
Insights into Recent Studies on Biotransformation and Pharmacological Activities of Ginsenoside Rd


Abstract
:1. Introduction
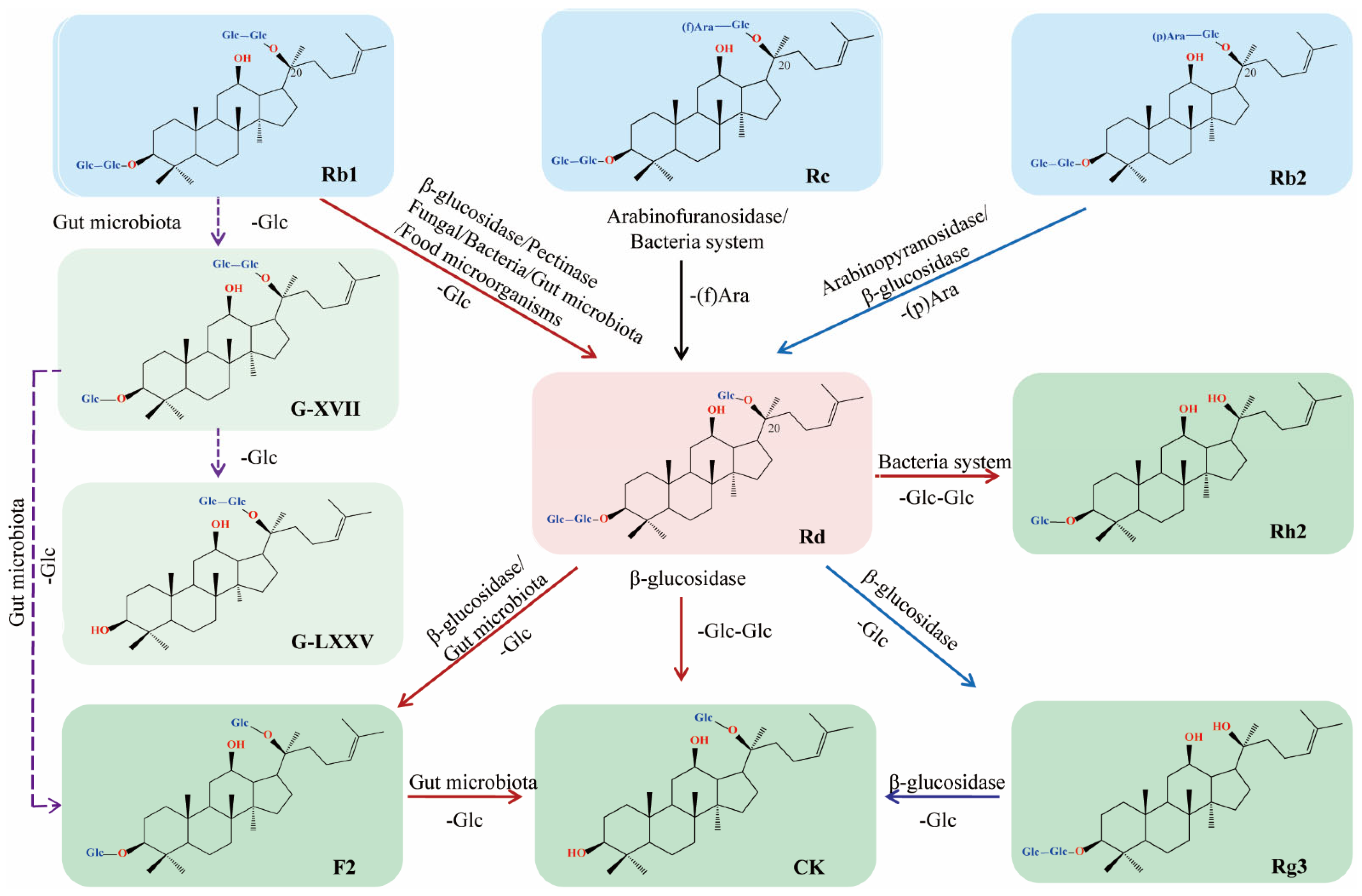
2. Biotransformation
2.1. Enzymatic Transformation
2.1.1. Arabinofuranosidase
2.1.2. β-glucosidase
2.1.3. Pectinase
2.2. Microbial Transformation
2.2.1. Fungal System
2.2.2. Bacteria System
2.2.3. Gut Microbiota and Food Microorganisms
3. Pharmacological Activity
3.1. Anti-Cancer
3.2. Anti-Diabetic
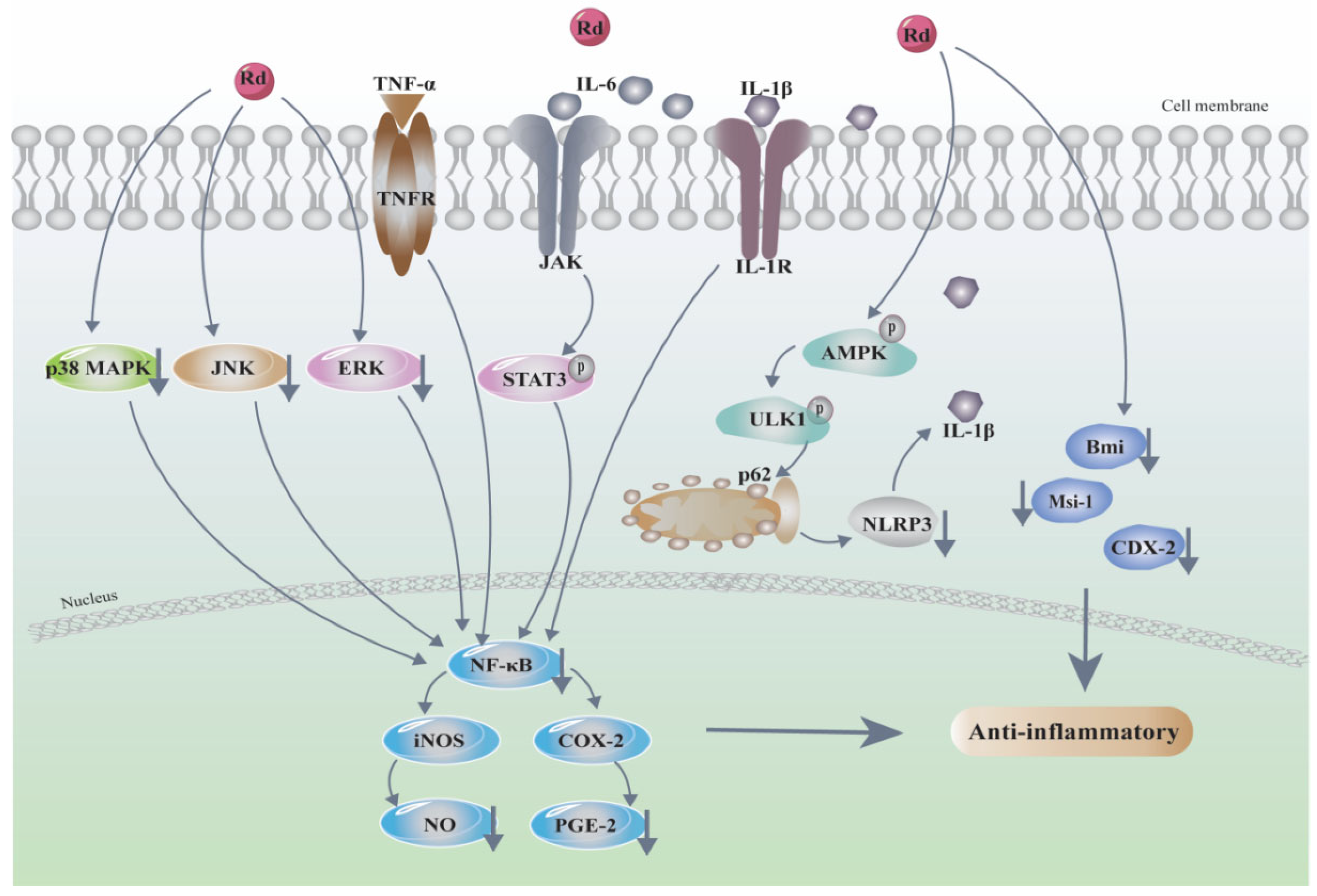
3.3. Anti-Inflammatory and Antioxidative
3.4. Cognition and Neuroprotection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
|---|---|---|---|---|---|---|
| Alzheimer’s disease (AD) | Animals: Aβ1–40 induced AD rat model | In vivo: Rd (10, 30 mg/kg/d, 30 days) | Protected cognitive impairment, improved memory function, alleviated Aβ1–40 induced inflammation | caspase-3↓, apoptosis↓ | [71] | 2012 |
| Cell lines: Aβ25–35 induced primary hippocampal neurons | In vitro: Rd (0.1, 1, 10 μM) | Ameliorated Aβ25–35 induced damage in primary cultured hippocampal neurons, inhibited Aβ25–35 induced apoptosis and oxidative stress, reversed Aβ 25–35 induced alterations | ROS↓, MDA↓, GSH-Px↑, SOD↑, Bcl-2↑, Bax↓, Cyt c↓, c-caspase-3↓ | [72] | 2015 | |
| Animals: APP transgenic mice | In vivo: Rd (10 mg/kg) | Improved learning and memory ability in APP transgenic mice | NF-kB↓ | [73] | 2015 | |
| Cell lines: cortical neurons from mice E17–18 embryosAnimals: Aβ1–40 induced AD rat model and APP transgenic mice | In vitro: Rd (2.5, 5 μM, 12 h) In vivo: Rd (5 mg/kg) | Inhibited OA-induced tau phosphorylation in vivo and in vitro | Altered the functional balance of GSK-3β and PP-2A | [74] | 2013 | |
| Cell lines: Neuro-2a | In vitro: Rd (2.5 to 5 µg/mL) | Enhanced the expression of cholinergic markers and neuronal differentiation | ChAT/VAChT↑, ERK and AKT↓; MAP-2↑, p75↑, p21↑, NGF-induced TrkA↑ | [75] | 2014 | |
| Animals: OA induced AD rat model | In vivo: Rd (2.5, 5 μM) | Protected SD rats and cultured cortical neurons against OA-induced toxicity | Decreased OA-induced the hyperphosphorylation of tau by the increase in activities of PP-2A | [76] | 2011 | |
| Animals: ovariectomy (OVX) rat model | In vivo: Rd (10 mg/kg, 2 months) | Enhanced learning and memory function of OVX rats and attenuated cognitive and memory impairment | α-Secretase and sAPPα↑, β-secretase and Aβ↓, p-ER-α at Ser118 residue↑ | [77] | 2017 | |
| Parkinson’s disease (PD) | Cell lines: SH-SY5Y | In vitro: Rd (0.5, 1 μM, 24 h) | Reduced oxidative stress, improved mitochondrial integrity and functions, and inhibited apoptosis | Bax/Bcl-2↓, Cyt c↓, caspase-3↓ | [78] | 2017 |
| Cell lines: SH-SY5Y | In vitro: Rd (1, 10 μM) | Exerted protective effect on neurodegenerative diseases, attenuated MPP+-induced cell death | Oxidative stress↓, mitochondrial function↑ and inhibited MPP+ induced ATP depletion, Bax/Bcl-2↓, Prevented p-AKT downregulation induced by MPP+ treatment | [79] | 2015 | |
| Cells: CCL4-treated primary dopaminergic cell cultures | In vitro: Rd (1, 5, 10 µM) | Protected dopaminergic neurons against CCl4-induced neurotoxicity; inhibited both oxidative stress and inflammation | LDH↓, NO↓, superoxide formation↓ | [80] | 2016 | |
| Neurotoxicity | Animals: lead (Pb)-treated old rat model | In vivo: Rd (50 mg/kg/d, 7 days) | Neuroprotective effects in old rats following acute Pb exposure | IL-1β↓, IL-6↓, TNF-α↓ | [81] | 2013 |
| Cells: TMT-treated hippocampal neurons | In vitro: Rd (1–40 µg/mL, 24 h) | Prevented TMT-induced cell apoptosis; attenuated the tremor seizures and cognitive decline; reduced neuronal loss | Bcl-2↑, Bcl-2↓, caspase-3↓ | [82] | 2017 | |
| Animals: KA-induced ICR mice | In vivo: Rd (50 mg/kg) | Attenuated the KA-induced lethal toxicity | p-ERK↑ and p-CREB↓ | [83] | 2003 | |
| Spinal cord injury (SCI) | Animals: spinal cord injury (SCI) rat model | In vivo: Rd (12.5, 25, 50 mg/kg) | Attenuated SCI-induced secondary injury through reversing the redox-state imbalance, inhibiting the inflammatory response and apoptosis | MAPK↓, MDA↓, GSH and SOD↑, TNF-α, IL-1β↓ | [84] | 2016 |
| Mitochondria isolated from mouse spinal cord tissues Animals: male C57BL/6J mice | In vitro: Rd (0.1, 1, 10 µM, 60 s) In vivo: Rd(10, 50 mg/kg, 7 days) | Protected isolated spinal cord mitochondria against Ca2+ induced MPT and cytochrome c release in a mitochondrial protein kinases-dependent manner | Ca2+ induced Cyt c↓, intramitochondrial AKT and ERK↑ | [85] | 2014 | |
| Stress-related disorders | Animals: immobilization stress (IS) or Escherichia coli (E. coli)-treated anxiety/depression mice model | In vivo: Rd (5 mg/kg/d, oral, 5 days) | Alleviated the IS-induced anxiety/depression and E. coli-induced anxiety/depression, colitis, and gut dysbiosis in mice | Myeloperoxidase activity↓, NF-κB↓, NF-κB+/CD11c+ cell population↓ | [86] | 2020 |
| Animals: CRS induced cognitive impairment mice model | In vivo: Rd (20, 40 mg/kg/d, 28 days) | Improved cognitive impairment subjected to chronic stress | Oxidative stress↓, inflammation↓, hippocampal BDNF-mediated CREB signaling pathway↑ | [87] | 2020 | |
| Animals: chronic cerebral hypoperfusion (CCH) mice model | In vivo: Rd (10, 30 mg/kg/d, 21 days) | Ameliorated CCH-induced impairment of learning and memory behaviors | Neuron survival↑, BDNF expression↑ | [88] | 2016 | |
| Cell lines: mouse adrenocortical tumor cell line Y1 | In vitro: Rd (2 μM) | Inhibited corticosterone secretion in the cells and impeded ACTH-induced corticosterone biosynthesis | cAMP/PKA/CREB signaling pathway↓; attenuated the induction of MC2R and MRAP by ACTH | [89] | 2020 | |
| Noise-induced hearing loss (NIHL) | Animals: noise-induced guinea pigs | In vivo: Rd (30 mg/kg, i.p.) | Exerted neuroprotective effects after noise-induced auditory system damage; ameliorated auditory cortex injury associated with military aviation NIHL | SIRT1/PGC-1α signaling pathway↑ | [90] | 2020 |
| Neural cells | Cells: neural stem cells Animals: male SD rats (180–220 g) | In vitro: Rd (0.1, 1, 10, 50 μM) In vivo: Rd (10, 30 mg/kg) | Had beneficial effects on learning and memory, promoted the size and number of neurospheres; but did not affect the differentiation of neural stem cells into neurons, astrocytes and oligodendrocytes | / | [91] | 2012 |
| Cells: neural stem cells | In vitro: Rd (0.1, 1 μM) | Promoted the differentiation of neurospheres into astrocytes and increased the production of astrocytes | Number of neurons↓, astrocytes↑ | [92] | 2005 | |
| Cell lines: PC12 | In vitro: Rd (10 µM) | Promoted the neurite outgrowth of PC12 cells | GAP-43↑ via ERK and ARK signaling pathways | [93] | 2016 | |
| Cells: rat cortical neurons | In vitro: Rd (1, 3, 10, 30 µM) | Prevented glutamate-induced apoptosis in rat cortical neurons | Inhibited voltage-independent Ca2+ entry | [94] | 2010 |
3.5. Ischemic Stroke
3.6. Cardiovascular Protection
3.7. Immunological Activities
3.8. Others
4. Pharmacokinetics and Clinical Studies
4.1. Preclinical Studies
4.2. Clinical Studies
| Compound | Subject | Dose | Pharmacokinetics Parameters | Ref. | Year | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cmax (ng/mL) | Tmax (h) | AUC (ng h/L) | MRT (h) | CL/F (L·h−1) | T1/2 (h) | |||||
| Preclinical Studies | ||||||||||
| Rd | Dogs | 2 mg/kg, i.g. | 81.0 ± 24.6 | 2.67 ± 1.17 | 1890.2 ± 668.6 | 25.5 ± 3.84 | 1.14 ± 0.40 | 24.2 ± 2.85 | [130] | 2007 |
| 0.2 mg/kg, i.v. | / | / | 76,403.4 ± 15,880.6 | 26.7 ± 1.63 | 0.0020 ± 0.0005 | 39.4 ± 12.0 | ||||
| Rd | Kunming mice Wistar rats | 20 mg/kg, 50 mg/kg, 150 mg/kg, i.v. | / | / | 305.0 ± 22.3 | / | 0.066 ± 0.005 | 14.19 ± 2.37 | [131] | 2012 |
| / | / | 293.2 ± 279.4 | / | 0.280 ± 0.172 | 12.83 ± 2.92 | |||||
| / | / | 312.6 ± 139.5 | / | 0.569 ± 0.306 | 14.02 ± 10.57 | |||||
| GTSSL | SD rats | 400 mg/kg, i.g. | 22.05 ± 2.21 | 2 | 2180.10 ± 18.69 | 12.43 ± 1.46 | / | 7.30 ± 3.32 | [132] | 2015 |
| RG | ICR mice | 2 g/kg/day, 7 days | 51.7 ± 24.7 | 2.8 ± 3.3 | 1145 ± 555.6 | / | / | 40.1 ± 6.1 | [133] | 2020 |
| SD rats | 6.5 ± 1.5 | 7.0 ± 2.0 | 257.8 ± 49.6 | / | / | 94.0 ± 23.7 | ||||
| Rd | Wistar rats—normal | 80 mg/kg, i.g. | 97.458 ± 1.80 | 1.00 ± 0.01 | 2061.658 ± 1011.618 | 13.997 ± 0.390 | 64.895 ± 2.255 | 9.631 ± 0.206 | [136] | 2021 |
| Wistar rats—depression model | 104.959 ± 5.0 | 1.00 ± 0.03 | 2583.439 ± 1254.680 | 15.126 ± 0.671 | 55.744 ± 2.366 | 10.198 ± 0.511 | ||||
| Clinical studies | ||||||||||
| Rd | 199 + 390 patients | 10, 20 mg, i.v. | / | / | / | / | / | / | [99] | 2016 |
| Rd | SD rats | 60 mg/mL, i.v. 150 mg/kg, i.g. | / | / | / | / | / | / | [137] | 2007 |
| 10 healthy Chinese volunteers | 10 mg, i.v. | 2841.18 ± 473.03 | 0.50 ± 0.00 | 27261.63 ± 8116.88 | 17.52 ± 3.73 | 0.39 ± 0.12 | 19.29 ± 3.44 | |||
| Rd | 24 healthy Chinese volunteers | 10 mg | 2.8 ± 0.5 | 0.5 ± 0.0 | 27.3 ± 8.1(mg·h/L) | 17.5 ± 3.7 | 0.39 ± 0.12 | 19.3 ± 3.4 | [138] | 2010 |
| 45 mg | 10.5 ± 1.7 | 0.5 ± 0.0 | 112.6 ± 24.1 (mg·h/L) | 18.3 ± 2.7 | 0.36 ± 0.08 | 18.4 ± 2.9 | ||||
| 75 mg, i.v. | 19.3 ± 2.6 | 0.5 ± 0.0 | 208.4 ± 51.4 (mg·h/L) | 18.6 ± 2.7 | 0.37 ± 0.09 | 17.7 ± 2.0 | ||||
| Rd | 199 patients | 10, 20 mg, i.v. | / | / | / | / | / | / | [139] | 2009 |
| Rd | 390 patients | 10 mg, i.v. | / | / | / | / | / | / | [140] | 2012 |
