
Emerging Roles for G Protein-Coupled Estrogen Receptor 1 in Cardio-Renal Health: Implications for Aging


 , ,
, ,  and
and
Abstract
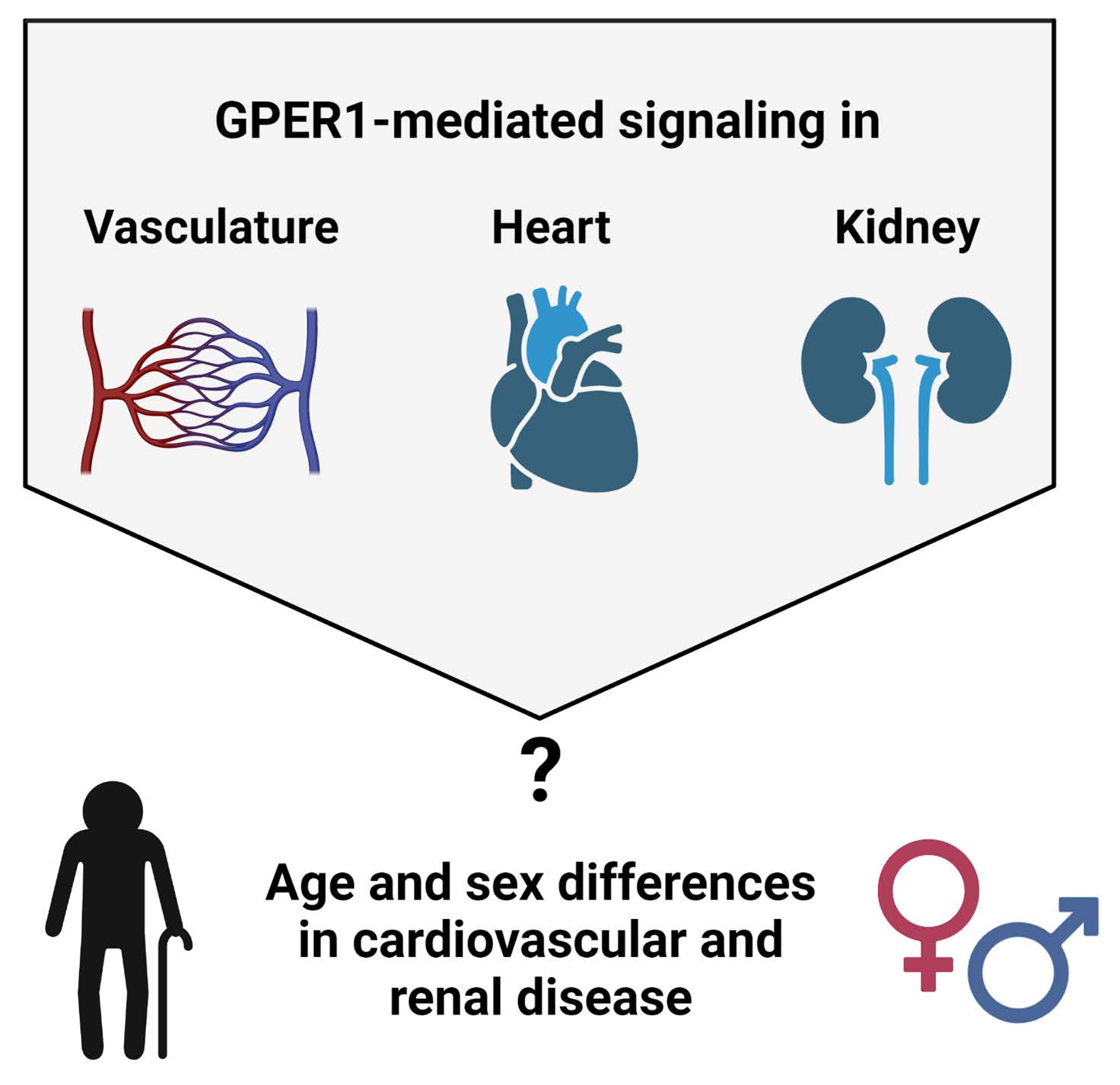
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. Cardio-Renal Health
1.2. Aging as a Contributor to Cardio-Renal Disease
1.3. Sex and Sex Hormones as Contributors to Cardio-Renal Disease
2. GPER1 as a Regulator for Cardio-Renal Health
2.1. Expression of GPER1 within the Cardiovascular and Renal Systems
2.2. Pharmacological and Genetic Tools to Study GPER1 Function
2.3. Mediators of GPER1 Actions within the Cardiovascular and Renal Systems
2.3.1. Renin-Angiotensin Aldosterone System
2.3.2. Endothelin-1
2.3.3. Nitric Oxide
2.3.4. Reactive Oxygen Species
2.3.5. Immunity and Inflammation
3. GPER1 Signaling in Cardiovascular and Renal Disease
3.1. Hypertension
3.2. Heart Disease
3.3. Kidney Disease
4. Future Directions and Translational Perspective
Author Contributions
Funding
Conflicts of Interest
References
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 20 January 2022).
