
Genes Responsible for H2S Production and Metabolism Are Involved in Learning and Memory in Drosophila melanogaster

 ,
,  , ,
, ,
Abstract
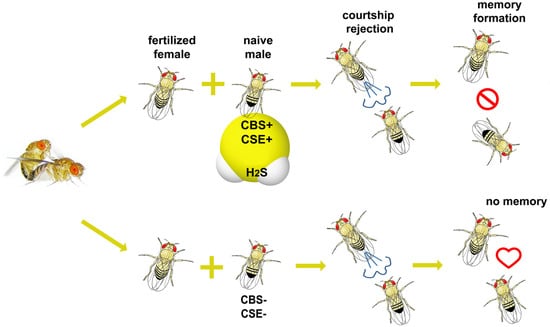
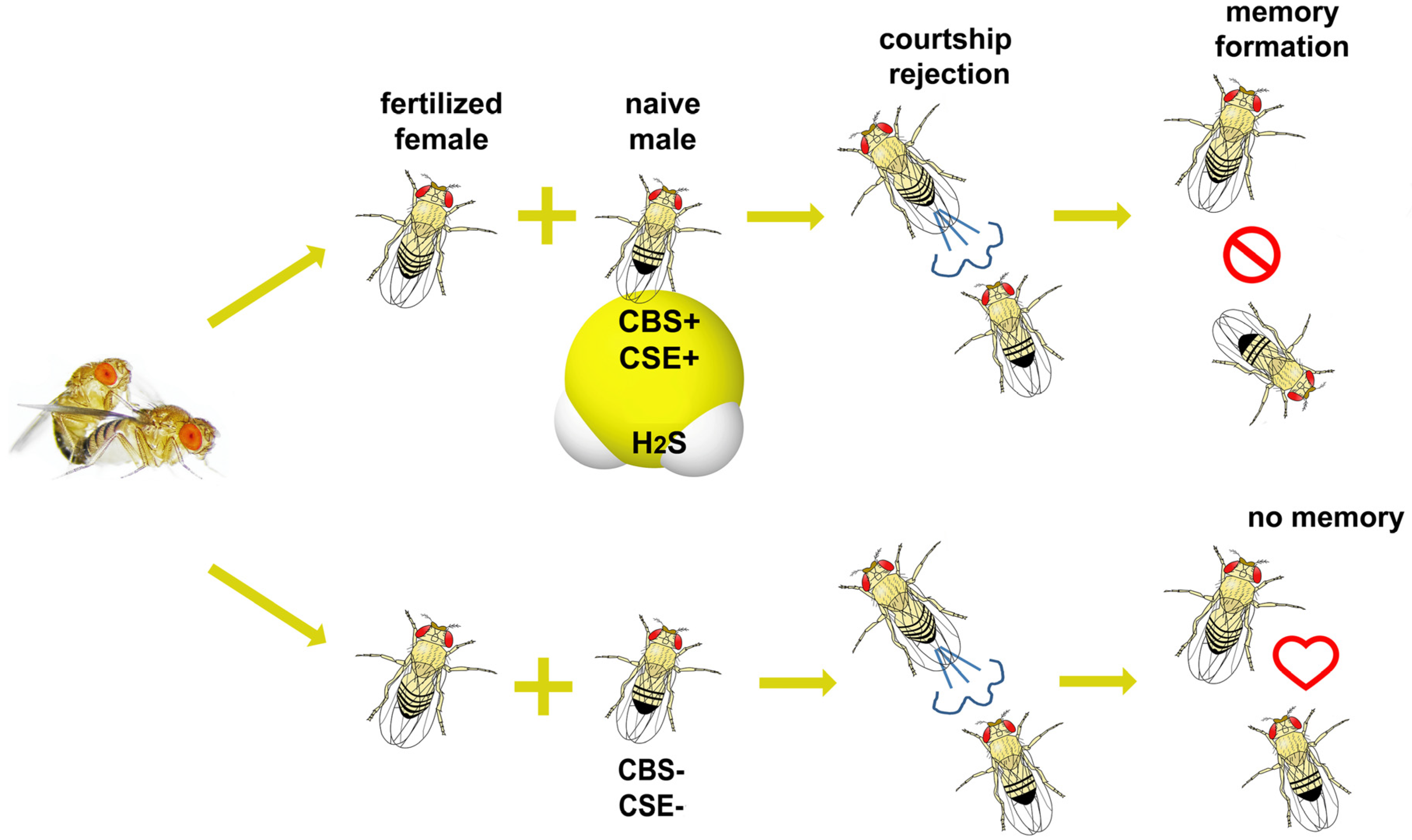
:
1. Introduction
2. Materials and Methods
2.1. Fly Strains and Maintenance
2.2. Test for Learning and Memory of Flies in Conditioned Courtship Suppression Paradigm
2.3. Olfactory Behavior Assay
2.4. RNA Extraction from Fly Heads for Library Preparations
2.5. cDNA Library Preparation and Data Analysis
2.6. Quantitative Real-Time PCR
2.7. Amino Acid Quantitative Analysis
3. Results
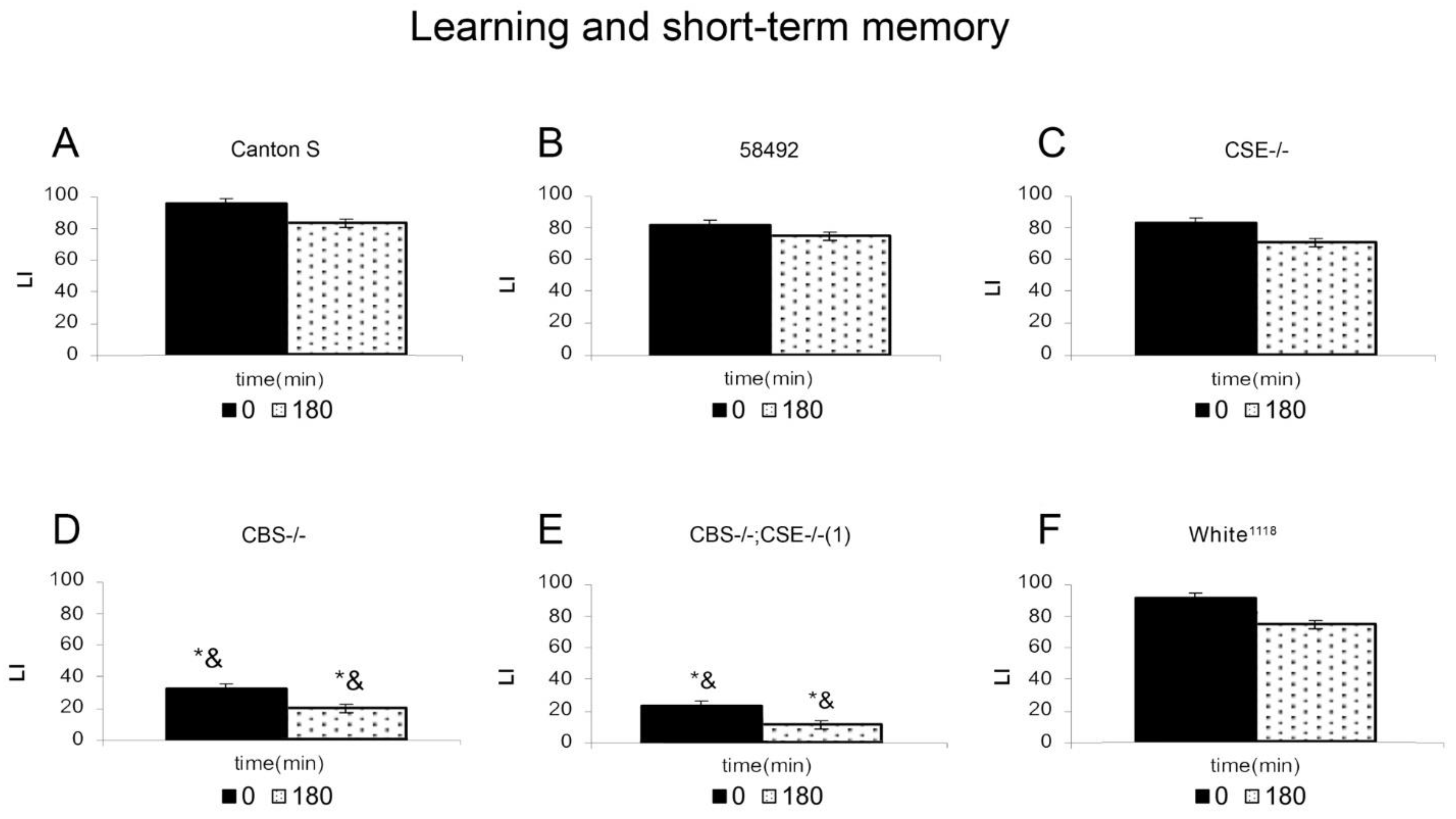
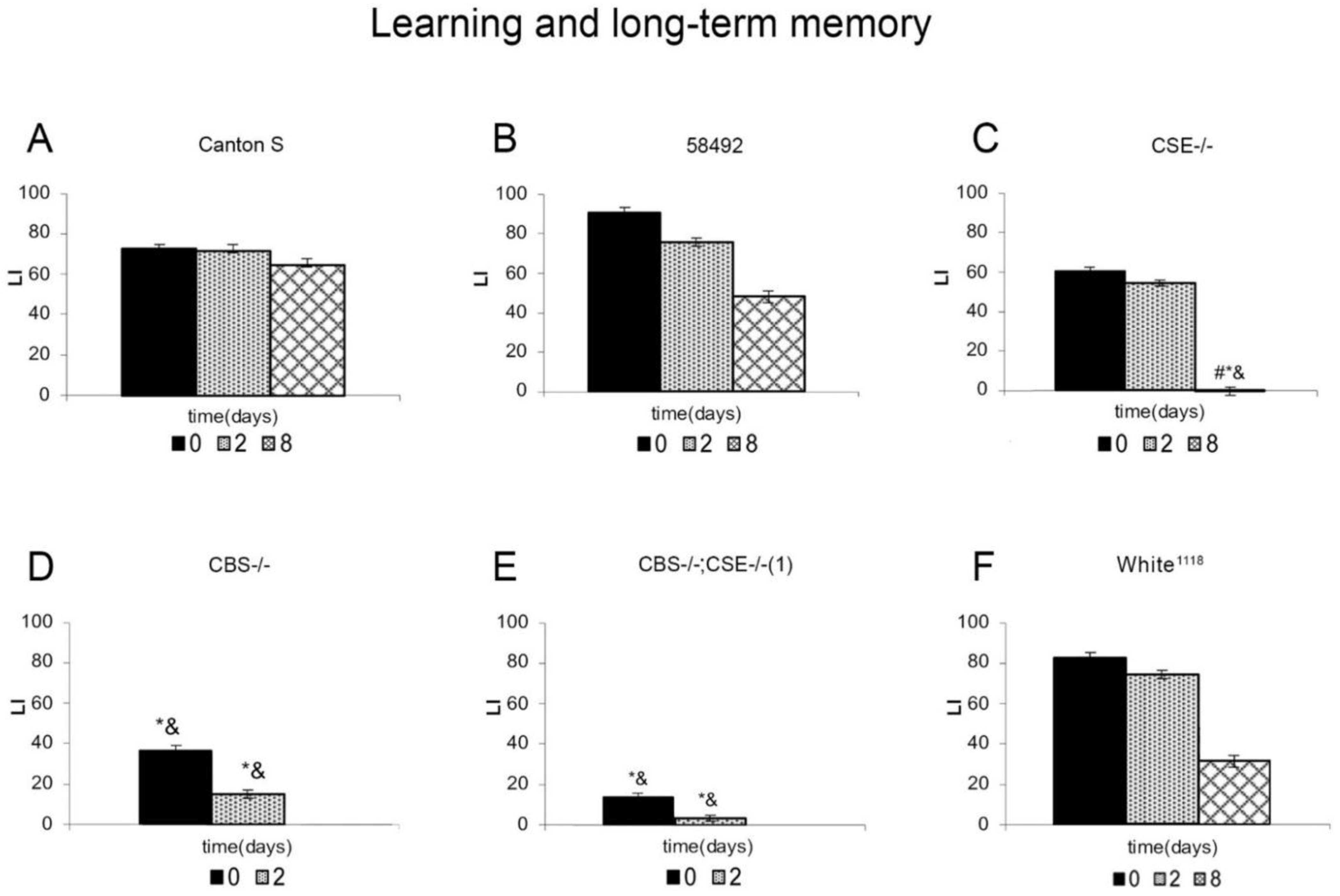
3.1. Learning and Memory of Flies in Conditioned Courtship Suppression Paradigm
3.2. The Monitoring of Olfactory Abilities of the Strains Used in the Study
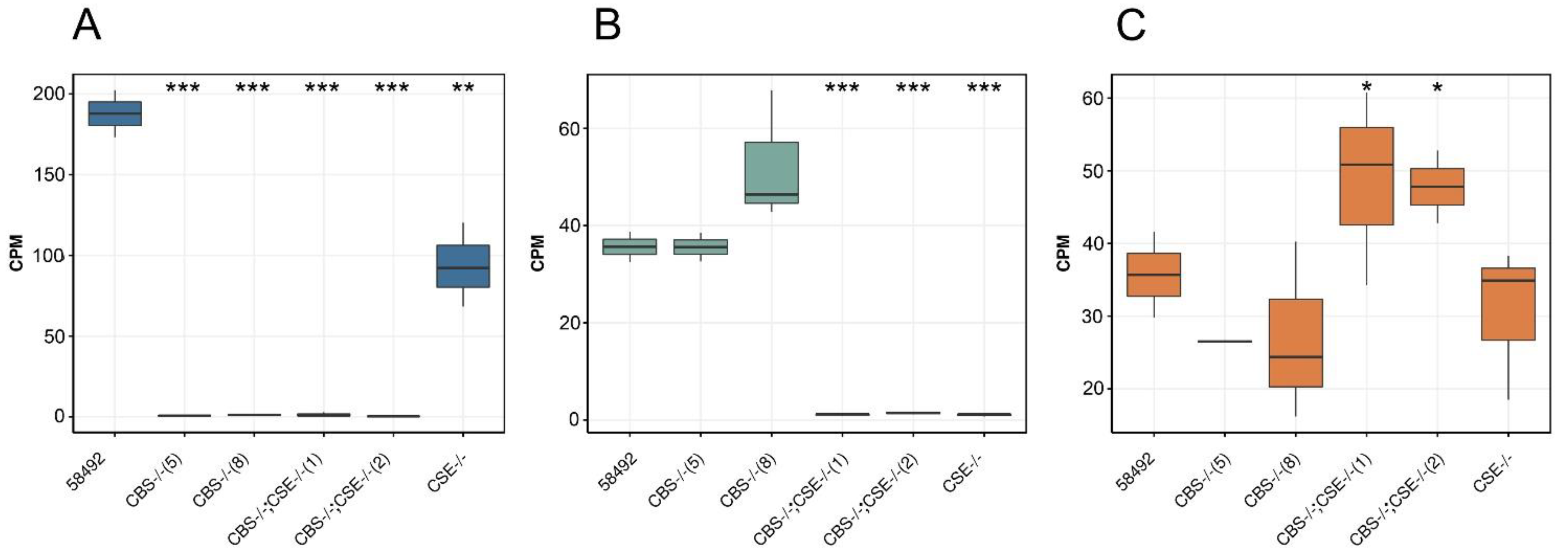
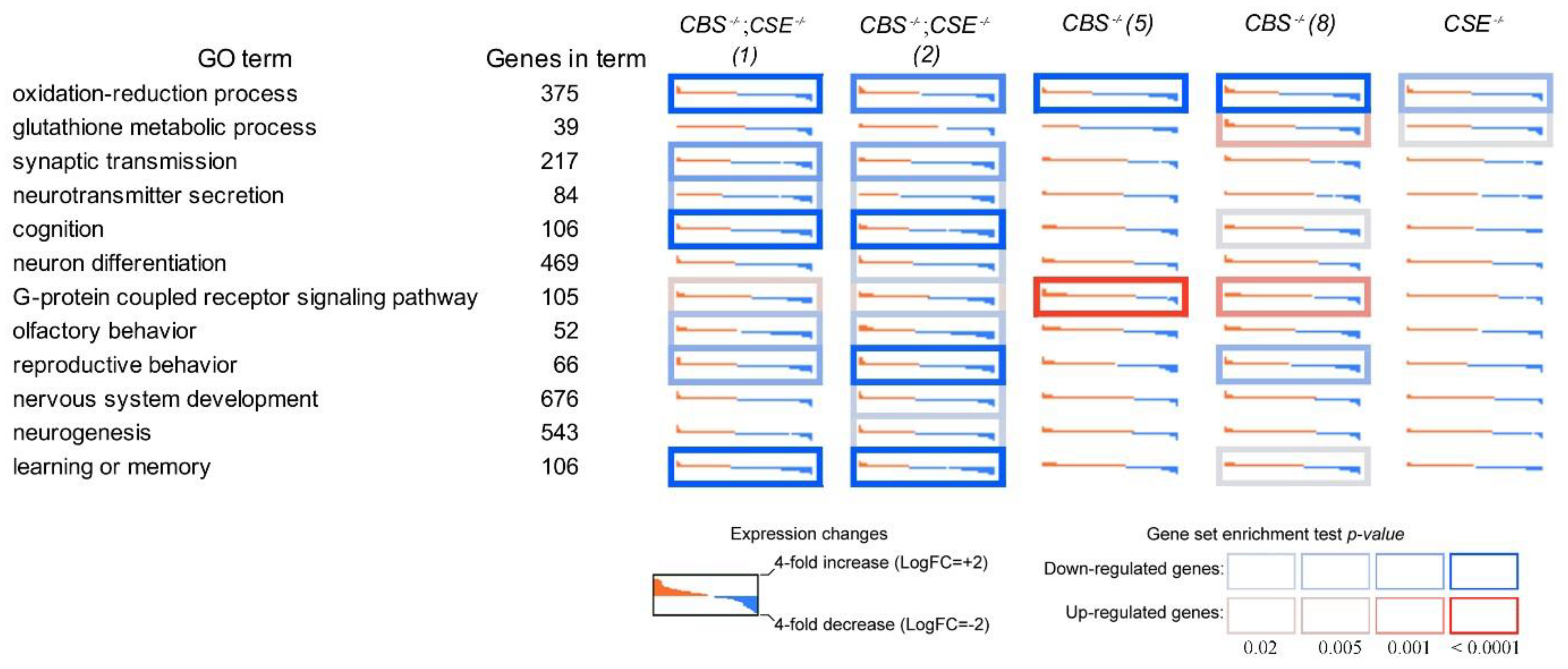
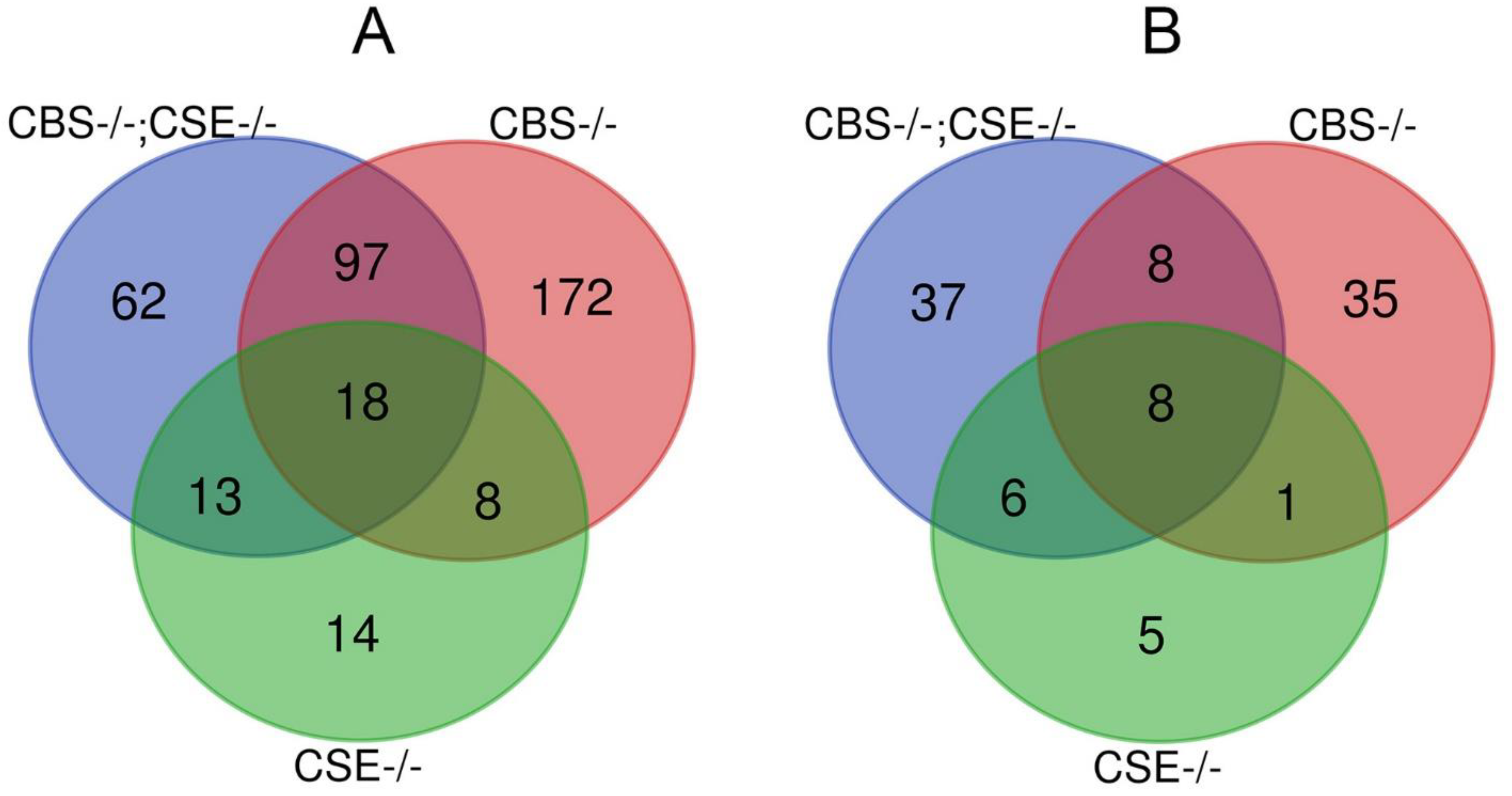
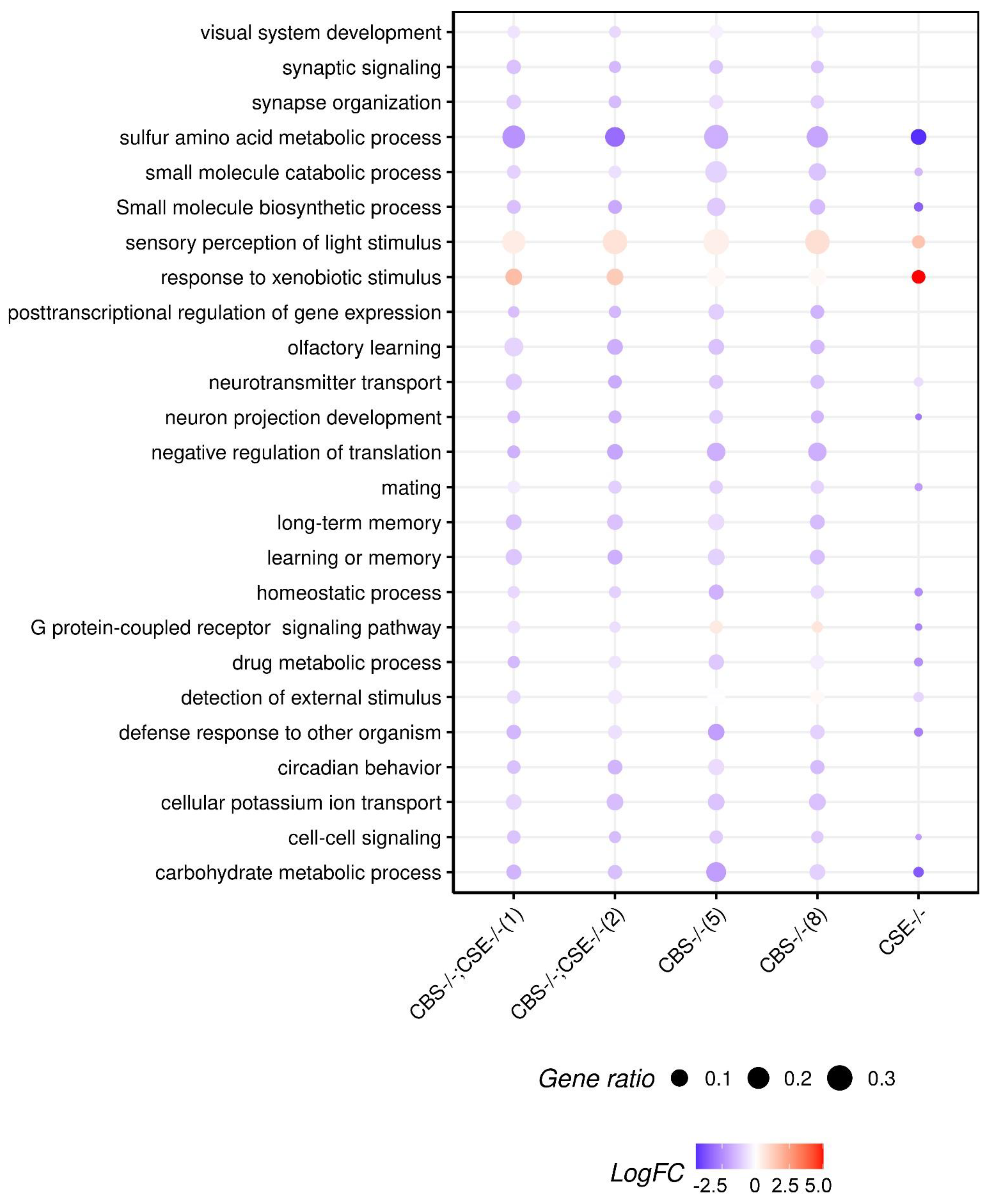
3.3. Transcriptome Analysis of the Strains with Deletions
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide and polysulfides as signaling molecules. Proc. Jpn. Acad. Ser. B 2015, 91, 131–159. [Google Scholar] [CrossRef] [Green Version]
- Paul, B.D.; Snyder, S.H. Gasotransmitter hydrogen sulfide signaling in neuronal health and disease. Biochem. Pharmacol. 2018, 149, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, H. Hydrogen Sulfide and Polysulfides as Biological Mediators. Molecules 2014, 19, 16146–16157. [Google Scholar] [CrossRef] [Green Version]
- Filipovic, M.R. Persulfidation (S-sulfhydration) and H2S. Handb. Exp. Pharmacol. 2015, 230, 29–59. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Enzymology of H2S Biogenesis, Decay and Signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Giovinazzo, D.; Bursac, B.; Sbodio, J.I.; Nalluru, S.; Vignane, T.; Snowman, A.M.; Albacarys, L.M.; Sedlak, T.W.; Torregrossa, R.; Whiteman, M.; et al. Hydrogen sulfide is neuroprotective in Alzheimer’s disease by sulfhydrating GSK3β and inhibiting Tau hyperphosphorylation. Proc. Natl. Acad. Sci. USA 2021, 118, e2017225118. [Google Scholar] [CrossRef]
- Peng, S.-Y.; Wu, X.; Lu, T.; Cui, G.; Chen, G. Research progress of hydrogen sulfide in Alzheimer′s disease from laboratory to hospital: A narrative review. Med. Gas Res. 2020, 10, 125–129. [Google Scholar] [CrossRef]
- Kshirsagar, V.; Thingore, C.; Gursahani, M.; Gawali, N.; Juvekar, A. Hydrogen Sulfide Ameliorates Lipopolysaccharide-Induced Memory Impairment in Mice by Reducing Apoptosis, Oxidative, and Inflammatory Effects. Neurotox. Res. 2021, 39, 1310–1322. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Tang, Y.-Y.; Jiang, L.; Lan, F.; Li, X.; Zhang, P.; Zou, W.; Chen, Y.-J.; Tang, X.-Q. H2S Attenuates Sleep Deprivation-Induced Cognitive Impairment by Reducing Excessive Autophagy via Hippocampal Sirt-1 in WISTAR RATS. Neurochem. Res. 2021, 46, 1941–1952. [Google Scholar] [CrossRef] [PubMed]
