
Matrix Metalloproteinases on Severe COVID-19 Lung Disease Pathogenesis: Cooperative Actions of MMP-8/MMP-2 Axis on Immune Response through HLA-G Shedding and Oxidative Stress

 ,
,  , , ,
, , ,  ,
,  , , , , , , , , ,
, , , , , , , , ,  and add
Show full author list
and add
Show full author list
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Design and Participants
2.3. Data Collection
2.4. Tracheal Aspirate Fluid (TAF) Collection and Processing
2.5. Soluble TREM-1 and MMPs Quantification
2.6. Quantification of Active MMPs by Zymography
2.7. Soluble HLA-G Quantification
2.8. Assessment of Lipid Peroxide Levels (MDA)
2.9. Detection of SARS-CoV-2 RNA
2.10. Statistical Analysis
2.11. Re-Analysis of Proteomics Data
3. Results
3.1. Participants Demographic Data and Clinical Characteristics
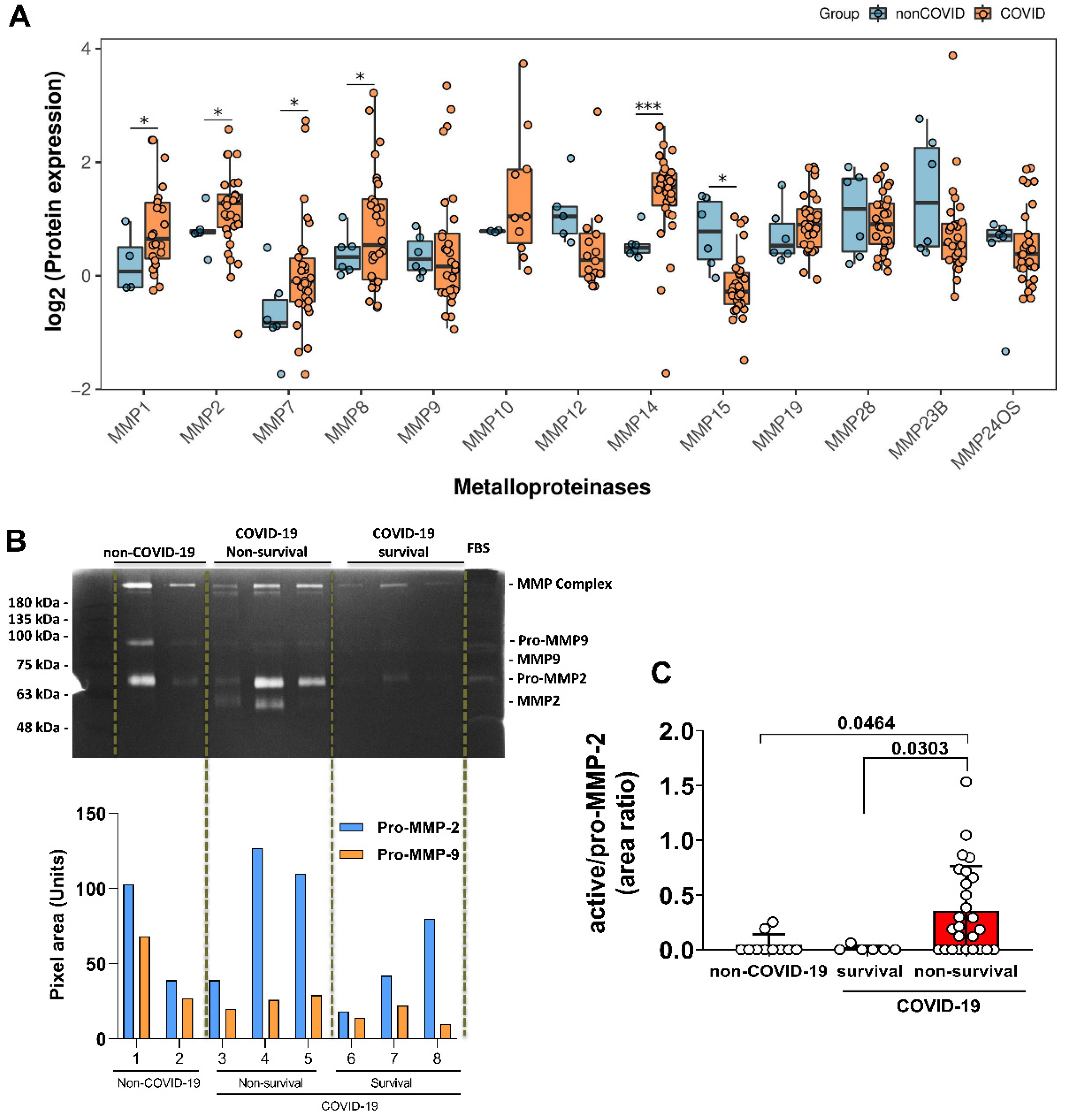
3.2. Elevated Expression of MMPs on Lung-Tissue and MMP-2 Active form on TAF Samples from COVID-19 Patients
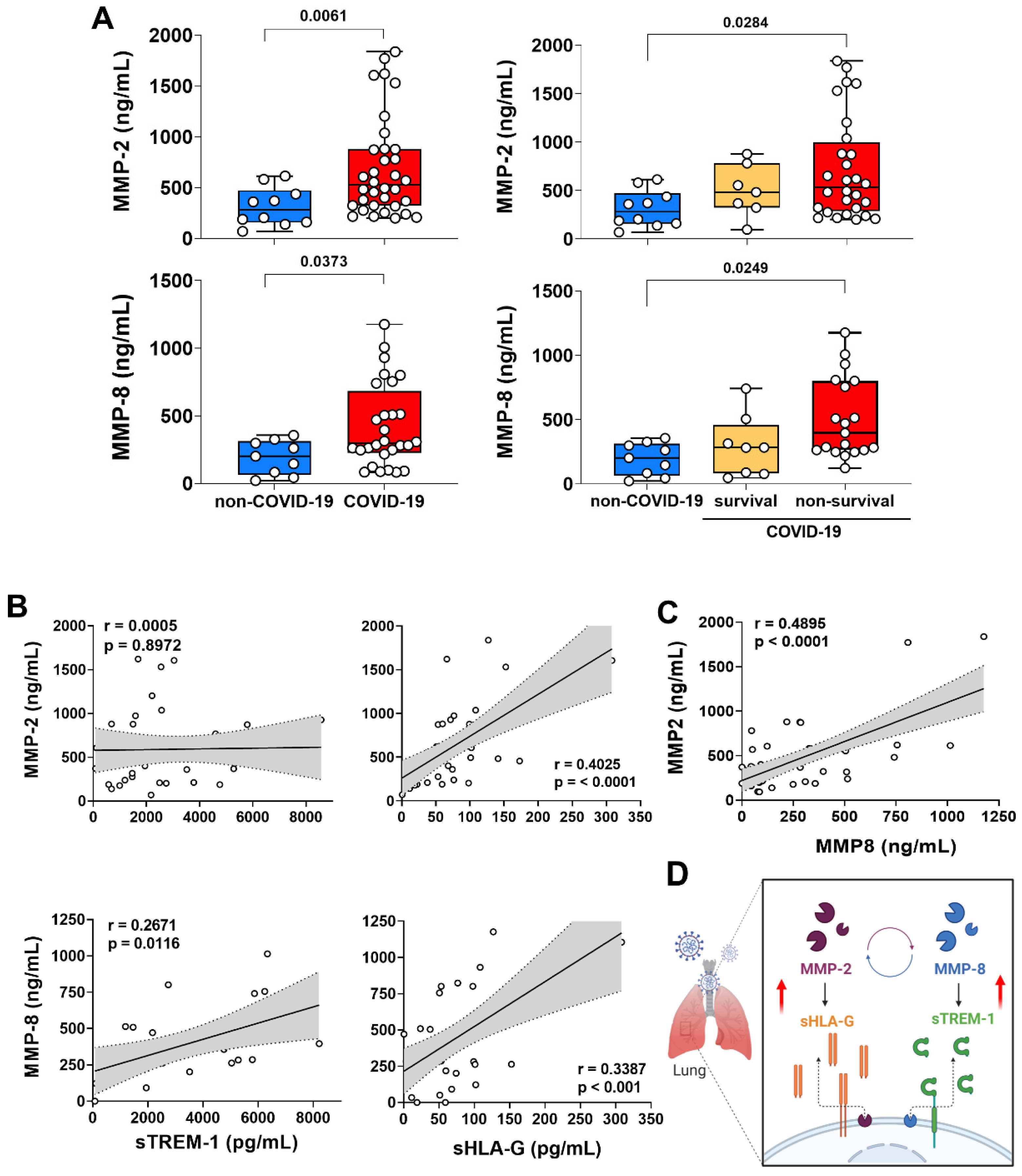
3.3. MMP-2 and MMP-8 Expression Increased in Lung from Patients with Non-Survival COVID-19 and Was Correlated with the Release of sTREM-1 and sHLA-G
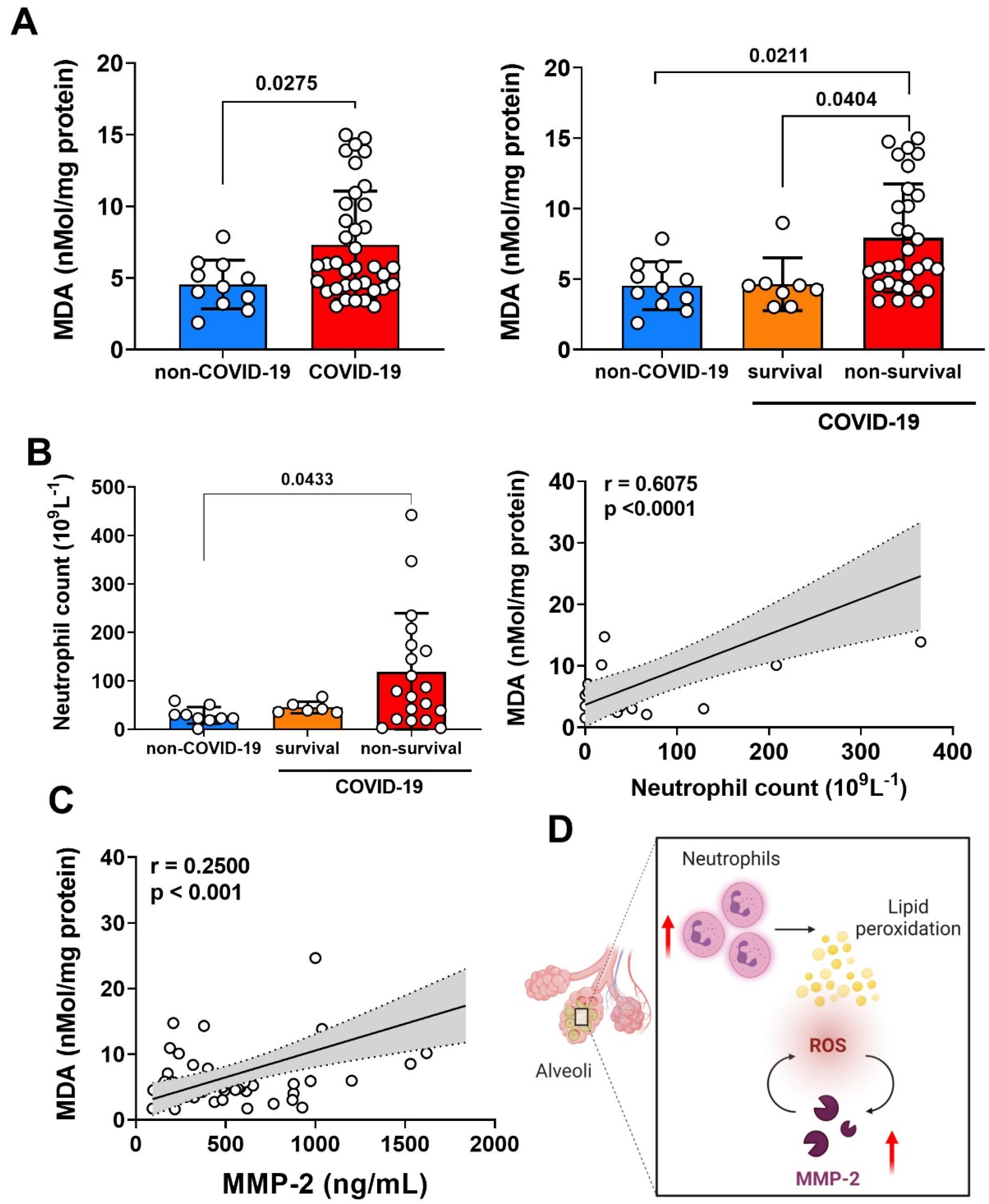
3.4. Relationship of MMP-2 Levels and Oxidative Stress in the Lung of Non-Survival COVID-19 Patients
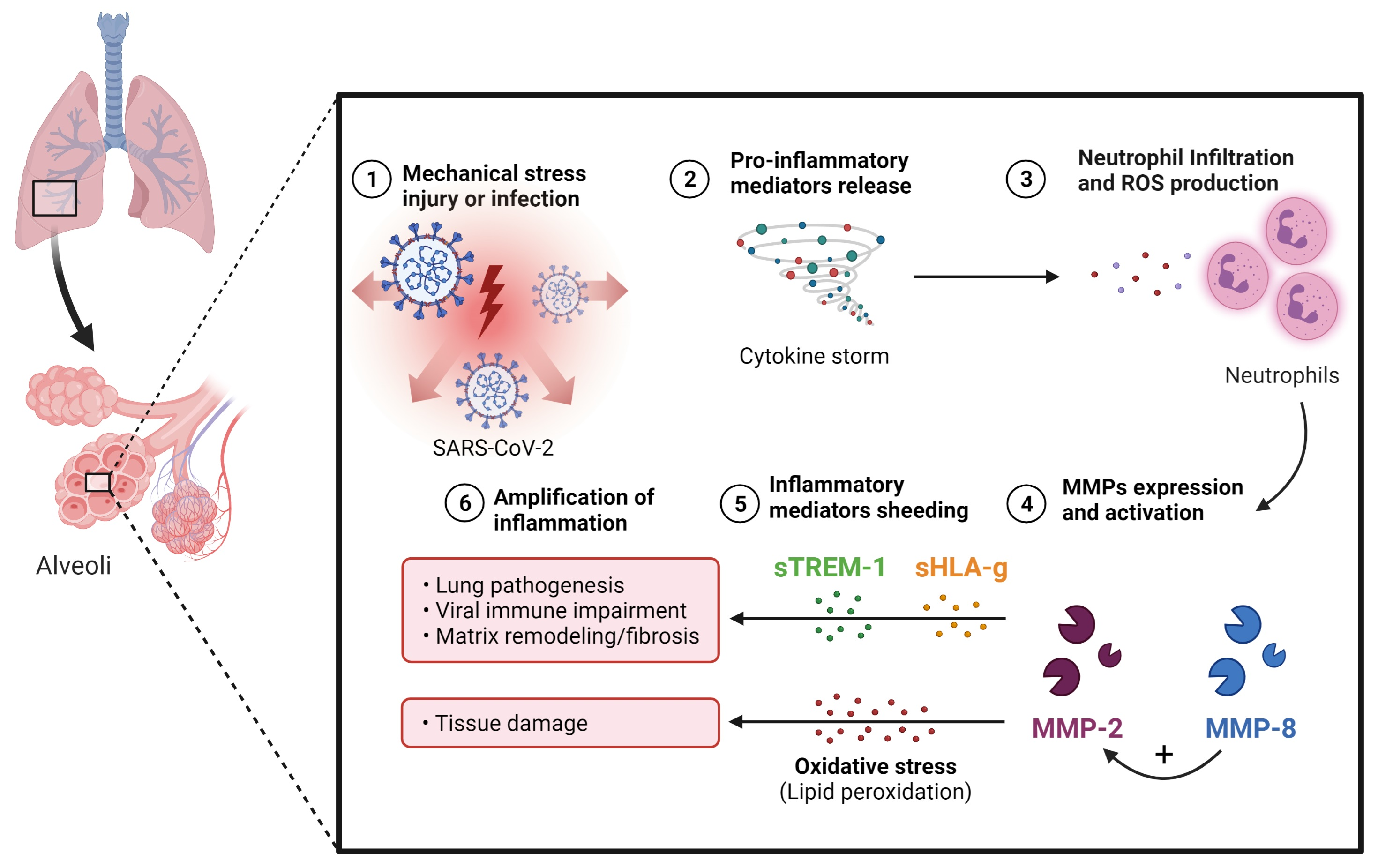
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Victora, C.G.; Castro, M.C.; Gurzenda, S.; Medeiros, A.C.; França, G.V.A.; Barros, A.J.D. Estimating the Early Impact of Vaccination against COVID-19 on Deaths among Elderly People in Brazil: Analyses of Routinely-Collected Data on Vaccine Coverage and Mortality. eClinicalMedicine 2021, 38, 101036. [Google Scholar] [CrossRef] [PubMed]
- Berlin, D.A.; Gulick, R.M.; Martinez, F.J. Severe COVID-19. N. Engl. J. Med. 2020, 383, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- CDC COVID-19 Response Team. Severe Outcomes Among Patients with Coronavirus Disease 2019 (COVID-19)—United States, 12 February–16 March 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Bhatraju, P.K.; Ghassemieh, B.J.; Nichols, M.; Kim, R.; Jerome, K.R.; Nalla, A.K.; Greninger, A.L.; Pipavath, S.; Wurfel, M.M.; Evans, L.; et al. COVID-19 in Critically Ill Patients in the Seattle Region—Case Series. N. Engl. J. Med. 2020, 382, 2012–2022. [Google Scholar] [CrossRef]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Xie, J.; Wu, W.; Li, S.; Hu, Y.; Hu, M.; Li, J.; Yang, Y.; Huang, T.; Zheng, K.; Wang, Y.; et al. Clinical Characteristics and Outcomes of Critically Ill Patients with Novel Coronavirus Infectious Disease (COVID-19) in China: A Retrospective Multicenter Study. Intensive Care Med. 2020, 46, 1863–1872. [Google Scholar] [CrossRef]
- Malemud, C.J. Matrix Metalloproteinases (MMPs) in Health and Disease: An Overview. Front. Biosci. 2006, 11, 1696–1701. [Google Scholar] [CrossRef]
- Fernandez-Patron, C.; Martinez-Cuesta, M.A.; Salas, E.; Sawicki, G.; Wozniak, M.; Radomski, M.W.; Davidge, S.T. Differential Regulation of Platelet Aggregation by Matrix Metalloproteinases-9 and -2. Thromb. Haemost. 1999, 82, 1730–1735. [Google Scholar] [CrossRef]