| RG | 34 healthy Korean volunteers | 3 g, i.g. | 1.77 ± 2.09 | 15.12 ± 9.35 | 7.85 ± 11.24 | / | / | / | [141] | 2020 |
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2k2c | two-kidney, two-clip; |
| 5-FU | 5-fluorouracil; |
| ACh | acetylcholine; |
| Ac-H3 | acetylated histone H3; |
| ACTH | adrenocorticotrophic hormone; |
| AD | Alzheimer’s disease; |
| ADR | Adriamycin; |
| ALI | acute lung injury; |
| AMP | adenosine monophosphate; |
| AMPK | adenosine 5‘-monophosphate (AMP)-activated protein kinase; |
| APP | Aβ-protein precursor; |
| Aβ | amyloid β; |
| BASMCs | basilar artery smooth muscle cells; |
| BAVSMCs | basilar arterial vascular smooth muscle cells; |
| BBB | blood–brain barrier; |
| Bax | Bcl2-Associated X; |
| Bcl-2 | B-cell lymphoma-2; |
| BDNF | brain-derived neurotrophic factor; |
| Bmi-1 | B cell-specific MLV insertion site-1; |
| C/EBP | CCAAT/enhancer binding protein; |
| CA4P | combretastatin A4 phosphate; |
| CaM | calmodulin; |
| CCH | chronic cerebral hypoperfusion; |
| CDX-2 | caudal type homeobox 2; |
| ChAT | choline O-Acetyltransferase; |
| COX | cyclooxygenase; |
| CREB | cAMP-response element binding protein; |
| CRS | chronic restraint stress; |
| CPA | combination of Panax ginseng and Angelica sinensis; |
| cTnI | cardiac troponin I; |
| DAPK1 | death associated protein kinase 1; |
| Drp1 | dynamin-related protein 1; |
| DSS | dextran sulfate sodium; |
| EAE | experimental autoimmune encephalomyelitis; |
| EAN | experimental autoimmune neuritis; |
| ERK | extracellular regulated protein kinases; |
| EGFR | epidermal growth factor receptor; |
| EMT | epithelial–mesenchymal transition; |
| E. coli | Escherichia coli; |
| FGA | floralginsenoside A; |
| fRG | fermented red ginseng; |
| GAP-43 | growth associated protein-43; |
| GBS | Guillain–Barré syndrome; |
| GS-E3D | pectin-lyase-modified ginseng; |
| GSH | glutathione; |
| GSSG | oxidized glutathione; |
| GSH-Px | glutathione peroxidase; |
| HDAC2 | histone deacetylase 2; |
| HO-1 | heme oxygenase-1; |
| HPLC | high performance liquid chromatography; |
| hiPSCs | human induced pluripotent stem cells; |
| hTERT | human telomerase reverse transcriptase; |
| HUVECs | human umbilical vascular endothelial cells; |
| HFs | hair follicles; |
| IBD | inflammatory bowel disease; |
| iNOS | inducible nitric-oxide synthase; |
| IRI | ischemia-reperfusion injury; |
| i.p. | intraperitoneal; |
| i.v. | intravenous; |
| i.g. | intragastrically; |
| IS | immobilization stress; |
| JNK | c-Jun N-terminal kinase; |
| KPCs | keratinocyte progenitor cells; |
| KA | kainic acid; |
| Cyt c | cytochrome c; |
| DMD | Duchenne muscular dystrophy; |
| LPS | lipopolysaccharide; |
| LVEF | left ventricular ejection fraction; |
| MDR1 | multidrug resistance protein 1; |
| MI/R | myocardial ischemia- reperfusion; |
| MMP | mitochondrial membrane potential; |
| MEK | methyl ethyl ketone; |
| MS | multiple sclerosis; |
| Msi-1 | Musashi-1; |
| MPO | myeloperoxidase; |
| MPP+ | 1-methyl-4-phenylpyridinium; |
| MPMC | mouse peritoneal mast cells; |
| MC2R | melanocortin 2 receptor; |
| MCAO | middle cerebral artery occlusion; |
| MDA | malondialdehyde; |
| mTOR | mammalian target of rapamycin; |
| NHA | normal human astrocytes; |
| NEIL | Nei-like DNA glycosylase; |
| NF-κB | nuclear factor kappa-B; |
| NGF | nerve growth factor; |
| NLRP3 | nod-like receptor protein 3; |
| NMDA | N-methyl-D-aspartic acid; |
| TMT | trimethyltin; |
| Tt-Afs | thermostable α-l-arabinofuranosidase; |
| TRPM7 | transient receptor potential melastatin 7; |
| NMDAR | N-methyl-D-aspartic acid receptor; |
| Nr4a1 | nuclear receptor subfamily 4 group A member 1; |
| NRCMs | neonatal rat cardiac myocytes; |
| NRF2 | nuclear factor erythroid 2-related factor 2; |
| NIHL | noise-induced hearing loss; |
| NSC | neural stem cell; |
| NSCLC | non-small-cell lung cancer; |
| OA | okadaic acid; |
| ox-LDL | oxidized low density lipoprotein; |
| PD | Parkinson’s disease; |
| LDH | lactate dehydrogenase; |
| LDL | low-density lipoprotein; |
| PGE2 | prostaglandin E2; |
| LC-ESI-MS/MS | liquid chromatography–electrospray ionization tandem mass spectrometry |
| PPD | protopanaxadiol; |
| Rd | ginsenoside Rd; |
| ROS | reactive oxygen species; |
| ROCC | receptor-operated Ca2+ channels; |
| sAPPα | soluble amyloid precursor protein alpha; |
| SCI | spinal cord injury; |
| SD rats | Sprague–Dawley rats; |
| SIRT1 | sirtuin 1 SAM, senescence-accelerated mice; |
| SR-A | scavenger receptor A; |
| STAT3 | signal transducer and activator of transcription 3; |
| SMI | Shenmai injection; |
| PKA | protein kinase A; |
| SFI | Shenfu injection; |
| SOCC | store-operated Ca2+ channels; |
| TUNEL | TdT-mediated dUTP nick-end labeling; |
| TG | total ginsenoside; |
| TNBS | 2,4,6-trinitrobenzenesulfonic acid; |
| TLC | thin-layer chromatography; |
| TNBS | 2,4,6-trinitrobenzenesulfonic acid; |
| Treg | regulatory T cells; |
| TRPM7 | transient receptor potential melastatin 7; |
| PP-2A | protein phosphatase 2A; |
| SOD | superoxide dismutase; |
| OVX | mice, ovariectomy mice; |
| PGC-1α | peroxisome proliferator-activated receptor-γ coactivator-1α; |
| TGF-β | transforming growth factor-β; |
| Th17 | T helper cell 17; |
| Tt-Afs | thermostable α-l-arabinofuranosidase; |
| TNF-α | tumor necrosis factor-α; |
| ULK1 | unc-51-like autophagy activating kinase 1; |
| VAChT | vesicular acetylcholine transporter; |
| VDCC | voltage dependent Ca2+ channel; |
| GTSSL | total saponins in the stems-leaves of Panax ginseng C. A. Meyer. |
References
- Sharma, A.; Lee, H.-J. Ginsenoside compound K: Insights into recent studies on pharmacokinetics and health-promoting activities. Biomolecules 2020, 10, 1028. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, J.-W.; Li, W.; Ma, H.; Sun, J.; Deng, M.-C.; Yang, L. Ginsenoside metabolites, rather than naturally occurring ginsenosides, lead to inhibition of human cytochrome P450 enzymes. Toxicol. Sci. 2006, 91, 356–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabavi, S.F.; Sureda, A.; Habtemariam, S. Ginsenoside Rd and ischemic stroke; a short review of literatures. J. Ginseng Res. 2015, 39, 299–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-Y.; Liu, Q.-P.; An, P.; Jia, M.; Luan, X.; Tang, J.-Y.; Zhang, H. Ginsenoside Rd: A promising natural neuroprotective agent. Phytomedicine 2022, 95, 153883. [Google Scholar] [CrossRef]
- An, D.-S.; Cui, C.-H.; Sung, B.H.; Yang, H.-C.; Kim, S.C.; Lee, S.-T.; Im, W.-T.; Kim, S.-G. Characterization of a novel ginsenoside-hydrolyzing α-l-arabinofuranosidase, AbfA, from Rhodanobacter ginsenosidimutans Gsoil 3054T. Appl. Microbiol. Biotechnol. 2012, 94, 673–682. [Google Scholar] [CrossRef]
- Liu, Q.-M.; Jung, H.-M.; Cui, C.-H.; Sung, B.-H.; Kim, J.-K.; Kim, S.-G.; Lee, S.-T.; Kim, S.-C.; Im, W.-T. Bioconversion of ginsenoside Rc into Rd by a novel α-l-arabinofuranosidase, Abf22-3 from Leuconostoc sp. 22-3: Cloning, expression, and enzyme characterization. Antonie Leeuwenhoek 2013, 103, 747–754. [Google Scholar] [CrossRef]
- Shin, K.-C. Production of Ginsenoside Rd from Ginsenoside Rc by α-L-Arabinofuranosidase from Caldicellulosiruptor saccharolyticus. J. Microbiol. Biotechnol. 2013, 23, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Zhao, D.; Zhao, L.; Pei, J.; Xiao, W.; Ding, G.; Wang, Z.; Xu, J. Characterization of a novel arabinose-tolerant α-l- arabinofuranosidase with high ginsenoside Rc to ginsenoside Rd bioconversion productivity. J. Appl. Microbiol. 2016, 120, 647–660. [Google Scholar] [CrossRef]
- Zhang, R.; Tan, S.; Zhang, B.; Guo, Z.; Tian, L.; Weng, P.; Luo, Z. Two key amino acids variant of α-L-arabinofuranosidase from Bacillus subtilis Str. 168 with altered activity for selective conversion ginsenoside Rc to Rd. Molecules 2021, 26, 1733. [Google Scholar] [CrossRef]
- Kim, J.-H.; Oh, J.-M.; Chun, S.; Park, H.Y.; Im, W.-T. Enzymatic Biotransformation of ginsenoside Rb2 into Rd by recombinant α-L-Arabinopyranosidase from Blastococcus saxobsidens. J. Microbiol. Biotechnol. 2020, 30, 391–397. [Google Scholar] [CrossRef]
- Zhao, L.; Xie, J.; Zhang, X.; Cao, F.; Pei, J. Overexpression and characterization of a glucose-tolerant β-glucosidase from Thermotoga thermarum DSM 5069T with high catalytic efficiency of ginsenoside Rb1 to Rd. J. Mol. Catal. B Enzym. 2013, 95, 62–69. [Google Scholar] [CrossRef]
- Quan, L.-H.; Min, J.-W.; Jin, Y.; Wang, C.; Kim, Y.-J.; Yang, D.-C. Enzymatic biotransformation of ginsenoside Rb1 to compound K by recombinant β-glucosidase from Microbacterium esteraromaticum. J. Agric. Food Chem. 2012, 60, 3776–3781. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.-H.; Wang, C.; Jin, Y.; Wang, T.-R.; Kim, Y.-J.; Yang, D.C. Isolation and characterization of novel ginsenoside-hydrolyzing glycosidase from Microbacterium esteraromaticum that transforms ginsenoside Rb2 to rare ginsenoside 20(S)-Rg3. Antonie Leeuwenhoek 2013, 104, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.-L.; Ma, R.; Jiang, M.; Dong, W.-W.; Jiang, J.; Wu, S.; Li, D.; Quan, L.-H. Cloning and characterization of ginsenoside-hydrolyzing β-glucosidase from Lactobacillus brevis that transforms ginsenosides Rb1 and F2 into ginsenoside Rd and compound K. J. Microbiol. Biotechnol. 2016, 26, 1661–1667. [Google Scholar] [CrossRef]
