
Exploring the Distinct Distribution of Archaeal Communities in Sites Contaminated with Explosives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Analytical Techniques
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Sequence and Statistical Analyses
2.5. Archaeal Communities Predictive Metabolic Profiling
3. Results
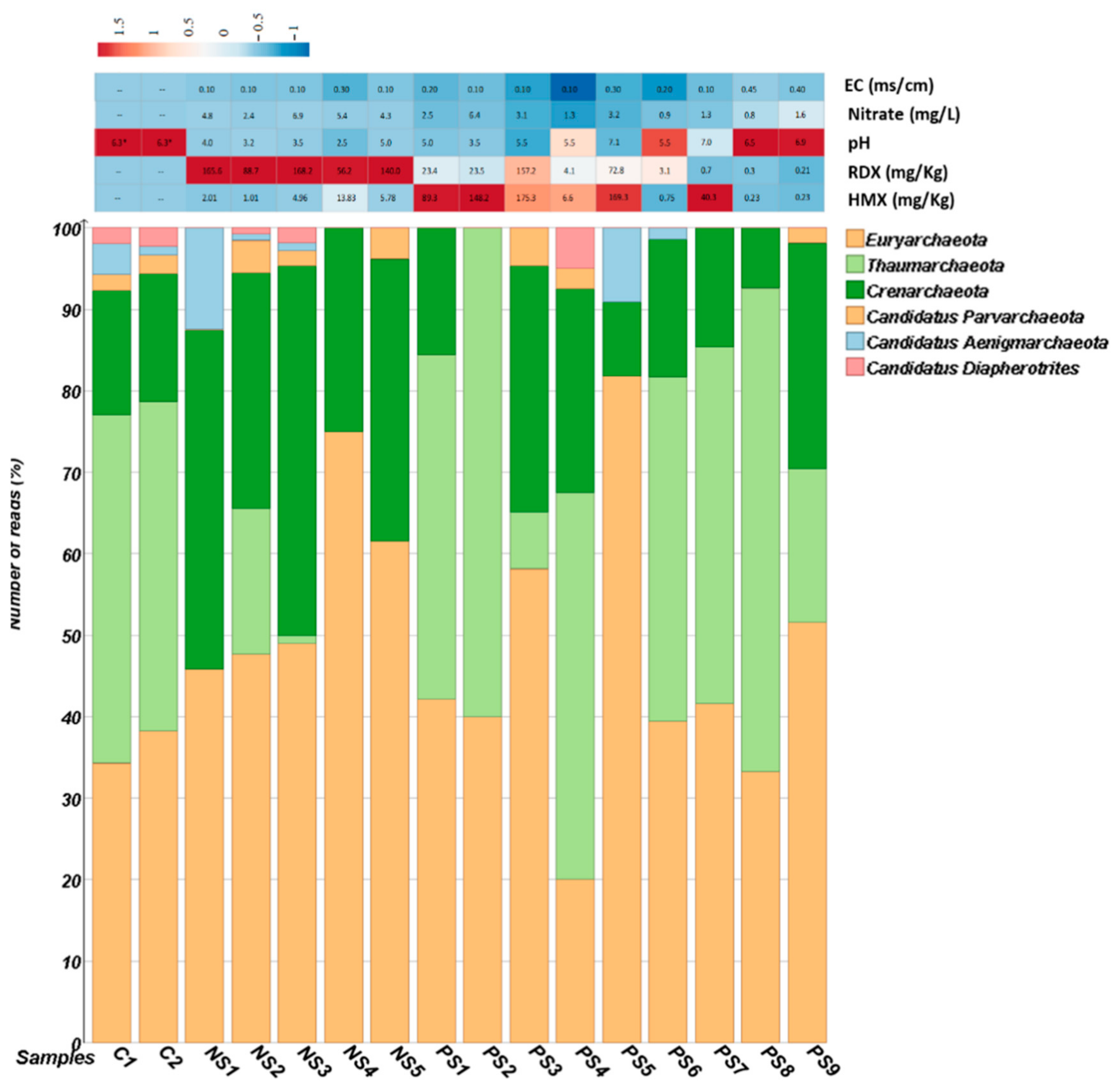
3.1. Physicochemical Analysis of Soil
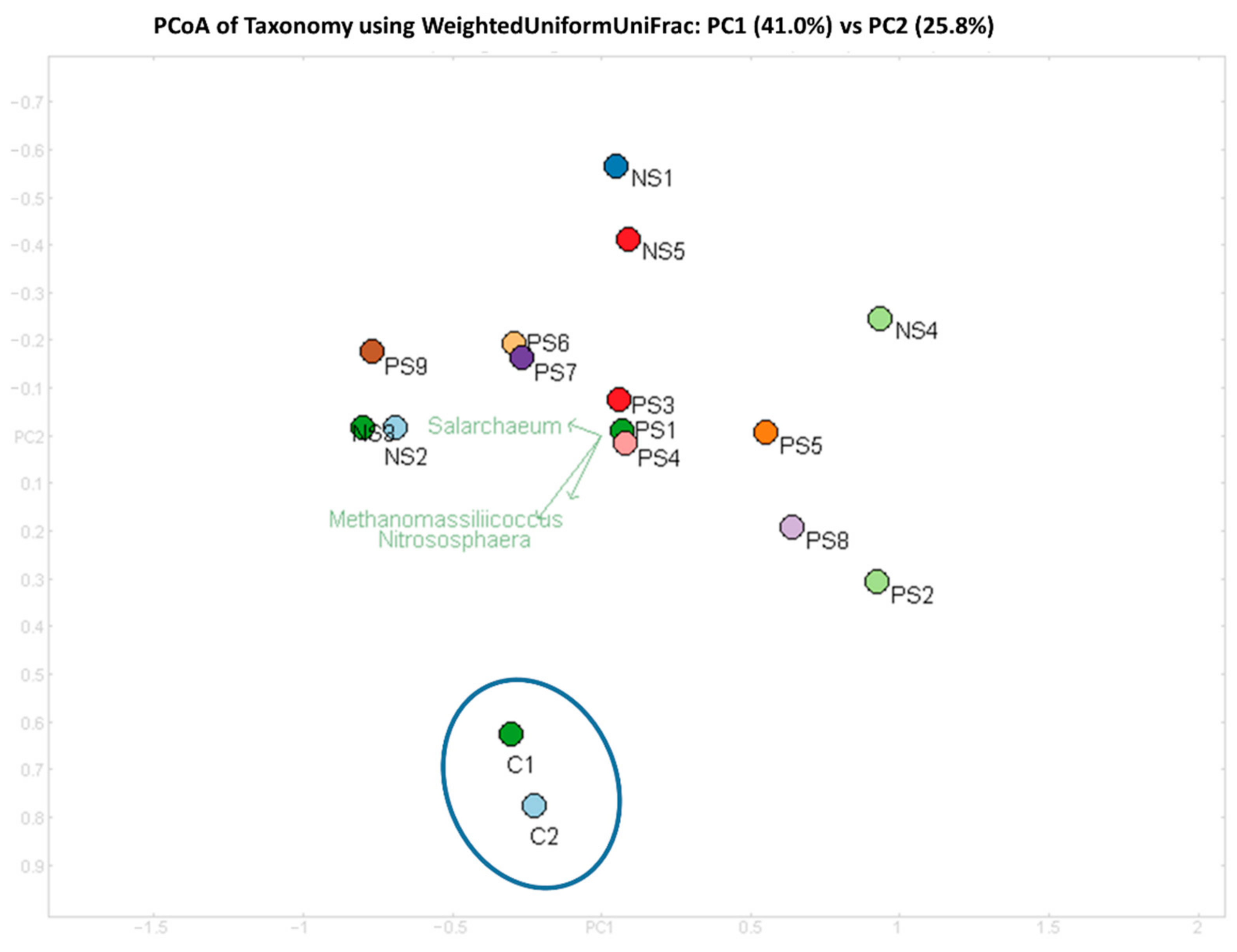
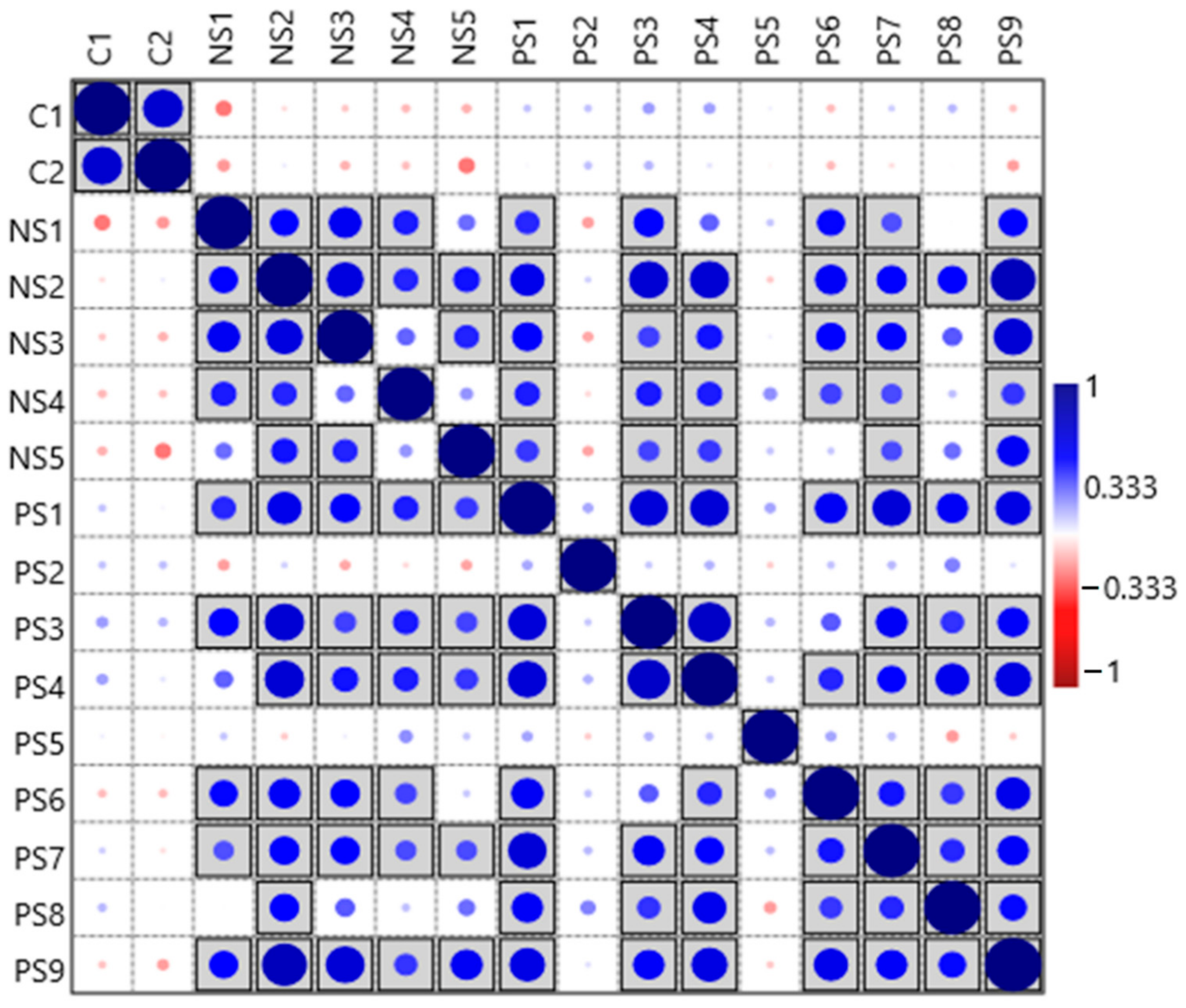
3.2. Diversity of Archaeal Community across SITE1 and SITE2
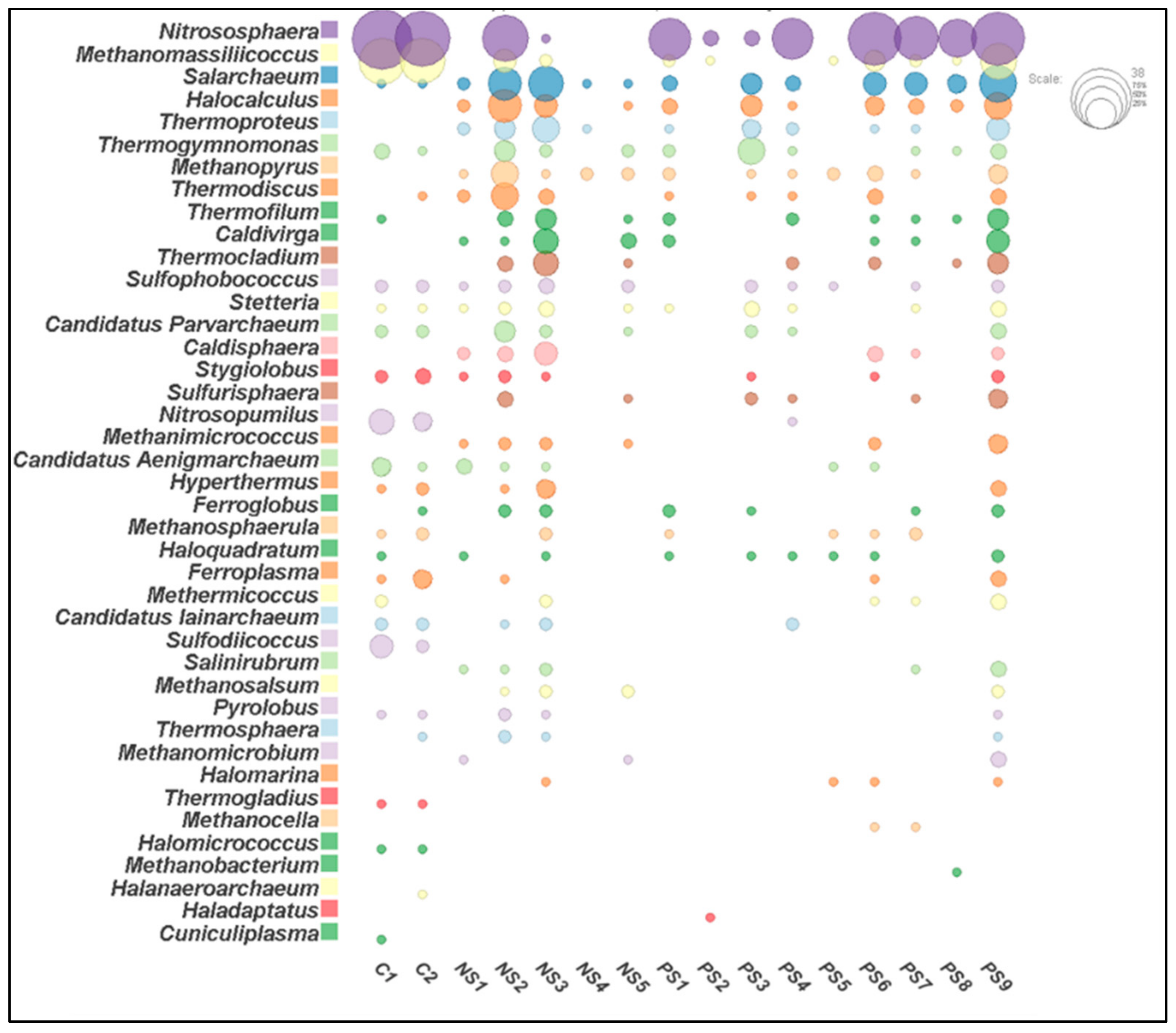
3.3. Archaeal Community Structure
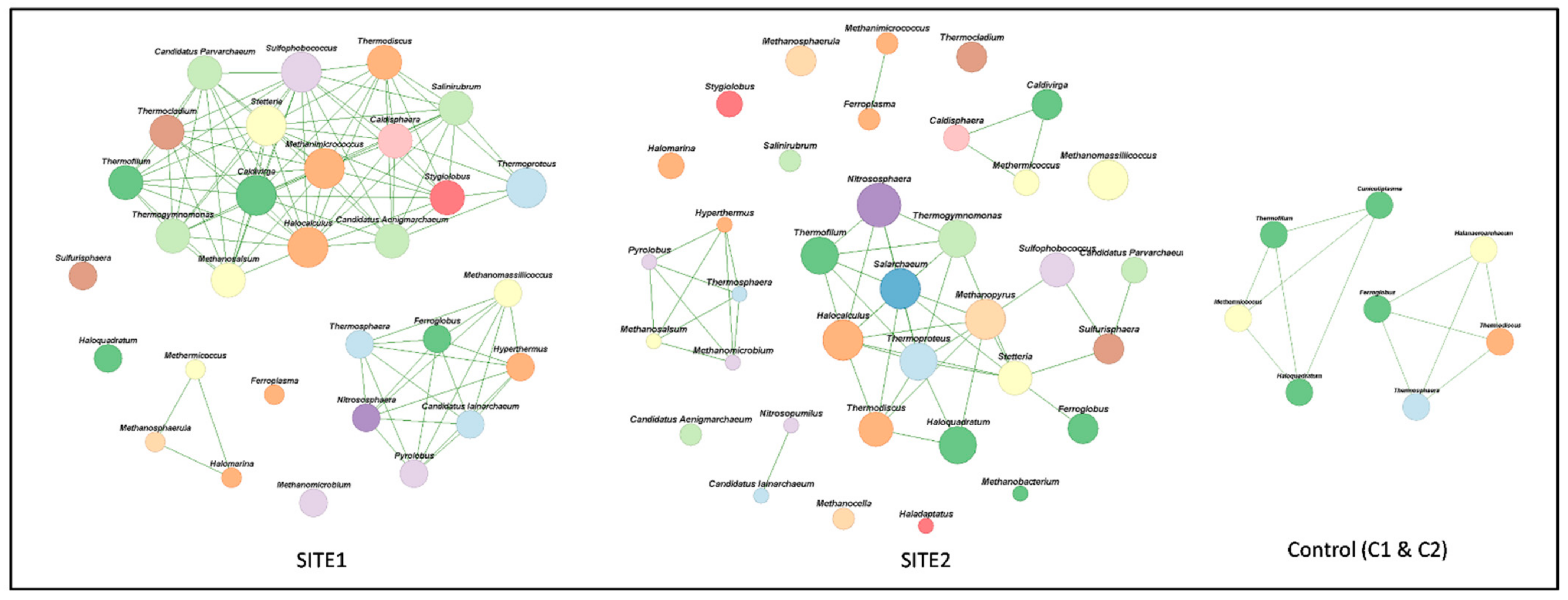
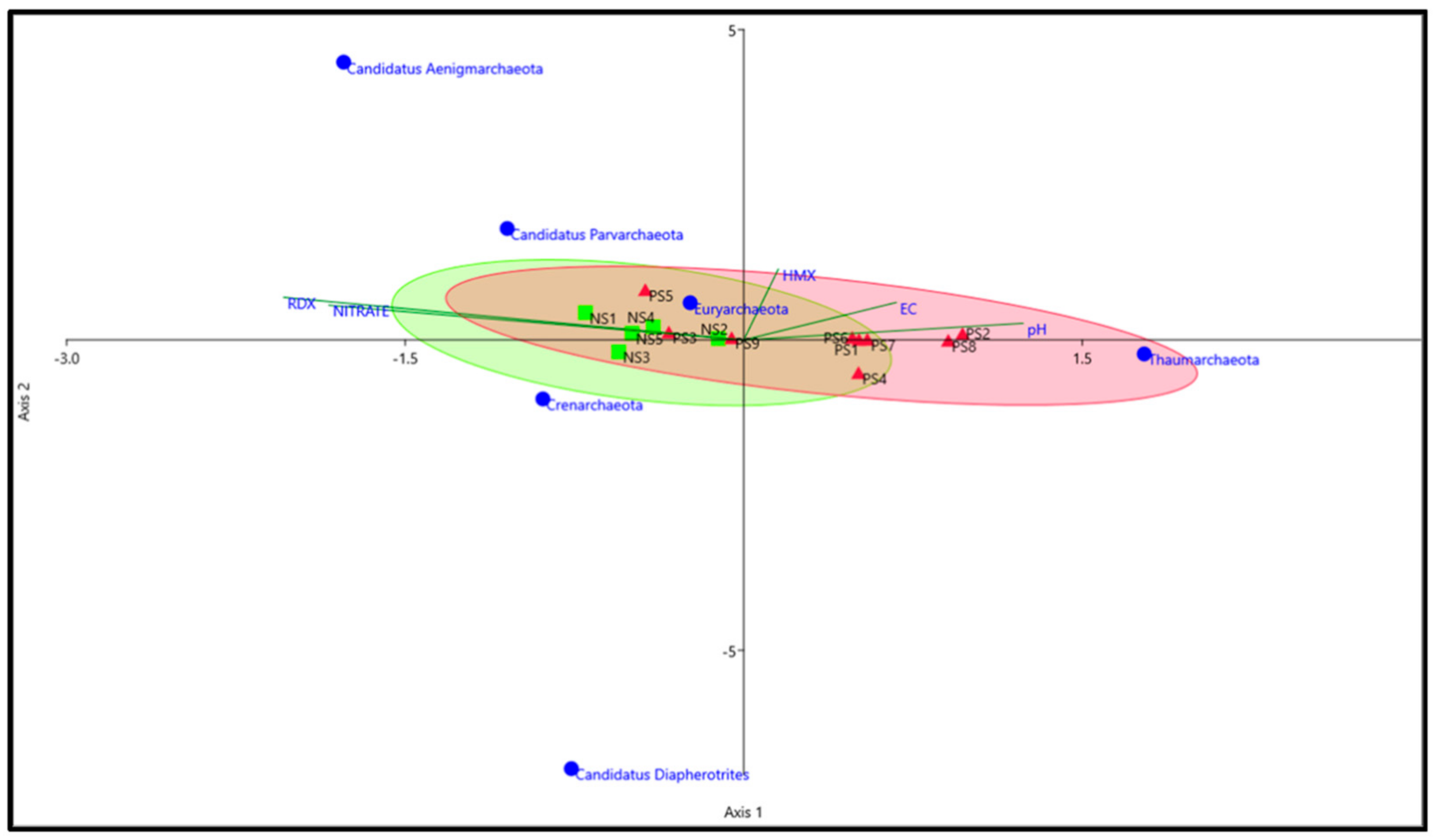