- Greenlee, K.J.; Werb, Z.; Kheradmand, F. Matrix Metalloproteinases in Lung: Multiple, Multifarious, and Multifaceted. Physiol. Rev. 2007, 87, 69–98. [Google Scholar] [CrossRef]
- D’Avila-Mesquita, C.; Couto, A.E.S.; Campos, L.C.B.; Vasconcelos, T.F.; Michelon-Barbosa, J.; Corsi, C.A.C.; Mestriner, F.; Petroski-Moraes, B.C.; Garbellini-Diab, M.J.; Couto, D.M.S.; et al. MMP-2 and MMP-9 Levels in Plasma Are Altered and Associated with Mortality in COVID-19 Patients. Biomed. Pharmacother. 2021, 142, 112067. [Google Scholar] [CrossRef]
- Nagaset, H.; Woessner, J.F. Matrix Metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.L.; Dyer, R.; Hupe, D.J. Matrix Metalloproteinases. Curr. Opin. Chem. Biol. 1998, 2, 466–471. [Google Scholar] [CrossRef]
- Fernandez-Patron, C.; Kassiri, Z.; Leung, D. Modulation of Systemic Metabolism by MMP-2: From MMP-2 Deficiency in Mice to MMP-2 Deficiency in Patients. Compr. Physiol. 2016, 6, 1935–1949. [Google Scholar] [CrossRef] [PubMed]
- Baricordi, O.; Stignani, M.; Melchiorri, L.; Rizzo, R. HLA-G and Inflammatory Diseases. Inflamm. Allergy Drug Targets 2008, 7, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Piña, V.; Soares-Schanoski, A.; Rodríguez-Rojas, A.; del Fresno, C.; García, F.; Vallejo-Cremades, M.T.; Fernández-Ruiz, I.; Arnalich, F.; Fuentes-Prior, P.; López-Collazo, E. Metalloproteinases Shed TREM-1 Ectodomain from Lipopolysaccharide-Stimulated Human Monocytes. J. Immunol. 2007, 179, 4065–4073. [Google Scholar] [CrossRef] [Green Version]
- Demaria, S.; Schwab, R.; Gottesman, S.R.S.; Bushkin, Y. Soluble Β2-Microglobulin-Free Class I Heavy Chains Are Released from the Surface of Activated and Leukemia Cells by a Metalloprotease. J. Biol. Chem. 1994, 269, 6689–6694. [Google Scholar] [CrossRef]
- Zidi, I.; Guillard, C.; Marcou, C.; Krawice-Radanne, I.; Sangrouber, D.; Rouas-Freiss, N.; Carosella, E.D.; Moreau, P. Increase in HLA-G1 Proteolytic Shedding by Tumor Cells: A Regulatory Pathway Controlled by NF-ΚB Inducers. Cell. Mol. Life Sci. 2006, 63, 2669–2681. [Google Scholar] [CrossRef]
- Dong, Y.; Lieskovska, J.; Kedrin, D.; Porcelli, S.; Mandelboim, O.; Bushkin, Y. Soluble Nonclassical HLA Generated by the Metalloproteinase Pathway. Hum. Immunol. 2003, 64, 802–810. [Google Scholar] [CrossRef]
- Bonam, S.R.; Kotla, N.G.; Bohara, R.A.; Rochev, Y.; Webster, T.J.; Bayry, J. Potential Immuno-Nanomedicine Strategies to Fight COVID-19 like Pulmonary Infections. Nano Today 2021, 36, 101051. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and Clinical Characteristics of 99 Cases of 2019 Novel Coronavirus Pneumonia in Wuhan, China: A Descriptive Study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Narasaraju, T.; Tang, B.M.; Herrmann, M.; Muller, S.; Chow, V.T.K.; Radic, M. Neutrophilia and NETopathy as Key Pathologic Drivers of Progressive Lung Impairment in Patients With COVID-19. Front. Pharmacol. 2020, 11, 870. [Google Scholar] [CrossRef] [PubMed]