- Murphy, S.L.; Kochanek, K.D.; Xu, J.Q.; Arias, E. Mortality in the United States, 2020; NCHS Data Brief, no 427; National Center for Health Statistics: Hyattsville, MD, USA, 2021. [CrossRef]
- Ronco, C.; Ronco, F. Cardio-renal syndromes: A systematic approach for consensus definition and classification. Heart Fail. Rev. 2011, 17, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 Update. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef] [PubMed]
- USRDS. 2018 USRDS Annual Data Report: Epidemiology of Kidney Disease in the United States; National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Disaeases: Bethesda, MD, USA, 2018.
- World Health Organization. GHE: Life Expectancy and Healthy Life Expectancy. Available online: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-life-expectancy-and-healthy-life-expectancy (accessed on 10 January 2022).
- Mather, M.; Scommegna, P.; Kilduff, L. Fact sheet: Aging in the United States. Popul. Ref. Bur. 2019, 70, 2. [Google Scholar]
- Crude Percentage with Cardiovascular Disease among All Adults by Age. Available online: https://nccd.cdc.gov/ckd/detail.aspx?Qnum=Q151 (accessed on 16 January 2022).
- Centers for Disease Control and Prevention. Chronic Kidney Disease in the United States, 2021; US Department of Health and Human Services, Centers for Disease Control and Prevention: Atlanta, GA, USA, 2021.
- Barzilai, N.; Huffman, D.M.; Muzumdar, R.H.; Bartke, A. The Critical Role of Metabolic Pathways in Aging. Diabetes 2012, 61, 1315–1322. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, T.B.L. Understanding ageing from an evolutionary perspective. J. Intern. Med. 2008, 263, 117–127. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münzel, T.; Camici, G.G.; Maack, C.; Bonetti, N.R.; Fuster, V.; Kovacic, J.C. Impact of Oxidative Stress on the Heart and Vasculature: Part 2 of a 3-Part Series. J. Am. Coll. Cardiol. 2017, 70, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Woollard, K. Atherosclerosis. Adv. Exp. Med. Biol. 2017, 1003, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.; Gona, P.; Larson, M.G.; Aragam, J.; Lee, D.S.; Mitchell, G.F.; Levy, D.; Cheng, S.; Benjamin, E.J.; Vasan, R.S. Aortic Root Remodeling and Risk of Heart Failure in the Framingham Heart Study. JACC Heart Fail. 2013, 1, 79–83. [Google Scholar] [CrossRef]
- Gramley, F.; Lorenzen, J.; Knackstedt, C.; Rana, O.R.; Saygili, E.; Frechen, D.; Stanzel, S.; Pezzella, F.; Koellensperger, E.; Weiss, C.; et al. Age-related atrial fibrosis. AGE 2008, 31, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obas, V.; Vasan, R.S. The aging heart. Clin. Sci. 2018, 132, 1367–1382. [Google Scholar] [CrossRef]
- Wang, X.; Vrtiska, T.J.; Avula, R.T.; Walters, L.R.; Chakkera, H.A.; Kremers, W.K.; Lerman, L.O.; Rule, A.D. Age, kidney function, and risk factors associate differently with cortical and medullary volumes of the kidney. Kidney Int. 2014, 85, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyengaard, J.R.; Bendtsen, T.F. Glomerular number and size in relation to age, kidney weight, and body surface in normal man. Anat. Rec. 1992, 232, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Poggio, E.D.; Rule, A.D.; Tanchanco, R.; Arrigain, S.; Butler, R.S.; Srinivas, T.; Stephany, B.R.; Meyer, K.H.; Nurko, S.; Fatica, R.A.; et al. Demographic and clinical characteristics associated with glomerular filtration rates in living kidney donors. Kidney Int. 2009, 75, 1079–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulladosa, X.; Moreso, F.; Narváez, J.A.; Grinyó, J.M.; Serón, D. Estimation of total glomerular number in stable renal transplants. J. Am. Soc. Nephrol. 