- Chang, K.H.; Na Jo, M.; Kim, K.-T.; Paik, H.-D. Purification and characterization of a ginsenoside Rb1-hydrolyzing β-glucosidase from Aspergillus niger KCCM 11239. Int. J. Mol. Sci. 2012, 13, 12140–12152. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Wei, Y.; Li, Y.; Zhou, G. One-pot process for the production of ginsenoside Rd by coupling enzyme-assisted extraction with selective enzymolysis. Biol. Pharm. Bull. 2020, 43, 1443–1447. [Google Scholar] [CrossRef]
- Ye, L.; Zhou, C.-Q.; Zhou, W.; Zhou, P.; Chen, D.-F.; Liu, X.-H.; Shi, X.-L.; Feng, M.-Q. Biotransformation of ginsenoside Rb1 to ginsenoside Rd by highly substrate-tolerant Paecilomyces bainier 229-7. Bioresour. Technol. 2010, 101, 7872–7876. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, C.; Li, J.; Shi, X.; Feng, M. Effects of external calcium on the biotransformation of ginsenoside Rb1 to ginsenoside Rd by Paecilomyces bainier 229-7. World J. Microbiol. Biotechnol. 2012, 28, 857–863. [Google Scholar] [CrossRef]
- Lin, F.; Guo, X.; Lu, W. Efficient biotransformation of ginsenoside Rb1 to Rd by isolated Aspergillus versicolor, excreting β-glucosidase in the spore production phase of solid culture. Antonie Leeuwenhoek 2015, 108, 1117–1127. [Google Scholar] [CrossRef]
- Feng, L.; Xu, C.; Li, Z.; Li, J.; Dai, Y.; Han, H.; Yu, S.; Liu, S. Microbial conversion of ginsenoside Rd from Rb1 by the fungus mutant Aspergillus niger strain TH-10a. Prep. Biochem. Biotechnol. 2016, 46, 336–341. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.-H.; Lee, P.Y.; Bae, K.-H.; Cho, S.; Park, B.C.; Shin, H.; Park, S.G. Ginsenoside Rb1 is transformed into Rd and Rh2 by Microbacterium trichothecenolyticum. J. Microbiol. Biotechnol. 2013, 23, 1802–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akter, S.; Huq, A. Biological synthesis of ginsenoside Rd using Paenibacillus horti sp. nov. isolated from vegetable garden. Curr. Microbiol. 2018, 75, 1566–1573. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Wang, X.; Lee, S.-Y.; Rahman, M.M.; Park, J.-H.; Siddiqi, M.Z.; Balusamy, S.R.; Nam, K.; Rahman, S.; Huq, A. Paenibacillus roseus sp. nov., a ginsenoside-transforming bacterium isolated from forest soil. Arch. Microbiol. 2021, 203, 3997–4004. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K. Novosphingobium ginsenosidimutans sp. nov., with the ability to convert ginsenoside. J. Microbiol. Biotechnol. 2013, 23, 444–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Xu, Y.; Gu, M.; Liu, Z.; Zhang, J.; Zeng, Q.; Zhu, D. Biotransformation of ginsenoside Rc to Rd by endophytic bacterium Bacillus sp. G9y isolated from Panax quinquefolius. Antonie Leeuwenhoek 2021, 114, 437–444. [Google Scholar] [CrossRef]
- Shen, H.; Leung, W.-I.; Ruan, J.-Q.; Li, S.-L.; Lei, J.P.-C.; Wang, Y.-T.; Yan, R. Biotransformation of ginsenoside Rb1 via the gypenoside pathway by human gut bacteria. Chin. Med. 2013, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, L.-H.; Piao, J.-Y.; Min, J.-W.; Kim, H.-B.; Kim, S.-R.; Yang, D.-U.; Yang, D.-C. Biotransformation of ginsenoside Rb1 to prosapogenins, gypenoside XVII, ginsenoside Rd, ginsenoside F2, and compound K by Leuconostoc mesenteroides DC102. J. Ginseng Res. 2011, 35, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Quan, L.-H.; Kim, Y.-J.; Li, G.H.; Choi, K.-T.; Yang, D.-C. Microbial transformation of ginsenoside Rb1 to compound K by Lactobacillus paralimentarius. World J. Microbiol. Biotechnol. 2013, 29, 1001–1007. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Duan, F.; Liu, A.; Li, S.; Zhong, W.; Sheng, W.; Chen, J.; Xu, J.; Xiao, S. Prebiotics enhance the biotransformation and bioavailability of ginsenosides in rats by modulating gut microbiota. J. Ginseng Res. 2021, 45, 334–343. [Google Scholar] [CrossRef]
- Ku, S.; You, H.J.; Park, M.S.; Ji, G.E. Whole-cell biocatalysis for producing ginsenoside Rd from Rb1 using Lactobacillus rhamnosus GG. J. Microbiol. Biotechnol. 2016, 26, 1206–1215. [Google Scholar] [CrossRef]
- Renchinkhand, G.; Cho, S.H.; Park, Y.W.; Song, G.-Y.; Nam, M.S. Biotransformation of major ginsenoside Rb1 to Rd by Dekkera anomala YAE-1 from mongolian fermented milk (Airag). J. Microbiol. Biotechnol. 2020, 30, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, Z.; Chen, H.; Brand, E.; Yi, T.; Qin, M.; Liang, Z. Determination of ginsenosides in Asian and American ginsengs by liquid chromatography–quadrupole/time-of-flight MS: Assessing variations based on morphological characteristics. J. Ginseng Res. 2017, 41, 10–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.-Z.; Liu, Y.-P.; Tian, S.-C.; Ge, S.-Y.; Wu, Y.-J.; Zhang, B.-L. Antitumor activity of ginsenoside Rd in gastric cancer via up-regulation of caspase-3 and caspase-9. Pharmazie 2020, 75, 147–150. [Google Scholar] [PubMed]
- Yang, Z.-G.; Sun, H.-X.; Ye, Y.-P. Ginsenoside Rd from Panax notoginseng is cytotoxic towards HeLa cancer cells and induces apoptosis. Chem. Biodivers. 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Wang, J.; Song, Y.; Wang, Q.; Wu, Q. The inhibitory effects of ginsenoside Rd on the human glioma U251 cells and its underlying mechanisms. J. Cell. Biochem. 2019, 120, 4444–4450. [Google Scholar] [CrossRef]
- Liu, G.-M.; Lu, T.-C.; Sun, M.-L.; Jia, W.-Y.; Ji, X.; Luo, Y.-G. Ginsenoside Rd inhibits glioblastoma cell proliferation by up-regulating the expression of miR-144-5p. Biol. Pharm. Bull. 2020, 43, 1534–1541. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, G.T.; Roh, S.H.; Song, J.S.; Kim, H.J.; Hong, S.S.; Kwon, S.W.; Park, J.H. Proteome changes related to the anti-cancer activity of HT29 cells by the treatment of ginsenoside Rd. Pharmazie 2009, 64, 242–247. [Google Scholar] [CrossRef]
- Yang, X.; Gao, M.; Miao, M.; Jiang, C.; Zhang, D.; Yin, Z.; Ni, Y.; Chen, J.; Zhang, J. Combining combretastatin A4 phosphate with ginsenoside Rd synergistically inhibited hepatocellular carcinoma by reducing HIF-1α via PI3K/AKT/mTOR signalling pathway. J. Pharm. Pharmacol. 2021, 73, 263–271. [Google Scholar] [CrossRef]
- Liu, H.; Dilger, J.P.; Lin, J. The role of transient receptor potential melastatin 7 (TRPM7) in cell viability: A potential target to suppress breast cancer cell cycle. Cancers 2020, 12, 131. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.J. Involvement of melastatin type transient receptor potential 7 channels in ginsenoside Rd-induced apoptosis in gastric and breast cancer cells. J. Ginseng Res. 2013, 37, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-H.; Choi, Y.-J.; Cha, S.-W.; Lee, S.-G. Anti-metastatic effects of ginsenoside Rd via inactivation of MAPK signaling and induction of focal adhesion formation. Phytomedicine 2012, 19, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Phi, L.; Sari, I.N.; Wijaya, Y.T.; Kim, K.S.; Park, K.C.; Cho, A.E.; Kwon, H.Y. Ginsenoside Rd inhibits the metastasis of colorectal cancer via epidermal growth factor receptor signaling axis. IUBMB Life 2019, 71, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Du, X.; Xiong, M.; Cui, J.; Yang, Q.; Wang, W.; Chen, Y.; Zhang, T. Ginsenoside Rd attenuates breast cancer metastasis implicating derepressing microRNA-18a-regulated Smad2 expression. Sci. Rep. 2016, 6, srep33709. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Shi, H.; Yang, L.; Wu, X.; Wang, Z. Ginsenoside Rd regulates the Akt/mTOR/p70S6K signaling cascade and suppresses angiogenesis and breast tumor growth. Oncol. Rep. 2017, 38, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, C.; Jiang, C.; Ni, S.; Wang, Q.; Cheng, L.; Wang, H.; Zhang, Q.; Liu, W.; Zhang, J.; Liu, J.; et al. Identification of bioactive anti-angiogenic components targeting tumor endothelial cells in Shenmai injection using multidimensional pharmacokinetics. Acta Pharm. Sin. B 2020, 10, 1694–1708. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, Y.R.; Kim, N.D.; Han, H.-K.; Oh, W.K.; Kang, K.W. Increased ubiquitination of multidrug resistance 1 by ginsenoside Rd. Nutr. Cancer 2010, 62, 252–259. [Google Scholar] [CrossRef]
- Chian, S.; Zhao, Y.; Xu, M.; Yu, X.; Ke, X.; Gao, R.; Yin, L. Ginsenoside Rd reverses cisplatin resistance in non-small-cell lung cancer A549 cells by downregulating the nuclear factor erythroid 2-related factor 2 pathway. Anti-Cancer Drugs 2019, 30, 838–845. [Google Scholar] [CrossRef]
- Huang, G.; Khan, I.; Li, X.; Chen, L.; Leong, W.; Ho, L.T.; Hsiao, W.L.W. Ginsenosides Rb3 and Rd reduce polyps formation while reinstate the dysbiotic gut microbiota and the intestinal microenvironment in ApcMin/+ mice. Sci. Rep. 2017, 7, 12552. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.M.; Lee, D.K.M.; Wong, D.P.; Wong, R.N.; Yung, K.K.; Cheng, C.H.-K.; Yue, K.K. Ginsenosides attenuate methylglyoxal-induced impairment of insulin signaling and subsequent apoptosis in primary astrocytes. Neuropharmacology 2014, 85, 215–223. [Google Scholar] [CrossRef]
- Kaviani, M.; Keshtkar, S.; Azarpira, N.; Aghdaei, M.H.; Geramizadeh, B.; Karimi, M.H.; Yaghobi, R.; Esfandiari, E.; Shamsaeefar, A.; Nikeghbalian, S.; et al. Cytoprotective effects of ginsenoside Rd on apoptosis-associated cell death in the isolated human pancreatic islets. EXCLI J. 2019, 18, 666–676. [Google Scholar] [CrossRef]
