
Upregulated Proteasome Subunits in COVID-19 Patients: A Link with Hypoxemia, Lymphopenia and Inflammation

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Peripheral Blood Mononuclear Cell Isolation
2.3. Hypoxia and PX-478 In Vitro Models
2.4. Lipopolysaccharide (LPS) Stimulation Model
2.5. mRNA Isolation and Quantification by Quantitative PCR
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Study Subjects
3.2. COVID-19 Patients Overexpress Certain Proteasome Subunits
3.3. Proteasome Subunits Gene Expresion Are Related to Hypoxia
3.4. Some Proteasome Subunits Are Related to Poor Clinical Outcome
3.5. Some Proteasome Subunits Are Associated with Inflammatory Markers
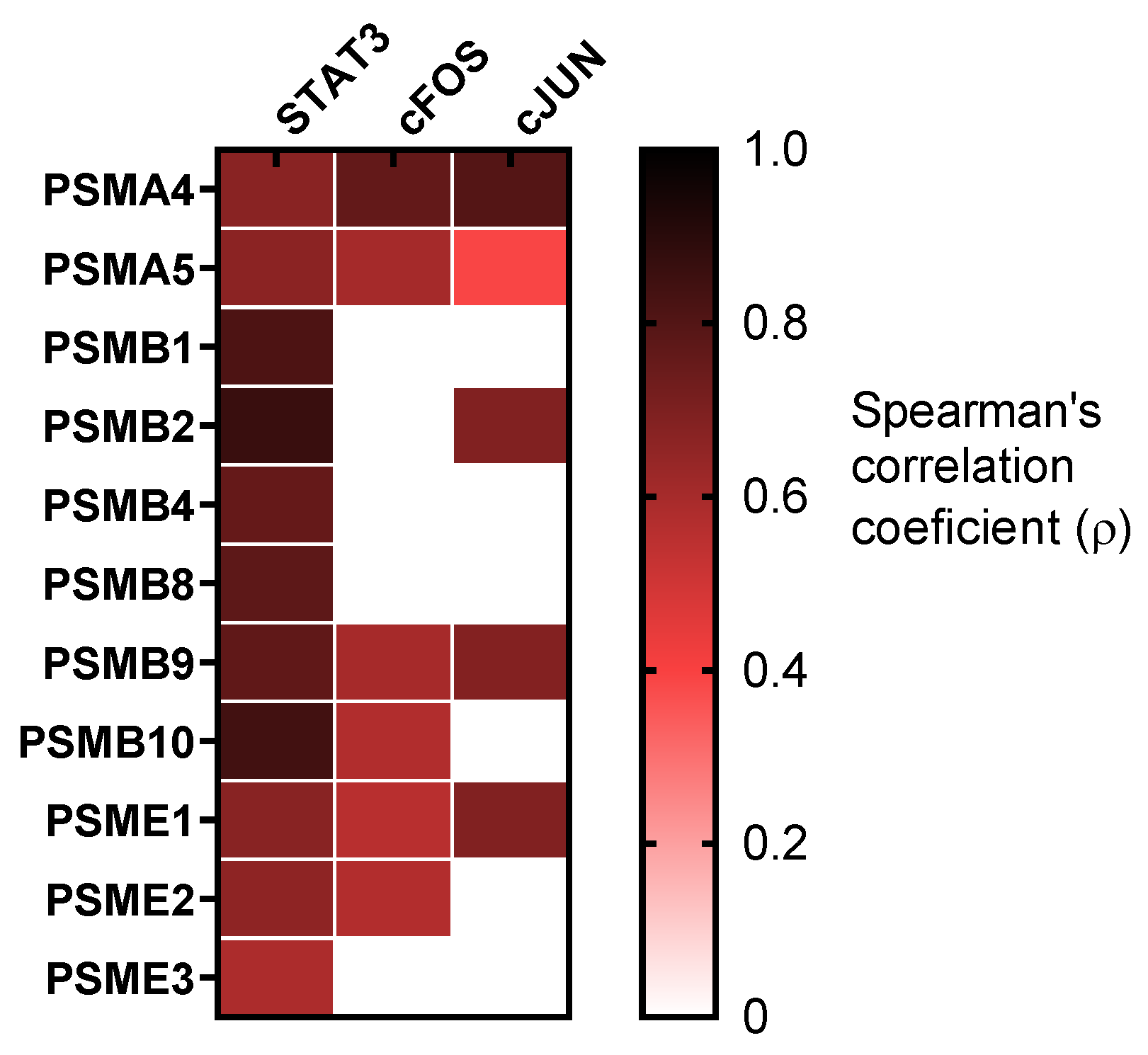
3.6. Proteasome Subunitswere Related with STAT3 Cascade
3.7. Proteasome Subunits Upregulation May Be a Cause and Not a Consequence of Inflammation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Jafarzadeh, S.; Nozari, P.; Mokhtari, P.; Nemati, M. Lymphopenia an important immunological abnormality in patients with COVID-19: Possible mechanisms. Scand. J. Immunol. 2021, 93, e12967. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Wang, G.; Yuan, Z.; et al. Reduction and Functional Exhaustion of T Cells in Patients With Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Meng, M.; Kumar, R.; Wu, Y.; Huang, J.; Deng, Y.; Weng, Z.; Yang, L. Lymphopenia is associated with severe coronavirus disease 2019 (COVID-19) infections: A systemic review and meta-analysis. Int. J. Infect. Dis. 2020, 96, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Qiu, Z.; Zhang, L.; Han, Y.; He, W.; Liu, Z.; Ma, X.; Fan, H.; Lu, W.; Xie, J.; et al. Significant changes of peripheral T lymphocyte subsets in patients with severe acute respiratory syndrome. J. Infect. Dis. 2004, 189, 648–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroemer, A.; Khan, K.; Plassmeyer, M.; Alpan, O.; Haseeb, M.A.; Gupta, R.; Fishbein, T.M. Inflammasome activation and pyroptosis in lymphopenic liver patients with COVID-19. J. Hepatol. 2020, 73, 1258–1262. [Google Scholar] [CrossRef] [PubMed]
- Fathi, N.; Rezaei, N. Lymphopenia in COVID-19: Therapeutic opportunities. Cell Biol. Int. 2020, 44, 1792–1797. [Google Scholar] [CrossRef]
- Cizmecioglu, A.; Akay Cizmecioglu, H.; Goktepe, M.H.; Emsen, A.; Korkmaz, C.; Esenkaya Tasbent, F.; Colkesen, F.; Artac, H. Apoptosis-induced T-cell lymphopenia is related to COVID-19 severity. J. Med. Virol. 2021, 93, 2867–2874. [Google Scholar] [CrossRef]
- Bucks, C.M.; Norton, J.A.; Boesteanu, A.C.; Mueller, Y.M.; Katsikis, P.D. Chronic antigen stimulation alone is sufficient to drive CD8+ T cell exhaustion. J. Immunol. 2009, 182, 6697–6708. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Li, X.W.; Mao, Y.; Wang, J.; Lu, H.Z.; Chen, Y.S.; Liang, Z.A.; Liang, L.; Zhang, S.J.; Zhang, B.; et al. Clinical features of the initial cases of 2009 pandemic influenza A (H1N1) virus infection in China. N. Engl. J. Med. 2009, 361, 2507–2517. [Google Scholar] [CrossRef] [Green Version]
- Perez-Padilla, R.; Fernandez, R.; Garcia-Sancho, C.; Franco-Marina, F.; Aburto, O.; Lopez-Gatell, H.; Bojorquez, I. Pandemic (H1N1) 2009 virus and Down syndrome patients. Emerg. Infect. Dis. 2010, 16, 1312–1314. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wo, J.; Shao, J.; Zhu, H.; Wu, N.; Li, M.; Yao, H.; Hu, M.; Dennin, R.H. SARS-coronavirus replicates in mononuclear cells of peripheral blood (PBMCs) from SARS patients. J. Clin. Virol. 2003, 28, 239–244. [Google Scholar] [CrossRef]