2003, 14, 2662–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuboi, N.; Kawamura, T.; Koike, K.; Okonogi, H.; Hirano, K.; Hamaguchi, A.; Miyazaki, Y.; Ogura, M.; Joh, K.; Utsunomiya, Y.; et al. Glomerular density in renal biopsy specimens predicts the long-term prognosis of iga nephropathy. Clin. J. Am. Soc. Nephrol. 2009, 5, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.C.; Workeneh, B.; Busque, S.; Blouch, K.; Derby, G.; Myers, B.D. Glomerular function, structure, and number in renal allografts from older deceased donors. J. Am. Soc. Nephrol. 2008, 20, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.C.; Busque, S.; Workeneh, B.; Ho, B.; Derby, G.; Blouch, K.L.; Sommer, F.G.; Edwards, B.; Myers, B.D. Effects of aging on glomerular function and number in living kidney donors. Kidney Int. 2010, 78, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Weidner, G. Why do men get more heart disease than women? an international perspective. J. Am. Coll. Health 2000, 48, 291–294. [Google Scholar] [CrossRef]
- Möller-Leimkühler, A.M. Gender differences in cardiovascular disease and comorbid depression. Dialogues Clin. Neurosci. 2007, 9, 71–83. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, Z.; Sun, A.; Deng, X. Gender differences in cardiovascular disease. Med. Nov. Technol. Devices 2019, 4, 100025. [Google Scholar] [CrossRef]
- Maas, A.; Appelman, Y. Gender differences in coronary heart disease. Neth. Heart J. 2010, 18, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.M.; West, J.H.; Korenman, S.G. The Menopausal Transition: Analysis of LH, FSH, Estradiol, and Progesterone Concentrations During Menstrual Cycles of Older Women. J. Clin. Endocrinol. Metab. 1976, 42, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Ferrini, R.L.; Barrett-Connor, E. Sex Hormones and Age: A Cross-sectional Study of Testosterone and Estradiol and Their Bioavailable Fractions in Community-dwelling Men. Am. J. Epidemiol. 1998, 147, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Kloner, R.A.; Carson, C., 3rd; Dobs, A.; Kopecky, S.; Mohler, E.R., 3rd. Testosterone and Cardiovascular Disease. J. Am. Coll. Cardiol. 2016, 67, 545–557. [Google Scholar] [CrossRef]
- Araujo, A.B.; Dixon, J.M.; Suarez, E.A.; Murad, M.H.; Guey, L.T.; Wittert, G.A. Clinical review: Endogenous Testosterone and Mortality in Men: A Systematic Review and Meta-Analysis. J. Clin. Endocrinol. Metab. 2011, 96, 3007–3019. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Rastrelli, G.; Monami, M.; Guay, A.; Buvat, J.; Sforza, A.; Forti, G.; Mannucci, E.; Maggi, M. Hypogonadism as a risk factor for cardiovascular mortality in men: A meta-analytic study. Eur. J. Endocrinol. 2011, 165, 687–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, H.A.; Longcope, C.; Derby, C.A.; Johannes, C.B.; Araujo, A.B.; Coviello, A.D.; Bremner, W.J.; McKinlay, J.B. Age Trends in the Level of Serum Testosterone and Other Hormones in Middle-Aged Men: Longitudinal Results from the Massachusetts Male Aging Study. J. Clin. Endocrinol. Metab. 