- Menter, T.; Haslbauer, J.D.; Nienhold, R.; Savic, S.; Hopfer, H.; Deigendesch, N.; Frank, S.; Turek, D.; Willi, N.; Pargger, H.; et al. Postmortem Examination of COVID-19 Patients Reveals Diffuse Alveolar Damage with Severe Capillary Congestion and Variegated Findings in Lungs and Other Organs Suggesting Vascular Dysfunction. Histopathology 2020, 77, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, D.; Sperhake, J.P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients with COVID-19: A Prospective Cohort Study. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial Cell Infection and Endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary Fibrosis and COVID-19: The Potential Role for Antifibrotic Therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Legrand, C.; Gilles, C.; Zahm, J.M.; Polette, M.; Buisson, A.C.; Kaplan, H.; Birembaut, P.; Tournier, J.M. Airway Epithelial Cell Migration Dynamics: MMP-9 Role in Cell- Extracellular Matrix Remodeling. J. Cell Biol. 1999, 146, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Tetley, T.D. Proteinase Imbalance: Its Role in Lung Disease. Thorax 1993, 48, 560–565. [Google Scholar] [CrossRef] [Green Version]
- Shields, M.D.; Riedler, J. Bronchoalveolar Lavage and Tracheal Aspirate for Assessing Airway Inflammation in Children. Am. J. Respir. Crit. Care Med. 2000, 162, S15–S17. [Google Scholar] [CrossRef]
- Gerlach, R.F.; Uzuelli, J.A.; Souza-Tarla, C.D.; Tanus-Santos, J.E. Effect of Anticoagulants on the Determination of Plasma Matrix Metalloproteinase (MMP)-2 and MMP-9 Activities. Anal. Biochem. 2005, 344, 147–149. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Martelli-Palomino, G.; Pancotto, J.A.; Muniz, Y.C.; Mendes-Junior, C.T.; Castelli, E.C.; Massaro, J.D.; Krawice-Radanne, I.; Poras, I.; Rebmann, V.; Carosella, E.D.; et al. Polymorphic Sites at the 3′ Untranslated Region of the HLA-G Gene Are Associated with Differential Hla-g Soluble Levels in the Brazilian and French Population. PLoS ONE 2013, 8, e71742. [Google Scholar] [CrossRef] [PubMed]
- Tsikas, D. Assessment of Lipid Peroxidation by Measuring Malondialdehyde (MDA) and Relatives in Biological Samples: Analytical and Biological Challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, M.F.; Pessa, L.R.; Gomes, V.A.; Desta, Z.; Flockhart, D.A.; Tanus-Santos, J.E. Assessment of Vascular Effects of Tamoxifen and Its Metabolites on the Rat Perfused Hindquarter Vascular Bed. Basic Clin. Pharmacol. Toxicol. 2009, 104, 400–407. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 Novel Coronavirus (2019-NCoV) by Real-Time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC 2019-Novel Coronavirus (2019-NCoV) Real-Time RT-PCR Diagnostic Panel For Emergency Use Only Instructions for Use; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2021.
- Pontelli, M.C.; Castro, I.A.; Martins, R.B.; Veras, F.P.; la Serra, L.; Nascimento, D.C.; Cardoso, R.S.; Rosales, R.; Lima, T.M.; Souza, J.P.; et al. Infection of Human Lymphomononuclear Cells by SARS-CoV-2. bioRxiv 2020, 1, 1–40. [Google Scholar] [CrossRef]