- Poutanen, S.M.; Low, D.E.; Henry, B.; Finkelstein, S.; Rose, D.; Green, K.; Tellier, R.; Draker, R.; Adachi, D.; Ayers, M.; et al. Identification of severe acute respiratory syndrome in Canada. N. Engl. J. Med. 2003, 348, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F.; et al. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Garvin, M.R.; Alvarez, C.; Miller, J.I.; Prates, E.T.; Walker, A.M.; Amos, B.K.; Mast, A.E.; Justice, A.; Aronow, B.; Jacobson, D. A mechanistic model and therapeutic interventions for COVID-19 involving a RAS-mediated bradykinin storm. Elife 2020, 9, e59177. [Google Scholar] [CrossRef] [PubMed]
- Shamaa, O.R.; Mitra, S.; Gavrilin, M.A.; Wewers, M.D. Monocyte Caspase-1 Is Released in a Stable, Active High Molecular Weight Complex Distinct from the Unstable Cell Lysate-Activated Caspase-1. PLoS ONE 2015, 10, e0142203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef] [Green Version]
- da Costa, L.S.; Outlioua, A.; Anginot, A.; Akarid, K.; Arnoult, D. RNA viruses promote activation of the NLRP3 inflammasome through cytopathogenic effect-induced potassium efflux. Cell Death Dis. 2019, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.; Moser, B.; Krenn, C.; Brunner, M.; Haisjackl, M.; Almer, G.; Gerlitz, S.; Wolner, E.; Boltz-Nitulescu, G.; Ankersmit, H.J. Susceptibility to programmed cell death in T-lymphocytes from septic patients: A mechanism for lymphopenia and Th2 predominance. Biochem. Biophys. Res. Commun. 2003, 308, 840–846. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Osmon, S.B.; Chang, K.C.; Wagner, T.H.; Coopersmith, C.M.; Karl, I.E. Accelerated lymphocyte death in sepsis occurs by both the death receptor and mitochondrial pathways. J. Immunol. 2005, 174, 5110–5118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankersmit, H.J.; Moser, B.; Roedler, S.; Teufel, I.; Zuckermann, A.; Roth, G.; Lietz, K.; Back, C.; Gerlitz, S.; Wolner, E.; et al. Death-inducing receptors and apoptotic changes in lymphocytes of patients with heart transplant vasculopathy. Clin. Exp. Immunol. 2002, 127, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, P.B.; Hrobjartsson, A.; Nielsen, D.L.; Henriksen, D.P.; Brabrand, M.; Lassen, A.T. Prevalence and prognosis of acutely ill patients with organ failure at arrival to hospital: Protocol for a systematic review. Syst. Rev. 2017, 6, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; Hlh Across Speciality Collaboration, U.K. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Xue, M.; Zhang, S.; Xie, J.; Zhang, X.; Liu, F.; Huang, Y.; Liu, L.; Liu, S.; Guo, F.; Yang, Y.; et al. Differential expression of genes associated with T lymphocytes function in septic patients with hypoxemia challenge. Ann. Transl. Med. 2019, 7, 810. [Google Scholar] [CrossRef] [PubMed]
- Yousef, A.A.; Suliman, G.A.; Mabrouk, M.M. The value of correlation of serum 20S proteasome concentration and percentage of lymphocytic apoptosis in critically ill patients: A prospective observational study. Crit. Care 2010, 14, R215. [Google Scholar] [CrossRef] [Green Version]
- Kravtsova-Ivantsiv, Y.; Ciechanover, A. The ubiquitin-proteasome system and activation of NF-kappaB: Involvement of the ubiquitin ligase KPC1 in p105 processing and tumor suppression. Mol. Cell Oncol. 2015, 2, e1054552. [Google Scholar] [CrossRef] [Green Version]