2002, 87, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.D.; Ruiz, A.L.; Yanes, L.L.; Iliescu, R.; Yuan, K.; Moulana, M.; Racusen, L.C.; Reckelhoff, J.F. Testosterone Supplementation in Male Obese Zucker Rats Reduces Body Weight and Improves Insulin Sensitivity but Increases Blood Pressure. Hypertension 2012, 59, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basaria, S.; Coviello, A.D.; Travison, T.G.; Storer, T.W.; Farwell, W.R.; Jette, A.; Eder, R.; Tennstedt, S.; Ulloor, J.; Zhang, A.; et al. Adverse Events Associated with Testosterone Administration. N. Engl. J. Med. 2010, 363, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmasso, C.; Patil, C.N.; Cardozo, L.L.Y.; Romero, D.G.; Maranon, R.O. Cardiovascular and Metabolic Consequences of Testosterone Supplements in Young and Old Male Spontaneously Hypertensive Rats: Implications for Testosterone Supplements in Men. J. Am. Heart Assoc. 2017, 6, e007074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojeda, N.; Grigore, D.; Robertson, E.B.; Alexander, B.T. Estrogen protects against increased blood pressure in postpubertal female growth restricted offspring. Hypertension 2007, 50, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Affinito, P.; Palomba, S.; Bonifacio, M.; Fontana, D.; Izzo, R.; Trimarco, B.; Nappi, C. Effects of hormonal replacement therapy in postmenopausal hypertensive patients. Maturitas 2001, 40, 75–83. [Google Scholar] [CrossRef]
- Chappell, M.C.; Gallagher, P.E.; Averill, D.B.; Ferrario, C.M.; Brosnihan, K.B. Estrogen or the AT1 antagonist olmesartan reverses the development of profound hypertension in the congenic mren2.lewis rat. Hypertension 2003, 42, 781–786. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa-Laborde, C.; Craig, T.; Zheng, W.; Ji, H.; Haywood, J.R.; Sandberg, K. Ovariectomy augments hypertension in aging female dahl salt-sensitive rats. Hypertension 2004, 44, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Barton, M.; Filardo, E.; Lolait, S.J.; Thomas, P.; Maggiolini, M.; Prossnitz, E.R. Twenty years of the G protein-coupled estrogen receptor GPER: Historical and personal perspectives. J. Steroid Biochem. Mol. Biol. 2017, 176, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Filardo, E.J.; Thomas, P. Minireview: G protein-coupled estrogen receptor-1, gper-1: Its mechanism of action and role in female reproductive cancer, renal and vascular physiology. Endocrinology 2012, 153, 2953–2962. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, B.-O.; Olde, B.; Leeb-Lundberg, L.F. G protein-coupled oestrogen receptor 1 (GPER1)/GPR30: A new player in cardiovascular and metabolic oestrogenic signalling. Br. J. Pharmacol. 2011, 163, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Qin, S.; Carrasco, G.A.; Dai, Y.; Filardo, E.J.; Prossnitz, E.R.; Battaglia, G.; Doncarlos, L.L.; Muma, N.A. Extra-nuclear estrogen receptor GPR30 regulates serotonin function in rat hypothalamus. Neuroscience 2009, 158, 1599–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echimento, A.; Esirianni, R.; Ecasaburi, I.; Epezzi, V. Role of estrogen receptors and g protein-coupled estrogen receptor in regulation of hypothalamus–pituitary–testis axis and spermatogenesis. Front. Endocrinol. 2014, 5, 1. [Google Scholar] [CrossRef]
- Isensee, J.; Meoli, L.; Zazzu, V.; Nabzdyk, C.; Witt, H.; Soewarto, D.; Effertz, K.; Fuchs, H.; Gailus-Durner, V.; Busch, D.; et al. Expression pattern of g protein-coupled receptor 30 in lacz reporter mice. Endocrinology 2009, 150, 1722–1730. [Google Scholar] [CrossRef] [Green Version]