- Vandini, E.; Ottani, A.; Zaffe, D.; Calevro, A.; Canalini, F.; Cavallini, G.M.; Rossi, R.; Guarini, S.; Giuliani, D. Mechanisms of Hydrogen Sulfide against the Progression of Severe Alzheimer’s Disease in Transgenic Mice at Different Ages. Pharmacology 2018, 103, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Chen, G.; Zhang, J.-Y.; Ding, Y.-P.; Wang, Z.; Kong, Y. Hydrogen sulfide therapy in brain diseases: From bench to bedside. Med Gas Res. 2017, 7, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Bashir, H.; Dekair, L.; Mahmoud, Y.; Ben-Omran, T. Neurodevelopmental and Cognitive Outcomes of Classical Homocystinuria: Experience from Qatar. JIMD Rep. 2014, 21, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Hoss, G.R.W.; Poloni, S.; Blom, H.J.; Schwartz, I.V.D. Three Main Causes of Homocystinuria: CBS, cblC and MTHFR Deficiency. What do they Have in Common? J. Inborn Errors Metab. Screen. 2019, 7, e20190007. [Google Scholar] [CrossRef]
- Keating, A.K.; Freehauf, C.; Jiang, H.; Brodsky, G.L.; Stabler, S.P.; Allen, R.H.; Graham, D.K.; Thomas, J.A.; Van Hove, J.L.; Maclean, K.N. Constitutive induction of pro-inflammatory and chemotactic cytokines in cystathionine beta-synthase deficient homocystinuria. Mol. Genet. Metab. 2011, 103, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Li, M.-H.; Tang, J.-P.; Zhang, P.; Li, X.; Wang, C.-Y.; Wei, H.-J.; Yang, X.-F.; Zou, W.; Tang, X.-Q. Disturbance of endogenous hydrogen sulfide generation and endoplasmic reticulum stress in hippocampus are involved in homocysteine-induced defect in learning and memory of rats. Behav. Brain Res. 2014, 262, 35–41. [Google Scholar] [CrossRef]
- Li, M.; Zhang, P.; Wei, H.-J.; Li, M.-H.; Zou, W.; Li, X.; Gu, H.-F.; Tang, X.-Q. Hydrogen Sulfide Ameliorates Homocysteine-Induced Cognitive Dysfunction by Inhibition of Reactive Aldehydes Involving Upregulation of ALDH2. Int. J. Neuropsychopharmacol. 2016, 20, 305–315. [Google Scholar] [CrossRef]
- Flores-Flores, M.; Moreno-García, L.; Castro-Martínez, F.; Nahmad, M. Cystathionine β-synthase Deficiency Impairs Vision in the Fruit Fly, Drosophila melanogaster. Curr. Eye Res. 2020, 46, 600–605. [Google Scholar] [CrossRef]
- Kuntz, S.; Poeck, B.; Strauss, R. Visual Working Memory Requires Permissive and Instructive NO/cGMP Signaling at Presynapses in the Drosophila Central Brain. Curr. Biol. 2017, 27, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, E.B.; Layne, J.E.; Elchert, A.R.; Rollmann, S.M. Behavioral and Transcriptional Response to Selection for Olfactory Behavior in Drosophila. G3 Genes Genomes Genet. 2020, 10, 1283–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatsepina, O.; Karpov, D.; Chuvakova, L.; Rezvykh, A.; Funikov, S.; Sorokina, S.; Zakluta, A.; Garbuz, D.; Shilova, V.; Evgen’Ev, M. Genome-wide transcriptional effects of deletions of sulphur metabolism genes in Drosophila melanogaster. Redox Biol. 2020, 36, 101654. [Google Scholar] [CrossRef] [PubMed]