- Murata, S.; Yashiroda, H.; Tanaka, K. Molecular mechanisms of proteasome assembly. Nat. Rev. Mol. Cell Biol. 2009, 10, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, A.G.; O’Brien, T. Therapeutic strategies within the ubiquitin proteasome system. Cell Death Differ. 2010, 17, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Rieser, E.; Cordier, S.M.; Walczak, H. Linear ubiquitination: A newly discovered regulator of cell signalling. Trends Biochem. Sci. 2013, 38, 94–102. [Google Scholar] [CrossRef]
- Filardy, A.A.; He, J.; Bennink, J.; Yewdell, J.; Kelsall, B.L. Posttranscriptional control of NLRP3 inflammasome activation in colonic macrophages. Mucosal. Immunol. 2016, 9, 850–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israel, A. The IKK complex, a central regulator of NF-kappaB activation. Cold Spring Harb. Perspect Biol. 2010, 2, a000158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayagaki, N.; Phung, Q.; Chan, S.; Chaudhari, R.; Quan, C.; O’Rourke, K.M.; Eby, M.; Pietras, E.; Cheng, G.; Bazan, J.F.; et al. DUBA: A deubiquitinase that regulates type I interferon production. Science 2007, 318, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, M.; Chu, H.; Zhang, H.; Wu, H.; Song, G.; Wang, P.; Zhao, K.; Hou, J.; Wang, X.; et al. The ubiquitin E3 ligase TRIM31 promotes aggregation and activation of the signaling adaptor MAVS through Lys63-linked polyubiquitination. Nat. Immunol. 2017, 18, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Ding, T.; Zuo, Z.; Xu, Z.; Deng, J.; Wei, Z. Regulation of MAVS Expression and Signaling Function in the Antiviral Innate Immune Response. Front. Immunol. 2020, 11, 1030. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Liu, B.; Huai, W.; Yu, Z.; Wang, W.; Zhao, J.; Han, L.; Jiang, G.; Zhang, L.; Gao, C.; et al. The E3 ubiquitin ligase TRIM31 attenuates NLRP3 inflammasome activation by promoting proteasomal degradation of NLRP3. Nat. Commun. 2016, 7, 13727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrington, D.A.; Gregerson, D.S. Immunoproteasomes: Structure, function, and antigen presentation. Prog. Mol. Biol. Transl. Sci. 2012, 109, 75–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groettrup, M.; Kirk, C.J.; Basler, M. Proteasomes in immune cells: More than peptide producers? Nat. Rev. Immunol. 2010, 10, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, E.; Nicola, A.V.; Liu, J. Multiscale perspectives of virus entry via endocytosis. Virol. J. 2013, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicola, A.V.; Aguilar, H.C.; Mercer, J.; Ryckman, B.; Wiethoff, C.M. Virus entry by endocytosis. Adv. Virol. 2013, 2013, 469538. [Google Scholar] [CrossRef] [PubMed]
- Randow, F.; Lehner, P.J. Viral avoidance and exploitation of the ubiquitin system. Nat. Cell Biol. 2009, 11, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Raaben, M.; Posthuma, C.C.; Verheije, M.H.; te Lintelo, E.G.; Kikkert, M.; Drijfhout, J.W.; Snijder, E.J.; Rottier, P.J.; de Haan, C.A. The ubiquitin-proteasome system plays an important role during various stages of the coronavirus infection cycle. J. Virol. 2010, 84, 7869–7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Elkin, B.A.; Knaap, R.C.; Johnson, G.G.; Dalebout, T.J.; Ninaber, D.K.; van Kasteren, P.B.; Bredenbeek, P.J.; Snijder, E.J.; Kikkert, M.; Mark, B.L. Crystal structure of the Middle East respiratory syndrome coronavirus (MERS-CoV) papain-like protease bound to ubiquitin facilitates targeted disruption of deubiquitinating activity to demonstrate its role in innate immune suppression. J. Biol. Chem. 2014, 289, 34667–34682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yang, X.; Zheng, Y.; Yang, Y.; Xing, Y.; Chen, Z. SARS coronavirus papain-like protease inhibits the type I interferon signaling pathway through