- Nie, X.; Qian, L.; Sun, R.; Huang, B.; Dong, X.; Xiao, Q.; Zhang, Q.; Lu, T.; Yue, L.; Chen, S.; et al. Multi-Organ Proteomic Landscape of COVID-19 Autopsies. Cell 2021, 184, 775–791.e14. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing: Reference Index; R Foundation for Statistical Computing: Vienna, Austria, 2010; ISBN 3900051070. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- da Silva-Neto, P.v.; de Carvalho, J.C.S.; Pimentel, V.E.; Pérez, M.M.; Toro, D.M.; Fraga-Silva, T.F.C.; Fuzo, C.A.; Oliveira, C.N.S.; Rodrigues, L.C.; Argolo, J.G.M.; et al. sTREM-1 Predicts Disease Severity and Mortality in COVID-19 Patients: Involvement of Peripheral Blood Leukocytes and MMP-8 Activity. Viruses 2021, 13, 2521. [Google Scholar] [CrossRef]
- Rizzo, R.; Trentini, A.; Bortolotti, D.; Manfrinato, M.C.; Rotola, A.; Castellazzi, M.; Melchiorri, L.; di Luca, D.; Dallocchio, F.; Fainardi, E.; et al. Matrix Metalloproteinase-2 (MMP-2) Generates Soluble HLA-G1 by Cell Surface Proteolytic Shedding. Mol. Cell. Biochem. 2013, 381, 243–255. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, Y.; Xiang, P.; Pu, L.; Xiong, H.; Li, C.; Zhang, M.; Tan, J.; Xu, Y.; Song, R.; et al. Neutrophil-to-Lymphocyte Ratio Predicts Critical Illness Patients with 2019 Coronavirus Disease in the Early Stage. J. Transl. Med. 2020, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Ragab, D.; Eldin, H.S.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Singh, K.; Mittal, S.; Gollapudi, S.; Butzmann, A.; Kumar, J.; Ohgami, R.S. A Meta-Analysis of SARS-CoV-2 Patients Identifies the Combinatorial Significance of D-Dimer, C-Reactive Protein, Lymphocyte, and Neutrophil Values as a Predictor of Disease Severity. Int. J. Lab. Hematol. 2021, 43, 324–328. [Google Scholar] [CrossRef]
- Matthay, M.A.; Wick, K.D. Corticosteroids, COVID-19 Pneumonia, and Acute Respiratory Distress Syndrome. J. Clin. Investig. 2020, 130, 6218–6221. [Google Scholar] [CrossRef]
- Parks, W.C.; Shapiro, S.D. Matrix Metalloproteinases in Lung Biology. Respir. Res. 2001, 2, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheradmand, F.; Rishi, K. The Role of Proteases in Airway Remodeling. In Therapeutic Targets in Airway Inflammation; CRC Press: Boca Raton, FL, USA, 2003; pp. 18–19. [Google Scholar]
- McKeown, S.; Richter, A.G.; O’Kane, C.; McAuley, D.F.; Thickett, D.R. MMP Expression and Abnormal Lung Permeability Are Important Determinants of Outcome in IPF. Eur. Respir. J. 2009, 33, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Elkington, P.T.G.; Friedland, J.S. Matrix Metalloproteinases in Destructive Pulmonary Pathology. Thorax 2006, 61, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugin, J.; Verghese, G.; Widmer, M.C.; Matthay, M.A. The Alveolar Space Is the Site of Intense Inflammatory and Profibrotic Reactions in the Early Phase of Acute Respiratory Distress Syndrome. Crit. Care Med. 1999, 27, 304–312. [Google Scholar] [CrossRef]
- Atkinson, J.J.; Senior, R.M. Matrix Metalloproteinase-9 in Lung Remodeling. Am. J. Respir. Cell Mol. Biol. 2003, 28, 12–24. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.O. Dysregulation of Pulmonary Responses in Severe COVID-19. Viruses 2021, 13, 957. [Google Scholar] [CrossRef]