- Jung, E.; Pyo, M.-K.; Kim, J. Pectin-lyase-modified ginseng extract and ginsenoside Rd inhibits high glucose-induced ROS production in mesangial cells and prevents renal dysfunction in db/db mice. Molecules 2021, 26, 367. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Qin, W.; Wei, R.; Jiang, Y.; Fan, L.; Wang, Z.; Tan, N. Ginsenoside Rd ameliorates high glucose-induced retinal endothelial injury through AMPK-STRT1 interdependence. Pharmacol. Res. 2022, 106123, in press. [Google Scholar] [CrossRef] [PubMed]
- Song, S.-B.; Tung, N.H.; Quang, T.H.; Ngan, N.T.T.; Kim, K.-E.; Kim, Y.-H. Inhibition of TNF-α-mediated NF-κB transcriptional activity in HepG2 cells by dammarane-type saponins from Panax ginseng leaves. J. Ginseng Res. 2012, 36, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.-M.; Zhang, Y.-M.; Moldzio, R.; Rausch, W.-D. Ginsenoside Rd attenuates neuroinflammation of dopaminergic cells in culture. J. Neural Suppl. 2007, 72, 105–112. [Google Scholar] [CrossRef]
- Shin, J.-S.; Park, N.; Ra, J.; Kim, Y.; Shin, M.; Hong, M.; Kim, S.-H.; Kwon, H.-J.; Hong, S.-P.; Kim, J.; et al. Panax ginseng C.A. Meyer modulates the levels of MMP3 in S12 murine articular cartilage cell line. J. Ethnopharmacol. 2009, 124, 397–403. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Sugimoto, S.; Nakamura, S.; Sakumae, H.; Matsuda, H. Medicinal flowers. XVI. new dammarane-type triterpene tetraglycosides and gastroprotective principles from flower buds of Panax ginseng. Chem. Pharm. Bull. 2007, 55, 1034–1038. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Chung, J.H.; Yoon, J.S.; Ha, Y.M.; Bae, S.; Lee, E.K.; Jung, K.J.; Kim, M.S.; Kim, Y.J.; Kim, M.K.; et al. Ginsenoside Rd inhibits the expressions of iNOS and COX-2 by suppressing NF-κB in LPS-stimulated RAW264.7 cells and mouse liver. J. Ginseng Res. 2013, 37, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M. Anti-inflammatory Effects of Ginsenosides Rg5, Rz1, and Rk1: Inhibition of TNF-α-induced NF-κB, COX-2, and iNOS transcriptional expression. Phytother. Res. 2014, 28, 1893–1896. [Google Scholar] [CrossRef]
- Liu, C.; Wang, J.; Yang, Y.; Liu, X.; Zhu, Y.; Zou, J.; Peng, S.; Le, T.H.; Chen, Y.; Zhao, S.; et al. Ginsenoside Rd ameliorates colitis by inducing p62-driven mitophagy-mediated NLRP3 inflammasome inactivation in mice. Biochem. Pharmacol. 2018, 155, 366–379. [Google Scholar] [CrossRef]
- Yang, X.-L.; Guo, T.-K.; Wang, Y.-H.; Gao, M.-T.; Qin, H.; Wu, Y.-J. Therapeutic effect of ginsenoside Rd in rats with TNBS-induced recurrent ulcerative colitis. Arch. Pharmacal Res. 2012, 35, 1231–1239. [Google Scholar] [CrossRef]
- Yang, X.-L.; Guo, T.-K.; Wang, Y.-H.; Huang, Y.-H.; Liu, X.; Wang, X.-X.; Li, W.; Zhao, X.; Wang, L.-P.; Yan, S.; et al. Ginsenoside Rd attenuates the inflammatory response via modulating p38 and JNK signaling pathways in rats with TNBS-induced relapsing colitis. Int. Immunopharmacol. 2012, 12, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Liang, G.; Lin, J.; Zhang, S.; Lin, Q.; Ji, X.; Chen, H.; Li, N.; Jin, S. Ginsenoside Rd therapy improves histological and functional recovery in a rat model of inflammatory bowel disease. Phytother. Res. 2020, 34, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Kim, J.-K.; Kim, J.-Y.; Han, M.J.; Kim, D.-H. Fermented red ginseng and ginsenoside Rd alleviate ovalbumin-induced allergic rhinitis in mice by suppressing IgE, interleukin-4, and interleukin-5 expression. J. Ginseng Res. 2019, 43, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Wang, Z.; Li, S.; Min, G.; Wang, L.; Chen, J.; Cheng, J.; Wu, Y. Inhibitory effect of ginsenoside-Rd on carrageenan-induced inflammation in rats. Can. J. Physiol. Pharmacol. 2012, 90, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-X.; Wang, L.; Xiao, E.-L.; Li, S.-J.; Chen, J.-J.; Gao, B.; Min, G.-N.; Wang, Z.-P.; Wu, Y.-J. Ginsenoside-Rd exhibits anti-inflammatory activities through elevation of antioxidant enzyme activities and inhibition of JNK and ERK activation in vivo. Int. Immunopharmacol. 2013, 17, 1094–1100. [Google Scholar] [CrossRef]
- Yokozawa, T.; Satoh, A.; Cho, E.J. Ginsenoside-Rd attenuates oxidative damage related to aging in senescence-accelerated mice. J. Pharm. Pharmacol. 2004, 56, 107–113. [Google Scholar] [CrossRef]
- Yu, X.; Li, H.; Lin, D.; Guo, W.; Xu, Z.; Wang, L.; Guan, S. Ginsenoside prolongs the lifespan of C. elegans via lipid metabolism and activating the stress response signaling pathway. Int. J. Mol. Sci. 2021, 22, 9668. [Google Scholar] [CrossRef]
- Ye, R.; Han, J.; Kong, X.; Zhao, L.; Cao, R.; Rao, Z.; Zhao, G. Protective effects of ginsenoside Rd on PC12 cells against hydrogen peroxide. Biol. Pharm. Bull. 2008, 31, 1923–1927. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.D.; Pokharel, Y.R.; Kang, K.W. Ginsenoside Rd enhances glutathione levels in H4IIE cells via NF-kappaB-dependent gamma-glutamylcysteine ligase induction. Pharmazie 2007, 62, 933–936. [Google Scholar]
- Ballard, C.; Gauthier, S.; Corbett, A.; Brayne, C.; Aarsland, D.; Jones, E. Alzheimer’s disease. Lancet 2011, 377, 1019–1031. [Google Scholar] [CrossRef]
- Liu, J.; Yan, X.; Li, L.; Zhu, Y.; Qin, K.; Zhou, L.; Sun, D.; Zhang, X.; Ye, R.; Zhao, G. Ginsennoside Rd attenuates cognitive dysfunction in a rat model of Alzheimer’s disease. Neurochem. Res. 2012, 37, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-F.; Yan, X.-D.; Qi, L.-S.; Li, L.; Hu, G.-Y.; Li, P.; Zhao, G. Ginsenoside Rd attenuates Aβ25–35-induced oxidative stress and apoptosis in primary cultured hippocampal neurons. Chem.-Biol. Interact. 2015, 239, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, X.; Li, L.; Li, Y.; Zhou, L.; Zhang, X.; Hu, X.; Zhao, G. Ginsenoside Rd improves learning and memory ability in APP transgenic mice. J. Mol. Neurosci. 2015, 57, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Z.; Liu, J.; Tai, X.; Hu, X.; Liu, X.; Wu, Z.; Zhang, G.; Shi, M.; Zhao, G. Ginsenoside Rd attenuates beta-amyloid-induced tau phosphorylation by altering the functional balance of glycogen synthase kinase 3beta and protein phosphatase 2A. Neurobiol. Dis. 2013, 54, 320–328. [Google Scholar] [CrossRef]
- Kim, M.S.; Yu, J.M.; Kim, H.J.; Kim, H.B.; Kim, S.T.; Kil Jang, S.; Choi, Y.W.; Lee, D.I.; Joo, S.S. Ginsenoside Re and Rd enhance the expression of cholinergic markers and neuronal differentiation in neuro-2a cells. Biol. Pharm. Bull. 2014, 37, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, J.; Yan, X.; Qin, K.; Shi, M.; Lin, T.; Zhu, Y.; Kang, T.; Zhao, G. Protective effects of ginsenoside Rd against okadaic acid-induced neurotoxicity in vivo and in vitro. J. Ethnopharmacol. 2011, 138, 135–141. [Google Scholar] [CrossRef]
- Yan, X.; Hu, G.; Yan, W.; Chen, T.; Yang, F.; Zhang, X.; Zhao, G.; Liu, J. Ginsenoside Rd promotes non-amyloidogenic pathway of amyloid precursor protein processing by regulating phosphorylation of estrogen receptor alpha. Life Sci. 2017, 168, 16–23. [Google Scholar] [CrossRef]
- González-Burgos, E.; Fernández-Moriano, C.; Lozano, R.; Iglesias, I.; Gómez-Serranillos, M. Ginsenosides Rd and Re co-treatments improve rotenone-induced oxidative stress and mitochondrial impairment in SH-SY5Y neuroblastoma cells. Food Chem. Toxicol. 2017, 109, 38–47. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, R.-Y.; Zhao, J.; Dong, Z.; Feng, D.-Y.; Wu, R.; Shi, M.; Zhao, G. Ginsenoside Rd protects SH-SY5Y cells against 1-methyl-4-phenylpyridinium induced injury. Int. J. Mol. Sci. 2015, 16, 14395–14408. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, Y.; Ma, C.; Yan, Y.; Yang, Y.; Wang, X.; Rausch, W.D. Ginsenoside Rd and ginsenoside Re offer neuroprotection in a novel model of Parkinson’s disease. Am. J. Neurodegener. Dis. 2016, 5, 52–61. [Google Scholar]
- Wang, B.; Feng, G.; Tang, C.; Wang, L.; Cheng, H.; Zhang, Y.; Ma, J.; Shi, M.; Zhao, G. Ginsenoside Rd maintains adult neural stem cell proliferation during lead-impaired neurogenesis. Neurol. Sci. 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Xue, J.; Lee, M.; Sung, C. Ginsenoside Rd as a potential neuroprotective agent prevents trimethyltin injury. Biomed. Rep. 2017, 6, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-K.; Choi, S.-S.; Lee, H.-K.; Han, K.-J.; Han, E.-J.; Suh, H.-W. Effects of ginsenoside Rd and decursinol on the neurotoxic responses induced by kainic acid in mice. Planta Med. 2003, 69, 230–234. [Google Scholar] [CrossRef]
- Cong, L.; Chen, W. Neuroprotective effect of ginsenoside Rd in spinal cord injury rats. Basic Clin. Pharmacol. Toxicol. 2016, 119, 193–201. [Google Scholar] [CrossRef]
- Zhou, J.-S.; Wang, J.-F.; He, B.-R.; Cui, Y.-S.; Fang, X.-Y.; Ni, J.-L.; Chen, J.; Wang, K.-Z. Ginsenoside Rd attenuates mitochondrial permeability transition and cytochrome c release in isolated spinal cord mitochondria: Involvement of kinase-mediated pathways. Int. J. Mol. Sci. 2014, 15, 9859–9877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.-K.; Joo, M.-K.; Kim, J.-K.; Jeung, W.; Kang, H.; Kim, D.-H. Bifidobacteria-fermented red ginseng and its constituents ginsenoside Rd and protopanaxatriol alleviate anxiety/depression in mice by the amelioration of gut dysbiosis. Nutrients 2020, 12, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Jiang, N.; Lv, J.; Huang, H.; Liu, X. Ginsenoside Rd reverses cognitive deficits by modulating BDNF-dependent CREB pathway in chronic restraint stress mice. Life Sci. 2020, 258, 118107. [Google Scholar] [CrossRef]