interaction with the STING-TRAF3-TBK1 complex. Protein. Cell 2014, 5, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak-Kroizman, T.R.; Hostetter, G.; Posner, R.; Aziz, M.; Hu, C.; Demeure, M.J.; Von Hoff, D.; Hingorani, S.R.; Palculict, T.B.; Izzo, J.; et al. Hypoxia triggers hedgehog-mediated tumor-stromal interactions in pancreatic cancer. Cancer Res. 2013, 73, 3235–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, D.L.; Powis, G.; Thitai-Kumar, A.; He, Y.; Bankson, J.; Williams, R.; Lemos, R.; Oh, J.; Volgin, A.; Soghomonyan, S.; et al. The selective hypoxia inducible factor-1 inhibitor PX-478 provides in vivo radiosensitization through tumor stromal effects. Mol. Cancer Ther. 2009, 8, 947–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, T.; Kubli, S.P.; Yoshinaga, S.K.; Pfeffer, K.; Mak, T.W. An aberrant STAT pathway is central to COVID-19. Cell Death Differ. 2020, 27, 3209–3225. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Nemati, M.; Jafarzadeh, S. Contribution of STAT3 to the pathogenesis of COVID-19. Microb. Pathog. 2021, 154, 104836. [Google Scholar] [CrossRef]
- Cui, R.; Wang, C.; Li, T.; Hua, J.; Zhao, T.; Ren, L.; Wang, Y.; Li, Y. Carboxypeptidase N1 is anticipated to be a synergy metrics for chemotherapy effectiveness and prognostic significance in invasive breast cancer. Cancer Cell Int. 2021, 21, 571. [Google Scholar] [CrossRef]
- Qureshi, N.; Morrison, D.C.; Reis, J. Proteasome protease mediated regulation of cytokine induction and inflammation. Biochim. Biophys. Acta 2012, 1823, 2087–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H. Interplay between the virus and the ubiquitin-proteasome system: Molecular mechanism of viral pathogenesis. Curr. Opin. Virol. 2016, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Madura, K. Increased proteasome activity, ubiquitin-conjugating enzymes, and eEF1A translation factor detected in breast cancer tissue. Cancer Res. 2005, 65, 5599–5606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlt, A.; Bauer, I.; Schafmayer, C.; Tepel, J.; Muerkoster, S.S.; Brosch, M.; Roder, C.; Kalthoff, H.; Hampe, J.; Moyer, M.P.; et al. Increased proteasome subunit protein expression and proteasome activity in colon cancer relate to an enhanced activation of nuclear factor E2-related factor 2 (Nrf2). Oncogene 2009, 28, 3983–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motosugi, R.; Murata, S. Dynamic Regulation of Proteasome Expression. Front. Mol. Biosci. 2019, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Fu, J.; Ha, S.W.; Ju, D.; Zheng, J.; Li, L.; Xie, Y. The CCAAT box-binding transcription factor NF-Y regulates basal expression of human proteasome genes. Biochim. Biophys. Acta 2012, 1823, 818–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilchez, D.; Boyer, L.; Morantte, I.; Lutz, M.; Merkwirth, C.; Joyce, D.; Spencer, B.; Page, L.; Masliah, E.; Berggren, W.T.; et al. Increased proteasome activity in human embryonic stem cells is regulated by PSMD11. Nature 2012, 489, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Vangala, J.R.; Dudem, S.; Jain, N.; Kalivendi, S.V. Regulation of PSMB5 protein and beta subunits of mammalian proteasome by constitutively activated signal transducer and activator of transcription 3 (STAT3): Potential role in bortezomib-mediated anticancer therapy. J. Biol. Chem. 2014, 289, 12612–12622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, S.K.; Lee, C.S.; Young, P.; Beskow, A.; Chan, J.Y.; Deshaies, R.J. Transcription factor Nrf1 mediates the proteasome recovery pathway after proteasome inhibition in mammalian cells. Mol. Cell 2010, 38, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.Y.; Moriyama, M.; Chang, M.F.; Ichinohe, T. Severe Acute Respiratory Syndrome Coronavirus Viroporin 3a Activates the NLRP3 Inflammasome. Front. Microbiol. 