- van Wart, H.E.; Birkedal-Hansen, H. The Cysteine Switch: A Principle of Regulation of Metalloproteinase Activity with Potential Applicability to the Entire Matrix Metalloproteinase Gene Family. Proc. Natl. Acad. Sci. USA 1990, 87, 5578–5582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix Metalloproteinases and the Regulation of Tissue Remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Yao, P.M.; Maitre, B.; Delacourt, G.; Buhler, J.M.; Harf, A.; Lafuma, C. Divergent Regulation of 92-KDa Gelatinase and TIMP-1 by HBECs in Response to IL-1β and TNF-α. Am. J. Physiol. Lung Cell. Mol. Physiol. 1997, 273, L866–L874. [Google Scholar] [CrossRef] [PubMed]
- Schnaper, H.W.; Grant, D.S.; Stetler-Stevenson, W.G.; Fridman, R.; D’Orazi, G.; Murphy, A.N.; Bird, R.E.; Hoythya, M.; Fuerst, T.R.; French, D.L.; et al. Type IV Collagenase(s) and TIMPs Modulate Endothelial Cell Morphogenesis in Vitro. J. Cell. Physiol. 1993, 156, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Corry, D.B.; Rishi, K.; Kanellis, J.; Kiss, A.; Song, L.-Z.; Xu, J.; Werb, Z.; Kheradmand, F. Decreased Allergic Lung Inflammatory Cell Egression and Increased Susceptibility to Asphyxiation in MMP2-Deficiency. Nat. Immunol. 2002, 3, 347–353. [Google Scholar] [CrossRef]
- Campbell, L.G.; Ramachandran, S.; Liu, W.; Shipley, M.M.; Itohara, S.; Rogers, J.G.; Moazami, N.; Senior, R.M.; Jaramillo, A. Different Roles for Matrix Metalloproteinase-2 and Matrix Metalloproteinase-9 in the Pathogenesis of Cardiac Allograft Rejection. Am. J. Transplant. 2005, 5, 517–528. [Google Scholar] [CrossRef]
- Owen, C.A.; Hu, Z.; Lopez-Otin, C.; Shapiro, S.D. Membrane-Bound Matrix Metalloproteinase-8 on Activated Polymorphonuclear Cells Is a Potent, Tissue Inhibitor of Metalloproteinase-Resistant Collagenase and Serpinase. J. Immunol. 2004, 172, 7791–7803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix Metalloproteinases as Modulators of Inflammation and Innate Immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Faurschou, M.; Borregaard, N. Neutrophil Granules and Secretory Vesicles in Inflammation. Microbes Infect. 2003, 5, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Cederqvist, K.; Sorsa, T.; Tervahartiala, T.; Maisi, P.; Reunanen, K.; Lassus, P.; Andersson, S. Matrix Metalloproteinases-2, -8, and -9 and TIMP-2 in Tracheal Aspirates from Preterm Infants with Respiratory Distress. Pediatrics 2001, 108, 686–692. [Google Scholar] [CrossRef]
- Kadry, R.; Newsome, A.S.; Somanath, P.R. Pharmacological Inhibition of MMP3 as a Potential Therapeutic Option for COVID-19 Associated Acute Respiratory Distress Syndrome. Infect. Disord. Drug Targets 2020, 21, e170721187996. [Google Scholar] [CrossRef] [PubMed]
- Gelzo, M.; Cacciapuoti, S.; Pinchera, B.; de Rosa, A.; Cernera, G.; Scialò, F.; Comegna, M.; Mormile, M.; Fabbrocini, G.; Parrella, R.; et al. Matrix Metalloproteinases (MMP) 3 and 9 as Biomarkers of Severity in COVID-19 Patients. Sci. Rep. 2022, 12, 1212. [Google Scholar] [CrossRef] [PubMed]