3.4. Archaeal Networks
3.5. Comparison of the Archaeal Community in Explosives Contaminated and Agricultural Soil
3.6. Predictive Archaeal Metabolic Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Offre, P.; Spang, A.; Schleper, C. Archaea in biogeochemical cycles. Annu. Rev. Microbiol. 2013, 67, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, M.; Montonen, L.; Münster, U.; Jurgens, G. Diversity and function of archaea in freshwater habitats. Curr. Trends Microbiol. 2008, 4, 61–89. [Google Scholar]
- Timonen, S.; Bomberg, M. Archaea in dry soil environments. Phytochem. Rev. 2009, 8, 505–518. [Google Scholar] [CrossRef]
- Schleper, C.; Nicol, G.W. Ammonia-oxidising archaea—Physiology, ecology and evolution. Adv. Microb. Physiol. 2010, 57, 1–41. [Google Scholar] [CrossRef]
- Chen, S.; Wang, P.; Liu, H.; Xie, W.; Wan, X.S.; Kao, S.J.; Phelps, T.J.; Zhang, C. Population dynamics of methanogens and methanotrophs along the salinity gradient in Pearl River Estuary: Implications for methane metabolism. Appl. Microbiol. Biotechnol. 2020, 104, 1331–1346. [Google Scholar] [CrossRef]
- Bertrand, J.C.; Almallah, M.; Acquaviva, M.; Mille, G. Biodegradation of hydrocarbons by an extremely halophilic archaeabacterium. Lett. Appl. Microbiol. 1990, 11, 260–263. [Google Scholar] [CrossRef]
