Podhorska-Okolow, M.; Dziegiel, P. Expression of Zyxin in Non-Small Cell Lung Cancer—A Preliminary Study. Biomolecules 2022, 12, 827. https://doi.org/10.3390/biom12060827
Partynska A, Gomulkiewicz A, Piotrowska A, Grzegrzolka J, Rzechonek A, Ratajczak-Wielgomas K, Podhorska-Okolow M, Dziegiel P. Expression of Zyxin in Non-Small Cell Lung Cancer—A Preliminary Study. Biomolecules. 2022; 12(6):827. https://doi.org/10.3390/biom12060827
Chicago/Turabian StylePartynska, Aleksandra, Agnieszka Gomulkiewicz, Aleksandra Piotrowska, Jedrzej Grzegrzolka, Adam Rzechonek, Katarzyna Ratajczak-Wielgomas, Marzenna Podhorska-Okolow, and Piotr Dziegiel. 2022. "Expression of Zyxin in Non-Small Cell Lung Cancer—A Preliminary Study" Biomolecules 12, no. 6: 827. https://doi.org/10.3390/biom12060827