2019, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Panfair, D.; Ramamurthy, A.; Kusmierczyk, A.R. Alpha-ring Independent Assembly of the 20S Proteasome. Sci. Rep. 2015, 5, 13130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendt, C.S.; Hochstrasser, M. Identification of the yeast 20S proteasome catalytic centers and subunit interactions required for active-site formation. Proc. Natl. Acad. Sci. USA 1997, 94, 7156–7161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinemeyer, W.; Ramos, P.C.; Dohmen, R.J. The ultimate nanoscale mincer: Assembly, structure and active sites of the 20S proteasome core. Cell Mol. Life Sci. 2004, 61, 1562–1578. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Takahama, Y.; Kasahara, M.; Tanaka, K. The immunoproteasome and thymoproteasome: Functions, evolution and human disease. Nat. Immunol. 2018, 19, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, N.; Hemmi, H.; Kinjo, N.; Ohnishi, H.; Hamazaki, J.; Mishima, H.; Kinoshita, A.; Mizushima, T.; Hamada, S.; Hamada, K.; et al. Heterozygous missense variant of the proteasome subunit beta-type 9 causes neonatal-onset autoinflammation and immunodeficiency. Nat. Commun. 2021, 12, 6819. [Google Scholar] [CrossRef] [PubMed]
- Sarrabay, G.; Mechin, D.; Salhi, A.; Boursier, G.; Rittore, C.; Crow, Y.; Rice, G.; Tran, T.A.; Cezar, R.; Duffy, D.; et al. PSMB10, the last immunoproteasome gene missing for PRAAS. J. Allergy Clin. Immunol. 2020, 145, 1015–1017e1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.; Aravind, L.; Oania, R.; McDonald, W.H.; Yates, J.R., III; Koonin, E.V.; Deshaies, R.J. Role of Rpn11 metalloprotease in deubiquitination and degradation by the 26S proteasome. Science 2002, 298, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; You, H.; Kong, D.; Zheng, K.; Tang, R. The interaction of hepatitis B virus with the ubiquitin proteasome system in viral replication and associated pathogenesis. Virol. J. 2019, 16, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sijts, A.; Sun, Y.; Janek, K.; Kral, S.; Paschen, A.; Schadendorf, D.; Kloetzel, P.M. The role of the proteasome activator PA28 in MHC class I antigen processing. Mol. Immunol. 2002, 39, 165–169. [Google Scholar] [CrossRef]
- McCarthy, M.K.; Weinberg, J.B. The immunoproteasome and viral infection: A complex regulator of inflammation. Front. Microbiol. 2015, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Su, F.; Zhang, L.; Liu, F.; Fan, W.; Li, Z.; Ma, J. Identifying Potential Diagnostic Genes for Diabetic Nephropathy Based on Hypoxia and Immune Status. J. Inflamm. Res. 2021, 14, 6871–6891. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yu, P.; Yang, F.; He, Q.; Jiang, B.; Zheng, L.; Wang, Q.; Wang, J.; Qiu, H.; Wang, H.; et al. PSMB4 inhibits cardiomyocyte apoptosis via activating NF-kappaB signaling pathway during myocardial ischemia/reperfusion injury. J. Mol. Histol. 2021, 52, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, L.; Yun, X.; Yang, K.; Liao, D.; Tian, L.; Jiang, H.; Lu, W. Proteomic analysis reveals that proteasome subunit beta 6 is involved in hypoxia-induced pulmonary vascular remodeling in rats. PLoS ONE 2013, 8, e67942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, G.A.; Moser, B.; Krenn, C.; Roth-Walter, F.; Hetz, H.; Richter, S.; Brunner, M.; Jensen-Jarolim, E.; Wolner, E.; Hoetzenecker, K.; et al. Heightened levels of circulating 20S proteasome in critically ill patients. Eur. J. Clin. Invest. 