- Mukherjee, A.; Chattopadhyay, D. Exploring environmental systems and processes through next-generation sequencing technologies: Insights into microbial response to petroleum contamination in key environments. Nucleus 2017, 60, 175–186. [Google Scholar] [CrossRef]
- Krzmarzick, M.J.; Taylor, D.K.; Fu, X.; McCutchan, A.L. Diversity and Niche of Archaea in Bioremediation. Archaea 2018, 2018, 3194108. [Google Scholar] [CrossRef] [Green Version]
- Kitts, C.L.; Cunningham, D.P.; Unkefer, P.J. Isolation of three hexahydro-1,3,5-trinitro-1,3,5-triazine-degrading species of the family Enterobacteriaceae from nitramine explosive-contaminated soil. Appl. Environ. Microbiol. 1994, 60, 4608–4711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichtel, J. Distribution and Fate of Military Explosives and Propellants in Soil: A Review. Appl. Environ. Soil Sci. 2012, 2012, 617236. [Google Scholar] [CrossRef] [Green Version]
- Juhasz, A.L.; Naidu, R. Explosives: Fate, dynamics, and ecological impact in terrestrial and marine environments. Rev. Environ. Contam. Toxicol. 2007, 191, 163–215. [Google Scholar] [CrossRef]
- EPA 505-F-14-008. Hexahydro-1,3,5-trinitro-1,3,5 triazine (RDX) 2014, (January). Technical fact sheet-RDX. United States Environmental Protection Agency. Available online: https://www.epa.gov/sites/default/files/2014-03/documents/ffrrofactsheet_contaminant_rdx_january2014_final.pdf (accessed on 11 January 2022).