- Tully, T.; Boynton, S.; Brandes, C.; Dura, J.; Mihalek, R.; Preat, T.; Villella, A. Genetic Dissection of Memory Formation in Drosophila melanogaster. Cold Spring Harb. Symp. Quant. Biol. 1990, 55, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Akalal, D.-B.G.; Yu, D.; Davis, R.L. A Late-Phase, Long-Term Memory Trace Forms in the Neurons of Drosophila Mushroom Bodies after Olfactory Classical Conditioning. J. Neurosci. 2010, 30, 16699–16708. [Google Scholar] [CrossRef] [PubMed]
- Mery, F.; Belay, A.T.; So, A.K.-C.; Sokolowski, M.B.; Kawecki, T.J. Natural polymorphism affecting learning and memory in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 13051–13055. [Google Scholar] [CrossRef] [Green Version]
- Zhuravlev, A.V.; Nikitina, E.; Savvateeva-Popova, E.V. Learning and memory in Drosophila: Physiologic and genetic bases. Usp Fiziol Nauk. 2015, 46, 76–92. [Google Scholar]
- Shaposhnikov, M.V.; Zakluta, A.S.; Zemskaya, N.V.; Guvatova, Z.G.; Shilova, V.Y.; Yakovleva, D.V.; Gorbunova, A.A.; Koval, L.A.; Ulyasheva, N.S.; Evgen’Ev, M.B.; et al. Deletions of the cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE) genes, involved in the control of hydrogen sulfide biosynthesis, significantly affect lifespan and fitness components of Drosophila melanogaster. Mech. Ageing Dev. 2022, 203, 111656. [Google Scholar] [CrossRef]
- Kamyshev, N.G.; Iliadi, K.G.; Bragina, J.V. Drosophila Conditioned Courtship: Two Ways of Testing Memory. Learn. Mem. 1999, 6, 1–20. [Google Scholar] [CrossRef]
- Raun, N.; Jones, S.; Kramer, J.M. Conditioned courtship suppression in Drosophila melanogaster. J. Neurogenet. 2021, 35, 154–167. [Google Scholar] [CrossRef]
- Keleman, K.; Vrontou, E.; Krüttner, S.; Yu, J.Y.; Kurtovic-Kozaric, A.; Dickson, B.J. Dopamine neurons modulate pheromone responses in Drosophila courtship learning. Nature 2012, 489, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Kuzin, B.A.; Nikitina, E.A.; Cherezov, R.O.; Vorontsova, J.E.; Slezinger, M.S.; Zatsepina, O.G.; Simonova, O.B.; Enikolopov, G.N.; Savvateeva-Popova, E.V. Combination of Hypomorphic Mutations of the Drosophila Homologues of Aryl Hydrocarbon Receptor and Nucleosome Assembly Protein Family Genes Disrupts Morphogenesis, Memory and Detoxification. PLoS ONE 2014, 9, e94975. [Google Scholar] [CrossRef] [PubMed]