- Wan, Q.; Ma, X.; Zhang, Z.-J.; Sun, T.; Xia, F.; Zhao, G.; Wu, Y.-M. Ginsenoside reduces cognitive impairment during chronic cerebral hypoperfusion through brain-derived neurotrophic factor regulated by epigenetic modulation. Mol. Neurobiol. 2017, 54, 2889–2900. [Google Scholar] [CrossRef]
- Jin, W.; Ma, R.; Zhai, L.; Xu, X.; Lou, T.; Huang, Q.; Wang, J.; Zhao, D.; Li, X.; Sun, L. Ginsenoside Rd attenuates ACTH-induced corticosterone secretion by blocking the MC2R-cAMP/PKA/CREB pathway in Y1 mouse adrenocortical cells. Life Sci. 2020, 245, 117337. [Google Scholar] [CrossRef]
- Chen, X.-M.; Ji, S.-F.; Liu, Y.-H.; Xue, X.-M.; Xu, J.; Gu, Z.-H.; Deng, S.-L.; Liu, C.-D.; Wang, H.; Chang, Y.-M.; et al. Ginsenoside Rd ameliorates auditory cortex injury associated with military aviation noise-induced hearing loss by activating SIRT1/PGC-1α signaling pathway. Front. Physiol. 2020, 11, 788. [Google Scholar] [CrossRef]
- Lin, T.; Liu, Y.; Shi, M.; Liu, X.; Li, L.; Liu, Y.; Zhao, G. Promotive effect of ginsenoside Rd on proliferation of neural stem cells in vivo and in vitro. J. Ethnopharmacol. 2012, 142, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Hao, Q.; Bouissac, J.; Lu, Y.; Tian, S.; Luu, B. Ginsenoside-Rd from Panax notoginseng enhances astrocyte differentiation from neural stem cells. Life Sci. 2005, 76, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-D.; Xia, F.; Lin, X.-M.; Duan, K.-L.; Wang, F.; Lu, Q.-L.; Cao, H.; Qian, Y.-H.; Shi, M. Ginsenoside-Rd promotes neurite outgrowth of PC12 cells through MAPK/ERK- and PI3K/AKT-dependent pathways. Int. J. Mol. Sci. 2016, 17, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-Y.; Liang, J.; Tang, Y.-B.; Zhou, J.-G.; Guan, Y.-Y. Ginsenoside Rd prevents glutamate-induced apoptosis in rat cortical neurons. Clin. Exp. Pharmacol. Physiol. 2010, 37, 199–204. [Google Scholar] [CrossRef]
- Ye, R.; Zhao, G.; Liu, X. Ginsenoside Rd for acute ischemic stroke: Translating from bench to bedside. Expert Rev. Neurother. 2013, 13, 603–613. [Google Scholar] [CrossRef]
- Yang, L.-X.; Zhang, X.; Zhao, G. Ginsenoside Rd attenuates DNA damage by increasing expression of DNA glycosylase endonuclease VIII-like proteins after focal cerebral ischemia. Chin. Med. J. 2016, 129, 1955–1962. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Xu, H.; Hu, G.; Zhang, X.; Xie, Z.; Feng, D.; Wu, R.; Zhao, G.; Shi, M. Protopanaxadiol ginsenoside Rd protects against NMDA receptor-mediated excitotoxicity by attenuating calcineurin-regulated DAPK1 activity. Sci. Rep. 2020, 10, 8078. [Google Scholar] [CrossRef]
- Xie, Z.; Shi, M.; Zhang, C.; Zhao, H.; Hui, H.; Zhao, G. Ginsenoside Rd protects against cerebral ischemia–reperfusion injury via decreasing the expression of the NMDA receptor 2B subunit and its phosphorylated product. Neurochem. Res. 2016, 41, 2149–2159. [Google Scholar] [CrossRef]
- Zhang, G.; Xia, F.; Zhang, Y.; Zhang, X.; Cao, Y.; Wang, L.; Liu, X.; Zhao, G.; Shi, M. Ginsenoside Rd is efficacious against acute ischemic stroke by suppressing microglial proteasome-mediated inflammation. Mol. Neurobiol. 2016, 53, 2529–2540. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Hu, G.; Zhang, G.; Zhao, G.; Shi, M. Ginsenoside Rd attenuates blood-brain barrier damage by suppressing proteasome-mediated signaling after transient forebrain ischemia. NeuroReport 2020, 31, 466–472. [Google Scholar] [CrossRef]
- Hu, J.; Zeng, C.; Wei, J.; Duan, F.; Liu, S.; Zhao, Y.; Tan, H. The combination of Panax ginseng and Angelica sinensis alleviates ischemia brain injury by suppressing NLRP3 inflammasome activation and microglial pyroptosis. Phytomedicine 2020, 76, 153251. [Google Scholar] [CrossRef]
- Guan, Y.-Y.; Zhou, J.-G.; Zhang, Z.; Wang, G.-L.; Cai, B.-X.; Hong, L.; Qiu, Q.-Y.; He, H. Ginsenoside-Rd from panax notoginseng blocks Ca2+ influx through receptor- and store-operated Ca2+ channels in vascular smooth muscle cells. Eur. J. Pharmacol. 2006, 548, 129–136. [Google Scholar] [CrossRef]
- Cai, B.-X.; Li, X.-Y.; Chen, J.-H.; Tang, Y.-B.; Wang, G.-L.; Zhou, J.-G.; Qui, Q.-Y.; Guan, Y.-Y. Ginsenoside-Rd, a new voltage-independent Ca2+ entry blocker, reverses basilar hypertrophic remodeling in stroke-prone renovascular hypertensive rats. Eur. J. Pharmacol. 2009, 606, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Y.; Wang, X.-G.; Ma, M.-M.; Liu, Y.; Du, Y.-H.; Lv, X.-F.; Zhou, J.-G.; Tang, Y.-B.; Guan, Y.-Y. Ginsenoside-Rd potentiates apoptosis induced by hydrogen peroxide in basilar artery smooth muscle cells through the mitochondrial pathway. Apoptosis 2012, 17, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, Z.-Z.; Tang, Y.-B.; Zhou, J.-G.; Guan, Y.-Y. Ginsenoside-Rd, a purified component from panax notoginseng saponins, prevents atherosclerosis in apoE knockout mice. Eur. J. Pharmacol. 2011, 652, 104–110. [Google Scholar] [CrossRef]
- Lu, C.; Sun, Z.; Wang, L. Inhibition of L-type Ca2+ current by ginsenoside Rd in rat ventricular myocytes. J. Ginseng Res. 2015, 39, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; An, X.; Lang, P.; Wang, F.; Xie, Y. Ginsenoside Rd contributes the attenuation of cardiac hypertrophy in vivo and in vitro. Biomed. Pharmacother. 2019, 109, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Wang, X.; Lau, W.; Wang, Y.; Xing, Y.; Zhang, X.; Ma, X.; Gao, F. Ginsenoside Rd attenuates myocardial ischemia/reperfusion injury via Akt/GSK-3β signaling and inhibition of the mitochondria-dependent apoptotic pathway. PLoS ONE 2013, 8, e70956. [Google Scholar] [CrossRef]
- Zeng, X.; Li, J.; Li, Z. Ginsenoside Rd mitigates myocardial ischemia-reperfusion injury via Nrf2/HO-1 signaling pathway. Int. J. Clin. Exp. Med. 2015, 8, 14497–14504. [Google Scholar]
- Zhang, B.; Hu, X.; Wang, H.; Wang, R.; Sun, Z.; Tan, X.; Liu, S.; Wang, H. Effects of a dammarane-type saponin, ginsenoside Rd, in nicotine-induced vascular endothelial injury. Phytomedicine 2020, 79, 153325. [Google Scholar] [CrossRef]
- Zhu, D.; Liu, M.; Yang, Y.; Ma, L.; Jiang, Y.; Zhou, L.; Huang, Q.; Pi, R.; Chen, X. Ginsenoside Rd ameliorates experimental autoimmune encephalomyelitis in C57BL/6 mice. J. Neurosci. Res. 2014, 92, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.; Zhang, C.; Geng, Y.; Liu, M. Therapeutic effect of ginsenoside Rd on experimental autoimmune encephalomyelitis model mice: Regulation of inflammation and Treg/Th17 cell balance. Mediat. Inflamm. 2020, 2020, 8827527. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Li, S.; Ding, J.; Zhao, S.; Liang, S.; Cao, X.; Su, C.; Guo, J. Ginsenoside Rd attenuates mouse experimental autoimmune neuritis by modulating monocyte subsets conversion. Biomed. Pharmacother. 2021, 138, 111489. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Chen, J.; Li, S.; Wang, Y.; Hu, L.; Wang, L.; Wu, Y. Immunosuppressive effects of ginsenoside-Rd on skin allograft rejection in rats. J. Surg. Res. 2012, 176, 267–274. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, A.; Sun, H.; Ye, Y.; Fang, W. Ginsenoside Rd elicits Th1 and Th2 immune responses to ovalbumin in mice. Vaccine 2007, 25, 161–169. [Google Scholar] [CrossRef]
- Han, Y.; Rhew, K.Y. Ginsenoside Rd induces protective anti-Candida albicans antibody through immunological adjuvant activity. Int. Immunopharmacol. 2013, 17, 651–657. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, Y.; Yang, Y.; Hu, Y.; Zou, X.; Yu, B.; Qi, J. Allergens in red ginseng extract induce the release of mediators associated with anaphylactoid reactions. J. Transl. Med. 2017, 15, 148. [Google Scholar] [CrossRef] [Green Version]
- Yokozawa, T.; Liu, Z.W. The role of ginsenoside-rd in cisplatin-induced acute renal failure. Ren. Fail. 2000, 22, 115–127. [Google Scholar] [CrossRef]
- Yokozawa, T.; Dong, E. Role of ginsenoside-Rd in cisplatin-induced renal injury: Special reference to DNA fragmentation. Nephron Exp. Nephrol. 2001, 89, 433–438. [Google Scholar] [CrossRef]
- Ren, K.; Jin, C.; Ma, P.; Ren, Q.; Jia, Z.; Zhu, D. Ginsenoside Rd alleviates mouse acute renal ischemia/reperfusion injury by modulating macrophage phenotype. J. Ginseng Res. 2016, 40, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Wang, R.; Ji, L.-L.; Li, X.-P.; Zhou, H.-G.; He, Z.-K.; Xu, H.-L.; Meng, F.-J.; Wang, G.-S. Exploration of the function of ginsenoside RD attenuates lipopolysaccharide-induced lung injury: A study of network pharmacology and experimental validation. Shock 2021, 57, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Satoh, K.; Murata, P.; Makino, B.; Sakakibara, I.; Kase, Y.; Ishige, A.; Higuchi, M.; Sasaki, H. Components of Panax ginseng that improve accelerated small intestinal transit. J. Ethnopharmacol. 2003, 84, 115–119. [Google Scholar] [CrossRef]
- Yao, L.; Han, Z.; Zhao, G.; Xiao, Y.; Zhou, X.; Dai, R.; Han, M.; Wang, Z.; Xin, R.; Wang, S. Ginsenoside Rd ameliorates high fat diet-induced obesity by enhancing adaptive thermogenesis in a cAMP-dependent manner. Obesity 2020, 28, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, J.; Jeong, Y.T.; Byun, G.H.; Kim, J.H. Melanogenesis inhibition activity of floralginsenoside A from Panax ginseng berry. J. Ginseng Res. 2017, 41, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Gu, L.J.; Zhang, D.L.; Wang, Y.B.; Sung, C.K. Ginsenosides Rb₁ and Rd regulate proliferation of mature keratinocytes through induction of p63 expression in hair follicles. Phytother. Res. 2013, 27, 1095–1101. [Google Scholar] [CrossRef]