- Ramakrishnan, B.; Lueders, T.; Dunfield, P.F.; Conrad, R.; Friedrich, M.W. Archaeal community structures in rice soils from different geographical regions before and after initiation of methane production. FEMS Microbiol. Ecol. 2001, 37, 175–186. [Google Scholar] [CrossRef]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Binks, P.R.; Nicklin, S.; Brouce, N.C. Degradation of hexahydro-I3, S-binitro-l,3, S-biazine (ROX) by Stenotrophomonas maltophilia PBI. Appl. Environ. Microbiol. 1995, 61, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, B.; Halasz, A.; Thiboutot, S.; Ampleman, G.; Hawari, J. Chemotaxis-mediated biodegradation of cyclic nitramine explosives RDX, HMX, and CL-20 by Clostridium sp. EDB2. Biochem. Biophys. Res. Commun. 2004, 316, 816–821. [Google Scholar] [CrossRef]
- Kanekar, P.; Dautpure, P.; Sarnaik, S. Biodegradation of nitro-explosives. Indian J. Exp. Biol. 2003, 41, 991–1001. [Google Scholar] [PubMed]
- Juck, D.; Driscoll, B.T.; Charles, T.C.; Greer, C.W. Effect of experimental contamination with the explosive hexahydro-1,3,5-trinitro-1,3,5-triazine on soil bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 255–262. [Google Scholar] [CrossRef]
- Jayamani, I.; Manzella, M.P.; Cupples, A.M. RDX degradation potential in soils previously unexposed to RDX and the identification of RDX-degrading species in one agricultural soil using stable isotope probing. Water. Air. Soil Pollut. 2013, 224, 1745. [Google Scholar] [CrossRef]
- Jayamani, I.; Cupples, A.M. Stable isotope probing reveals the importance of Comamonas and Pseudomonadaceae in RDX degradation in samples from a Navy detonation site. Environ. Sci. Pollut. Res. Int. 2015, 22, 10340–10350. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Boukhalfa, H.; Marina, O.; Ware, D.S.; Goering, T.J.; Sun, F.; Daligault, H.E.; Lo, C.C.; Vuyisich, M.; Starkenburg, S.R. Biostimulation and microbial community profiling reveal insights on RDX transformation in groundwater. Microbiol. Open 2016, 6, 423. [Google Scholar] [CrossRef] [PubMed]
- Lizbeth, D.S.; Natasha, D.R.; Katia, L.P.; Janet, K.H.; Zohre, K.; Arturo, M.D.; Konstantinos, T.K. Microbial Diversity in a Military Impacted Lagoon (Vieques, Puerto Rico) as Revealed by Metagenomics. bioRxiv 2018, 389379. [Google Scholar] [CrossRef]
- Limane, B.; Muter, O.; Juhanson, J.; Truu, M.; Truu, J.; Nolvak, H. Characterization of microbial community structure after application of different bioremediation approaches in tnt contaminated soil. In Proceedings of the 8th International Conference Environmental Engineering, Vilnius, Lithuania, 19–20 May 2011; pp. 188–194. [Google Scholar]
- Pal, Y.; Mayilraj, S.; Paul, M.; Schumann, P.; Krishnamurthi, S. Indiicoccus explosivorum gen. nov., sp. nov., isolated from an explosives waste contaminated site. Int. J. Syst. Evol. Microbiol. 2019, 69, 2555–2564. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bier, R.; Zgleszewski, L.; Peipoch, M.; Omondi, E.; Mukherjee, A.; Chen, F.; Zhang, C.; Kan, J. Distinct Distribution of Archaea from Soil to Freshwater to Estuary: Implications of Archaeal Composition and Function in Different Environments. Front. Microbiol. 2020, 11, 576661. [Google Scholar] [CrossRef] [PubMed]
- Baruah, T.C.; Barthakur, H.P. A Text Book of Soil Analysis; Vikas Publishing House Pvt Ltd.: New Delhi, India, 1997. [Google Scholar]
- AOAC; Cunniff, P. Official Methods of Analysis of AOAC International, 16th ed.; 4th Revision; AOAC: Rockville, MD, USA, 1998; Volume 1. [Google Scholar]
- US EPA. Method 8330, Nitroaromatics and Nitramines by High Performance Liquid Chromatography (HPLC) 2012, pp. 8330–8330. Available online: www.epa.gov/sw-846/pdfs/8330.pdf (accessed on 2 January 2022).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Wang, Q.; Cole, J.R.; Rosen, G.L. Using the RDP classifier to predict taxonomic novelty and reduce the search space for finding novel organisms. PLoS ONE 2012, 7, e32491. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.J.; Tappu, R. MEGAN Community Edition—Interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotech. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.E.; McClay, K.; Hawari, J.; Paquet, L.; Malone, T.E.; Fox, B.G.; Steffan, R.J. Transformation of RDX and other energetic compounds by xenobiotic reductases XenA and XenB. Appl. Microbiol. Biotechnol. 2009, 84, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, M.E.; Perreault, N.; Hawari, J. Microaerophilic Degradation of Hexahydro-1,3,5-Trinitro-1,3,5-Triazine (RDX) by Three Rhodococcus Strains. Lett. Appl. Microbiol. 2010, 51, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cupples, A.M. RDX degrading microbial communities and the prediction of microorganisms responsible for RDX bioremediation. Int. Biodeterior. Biodegrad. 2013, 85, 260–270. [Google Scholar] [CrossRef]