- Savvateeva-Popova, E.; Popov, A.; Grossman, A.; Nikitina, E.; Medvedeva, A.; Peresleni, A.; Korochkin, L.; Moe, J.G.; Davidowitz, E.; Pyatkov, K.; et al. Pathogenic chaperone-like RNA induces congophilic aggregates and facilitates neurodegeneration in Drosophila. Cell Stress Chaperon 2007, 12, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvateeva-Popova, E.; Popov, A.; Grossman, A.; Nikitina, E.; Medvedeva, A.; Molotkov, D.; Kamyshev, N.; Pyatkov, K.; Zatsepina, O.; Schostak, N.; et al. Non-coding RNA as a trigger of neuropathologic disorder phenotypes in transgenic Drosophila. J. Neural Transm. 2008, 115, 1629–1642. [Google Scholar] [CrossRef]
- Zatsepina, O.G.; Nikitina, E.A.; Shilova, V.Y.; Chuvakova, L.N.; Sorokina, S.; Vorontsova, J.E.; Tokmacheva, E.V.; Funikov, S.Y.; Rezvykh, A.P.; Evgen’Ev, M.B. Hsp70 affects memory formation and behaviorally relevant gene expression in Drosophila melanogaster. Cell Stress Chaperon 2021, 26, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Koemans, T.S.; Oppitz, C.; Donders, R.A.T.; van Bokhoven, H.; Schenck, A.; Keleman, K.; Kramer, J.M. Drosophila Courtship Conditioning as a Measure of Learning and Memory. J. Vis. Exp. 2017, e55808. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, M.; Rapp, P.R. The use of animal models to study the effects of aging on cognition. Annu. Rev. Psychol. 1997, 48, 339–370. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J.; Sokal, R.R. Biometry: The Principles and Practice of Statistics in Biological Research; Freeman: New York, NY, USA, 1981. [Google Scholar]
- Tully, T.; Quinn, W.G. Classical conditioning and retention in normal and mutant Drosophila melanogaster. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1985, 157, 263–277. [Google Scholar] [CrossRef]
- Zhou, X.; Yuan, C.; Guo, A. Drosophila Olfactory Response Rhythms Require Clock Genes but Not Pigment Dispersing Factor or Lateral Neurons. J. Biol. Rhythm. 2005, 20, 237–244. [Google Scholar] [CrossRef]
- Krasnov, G.S.; Dmitriev, A.A.; Kudryavtseva, A.V.; Shargunov, A.V.; Karpov, D.S.; Uroshlev, L.A.; Melnikova, N.V.; Blinov, V.M.; Poverennaya, E.V.; Archakov, A.I.; et al. PPLine: An Automated Pipeline for SNP, SAP, and Splice Variant Detection in the Context of Proteogenomics. J. Proteome Res. 2015, 14, 3729–3737. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and the novel “gene expression’s C T difference” formula. Klin. Wochenschr. 2006, 84, 901–910. [Google Scholar] [CrossRef]
- Ponton, F.; Chapuis, M.-P.; Pernice, M.; Sword, G.A.; Simpson, S.J. Evaluation of potential reference genes for reverse transcription-qPCR studies of physiological responses in Drosophila melanogaster. J. Insect Physiol. 2011, 57, 840–850. [Google Scholar] [CrossRef]