2005, 35, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Wendt, R.; Lingitz, M.T.; Laggner, M.; Mildner, M.; Traxler, D.; Graf, A.; Krotka, P.; Moser, B.; Hoetzenecker, K.; Kalbitz, S.; et al. Clinical Relevance of Elevated Soluble ST2, HSP27 and 20S Proteasome at Hospital Admission in Patients with COVID-19. Biology 2021, 10, 1186. [Google Scholar] [CrossRef] [PubMed]
- Palombella, V.J.; Rando, O.J.; Goldberg, A.L.; Maniatis, T. The ubiquitin-proteasome pathway is required for processing the NF-kappa B1 precursor protein and the activation of NF-kappa B. Cell 1994, 78, 773–785. [Google Scholar] [CrossRef]
- Guan, K.; Wei, C.; Zheng, Z.; Song, T.; Wu, F.; Zhang, Y.; Cao, Y.; Ma, S.; Chen, W.; Xu, Q.; et al. MAVS Promotes Inflammasome Activation by Targeting ASC for K63-Linked Ubiquitination via the E3 Ligase TRAF3. J. Immunol. 2015, 194, 4880–4890. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.S.; de Sa, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Goncalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Wu, S.; Zhang, L.; Yang, M.; Lin, Q.; Xie, Q.; Ding, H.; Lian, N.; Gao, S.; Huang, Y.; et al. Role of monocyte tissue factor on patients with non-small cell lung cancer. Clin. Respir. J. 2018, 12, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Iness, A.; Yoon, J.; Grider, J.R.; Murthy, K.S.; Kellum, J.M.; Kuemmerle, J.F. Noncanonical STAT3 activation regulates excess TGF-beta1 and collagen I expression in muscle of stricturing Crohn’s disease. J. Immunol. 2015, 194, 3422–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinoco, R.; Alcalde, V.; Yang, Y.; Sauer, K.; Zuniga, E.I. Cell-intrinsic transforming growth factor-beta signaling mediates virus-specific CD8+ T cell deletion and viral persistence in vivo. Immunity 2009, 31, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, K.; Kaisho, T.; Yoshida, N.; Takeda, J.; Kishimoto, T.; Akira, S. Stat3 activation is responsible for IL-6-dependent T cell proliferation through preventing apoptosis: Generation and characterization of T cell-specific Stat3-deficient mice. J. Immunol. 1998, 161, 4652–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients. Emerg. Microbes. Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, P.; Wen, W.; James, M.A.; Wang, Y.; Bailey-Wilson, J.E.; Amos, C.I.; Pinney, S.M.; Yang, P.; de Andrade, M.; et al. Haplotype and cell proliferation analyses of candidate lung cancer susceptibility genes on chromosome 15q24-25.1. Cancer Res. 2009, 69, 7844–7850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, J.; Zhang, S.; Wu, H.; Lu, J.; Lu, Y.; Wang, F.; Zhao, W.; Zhan, P.; Lu, J.; Fang, Q.; et al. Deubiquitinase PSMD14 enhances hepatocellular carcinoma growth and metastasis by stabilizing GRB2. Cancer Lett. 2020, 469, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Walter, R.F.H.; Sydow, S.R.; Berg, E.; Kollmeier, J.; Christoph, D.C.; Christoph, S.; Eberhardt, W.E.E.; Mairinger, T.; Wohlschlaeger, J.; Schmid, K.W.; et al. Bortezomib sensitivity is tissue dependent and high expression of the 20S proteasome precludes good response in malignant pleural mesothelioma. Cancer Manag. Res. 2019, 11, 8711–8720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConkey, D.J.; Zhu, K. Mechanisms of proteasome inhibitor action and resistance in cancer. Drug Resist. Updat. 