- van Lint, P.; Libert, C. Chemokine and Cytokine Processing by Matrix Metalloproteinases and Its Effect on Leukocyte Migration and Inflammation. J. Leukoc. Biol. 2007, 82, 1375–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, D.; Morrison, C.J.; Overall, C.M. Matrix Metalloproteinases: What Do They Not Do? New Substrates and Biological Roles Identified by Murine Models and Proteomics. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morandi, F.; Rizzo, R.; Fainardi, E.; Rouas-Freiss, N.; Pistoia, V. Recent Advances in Our Understanding of HLA-G Biology: Lessons from a Wide Spectrum of Human Diseases. J. Immunol. Res. 2016, 2016, 4326495. [Google Scholar] [CrossRef] [Green Version]
- Al-Bayatee, N.T.; Ad’hiah, A.H. Soluble HLA-G Is Upregulated in Serum of Patients with Severe COVID-19. Hum. Immunol. 2021, 82, 726–732. [Google Scholar] [CrossRef]
- Zheng, H.; Heiderscheidt, C.A.; Joo, M.; Gao, X.; Knezevic, N.; Mehta, D.; Sadikot, R.T. MYD88-Dependent and -Independent Activation of TREM-1 via Specific TLR Ligands. Eur. J. Immunol. 2010, 40, 162–171. [Google Scholar] [CrossRef]
- Weiss, G.; Lai, C.; Fife, M.E.; Grabiec, A.M.; Tildy, B.; Snelgrove, R.J.; Xin, G.; Lloyd, C.M.; Hussell, T. Reversal of TREM-1 Ectodomain Shedding and Improved Bacterial Clearance by Intranasal Metalloproteinase Inhibitors. Mucosal Immunol. 2017, 10, 1021–1030. [Google Scholar] [CrossRef]
- Strongin, A.Y.; Collier, I.; Bannikov, G.; Marmer, B.L.; Grant, G.A.; Goldberg, G.I. Mechanism of Cell Surface Activation of 72-KDa Type IV Collagenase. J. Biol. Chem. 1995, 270, 5331–5338. [Google Scholar] [CrossRef] [Green Version]
- Dozier, S.; Escobar, G.; Lindsey, M. Matrix Metalloproteinase (MMP)-7 Activates MMP-8 But Not MMP-13. Med. Chem. 2006, 2, 523–526. [Google Scholar] [CrossRef]
- Solun, B.; Shoenfeld, Y. Inhibition of Metalloproteinases in Therapy for Severe Lung Injury Due to COVID-19. Med. Drug Discov. 2020, 7, 100052. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Rios, F.J.; Alves-Lopes, R.; Neves, K.B.; Camargo, L.L.; Montezano, A.C. Oxidative Stress: A Unifying Paradigm in Hypertension. Can. J. Cardiol. 2020, 36, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopalan, S.; Meng, X.P.; Ramasamy, S.; Harrison, D.G.; Galis, Z.S. Reactive Oxygen Species Produced by Macrophage-Derived Foam Cells Regulate the Activity of Vascular Matrix Metalloproteinases in Vitro: Implications for Atherosclerotic Plaque Stability. J. Clin. Investig. 1996, 98, 2572–2579. [Google Scholar] [CrossRef] [Green Version]
- Migita, K.; Maeda, Y.; Abiru, S.; Komori, A.; Yokoyama, T.; Takii, Y.; Nakamura, M.; Yatsuhashi, H.; Eguchi, K.; Ishibashi, H. Peroxynitrite-Mediated Matrix Metalloproteinase-2 Activation in Human Hepatic Stellate Cells. FEBS Lett. 2005, 579, 3119–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viappiani, S.; Nicolescu, A.C.; Holt, A.; Sawicki, G.; Crawford, B.D.; León, H.; van Mulligen, T.; Schulz, R. Activation and Modulation of 72 KDa Matrix Metalloproteinase-2 by Peroxynitrite and Glutathione. Biochem. Pharmacol. 2009, 77, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Prado, A.F.; Pernomian, L.; Azevedo, A.; Costa, R.A.P.; Rizzi, E.; Ramos, J.; Paes Leme, A.F.; Bendhack, L.M.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix Metalloproteinase-2-Induced Epidermal Growth Factor Receptor Transactivation Impairs Redox Balance in Vascular Smooth Muscle Cells and Facilitates Vascular Contraction. Redox Biol. 2018, 18, 181–190. [Google Scholar] [CrossRef]