- Parkhitko, A.A.; Binari, R.; Zhang, N.; Asara, J.M.; Demontis, F.; Perrimon, N. Tissue-specific down-regulation of S-adenosyl-homocysteine via suppression of dAhcyL1/dAhcyL2 extends health span and life span in Drosophila. Genes Dev. 2016, 30, 1409–1422. [Google Scholar] [CrossRef] [Green Version]
- Ksenofontov, A.L.; Boyko, A.I.; Mkrtchyan, G.V.; Tashlitsky, V.N.; Timofeeva, A.V.; Graf, A.V.; Bunik, V.I.; Baratova, L.A. Analysis of free amino acids in mammalian brain extracts. Biochemistry 2017, 82, 1183–1192. [Google Scholar] [CrossRef]
- Kaminskaya, A.N.; Nikitina, E.A.; Medvedeva, A.V.; Gerasimenko, M.S.; Chernikova, D.A.; Savateeva-Popova, E.V. Influence of limk1 Gene Polymorphism on Learning Acquisition and Memory Formation with pCREB Distribution and Aggregate Formation in Neuromuscular Junctions in Drosophila melanogaster. Genetika 2015, 51, 685–693. [Google Scholar] [CrossRef]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate Sulfurtransferase Produces Hydrogen Sulfide and Bound Sulfane Sulfur in the Brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef]
- Kumar, M.; Tyagi, N.; Moshal, K.S.; Sen, U.; Pushpakumar, S.; Vacek, T.; Lominadze, D.; Tyagi, S.C. GABAA receptor agonist mitigates homocysteine-induced cerebrovascular remodeling in knockout mice. Brain Res. 2008, 1221, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, S. Disorders of transsulfuration. In The Metabolic and Molecular Bases of Inherited Disease; McGraw-Hill Professiona: New York, NY, USA, 2001. [Google Scholar] [CrossRef]
- Davis, M.Y.; Trinh, K.; Thomas, R.E.; Yu, S.; Germanos, A.A.; Whitley, B.N.; Sardi, S.P.; Montine, T.J.; Pallanck, L.J. Glucocerebrosidase Deficiency in Drosophila Results in α-Synuclein-Independent Protein Aggregation and Neurodegeneration. PLoS Genet. 2016, 12, e1005944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didelot, G.; Molinari, F.; Tchénio, P.; Comas, D.; Milhiet, E.; Munnich, A.; Colleaux, L.; Preat, T. Tequila, a Neurotrypsin Ortholog, Regulates Long-Term Memory Formation in Drosophila. Science 2006, 313, 851–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhakaran, I.P.; Hillebrand, J.; Dervan, A.; Das, S.; Holohan, E.E.; Hülsmeier, J.; Sarov, M.; Parker, R.; VijayRaghavan, K.; Ramaswami, M. FMRP and Ataxin-2 function together in long-term olfactory habituation and neuronal translational control. Proc. Natl. Acad. Sci. USA 2013, 111, E99–E108. [Google Scholar] [CrossRef] [Green Version]
- Levin, L.; Han, P.-L.; Hwang, P.M.; Feinstein, P.G.; Davis, R.; Reed, R.R. The Drosophila learning and memory gene rutabaga encodes a Ca2+calmodulin-responsive adenylyl cyclase. Cell 1992, 68, 479–489. [Google Scholar] [CrossRef]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative Contributions of Cystathionine β-Synthase and γ-Cystathionase to H2S Biogenesis via Alternative Trans-sulfuration Reactions. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef] [Green Version]