2008, 11, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Bazzaro, M.; Lee, M.K.; Zoso, A.; Stirling, W.L.; Santillan, A.; Shih Ie, M.; Roden, R.B. Ubiquitin-proteasome system stress sensitizes ovarian cancer to proteasome inhibitor-induced apoptosis. Cancer Res. 2006, 66, 3754–3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nencioni, A.; Garuti, A.; Schwarzenberg, K.; Cirmena, G.; Dal Bello, G.; Rocco, I.; Barbieri, E.; Brossart, P.; Patrone, F.; Ballestrero, A. Proteasome inhibitor-induced apoptosis in human monocyte-derived dendritic cells. Eur. J. Immunol. 2006, 36, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neutzner, A.; Li, S.; Xu, S.; Karbowski, M. The ubiquitin/proteasome system-dependent control of mitochondrial steps in apoptosis. Semin. Cell Dev. Biol. 2012, 23, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Proulx, J.; Borgmann, K.; Park, I.W. Role of Virally-Encoded Deubiquitinating Enzymes in Regulation of the Virus Life Cycle. Int. J. Mol. Sci. 2021, 22, 4438. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Ackermann, K.; Stuart, M.; Wex, C.; Protzer, U.; Schatzl, H.M.; Gilch, S. Severe acute respiratory syndrome coronavirus replication is severely impaired by MG132 due to proteasome-independent inhibition of M-calpain. J. Virol. 2012, 86, 10112–10122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.Z.; Bartczak, A.; Zhang, J.; Khattar, R.; Chen, L.; Liu, M.F.; Edwards, A.; Levy, G.; McGilvray, I.D. Proteasome inhibition in vivo promotes survival in a lethal murine model of severe acute respiratory syndrome. J. Virol. 2010, 84, 12419–12428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaben, M.; Grinwis, G.C.; Rottier, P.J.; de Haan, C.A. The proteasome inhibitor Velcade enhances rather than reduces disease in mouse hepatitis coronavirus-infected mice. J. Virol. 2010, 84, 7880–7885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhitano, L.; Tibullo, D.; Giallongo, C.; Lazzarino, G.; Tartaglia, N.; Galimberti, S.; Volti, G.L.; Palumbo, G.A.; Liso, A. Proteasome Inhibitors as a Possible Therapy for SARS-CoV-2. Int. J. Mol. Sci. 2020, 21, 3622. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age, year | 57 ± 13 | |
| Sex, Male/Female | 30/14 | |
| Days since onset of symptoms | 7.7 ± 4.0 | |
| Symptoms at admission | ||
| Cough | 28 (64) | |
| Active fever | 32 (73) | |
| Dyspnea | 31 (70) | |
| Myalgia | 15 (34) | |
| Sputum production | 8 (18) | |
| Chest tightness | 3 (7) | |
| Headache | 8 (18) | |
| Fatigue | 12 (27) | |
| Anorexia | 5 (11) | |
| Nausea | 3 (7) | |
| Diarrhea | 7 (16) | |
| Chest pain | 5 (11) | |
| Anosmia | 6 (14) | |
| Comorbidities | ||
| Hypertension | 12 (27) | |
| Coronary artery disease | 3 (7) | |
| Diabetes mellitus | 13 (30) | |
| Obesity | 10 (23) | |
| Chronic lung disease | 4 (9) | |
| Chronic kidney disease | 1 (2) | |
| Hypothyroidism | 2 (3) | |
| Smoking history | ||
| Current, n (%) | 31 (70) | |
| Former, (%) | 3 (7) | |
| Never, n (%) | 10 (23) | |
| Pneumonia severity scores | ||
| CURB-65 | 0.84 ± 0.88 | |
| Fine risk class | 2.4 ± 1.3 | |
| Laboratory findings | ||
| PaO2, mmHg | 65.0 ± 25.9 | |
| PaO2/FiO2 ratio | 282.5 ± 113.5 | |
| PaCO2, mmHg | 36.8 ± 14.7 | |
| White cell count, 103 cells/µL | 6.74 ± 4.41 | |
| Neutrophils, 103 cells/µL | 4.90 ± 3.10 | |
| Lymphocytes, 103 cells/µL | 1.31 ± 2.04 | |
| Eosinophils, 103 cells/µL | 0.02 ± 0.03 | |
| Platelets, 103 cells/µL | 227.7 ± 101.8 | |
| Hemoglobin, g/dL | 13.9 ± 4.7 | |
| C-reactive protein, mg/L | 68.8 ± 57.7 | |
| Aspartate aminotransferase, U/L | 48.8 ± 38.5 | |
| Alanine aminotransferase, IU/L | 43.7 ± 29.6 | |