- Galis, Z.S.; Khatri, J.J. Matrix Metalloproteinases in Vascular Remodeling and Atherogenesis: The Good, the Bad, and the Ugly. Circ. Res. 2002, 90, 251–262. [Google Scholar] [CrossRef]
- Stephenson, E.; Reynolds, G.; Botting, R.A.; Calero-Nieto, F.J.; Morgan, M.D.; Tuong, Z.K.; Bach, K.; Sungnak, W.; Worlock, K.B.; Yoshida, M.; et al. Single-Cell Multi-Omics Analysis of the Immune Response in COVID-19. Nat. Med. 2021, 27, 904–916. [Google Scholar] [CrossRef]
- Ren, X.; Wen, W.; Fan, X.; Hou, W.; Su, B.; Cai, P.; Li, J.; Liu, Y.; Tang, F.; Zhang, F.; et al. COVID-19 Immune Features Revealed by a Large-Scale Single-Cell Transcriptome Atlas. Cell 2021, 184, 1895–1913.e19. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Kuwano, K. Epithelial Cell Apoptosis and Lung Remodeling. Cell. Mol. Immunol. 2007, 4, 419–429. [Google Scholar] [PubMed]
- Gropper, M.A.; Wiener-Kronish, J. The Epithelium in Acute Lung Injury/Acute Respiratory Distress Syndrome. Curr. Opin. Crit. Care 2008, 14, 11–15. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in Sepsis: A Novel Understanding of the Disorder and a New Therapeutic Approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Amodio, G.; de Albuquerque, R.S.; Gregori, S. New Insights into HLA-G Mediated Tolerance. Tissue Antigens 2014, 84, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fraga-Silva, T.F.d.C.; Maruyama, S.R.; Sorgi, C.A.; Russo, E.M.d.S.; Fernandes, A.P.M.; de Barros Cardoso, C.R.; Faccioli, L.H.; Dias-Baruffi, M.; Bonato, V.L.D. COVID-19: Integrating the Complexity of Systemic and Pulmonary Immunopathology to Identify Biomarkers for Different Outcomes. Front. Immunol. 2021, 11, 599736. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Variable | Non-COVID-19 N = 13 | COVID-19 N = 39 | COVID-19 Survival N = 6 | COVID-19 Non-Survival N = 33 | p Value |
|---|---|---|---|---|---|
| Demographic characteristics (median ± SD) | |||||
| Age | 61 ± 17.8 | 66 ± 16.0 | 61 ± 24.1 | 66 ± 14.3 | 0.5200 a 0.5488 b |
| BMI (kg/m2) | 26.8 ± 6.5 | 29.4 ± 7.0 | 27.4 ± 6.7 | 29.5 ± 7.1 | 0.1528 a 0.3915 b |
| Sex, No. (%) | |||||
| Male | 4 (30.8) | 21 (55.2) | 1 (16.7) | 17 (51.5) | - |
| Female | 9 (69.2) | 18 (44.7) | 5 (83.3) | 16 (48.5) | - |
| Comorbidities or coexisting disorders, No. (%) | |||||
| Hypertension | 3 (23.1) | 23(59) | 3 (50) | 20 (60.6) | 0.0250 a 0.6271 b |
| Dyslipidemia | - | 2 (5.1) | - | 2 (6.0) | - |
| Diabetes mellitus | 2 (15.4) | 15 (38.4) | 1 (16.7) | 14 (42.4) | 0.1245 a 0.2329 b |
| Obesity | 3 (23.1) | 17 (43.6) | 1 (16.7) | 16 (48.5) | 0.1880 a 0.1482 b |
| Neurological Disease | 2 (15.4) | 3 (7.7) | - | 3 (9.1) | 0.4152 a |
| Respiratory Disorders | 1 (16.7) | 5 (12.8) | - | 5 (21.7) | 0.6162 a |
| Presenting symptoms, No. (%) | |||||
| Dyspnea | 4 (30.7) | 27 (69.2) | 4 (66.7) | 23 (69.7) | 0.0144a 0.8824 b |
| Fever | - | 12 (30.8) | 2 (33.3) | 10 (30.3) | 0.8824 b |
| Myalgia | - | 17 (43.6) | 1 (16.7) | 16 (48.5) | 0.0037a 0.1482 b |