- Robert, K.; Vialard, F.; Thiery, E.; Toyama, K.; Sinet, P.-M.; Janel, N.; London, J. Expression of the Cystathionine β Synthase (CBS) Gene During Mouse Development and Immunolocalization in Adult Brain. J. Histochem. Cytochem. 2003, 51, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.-F.; Lu, M.; Hon Wong, P.T.; Bian, J.-S. Hydrogen Sulfide: Neurophysiology and Neuropathology. Antioxid. Redox Signal. 2011, 15, 405–419. [Google Scholar] [CrossRef]
- Enokido, Y.; Suzuki, E.; Iwasawa, K.; Namekata, K.; Okazawa, H.; Kimura, H. Cystathionine β-synthase, a key enzyme for homocysteine metabolism, is preferentially expressed in the radial glia/astrocyte lineage of developing mouse CNS. FASEB J. 2005, 19, 1854–1856. [Google Scholar] [CrossRef]
- Lee, M.; Schwab, C.; Yu, S.; McGeer, E.; McGeer, P.L. Astrocytes produce the antiinflammatory and neuroprotective agent hydrogen sulfide. Neurobiol. Aging 2009, 30, 1523–1534. [Google Scholar] [CrossRef]
- Kimura, Y.; Kimura, H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004, 18, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Ishii, I.; Akahoshi, N.; Yu, X.-N.; Kobayashi, Y.; Namekata, K.; Komaki, G.; Kimura, H. Murine cystathionine γ-lyase: Complete cDNA and genomic sequences, promoter activity, tissue distribution and developmental expression. Biochem. J. 2004, 381, 113–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, N.Y.; Jung, J.; Tabassum, R. Therapeutic importance of hydrogen sulfide in age-associated neurodegenerative diseases. Neural Regen. Res. 2020, 15, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Nagpure, B.V.; Bian, J.-S. Brain, Learning, and Memory: Role of H2S in Neurodegenerative Diseases. Handb. Exp. Pharmacol. 2015, 230, 193–215. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Kandel, E.R. The Molecular Biology of Memory Storage: A Dialogue Between Genes and Synapses. Science 2001, 294, 1030–1038. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.H.; Bartsch, D.; Kandel, E.R. Toward a molecular definition of long-term memory storage. Proc. Natl. Acad. Sci. USA 1996, 93, 13445–13452. [Google Scholar] [CrossRef] [Green Version]
- Tully, T. Discovery of genes involved with learning and memory: An experimental synthesis of Hirschian and Benzerian perspectives. Proc. Natl. Acad. Sci. USA 1996, 93, 13460–13467. [Google Scholar] [CrossRef] [Green Version]
- Cowan, T.M.; Siegel, R.W. Drosophila Mutations that Alter Ionic Conduction Disrupt Acquisition and Retention of a Conditioned Odor Avoidance Response. J. Neurogenet. 1986, 3, 187–201. [Google Scholar] [CrossRef]