| Υ-Glutamyltransferase, IU/L | 84.8 ± 81.0 | |
| Bilirubin, µmol/L | 0.51 ± 0.28 | |
| Albumin, g/L | 4.34 ± 1.82 | |
| Ferritin, ng/mL | 839.1 ± 899.4 | |
| Lactate dehydrogenase, U/L | 304.2 ± 163.6 | |
| D-dimer, ng/mL | 697.4 ± 677.4 | |
| Fibrinogen, mg/dL | 672.1 ± 270.9 | |
| Evolution results | ||
| Duration of hospital admission, days | 9.9 ± 5.6 | |
| ICU admission, n (%) | 2 (4.5) | |
| Exitus, n (%) | 2 (4.5) | |
| Gene | Lymphocytes (×103 Cells/mm3) | PaO2/FiO2 Ratio | C-Reactive Protein (mg/L) | Ferritin (mg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | High | p | Low | High | p | Low | High | p | Low | High | p | |
| PSMA4 | 1.018 ± 0.293 | 0.740 ± 0.168 | 0.0084 | 320.0 ± 92.90 | 232.9 ± 105.0 | 0.028 | 46.97 ± 32.63 | 90.42 ± 67.20 | 0.044 | 486.1 ± 532.2 | 1361 ± 1158 | 0.024 |
| PSMA5 | 1.035 ± 0.476 | 0.734 ± 0.164 | 0.0498 | 302.2 ± 75.82 | 229.5 ± 88.98 | 0.0429 | 50.36 ± 34.32 | 94.5 ± 66.77 | 0.0470 | 473.8 ± 475.6 | 927.6 ± 471 | 0.0339 |
| PSMB1 | 0.983 ± 0.306 | 0.655 ± 0.199 | 0.056 | 288.3 ± 100.8 | 247.5 ± 82.38 | 0.438 | 41.93 ± 38.4 | 47.1 ± 29.8 | 0.791 | 707.7 ± 684.4 | 876.5 ± 696.4 | 0.659 |
| PSMB2 | 0.983 ± 0.306 | 0.626 ± 0.207 | 0.048 | 288.3 ± 100.8 | 245.7 ± 88.81 | 0.441 | 41.93 ± 38.4 | 47.20 ± 32.2 | 0.796 | 707.7 ± 684.4 | 796.3 ± 711.2 | 0.824 |
| PSMB4 | 1.030 ± 0.380 | 0.784 ± 0.360 | 0.301 | 295.7 ± 106.5 | 247.7 ± 90.97 | 0.407 | 48.25 ± 35.95 | 37.51 ± 30.14 | 0.576 | 772.3 ± 621.6 | 775.3 ± 734.7 | 0.994 |
| PSMB8 | 0.957 ± 0.287 | 0.919 ± 0.528 | 0.164 | 284.3 ± 92.6 | 262.6 ± 95.12 | 0.663 | 42.57 ± 35.08 | 47.81 ± 29.88 | 0.763 | 812.0 ± 683.0 | 747.5 ± 672.7 | 0.857 |
| PSMB9 | 0.873 ± 0.317 | 0.976 ± 0.471 | 0.623 | 302.0 ± 85.14 | 244.4 ± 88.27 | 0.209 | 52.13 ± 35.46 | 38.77 ± 26.52 | 0.423 | 804.9 ± 690.5 | 707.1 ± 639.2 | 0.777 |
| PSMB10 | 0.798 ± 0.272 | 0.963 ± 0.456 | 0.441 | 296.2 ± 91.72 | 241.6 ± 85.97 | 0.283 | 58.45 ± 34.26 | 34.71 ± 29.01 | 0.202 | 936.0 ± 654.0 | 705.3 ± 708.5 | 0.541 |
| PSME1 | 0.920 ± 0.311 | 0.885 ± 0.489 | 0.867 | 285.6 ± 89.74 | 249.2 ± 86.46 | 0.409 | 56.75 ± 37.74 | 42.46 ± 33.01 | 0.422 | 614.1 ± 618.9 | 795.3 ± 687.2 | 0.576 |
| PSME2 | 0.831 ± 0.381 | 0.974 ± 0.423 | 0.491 | 289.4 ± 93.14 | 245.9 ± 81.42 | 0.325 | 58.65 ± 37.17 | 40.77 ± 32.65 | 0.312 | 641.8 ± 621.7 | 770.8 ± 691.2 | 0.691 |
| PSME3 | 0.939 ± 0.370 | 0.866 ± 0.443 | 0.728 | 294.6 ± 91.56 | 241.2 ± 79.86 | 0.223 | 52.24 ± 32.69 | 46.47 ± 38.63 | 0.743 | 779.6 ± 626.1 | 648.2 ± 687.5 | 0.686 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfaro, E.; Díaz-García, E.; García-Tovar, S.; Zamarrón, E.; Mangas, A.; Galera, R.; López-Collazo, E.; García-Rio, F.; Cubillos-Zapata, C. Upregulated Proteasome Subunits in COVID-19 Patients: A Link with Hypoxemia, Lymphopenia and Inflammation. Biomolecules 2022, 12, 442. https://doi.org/10.3390/biom12030442
Alfaro E, Díaz-García E, García-Tovar S, Zamarrón E, Mangas A, Galera R, López-Collazo E, García-Rio F, Cubillos-Zapata C. Upregulated Proteasome Subunits in COVID-19 Patients: A Link with Hypoxemia, Lymphopenia and Inflammation. Biomolecules. 2022; 12(3):442. https://doi.org/10.3390/biom12030442
Chicago/Turabian StyleAlfaro, Enrique, Elena Díaz-García, Sara García-Tovar, Ester Zamarrón, Alberto Mangas, Raúl Galera, Eduardo López-Collazo, Francisco García-Rio, and Carolina Cubillos-Zapata. 2022. "Upregulated Proteasome Subunits in COVID-19 Patients: A Link with Hypoxemia, Lymphopenia and Inflammation" Biomolecules 12, no. 3: 442. https://doi.org/10.3390/biom12030442