- Kim, D.Y.; Park, Y.G.; Quan, H.-Y.; Kim, S.J.; Jung, M.S.; Chung, S.H. Ginsenoside Rd stimulates the differentiation and mineralization of osteoblastic MC3T3-E1 cells by activating AMP-activated protein kinase via the BMP-2 signaling pathway. Fitoterapia 2012, 83, 215–222. [Google Scholar] [CrossRef]
- Sun, C.; Choi, I.Y.; Gonzalez, Y.I.R.; Andersen, P.; Talbot, C.C., Jr.; Iyer, S.R.; Lovering, R.M.; Wagner, K.R.; Lee, G. Duchenne muscular dystrophy hiPSC-derived myoblast drug screen identifies compounds that ameliorate disease in mdx mice. JCI Insight 2020, 5, e134287. [Google Scholar] [CrossRef]
- Kim, W.-K.; Song, S.-Y.; Oh, W.K.; Kaewsuwan, S.; Tran, T.L.; Kim, W.-S.; Sung, J.-H. Wound-healing effect of ginsenoside Rd from leaves of Panax ginseng via cyclic AMP-dependent protein kinase pathway. Eur. J. Pharmacol. 2013, 702, 285–293. [Google Scholar] [CrossRef]
- Tamura, T.; Cui, X.; Sakaguchi, N.; Akashi, M. Ginsenoside Rd prevents and rescues rat intestinal epithelial cells from irradiation-induced apoptosis. Food Chem. Toxicol. 2008, 46, 3080–3089. [Google Scholar] [CrossRef]
- Wang, W.; Wang, G.-J.; Xie, H.-T.; Sun, J.-G.; Zhao, S.; Jiang, X.-L.; Li, H.; Lv, H.; Xu, M.-J.; Wang, R. Determination of ginsenoside Rd in dog plasma by liquid chromatography–mass spectrometry after solid-phase extraction and its application in dog pharmacokinetics studies. J. Chromatogr. B 2007, 852, 8–14. [Google Scholar] [CrossRef]
- Sun, D.; Wang, B.; Shi, M.; Zhang, Y.-X.; Zhou, L.-F.; Liu, Z.-R.; Wu, Z.-L.; Jiang, W.; Han, J.-L.; Xiong, L.-Z.; et al. Pharmacokinetic, tissue distribution and excretion of ginsenoside-Rd in rodents. Phytomedicine 2012, 19, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.-Y.; Zhang, Y.-B.; Zhou, Q.-L.; Yang, Y.-F.; Yang, X.-W. Simultaneous determination of eight ginsenosides in rat plasma by liquid chromatography–electrospray ionization tandem mass spectrometry: Application to their pharmacokinetics. Molecules 2015, 20, 21597–21608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, J.-H.; Lee, J.; Choi, M.-K.; Song, I.-S. Pharmacokinetics of ginsenosides following repeated oral administration of red ginseng extract significantly differ between species of experimental animals. Arch. Pharmacal Res. 2020, 43, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Lee, E.K.; Bae, C.H.; Park, S.-D.; Shim, J.-J.; Lee, J.-L.; Yoo, H.H.; Kim, D.-H. The impact of gut microbiome on the pharmacokinetics of ginsenosides Rd and Rg3 in mice after oral administration of red ginseng. Am. J. Chin. Med. 2021, 49, 1897–1912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, R.; Xu, C.; Zhang, Y.; Tian, Q.; Wang, B.; Zhang, G.; Guan, Y.; Yan, J. Simultaneous determination of saponins and lignans in rat plasma by UPLC- MS/MS and its application to a pharmacokinetic study of Shenqi Jiangtang granule. Curr. Drug Metab. 2021, 22, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Du, L.-Y.; Jiang, T.; Wei, K.; Zhu, S.; Shen, Y.-L.; Ye, P.; Zhang, H.-E.; Chen, C.-B.; Wang, E.-P. Simultaneous quantification of four ginsenosides in rat plasma and its application to a comparative pharmacokinetic study in normal and depression rats using UHPLC-MS/MS. J. Anal. Methods Chem. 2021, 2021, 4488822. [Google Scholar] [CrossRef]
- Yang, L.; Deng, Y.; Xu, S.; Zeng, X. In vivo pharmacokinetic and metabolism studies of ginsenoside Rd. J. Chromatogr. B 2007, 854, 77–84. [Google Scholar] [CrossRef]
- Zeng, X.; Deng, Y.; Feng, Y.; Liu, Y.; Yang, L.; Huang, Y.; Sun, J.; Liang, W.; Guan, Y. Pharmacokinetics and safety of ginsenoside Rd following a single or multiple intravenous dose in healthy Chinese volunteers. J. Clin. Pharmacol. 2010, 50, 285–292. [Google Scholar] [CrossRef]
- Liu, X.; Xia, J.; Wang, L.; Song, Y.; Yang, J.; Yan, Y.; Ren, H.; Zhao, G. Efficacy and safety of ginsenoside-Rd for acute ischaemic stroke: A randomized, double-blind, placebo-controlled, phase II multicenter trial. Eur. J. Neurol. 2009, 16, 569–575. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Wen, A.; Yang, J.; Yan, Y.; Song, Y.; Ren, H.; Wu, Y.; Li, Z.; Chen, W.; et al. Ginsenoside-Rd improves outcome of acute ischaemic stroke—A randomized, double-blind, placebo-controlled, multicenter trial. Eur. J. Neurol. 2012, 19, 855–863. [Google Scholar] [CrossRef]
- Kim, J.-K.; Choi, M.S.; Jeung, W.; Ra, J.; Yoo, H.H.; Kim, D.-H. Effects of gut microbiota on the pharmacokinetics of protopanaxadiol ginsenosides Rd, Rg3, F2, and compound K in healthy volunteers treated orally with red ginseng. J. Ginseng Res. 2020, 44, 611–618. [Google Scholar] [CrossRef] [PubMed]




| Enzymes | Transformation Pathways | Optimum Conditions | Yield and Reaction Scale | Ref. | Year |
|---|---|---|---|---|---|
| Enzymatic Transformation | |||||
| Arabinofuranosidase | |||||
| α-l-arabinofuranosidase AbfA from Rhodanobacter ginsenosidimutans strain Gsoil 3054T | Rc ⤏ Rd | pH 7.5, 37 °C | / | [5] | 2012 |
| α-l-arabinofuranosidase, Abf22-3 from Leuconostoc sp. 22-3 | Rc ⤏ Rd | pH 6.0, 30 °C | 99.50% | [6] | 2013 |
| α-l-arabinofuranosidase from Caldicellulosiruptor saccharolyticus | Rc ⤏ Rd | pH 5.5, 80 °C, 227 U enzyme/mL | a molar yield of 100% | [7] | 2013 |
| α-l-arabinofuranosidase (Tt-Afs) from Thermotoga thermarum DSM5069 | Rc ⤏ Rd | pH 5.0, 85 °C | 99.40% | [8] | 2016 |
| α-l-arabinofuranosidase from Bacillus subtilis Str. 168 | Rc ⤏ Rd | pH 5.0, 40 °C | 90% | [9] | 2021 |
| Arabinopyranosidase | |||||
| α-l-Arabinopyranosidase from Blastococcus saxobsidens (AbpBs) | Rb2 ⤏ Rd | pH 7.0, 40 °C | / | [10] | 2020 |
| β-glucosidase | |||||
| β-glucosidase Tt-BGL from Thermotoga thermarum DSM 5069T | Rb1 ⤏ Rd | pH 4.8, 90 °C | 95% | [11] | 2013 |
| β-glucosidase Bgp3 from Microbacterium esteraromaticum | Rb1 ⤏ Rd ⤏ CK | pH 7.0, 40 °C | 77% | [12] | 2012 |
| glycosidase Bgp2 from Microbacterium esteraromaticum | Rb2 ⤏ Rd ⤏ 20(S)-Rg3 | pH 7.0, 40 °C | 65% | [13] | 2013 |
| β-Glucosidase Bgy2 from Lactobacillus brevis | Rb1 ⤏ Rd⤏F2 ⤏ CK | pH 7.0, 30 °C | 69%91% | [14] | 2016 |
| β-glucosidase from Aspergillus niger KCCM 11239 | Rb1 ⤏ Rd ⤏ Rg3 Rb1 ⤏ Rd ⤏ F2 | pH 4.0, 70 °C | / | [15] | 2012 |
| Pectinase | |||||
| Pectinase coupled with one-pot process | Rb1 ⤏ Rd | pH 6.0, 52.5 °C | 83.14% | [16] | 2020 |
| Microbial Transformation | |||||
| Fungal System | |||||
| Paecilomyces bainier 229-7 | Rb1 ⤏ Rd | / | 94.9% in shake flasks, 89% in 10 L fermenter | [17] | 2010 |
| Paecilomyces bainier 229-7 | Rb1 ⤏ Rd | / | 92.44% | [18] | 2012 |
| Aspergillus versicolor strain LFJ1403 | Rb1 ⤏ Rd | pH 5.0, 37 °C | 94.9% in shake flasks85% in 2 L fermenter | [19] | 2015 |
| Aspergillus niger strain TH-10a | Rb1 ⤏ Rd | pH 5.0, 32 °C | 86% | [20] | 2016 |
| Bacteria system | / | ||||
| M. trichothecenolyticum | Rb1 ⤏ Rd⤏ Rh2 | / | / | [21] | 2013 |
| Bacterial strain MAH-16T | Rb1 ⤏ Rd | pH 5.0–7.0, 20–40 °C | / | [22] | 2018 |
| Bacterial strain MAHUQ-46T | Rb1 ⤏ Rd | pH 7.5, 30 °C | / | [23] | 2021 |
| Bacterial strain FW-6T | Rb1 ⤏ Rd | / | / | [24] | 2013 |
| Bacterium G9y | Rc ⤏ Rd | pH 7.0, 45 °C | / | [25] | 2021 |
| Gut microbiota | |||||
| Gut bacteria | Rb1 ⤏Rd ⤏ F2 ⤏ CK Rb1 ⤏ G-XVII ⤏ G-LXXV ⤏ CK | / | / | [26] | 2013 |
| Leuconostoc mesenteroides DC102 | Rb1 ⤏ G-XVII and Rd ⤏ F2⤏ CK | pH 6.0–8.0, 30 °C | 99% | [27] | 2011 |
| Lactobacillus paralimentarius LH4 | Rb1 ⤏ G-XVII and Rd ⤏ F2 ⤏ CK | pH 6.0, 30 °C | 88% | [28] | 2013 |
| Probiotics | Rb1 ⤏ Rd ⤏ F2⤏ CK | / | / | [29] | 2021 |
| Lactobacillus rhamnosus GG | Rb1 ⤏ Rd | pH 6.0, 40 °C | / | [30] | 2016 |
| Food microorganisms | |||||
| Dekkera anomala YAE-1 | Rb1 ⤏ Rd | pH 5.0, 40 °C | / | [31] | 2020 |
| Anti-Cancer | ||||||
|---|---|---|---|---|---|---|
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
| Cervical cancer | Cell lines: HeLa | In vitro: IC50 = 150.5 ± 0.8 μg/mL (48 h) | Inhibited proliferation and induced cell apoptosis | Bcl-2↓, Bax↑, mitochondrial transmembrane potential↓, caspase-3↑ | [34] | 2006 |
| Glioblastoma | Cell lines: U251 | In vitro: IC50 = 88.89 μM (24 h); IC50 = 13.20 μM (28 h); IC50 = 9.55 μM (72 h) | Inhibited proliferation, promoted cell apoptosis, enhanced the expression of telomerase | caspase-3↑, Bcl-2↓, hTERT↓ | [35] | 2019 |
| Cell lines: U251, H4 (HTB148), U87 MG (HTB-14) cells, NHA | In vitro: Rd (100, 200 µM) | Reduced proliferation and migration | miR-144-5p↑ | [36] | 2020 | |
| Gastric cancer | Cell lines: SGC-7901Cell lines: MKN-45 | In vitro: IC50 = 86.96 ± 0.23 μg/mL (SGC-7901, 48 h) and 71.70 ± 2.16 μg/mL (MKN-45, 48 h) | Inhibited proliferation, induced apoptosis and cell cycle arrest at G0/G1 phase | Cyclin D1↓, caspase-3↑, caspase-9↑, Bax/Bcl-2↑ | [33] | 2020 |
| Cell lines: AGS, MCF-7 | In vitro: IC50 =131.2 μM (AGS) IC50 = 154.3 µM (MCF-7) | Inhibited proliferation | TRPM7 channel activity↓ | [40] | 2013 | |
| Liver cancer | Cell lines: HepG2 | In vitro: EC50 = 18.26 μM | Combination of CA4P and Rd inhibited proliferation and induced apoptosis | HIF-1α↓, PI3K/AKT/mTOR↓ | [38] | 2021 |