- Lichtinghagen, R.; Stocker, M.; Wittka, R.; Boheim, G.; Stühmer, W.; Ferrus, A.; Pongs, O. Molecular basis of altered excitability in Shaker mutants of Drosophila melanogaster. EMBO J. 1990, 9, 4399–4407. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, F.; Zou, B.; Xu, X.; Ordway, R.W. Active Zone Localization of Presynaptic Calcium Channels Encoded by the cacophony Locus of Drosophila. J. Neurosci. 2004, 24, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.A.; Peixoto, A.A.; Kramer, E.M.; Villella, A.; Hall, J.C. Courtship and Visual Defects of cacophony Mutants Reveal Functional Complexity of a Calcium-Channel α1 Subunit in Drosophila. Genetics 1998, 149, 1407–1426. [Google Scholar] [CrossRef] [PubMed]
- Mayford, M.; Siegelbaum, S.A.; Kandel, E.R. Synapses and Memory Storage. Cold Spring Harb. Perspect. Biol. 2012, 4, a005751. [Google Scholar] [CrossRef] [PubMed]
- Flexner, J.B.; Flexner, L.B.; Stellar, E. Memory in Mice as Affected by Intracerebral Puromycin. Science 1963, 141, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Bliss, T.V.P.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.A. Long-Term Potentiation and Memory. Physiol. Rev. 2004, 84, 87–136. [Google Scholar] [CrossRef]
- Elgersma, Y.; Fedorov, N.B.; Ikonen, S.; Choi, E.S.; Elgersma, M.; Carvalho, O.; Giese, K.P.; Silva, A.J. Inhibitory Autophosphorylation of CaMKII Controls PSD Association, Plasticity, and Learning. Neuron 2002, 36, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.J.; Paylor, R.; Wehner, J.M.; Tonegawa, S. Impaired Spatial Learning in α-Calcium-Calmodulin Kinase II Mutant Mice. Science 1992, 257, 206–211. [Google Scholar] [CrossRef]
- Lisman, J.; Yasuda, R.; Raghavachari, S. Mechanisms of CaMKII action in long-term potentiation. Nat. Rev. Neurosci. 2012, 13, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.-Y.; Nakahata, Y.; Hayano, Y.; Yasuda, R. Mechanisms of Ca2+/calmodulin-dependent kinase II activation in single dendritic spines. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.; Lee, J.; Chen, X.; Díaz-Alonso, J.; Zhou, J.; Pleasure, S.; Nicoll, R.A. Synaptic memory requires CaMKII. eLife 2021, 10, e60360. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Tsugane, M.; Oka, J.-I.; Kimura, H. Hydrogen sulfide induces calcium waves in astrocytes. FASEB J. 2004, 18, 557–559. [Google Scholar] [CrossRef] [PubMed]
- Kamat, P.K.; Kalani, A.; Tyagi, N. Role of Hydrogen Sulfide in Brain Synaptic Remodeling. Methods Enzymol. 2015, 555, 207–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Moya, S.M.; Bauer, K.E.; Kiebler, M.A. Meet the players: Local translation at the synapse. Front. Mol. Neurosci. 2014, 7, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melchor, J.P.; Strickland, S. Tissue plasminogen activator in central nervous system physiology and pathology. Thromb. Haemost. 2005, 93, 655–660. [Google Scholar] [CrossRef]