| Diarrhea | - | 7 (18) | - | 7 (21.2) | - |
| Cough | - | 13 (33.3) | 4 (66.7) | 9 (27.3) | 0.0597 b |
| Anosmia | - | 4 (10.3) | 1 (16.7) | 3 (9.1) | 0.5737 b |
| Dysgeusia | - | 3 (7.7) | - | 3 (9.1) | - |
| Laboratory findings (median ± SD) | |||||
| Erythrocytes × 109/L | 2.7 ± 0.9 | 3.7 ± 0.8 | 3.0 ± 0.7 | 3.9 ± 0.7 | 0.0037 a 0.0136 b |
| Hemoglobin (g/dL) | 8.7 ± 2.9 | 10.7 ± 2.2 | 9.4 ± 1.3 | 10.9 ± 1.9 | 0.0090 a 0.0115 b |
| Leukocytes × 109/L | 11.5 ± 15.4 | 15.1 ± 16.7 | 11.2 ± 4.2 | 16.2 ± 18.0 | 0.5318 a 0.1850 b |
| Neutrophils × 109/L | 8.7 ± 11.1 | 12.8 ± 5.5 | 7.6 ± 4.1 | 13.4 ± 5.5 | 0.8028 a 0.1327 b |
| Lymphocytes × 109/L | 1.1 ± 1.1 | 0.9 ± 0.5 | 1.4 ± 0.8 | 0.9 ± 0.5 | 0.5117 a 0.5820 b |
| NLR | 7.0 ± 21.1 | 12.3 ± 10.5 | 5.0 ± 10.0 | 12.8 ± 10.6 | 0.1879 a 0.3311 b |
| Monocytes × 109/L | 0.48 ± 0.4 | 0.52 ± 0.5 | 0.6 ± 0.4 | 0.5 ± 0.5 | 0.7854 a 0.6482 b |
| Platelets × 109/L | 250 ± 165.8 | 229 ± 9.15 | 286 ± 80.7 | 228 ± 92.3 | 0.3112 a 0.6693 b |
| Hospital support, No. (%) | |||||
| Intensive care unit (ICU) | 13 (100) | 39 (100) | 6 (100) | 33 (100) | - |
| Hospitalization data, No. (median ± SD) | |||||
| Hospitalization days | 23 ± 19.6 | 6.5 ± 2.6 | 23.5 ± 8.2 | 17 ± 8.4 | <0.0001 a 0.0374 b |
| Respiratory support received | |||||
| Invasive mechanical ventilation (%) | 13 (100) | 39 (100) | 6 (100) | 33 (100) | - |
| Oxygen Saturation (median ± SD) | 94 ± 6.3 | 91 ± 10.7 | 91.5 ± 4.2 | 91 ± 11.3 | 0.0125 a 0.3240 b |
| PaO2/FiO2 ratio (median ± SD) | 156.3 ± 58.3 | 140.3 ± 94.8 | 106.4 ± 165 | 140.3 ± 76.4 | 0.9335 a 0.9245 b |
| Denouement, No (%) | |||||
| Survival | 5 (38.5) | 6 (15.4) | - | - | - |
| Non-survival | 8 (61.5) | 33 (84.6) | - | - | - |
| Viral charge | |||||
| ∆CT | - | 57.8 ± 3402 | 30.2 ± 3899 | 52.6 ± 10,455 | 0.3706 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva-Neto, P.V.; do Valle, V.B.; Fuzo, C.A.; Fernandes, T.M.; Toro, D.M.; Fraga-Silva, T.F.C.; Basile, P.A.; de Carvalho, J.C.S.; Pimentel, V.E.; Pérez, M.M.; et al. Matrix Metalloproteinases on Severe COVID-19 Lung Disease Pathogenesis: Cooperative Actions of MMP-8/MMP-2 Axis on Immune Response through HLA-G Shedding and Oxidative Stress. Biomolecules 2022, 12, 604. https://doi.org/10.3390/biom12050604
da Silva-Neto PV, do Valle VB, Fuzo CA, Fernandes TM, Toro DM, Fraga-Silva TFC, Basile PA, de Carvalho JCS, Pimentel VE, Pérez MM, et al. Matrix Metalloproteinases on Severe COVID-19 Lung Disease Pathogenesis: Cooperative Actions of MMP-8/MMP-2 Axis on Immune Response through HLA-G Shedding and Oxidative Stress. Biomolecules. 2022; 12(5):604. https://doi.org/10.3390/biom12050604
Chicago/Turabian Styleda Silva-Neto, Pedro V., Valéria B. do Valle, Carlos A. Fuzo, Talita M. Fernandes, Diana M. Toro, Thais F. C. Fraga-Silva, Patrícia A. Basile, Jonatan C. S. de Carvalho, Vinícius E. Pimentel, Malena M. Pérez, and et al. 2022. "Matrix Metalloproteinases on Severe COVID-19 Lung Disease Pathogenesis: Cooperative Actions of MMP-8/MMP-2 Axis on Immune Response through HLA-G Shedding and Oxidative Stress" Biomolecules 12, no. 5: 604. https://doi.org/10.3390/biom12050604