| Cell lines: HepG2 | In vitro: IC50 = 256.3 μM (24 h) and 172 μM (48 h) | Inhibited migration and invasion | MMP↓, MAPK↓ | [41] | 2012 | |
| Colorectal cancer | Cell lines: HT29 | In vitro: IC50 = 277 μg/mL (48 h) | Inhibited proliferation | caspase 3↑, stathmin 1c, PCNA↓, rho GDP dissociation inhibitor (GDI) alpha↓, reticulocalbin 1 precursor↓, nudix hydrolase NUDT5↓, microtubule-associated protein RP/EB family↓, proteasome β 6 subunit↓, tyrosine 3/tryptophan 5-monooxygenase activation protein, epsilon↓, tropomyosin 1 (α)↑, glutathione S-transferase-P1↑, annexin 5↑, Nm23 protein↑, tropomodulin 3↑, and stratifin ↑ | [37] | 2009 |
| Cell lines: HT29 and SW620 | In vitro: 0, 10, 50, 100 μM (72 h) | Inhibited metastasis | Bound to EGFR with a high binding affinity, stemness- and EMT-related genes↓ | [42] | 2019 | |
| Cell lines: HUVEC animals: LoVo xenograft BALB/C mice | In vitro: Rd (2, 10, 50 µM In vivo: SMI (10 mL/kg/day, 13 days) | Suppressed neovascularization in tumors, normalized the structure of tumor vessels, and improved the anti-tumor effect of 5-FU | / | [45] | 2019 | |
| Animals: heterozygous C57BL/6J-ApcMin/+ mice | In vivo: Rd (20 mg/kg, 8 weeks) | suppressed cancer-promoting signaling markers, reduced the size and the number of the polyps, and improved intestinal barrier | iNOS↓, STAT3/pSTAT3↓, Src/pSrc↓, reinstated mucosal architecture, improved mucosal immunity, promoted beneficial bacteria, cancer cachexia associated bacteria↓ | [48] | 2017 | |
| Breast cancer | Cell lines: HEK293, MDA-MB-231, AU565, and T47D | In vitro: Rd (100–400 µM) | Suppressed the viability of TRPM7-expressing breast cancer cells | S phase↑, G0/G1 phase↓ | [39] | 2020 |
| Cell lines: AGS, MCF-7 | In vitro: IC50 = 131.2 µM (AGS) and 154.3 µM (MCF-7) | Inhibited proliferation, induced cell apoptosis | TRPM7 channel activity↓ | [40] | 2013 | |
| Cell lines: 4T1, MDA-MB-231 | In vitro: Rd (50, 100, 150 μM, 72 h) | Suppressed cell migration and invasion | miR-18a-mediated Smad2↓ | [41] | 2016 | |
| Cell lines: HUVECs, MDA-MB-231 | In vitro: Rd (5, 10, 25, 50 µM) | Inhibited VEGF-induced migration, tube formation and proliferation of HUVECs, Inhibited proliferation and induced apoptosis | AKT/mTOR/P70S6↓ | [42] | 2017 | |
| Cell lines: MCF-7, MCF-7/ADR | In vitro: Rd (10, 100 µg/mL, 24 h) | Reversed doxorubicin resistance in MCF-7/ADR cells | MDR1 protein↓ | [44] | 2010 | |
| Lung cancer | Cell lines: A549 | In vitro: IC50 = 246.4 µM (24 h) IC50 = 149.0 µM (48 h) IC50 = 93.7 µM (72 h) | Inhibited proliferation, induced G0/G1 phase arrest, reversed cisplatin resistance | NRF2 pathway↓ | [47] | 2019 |
| Anti-diabetic | ||||||
| Diabetes | Animals: postnatal day 1 SD rats | In vivo: Rd (5, 10, 20, 50 μM) | Ameliorated the cell viability of MG-treated astrocytes | Improved insulin signaling and inhibited apoptosis | [49] | 2014 |
| Cell lines: human pancreatic islets | In vitro: Rd (0.1,1,10 μM, 72 h) | Inhibited the progress of death of cultured human pancreatic islets, no effects on glucose-induced insulin and C-peptide stimulation secretion | Apoptosis of the islet cells↓, Bax↓, Bcl2↑, and caspase-3↓ | [50] | 2019 | |
| Animals: type-2 diabetic db/db mice | In vivo: GS-E3D (100 or 250 mg/kg/d, oral, 6 weeks) | Renal protective roles | ROS↓ | [51] | 2021 | |
| Diabetic retinopathy (DR) | Cell lines: HUVEC Animals: STZ-induced diabetic mouse model | In vitro: Rd (1, 3, 10, 30 μM, 24 h) In vivo: Rd (100 mg/kg, 1 month) | Ameliorated diabetes-driven vascular damage, modulated oxidative stress and apoptosis | AMPK↑, SIRT1↑, AMPK/SIRT1 interaction↑ | [52] | 2022 |
| Anti-Inflammatory | ||||||
|---|---|---|---|---|---|---|
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
| Chronic hepatitis | Cell lines: HepG2 | In vitro: Rd (IC50 = 12.05 ± 0.82 µM) | Anti-inflammatory activity | NF-kB↓, iNOS↓, COX-2↓ | [53] | 2012 |
| Neuroinflammation | Cell lines: mouse primary neuron-glia Animals: pregnant OF1/SPF mice | In vivo: Rd (1, 10, 50 µM) | Protected dopaminergic neurons against LPS-neurotoxicity | iNOS↓, COX-2↓, iNOS↓, PGE2↓ | [54] | 2007 |
| Osteoarthritis | Cell lines: S12 | In vitro: Rd (100 μg/mL) | Exerted a protective effect against the cartilage degradation of OA | p-p38↓, MMP3↓ | [55] | 2009 |
| Gastritis | Animals: ethanol- or indomethacin-induced gastric mucosal lesions in rat model | In vivo: Rd (100 mg/kg) | Showed gastroprotective effects on ethanol- and indomethacin-induced gastric mucosal lesions | / | [56] | 2007 |
| Colitis | Animals: DSS-induced murine colitis model | In vivo: Rd (10, 20, 40 mg/kg) | Ameliorated DSS-induced colitis, inhibited inflammatory cell recruitment into colonic tissue | p62-driven mitophagy-mediated NLRP3 inflammasome↓, AMPK/ULK1↑ | [59] | 2018 |
| Animals: TNBS-induced ulcerative colitis rat model | In vivo: Rd (10, 20, 40 mg/kg/d, orally | Against TNBS-induced recurrent ulcerative colitis and increased superoxide dismutase and glutathione peroxidase activities | Inhibited neutrophil infiltration and promoted the antioxidant capacity of the damaged colonic tissue | [60] | 2012 | |
| Animals: TNBS-induced ulcerative colitis rat model | In vivo: Rd (10, 20, 40 mg/kg/d, 7 days) | Attenuated the inflammatory response to TNBS-induced relapsing colitis | MPO↓, proinflammatory cytokine TNF-α, IL-1β, and IL-6↓, p-p38↓, JNK↓ | [61] | 2012 | |
| Inflammatory bowel diseases(IBD) | Animals: indomethacin-induced IBD rat model | In vivo: Rd (10, 20, 40 mg/kg, 7 days) | Stimulated the proliferation and differentiation of endogenous intestinal stem cells in IBD model rats, improved recovery of intestinal function | Bmi, CDX-2, and Msi-1↑ | [62] | 2020 |
| Allergic rhinitis | Cell lines: RBL-2H3Animals: ovalbumin-induced AR mice model | In vivo: Rd (10 μM, 18 h) | Alleviated ovalbumin-induced allergic rhinitis in mice | IgE, IL-4, IL-5, and IL-13↓, restored the composition of gut microbiota | [63] | 2019 |
| Inflammatory | Cell lines: RAW264.7Animals: ICR mouse | In vitro: LPS (5 mg/kg) + Rd (2, 10, 50 mg/kg) | Anti-inflammatory effects | NF-kB↓, iNOS↓, COX-2↓, NO↓, PGE2↓ | [57] | 2013 |
| Cell lines: HepG2 | In vitro: Rd (IC50 = 3.47 μM) | Suppressed inflammatory responses | NF-κB↓, COX-2↓ and iNOS↓ | [58] | 2014 | |
| Animals: carrageenan-induced hind paw edema rat model | In vivo: Rd (12.5, 25, 50 mg/kg, i.m.) | Anti-inflammatory effects against carrageenan-induced edema | NF-kB↓ | [64] | 2012 | |
| Animals: carrageenan -induced rat paw edema rat model | In vivo: Rd (12.5, 25, 50 mg/kg) | Reduced the inflammatory cell infiltration into inflammatory sites, inhibited the tissue lipid peroxidation, increased the antioxidant enzyme activities, and suppressed the proinflammatory enzyme expressions | NF-κB↓, p-ERK↓, p- JNK↓ | [65] | 2013 | |
| Antioxidative | ||||||
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
| Antioxidative | Animals: senescence-accelerated mice (SAM) of 10 months | In vivo: Rd (1 or 5 mg/kg/d, 30 days) | Attenuated the oxidative damage and enhanced the antioxidative defense system | Regulated the GSH/GSSG redox status | [66] | 2004 |
| Animals: synchronized L4 larvae worms | In vivo: TG (10 μg/mL) | Has antiaging effects and only Rd prolonged the lifespan of C. elegans to levels comparable to total ginsenoside (TG) | Via lipid metabolism and activating the stress response signaling pathway | [67] | 2021 | |
| Cell lines: PC12 | In vitro: Rd (1, 10 μM) | Antioxidative properties | ROS↓, MDA↓, SOD↑, GSH-Px↑, stabilized the mitochondrial membrane potential | [68] | 2008 | |
| Cell lines: H4IIE | In vitro: Rd (1–30 μg/mL) | Antioxidative effects; increased both cellular glutathione (GSH) content and the protein level of γ-glutamylcysteine ligase heavy chain | p65↑ via NF-κB-dependent γ-glutamylcysteine ligase induction | [69] | 2007 | |
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
|---|---|---|---|---|---|---|
| Ischemic stroke | Animals: MCAO rat models | In vivo: Rd (30 mg/kg) | Reduced mtDNA and nDNA damages and had the neuroprotective function | Survival rate and neurological function↑, cell apoptosis↓, cleaved caspase-3↓, NEIL1 and NEIL3↑ | [96] | 2016 |
| Cell lines: cortical neurons cells from embryonic day 18 SD ratsAnimals: MCAO rat models | In vitro: Rd (1, 3, 10, 30, 100 μM) In vivo: 10 mg/kg | Neuroprotectant for the treatment of ischemic stroke; exerted an inhibitive effect on NMDAR-triggered currents and sequential excitotoxicity | DAPK1-mediated NR2B phosphorylation↓, calcineurin activity↓ | [97] | 2020 | |
| Cell lines: cortical neuronsAnimals: MCAO rat models | In vitro: Rd (10 μM) In vivo: Rd (50 mg/kg) | Improved the behavior score, infarct volume, and viability of the cultured neurons after ischemia | Hyperphosphorylation of NR2B subunit↓ and expression levels of NR2B subunit in cell membrane↓ | [98] | 2016 | |
| Cells: microglia from P1 newborn SD rats, BV2, MC3T3-E1 Animals: MCAO rat models | In vitro: Rd (1, 10, 50, 100 μM) In vivo: 10 mg/kg, i.p. | Improved the outcome of patients with ischemic stroke | Microglial proteasome activity and sequential inflammation↓ | [99] | 2016 | |
| Animals: MCAO rat models | In vivo: Rd (1, 10, 100 μM) In vivo: 30 mg/kg, i.p. | Attenuated the pathogenesis of cerebral ischemia-induced BBB damage, suppressed proteasome-mediated inflammation | Proteasome activity and NF-κB/MMP-9 pathway↓ | [100] | 2020 | |