- Sontag, E.; Nunbhakdi-Craig, V.; Sontag, J.-M.; Diaz-Arrastia, R.; Ogris, E.; Dayal, S.; Lentz, S.; Arning, E.; Bottiglieri, T. Protein Phosphatase 2A Methyltransferase Links Homocysteine Metabolism with Tau and Amyloid Precursor Protein Regulation. J. Neurosci. 2007, 27, 2751–2759. [Google Scholar] [CrossRef] [Green Version]
- Siegel, G.; Obernosterer, G.; Fiore, R.; Oehmen, M.; Bicker, S.; Christensen, M.; Khudayberdiev, S.; Leuschner, P.F.; Busch, C.J.L.; Kane, C.; et al. A functional screen implicates microRNA-138-dependent regulation of the depalmitoylation enzyme APT1 in dendritic spine morphogenesis. Nat. Cell Biol. 2009, 11, 705–716. [Google Scholar] [CrossRef]
- Phillis, J.W. Acetylcholine Release from the Central Nervous System: A 50-Year Retrospective. Crit. Rev. Neurobiol. 2005, 17, 161–217. [Google Scholar] [CrossRef]
- Blankman, J.L.; Cravatt, B.F. Chemical Probes of Endocannabinoid Metabolism. Pharmacol. Rev. 2013, 65, 849–871. [Google Scholar] [CrossRef] [Green Version]
- Viader, A.; Ogasawara, D.; Joslyn, C.M.; Sanchez-Alavez, M.; Mori, S.; Nguyen, W.; Conti, B.; Cravatt, B.F. A chemical proteomic atlas of brain serine hydrolases identifies cell type-specific pathways regulating neuroinflammation. eLife 2016, 5, e12345. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
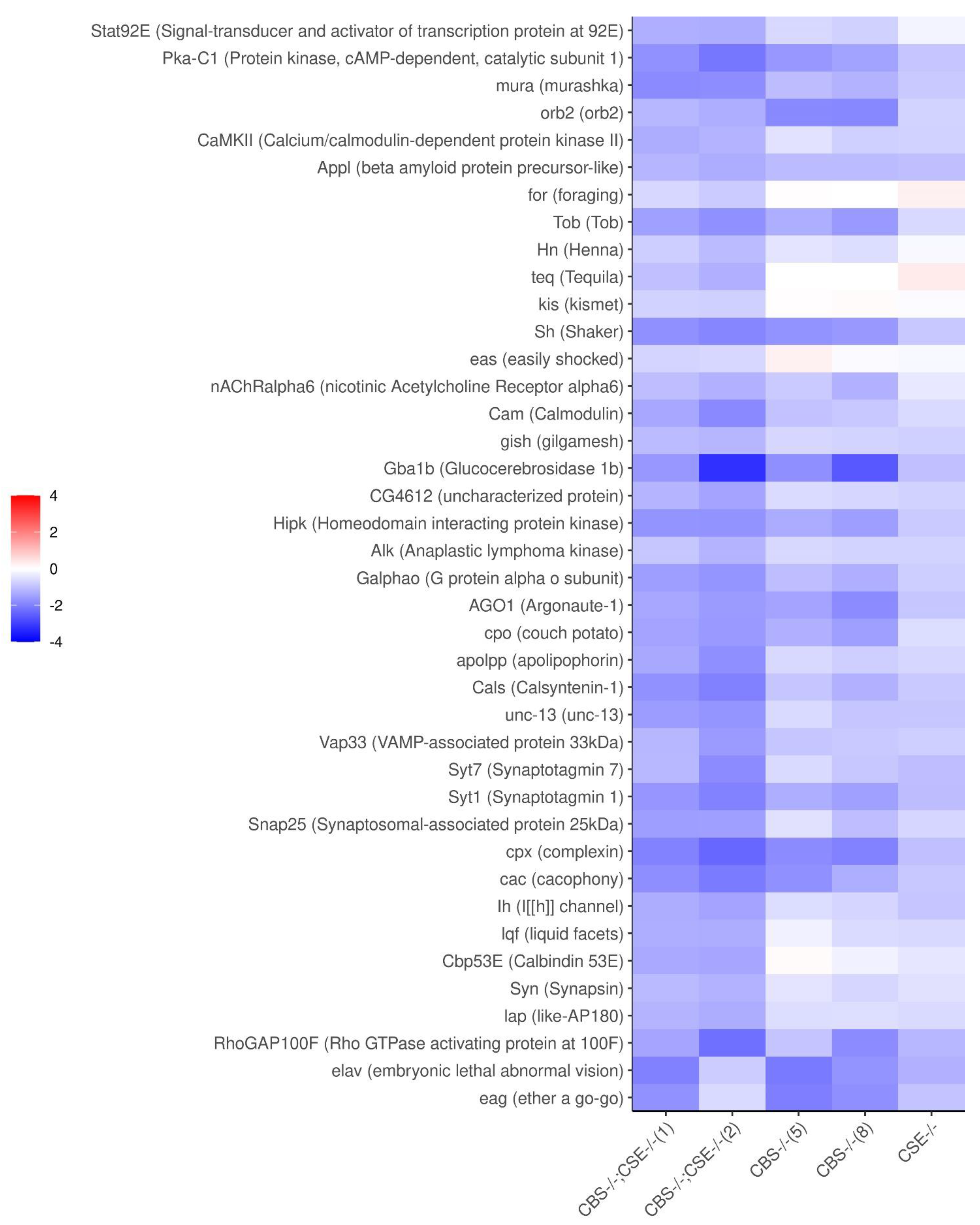{kind=link}
| Category | Number of Genes | Category | Number of Genes |
|---|---|---|---|
| CBS-/- genes down-regulated | 295 | CBS-/- genes up-regulated | 72 |
| CBS-/-; CSE-/- genes down-regulated | 190 | CBS-/-; CSE-/- genes up-regulated | 61 |
| CSE-/- genes down-regulated | 53 | CSE-/- genes up- regulated | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zatsepina, O.G.; Chuvakova, L.N.; Nikitina, E.A.; Rezvykh, A.P.; Zakluta, A.S.; Sarantseva, S.V.; Surina, N.V.; Ksenofontov, A.L.; Baratova, L.A.; Shilova, V.Y.; et al. Genes Responsible for H2S Production and Metabolism Are Involved in Learning and Memory in Drosophila melanogaster. Biomolecules 2022, 12, 751. https://doi.org/10.3390/biom12060751
Zatsepina OG, Chuvakova LN, Nikitina EA, Rezvykh AP, Zakluta AS, Sarantseva SV, Surina NV, Ksenofontov AL, Baratova LA, Shilova VY, et al. Genes Responsible for H2S Production and Metabolism Are Involved in Learning and Memory in Drosophila melanogaster. Biomolecules. 2022; 12(6):751. https://doi.org/10.3390/biom12060751
Chicago/Turabian StyleZatsepina, Olga G., Lyubov N. Chuvakova, Ekaterina A. Nikitina, Alexander P. Rezvykh, Alexey S. Zakluta, Svetlana V. Sarantseva, Nina V. Surina, Alexander L. Ksenofontov, Ludmila A. Baratova, Viktoria Y. Shilova, and et al. 2022. "Genes Responsible for H2S Production and Metabolism Are Involved in Learning and Memory in Drosophila melanogaster" Biomolecules 12, no. 6: 751. https://doi.org/10.3390/biom12060751