- Jayamani, I.; Cupples, A.M. A comparative study of microbial communities in four soil slurries capable of RDX degradation using illumina sequencing. Biodegradation 2015, 26, 247–257. [Google Scholar] [CrossRef]
- Eberly, J.O.; Indest, K.J.; Hancock, D.E.; Jung, C.M.; Crocker, F.H. Metagenomic analysis of denitrifying wastewater enrichment cultures able to transform the explosive, 3-nitro-1,2,4-triazol-5-one (NTO). J. Ind. Microbiol. Biotechnol. 2016, 43, 795–805. [Google Scholar] [CrossRef]
- Fuller, M.E.; Koster van Groos, P.G.; Jarrett, M.; Kucharzyk, K.H.; Minard-Smith, A.; Heraty, L.J.; Sturchio, N.C. Application of a multiple lines of evidence approach to document natural attenuation of hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) in groundwater. Chemosphere 2020, 250, 126210. [Google Scholar] [CrossRef] [PubMed]
- US EPA. IRIS Toxicological Review of Hexahydro-1,3,5-Trinitro-1,3,5-Triazine (RDX); (Public Comment Draft); U.S. Environmental Protection Agency: Washington, DC, USA, 2015; EPA/635/R-15/034a. [Google Scholar]
- Pennington, J.C.; Bowen, R.; Brannon, J.M.; Zakikhani, M.; Harrelson, D.W.; Gunnison, D.; Mahannah, J.; Clarke, J.; Jenkins, T.F.; Gnewuch, S. Draft Protocol for Evaluating, Selecting, and Implementing Monitored Natural Attenuation at Explosives-Contaminated Sites; Tech. Rep. EL-99-10 U.S. Army Corps Eng.; Engineer Research and Development Center: Vicksburg, MS, USA, 1999. [Google Scholar]
- Glover, D.J.; Hoffsommer, J.C. Photolysis of RDX in Aqueous Solution with and Without Ozone; Rep. No. NSWC/WOL-TR-78-175 (AD-A080195); Naval Surface Warfare Center-White Oak: Silver Spring, MD, USA, 1979. [Google Scholar]
- Balakrishnan, V.K.; Halasz, A.; Hawari, J. Alkaline hydrolysis of the cyclic nitramine explosives RDX, HMX, and CL-20: New insights into degradation pathways obtained by the observation of novel intermediates. Environ. Sci. Technol. 2003, 37, 1838–1843. [Google Scholar] [CrossRef] [PubMed]
- Kalderis, D.; Juhasz, A.L.; Boopathy, R.; Comfort, S. Soils contaminated with explosives: Environmental fate and evaluation of state-of-the-art remediation processes (IUPAC technical report). Pure Appl. Chem. 2011, 83, 1407–1484. [Google Scholar] [CrossRef]
- Meyers, S.K.; Deng, S.; Basta, N.T.; Clarkson, W.W.; Wilber, G.G. Long-term explosive contamination in soil: Effects on soil microbial community and bioremediation. Soil Sediment Contam. 2007, 16, 61–77. [Google Scholar] [CrossRef]
- Mayer, F.; Müller, V. Adaptations of anaerobic archaea to life under extreme energy limitation. FEMS Microbiol. Rev. 2014, 38, 449–472. [Google Scholar] [CrossRef] [Green Version]
- Abadin, H.; Smith, C.; Ingerman, L.; Llados, F.T.; Barber, L.E.; Plewak, D.; Diamond, G.L. Toxicological Profile RDX; Agency for Toxic Substances and Disease Registry (US): Atlanta, GA, USA, 2012. [Google Scholar]
- Biller, S.J.; Mosier, A.C.; Wells, G.F.; Francis, C.A. Global biodiversity of aquatic ammonia-oxidizing archaea is partitioned by habitat. Front. Microbiol. 2012, 3, 252. [Google Scholar] [CrossRef] [Green Version]
- Reichenberger, E.R.; Rosen, G.; Hershberg, U.; Hershberg, R. Prokaryotic nucleotide composition is shaped by both phylogeny and the environment. Genome Biol. Evol. 2015, 7, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.J.E.; Minh, B.Q.; Urich, T.; Von Haeseler, A.; Schleper, C. Unifying the global phylogeny and environmental distribution of ammonia-oxidising archaea based on amoA genes. Nat. Commun. 2018, 9, 1517. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Mazel, F.; Doebeli, M.; Parfrey, L.W. A census-based estimate of Earth’s bacterial and archaeal diversity. PLoS Biol. 2019, 17, e3000106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.X.; Rao, Y.Z.; Qi, Y.L.; Qu, Y.N.; Chen, Y.T.; Jiao, J.Y.; Shu, W.S.; Jiang, H.; Hedlund, B.P.; Hua, Z.S.; et al. Deciphering Symbiotic Interactions of ”Candidatus Aenigmarchaeota” with Inferred Horizontal Gene Transfers and Co-occurrence Networks. mSystems 2021, 6, e0060621. [Google Scholar] [CrossRef]
- Porat, I.; Vishnivetskaya, T.A.; Mosher, J.J.; Brandt, C.C.; Yang, Z.K.; Brooks, S.C.; Liang, L.; Drake, M.M.; Podar, M.; Brown, S.D.; et al. Characterization of archaeal community in contaminated and uncontaminated surface stream sediments. Microb. Ecol. 2010, 60, 784–795. [Google Scholar] [CrossRef] [Green Version]
- Boopathy, R.; Kulpa, C.F. Biotransformation of 2,4,6-trinitrotoluene (TNT) by a Methanococcus sp. (strain B) isolated from a lake sediment. Can. J. Microbiol. 1994, 40, 273–278. [Google Scholar] [CrossRef]