| Cell lines: BV-2 Animals: MCAO rat models | In vitro: Rd (0.1, 1, 10 μM) In vivo: CPA (4.5, 9 g/kg) | Attenuated cerebral injury after stroke | NLRP3↓, OGD/R-induced BV-2 cell injury↓, Drp1-mediated mitochondrial fission↓, Drp1↓ | [101] | 2020 | |
| Cardiovascular diseases | Cell lines: A10 embryonic rat thoracic aortic, rat aorta smooth muscle cells prepared from rat thoracic aorta | In vitro: Rd (100 μM) | Had an effect on cardiovascular diseases and inhibited Ca2+ entry | Through ROCC and SOCC without effects on VDCC and Ca2+ release | [102] | 2006 |
| Cerebrovascular remodeling | Cell lines: BAVSMCs from rat basilar arteries Animals: two-kidney, two-clip (2k2c) stroke-prone hypertensive rat model | In vitro: Rd(2.5, 5, 10, 20, 40 μM, 48 h) In vivo: Rd (20 mg in 2 mL saline solution containing 20% propylene glycol/kg/d) | Attenuated basilar hypertrophic inward remodeling in 2k2c hypertensive rats without affecting systemic blood pressure; attenuated hypertensive cerebrovascular remodeling | Inhibited voltage-independent Ca2+ entry and BAVSMC proliferation, but not with VDCC-mediated Ca2+ entry | [103] | 2009 |
| Cell lines: BASMCs from rat basilar arteries | In vivo: Rd (10 μM) | Potentiated H2O2-induced cell death and cell apoptosis | Cyt c release↑, caspase-9/caspase-3↑, Bcl-2/Bax↓ | [104] | 2011 | |
| Cell lines: RAW264.7 Animals: apolipoprotein E deficient (ApoE−/−) mice | In vitro, Rd (20 μM) In vivo: Rd (20 mg/kg/d) | Prevented the development of atherosclerosis | Through voltage-independent Ca2+ channels, SR-A↓, ox-LDL↓, cholesterol ↓ | [105] | 2011 | |
| Cell lines: ventricular myocytes from the hearts of male SD rats | In vitro: Rd (IC50 = 32.4 ± 7.1 μM) | Protected the heart and inhibited ICa,L | ICa,L peak amplitude↓, the current-voltage (I-V) curve↑, changed the steady-state activation curve of ICa,L and slowed down the recovery of ICa,L from inactivation | [106] | 2015 | |
| Cardiac hypertrophy | Cells: rat neonatal cardiac myocytes (NRCMs) from 24 h old SD ratsAnimal: C57BL/6 mice | In vitro: Rd (150 µg/mL) In vivo: Rd (50 μg/kg/d, i.v., 14 days) | Improved cardiac dysfunction and remodeling induced by pressure overload | AKT↓, calcineurin A↓, ERK1/2 and TGF-β1↓ | [107] | 2019 |
| Myocardial I/R injury | Cells: neonatal rat cardiomyocytes (NRCs) Animals: MI/R injury rat model | In vitro: Rd (10 µM) In vivo: Rd (50 mg/kg) | Augmented rat cardiac function, reduced myocardial infarct size, apoptotic cell death | Left ventricular ejection fraction (LVEF)↑, ±dP/dt↑;inhibited caspase-9 and caspase-3, p-AKT and GSK-3β↑, and Bcl-2/Bax ratio↑ | [108] | 2013 |
| Cells: neonatal rat cardiomyocyte (NRCs) Animals: MI/R injury rat model | In vivo: Rd (50 mg/kg) | Improved cardiac function and attenuated myocardial infarction | Serum creatine kinase, LDH and cTnI↓, Nrf2, HO-1 and NQO1↑ | [109] | 2015 | |
| Vascular endothelial injury | Cell lines: HUVECs, THP-1Animal: nicotine-administered SD rat model | In vitro: Rd (30 μM, 24 h) In vivo: Rd (25, 50 mg/kg/d, 4 weeks) | Prevented nicotine-induced cardiovascular diseases | Vascular endothelial NO signaling↑, platelet aggregation and vasoconstriction↓, endothelial cell adhesion↓ | [110] | 2020 |
| Multiple sclerosis (MS) | Animals: MOG35–55 induced EAE mouse model | In vivo: Rd (40 mg/kg/d, 35 days) | Ameliorated clinical severity and improved histopathology, reduced BBB dysfunction | IFN-γ↓, IL-4↑; BDNF and NGF↑ | [111] | 2014 |
| Cells: Mouse bone marrow stem cellsAnimals: EAE C57BL/6 mice | In vivo: 50 μM | Ameliorated the severity of EAE and attenuated the characteristic signs of disease; had modulation potential on gut microbiota in EAE mice | IL-6 and IL-17↓, TGF-β and IL-10↑, modulated Treg/Th17 imbalance | [112] | 2020 | |
| Guillain–Barré syndrome (GBS) | Cells: mouse bone marrow stem cells Animals: P0180–199 induced EAN mouse model | In vitro: Rd(10, 30, 50 μM) In vivo: Rd (20, 50, 100 mg/kg, 30 days) | Preventive function on GBS, attenuated experimental autoimmune neuritis in mice | Modulated monocytes infiltration and macrophage polarization, regulated monocyte phenotype | [113] | 2021 |
| Immunosuppressive | Cells: mouse spleen T lymphocytesAnimals: allo-skin transplantation rat model | In vivo: Rd (25 mg/kg) | Antagonized transplant rejection | Th1 cytokines IL-2↓, IFN-γ↓, TNF-α↓, IL-12↓, Th2 cytokine IL-10↑ | [114] | 2012 |
| Immunoadjuvant | Animals: OVA-immunized mouse model | In vivo: Rd (25 μg, 2 weeks) | Had immunological adjuvant activity, and elicited a Th1 and Th2 immune response, enhanced the Con A-, LPS-, and OVA-induced splenocyte proliferation | Regulated production and gene expression of Th1 cytokines and Th2 cytokines | [115] | 2007 |
| Strains: C. albicans strains Animals: vaccinated BALB/c mice | In vitro: Rd (1 mg/mL) In vivo: Rd (1 mg/mL, i.p., 10 days) | Protected mice against disseminated candidiasis and enhanced Th1 immunity | Elicited higher titers of Th1 type antibody and a Th1-dominant immune response | [116] | 2013 | |
| Anaphylactoid reactions | Cells: RBL-2H3 MCs, mouse peritoneal mast cells (MPMC) isolated from mouse, LAD2 cells Animals: ICR male mice (18–22 g) | In vitro: Rd (0.11, 0.21, 0.42 mM) In vivo: Rd (10, 20, 40 mg/kg) | Potential allergens, induced the release of mediators associated with anaphylactoid reactions | β-hexosaminidase↑, histamine↑, translocation of phosphatidylserine↑, Ca2+↑ | [117] | 2017 |
| Disease Type | Cell Lines/Animal | Effective Concentration/Dose | Effects | Mechanisms of Action | Refs. | Year |
|---|---|---|---|---|---|---|
| Renal injury | Animals: cisplatin-induced acute renal failure rat model | In vivo: Rd (1, 5 mg/kg/d, 30 days) | Decreased the severity of renal injury induced by cisplatin | MDA↓, blood urea nitrogen↓, Cr↓, urinary excretion of glucose↓ | [118] | 2000 |
| Cell lines: LLC-PK1 cells cultured with cisplatin Animals: cisplatin-induced acute renal failure rat model | In vitro: Rd (125 μM) In vivo: Rd (1, 5 mg/kg/d, 30 days) | Ameliorated cisplatin-induced renal injury, caused restoration of the renal function | DNA fragmentation↓, apoptosis↓, urea nitrogen and creatinine↓ | [119] | 2001 | |
| Cell lines: mouse polarized macrophagesAnimals: renal IRI mouse model | In vitro: Rd (10, 20, 50, 100 μg/mL) In vivo: Rd (10, 20, 50, 100 mg/kg) | Alleviated mouse acute renal ischemia/reperfusion injury | M1 macrophage polarization↓ | [120] | 2016 | |
| Acute lung injury (ALI) | Animals: LPS-induced ALI mouse model | In vivo: Rd (25, 50 mg/kg) | Protected mice against LPS-induced ALI; improved survival in endotoxemic mice | PI3K/AKT↓ | [121] | 2021 |
| Small intestinal transport | Animals: carbachol/BaCl2-induced accelerated small intestinal transit mouse model | In vivo: Rd (0.4, 1.0, 2.0 mg/kg) | Ameliorative effects on the carbachol-induced accelerated small intestinal transport | Intestinal motility↓, cholinergic nervous system↓ | [122] | 2003 |
| Anti-obesity | Animal: high-fat diet-induced obese mouse model | In vivo: Rd (15 mg/kg/d, 23 days) | Ameliorated obesity and insulin resistance | Cyclic adenosine monophosphate (cAMP)↑ | [123] | 2020 |
| Whitening activity | Cell lines: Melan-a cellsAnimal: zebrafish | In vitro: Rd (10, 20 μM) Re (20, 40, 80 μM) FGA (20, 40, 80, 160 μM) In vivo: FGA (80, 160 μM) | Inhibited melanin biosynthesis | AKT↑, ERK↑ | [124] | 2017 |
| Anti-alopecia | Cells: HFsAnimals: shaved skin B57CL/6 mouse model | In vivo: Rd and Rb1 (300 mg/kg/d, 35 days) | Promoted hair growth | p63 expression↑ in hair follicles | [125] | 2012 |
| Anti-osteoporotic | Cell lines: MC3T3-E1 | In vitro: Rd (10, 20, 40 μM) | Stimulated osteoblastic differentiation and mineralization | AMPK/BMP-2/Smad signaling pathways↑ | [126] | 2012 |
| Duchenne muscular dystrophy (DMD) | Cells: D2325 fibroblasts from a DMD patient Animals: mdx5cv mice | In vitro: Rd (5 μM) In vivo: Rd (10 mg/kg) | Ameliorated some of the skeletal muscle phenotypes caused by dystrophin deficiency | FLT3 signaling↑ | [127] | 2020 |
| Wound healing effects | Cell lines: KPCs, HDFsAnimal: hairless wound mice model | In vitro: Rd (0.1, 1, 10 μM) In vivo: Rd (10 μM, every 2 days, 10 days) | Promoted skin regeneration | Collagen type 1↑, matrix metalloproteinase-1 (MMP-1) ↓, cAMP-dependent protein kinase pathway↑ | [128] | 2013 |
| Irradiation-induced damage | Cell lines: rat intestinal epithelial IEC-6 cells | In vitro: Rd (2.5, 5, 10, 20, 40 μM, 24 h) | Protected and rescued rat intestinal epithelial cells from irradiation-induced apoptosis | Bax/Bcl-xL↓, Cyt c↓, cleaved-caspase-3↓, PI3K/AKT↑, MEK↓, mitochondria/caspase pathway↓ | [129] | 2008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Wang, L.; Fan, D. Insights into Recent Studies on Biotransformation and Pharmacological Activities of Ginsenoside Rd. Biomolecules 2022, 12, 512. https://doi.org/10.3390/biom12040512
Song X, Wang L, Fan D. Insights into Recent Studies on Biotransformation and Pharmacological Activities of Ginsenoside Rd. Biomolecules. 2022; 12(4):512. https://doi.org/10.3390/biom12040512
Chicago/Turabian StyleSong, Xiaoping, Lina Wang, and Daidi Fan. 2022. "Insights into Recent Studies on Biotransformation and Pharmacological Activities of Ginsenoside Rd" Biomolecules 12, no. 4: 512. https://doi.org/10.3390/biom12040512