- Pester, M.; Schleper, C.; Wagner, M. The Thaumarchaeota: An emerging view of their phylogeny and ecophysiology. Curr. Opin. Microbiol. 2011, 14, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Gubry-Rangin, C.; Kratsch, C.; Williams, T.A.; McHardy, A.C.; Embley, T.M.; Prosser, J.I.; Macqueen, D.J. Coupling of diversification and pH adaptation during the evolution of terrestrial Thaumarchaeota. Proc. Natl. Acad. Sci. USA 2015, 112, 9370–9375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.; Lee, J.; Park, J. Microbial degradation and toxicity of hexahydro-1,3,5-trinitro-1,3,5-triazine. J. Microbiol. Biotechnol. 2012, 22, 1311–1323. [Google Scholar] [CrossRef] [Green Version]
- Das, R.; Kazy, S.K. Microbial diversity, community composition and metabolic potential in hydrocarbon contaminated oily sludge: Prospects for in situ bioremediation. Environ. Sci. Pollut. Res. Int. 2014, 21, 7369–7389. [Google Scholar] [CrossRef]
- Men, Y.; Han, P.; Helbling, D.E.; Jehmlich, N.; Herbold, C.; Gulde, R.; Onnis-Hayden, A.; Gu, A.Z.; Johnson, D.R.; Wagner, M.; et al. Biotransformation of two pharmaceuticals by the ammonia-oxidizing archaeon Nitrososphaeria gargensis. Environ. Sci. Technol. 2016, 50, 4682–4692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebron, A.; Berthe, T.; Garnier, J. Nitrification and nitrifying bacteria in the Lower Seine River and Estuary (France). Appl. Env. Microbiol. 2003, 69, 7091–7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimane, Y.; Hatada, Y.; Minegishi, H.; Echigo, A.; Nagaoka, S.; Miyazaki, M.; Ohta, Y.; Maruyama, T.; Usami, R.; Grant, W.D.; et al. Salarchaeum japonicum gen. nov., sp. nov., an aerobic, extremely halophilic member of the Archaea isolated from commercial salt. Int. J. Syst. Evol. Microbiol. 2011, 61, 2266–2270. [Google Scholar] [CrossRef] [PubMed]
- Anton, B.P.; DasSarma, P.; Martinez, F.L.; DasSarma, S.L.; Al Madadha, M.; Roberts, R.J.; DasSarma, S. Genome Sequence of Salarchaeum sp. Strain JOR-1, an Extremely Halophilic Archaeon from the Dead Sea. Microbiol. Resour. Announc. 2020, 9, e01505-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapilatu, Y.H.; Grossi, V.; Acquaviva, M.; Militon, C.; Bertrand, J.C.; Cuny, P. Isolation of hydrocarbon-degrading extremely halophilic Archaea from an uncontaminated hypersaline pond (Camargue, France). Extremophiles 2010, 14, 225–231. [Google Scholar] [CrossRef]
- An, B.A.; Shen, Y.; Voordouw, J.; Voordouw, G. Halophilic methylotrophic methanogens may contribute to the high ammonium concentrations found in shale oil and shale gas reservoirs. Front. Energy Res. 2019, 7, 23. [Google Scholar] [CrossRef]
- Bae, H.S.; Morrison, E.; Chanton, J.P.; Ogram, A. Methanogens are major contributors to nitrogen fixation in soils of the Florida Everglades. Appl. Environ. Microbiol. 2018, 84, e02222-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, H.L.; De Lorme, M.; Chaney, R.L.; Craig, A.M. Ovine ruminal microbes are capable of biotransforming hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX). Microb. Ecol. 2011, 62, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.E.L.; Henneberger, R.; Huber, H.; Moissl-Eichinger, C. Microbial syntrophy: Interaction for the common good. FEMS Microbiol. Rev. 2013, 37, 384–406. [Google Scholar] [CrossRef]
- Wegener, G.; Krukenberg, V.; Riedel, D.; Tegetmeyer, H.E.; Boetius, A. Intercellular wiring enables electron transfer between methanotrophic archaea and bacteria. Nature 2015, 526, 587–590. [Google Scholar] [CrossRef]
- Moissl-Eichinger, C.; Pausan, M.; Taffner, J.; Berg, G.; Bang, C.; Schmitz, R.A. Archaea are interactive components of complex microbiomes. Trends Microbiol. 2018, 26, 70–85. [Google Scholar] [CrossRef]
- Itoh, T.; Suzuki, K.; Nakase, T. Thermocladium modestius gen. nov., sp. nov., a new genus of rod-shaped, extremely thermophilic Crenarchaeote. Int. J. Syst. Bacteriol. 1998, 48, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef]
- Cuadros-Orellana, S.; Pohlschröder, M.; Grossman, J.; Durrant, L.R. Biodegradation of aromatic compounds by a halophilic archaeon isolated from the dead sea. Chem. Eng. Trans. 2012, 27, 13–18. [Google Scholar] [CrossRef]
- Enzmann, F.; Mayer, F.; Rother, M.; Holtmann, D. Methanogens: Biochemical background and biotechnological applications. AMB Express 2018, 8, 1. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, Y.; Mayilraj, S.; Krishnamurthi, S. Exploring the Distinct Distribution of Archaeal Communities in Sites Contaminated with Explosives. Biomolecules 2022, 12, 489. https://doi.org/10.3390/biom12040489
Pal Y, Mayilraj S, Krishnamurthi S. Exploring the Distinct Distribution of Archaeal Communities in Sites Contaminated with Explosives. Biomolecules. 2022; 12(4):489. https://doi.org/10.3390/biom12040489
Chicago/Turabian StylePal, Yash, Shanmugam Mayilraj, and Srinivasan Krishnamurthi. 2022. "Exploring the Distinct Distribution of Archaeal Communities in Sites Contaminated with Explosives" Biomolecules 12, no. 4: 489. https://doi.org/10.3390/biom12040489