- Martensson, U.E.A.; Salehi, A.; Windahl, S.; Gomez, M.F.; Swärd, K.; Daszkiewicz-Nilsson, J.; Wendt, A.; Andersson, N.; Hellstrand, P.; Grände, P.-O.; et al. Deletion of the G protein-coupled receptor 30 impairs glucose tolerance, reduces bone growth, increases blood pressure, and eliminates estradiol-stimulated insulin release in female mice. Endocrinology 2009, 150, 687–698. [Google Scholar] [CrossRef]
- Olde, B.; Leeb-Lundberg, L.F. GPR30/GPER1: Searching for a role in estrogen physiology. Trends Endocrinol. Metab. 2009, 20, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Carreau, S.; Bouraima-Lelong, H.; Delalande, C. Estrogens–new players in spermatogenesis. Reprod. Biol. 2011, 11, 174–193. [Google Scholar] [CrossRef]
- Carreau, S.; Bouraima-Lelong, H.; Delalande, C. Estrogen, a female hormone involved in spermatogenesis. Adv. Med. Sci. 2012, 57, 31–36. [Google Scholar] [CrossRef]
- Wang, H.; Alencar, A.; Lin, M.; Sun, X.; Sudo, R.T.; Zapata-Sudo, G.; Lowe, D.A.; Groban, L. Activation of GPR30 improves exercise capacity and skeletal muscle strength in senescent female Fischer344 × Brown Norway rats. Biochem. Biophys. Res. Commun. 2016, 475, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLeon, C.; Wang, D.Q.-H.; Arnatt, C.K. G Protein-Coupled Estrogen Receptor, GPER1, Offers a Novel Target for the Treatment of Digestive Diseases. Front. Endocrinol. 2020, 11, 578536. [Google Scholar] [CrossRef] [PubMed]
- Nadal, A.; Alonso-Magdalena, P.; Soriano, S.; Ripoll, C.; Fuentes, E.; Quesada, I.; Ropero, A.B. Role of estrogen receptors alpha, beta and GPER1/GPR30 in pancreatic beta-cells. Front. Biosci. 2011, 16, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, E.; Meyer, M.; Schurr, U.; Bhattacharya, I.; Minotti, R.; Nguyen, H.H.; Heigl, A.; Lachat, M.; Genoni, M.; Barton, M. Differential effects of 17β-estradiol on function and expression of estrogen receptor α, estrogen receptor β, and gpr30 in arteries and veins of patients with atherosclerosis. Hypertension 2007, 49, 1358–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohar, E.Y.; Daugherty, E.M.; Aceves, J.O.; Sedaka, R.; Obi, I.E.; Allan, J.M.; Soliman, R.H.; Jin, C.; De Miguel, C.; Lindsey, S.; et al. Evidence for G-Protein–Coupled estrogen receptor as a pronatriuretic factor. J. Am. Heart Assoc. 2020, 9, e015110. [Google Scholar] [CrossRef]
- Meyer, M.R.; Amann, K.; Field, A.S.; Hu, C.; Hathaway, H.J.; Kanagy, N.L.; Walker, M.K.; Barton, M.; Prossnitz, E.R. Deletion of G Protein–Coupled estrogen receptor increases endothelial vasoconstriction. Hypertension 2012, 59, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Takada, Y.; Kato, C.; Kondo, S.; Korenaga, R.; Ando, J. Cloning of cDNAs encoding g protein-coupled receptor expressed in human endothelial cells exposed to fluid shear stress. Biochem. Biophys. Res. Commun. 1997, 240, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Holm, A.; Baldetorp, B.; Olde, B.; Leeb-Lundberg, L.F.; Nilsson, B.-O. The GPER1 agonist g-1 attenuates endothelial cell proliferation by inhibiting dna synthesis and accumulating cells in the s and g2 phases of the cell cycle. J. Vasc. Res. 2011, 48, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Haas, E.; Bhattacharya, I.; Brailoiu, E.; Damjanović, M.; Brailoiu, G.C.; Gao, X.; Mueller-Guerre, L.; Marjon, N.A.; Gut, A.; Minotti, R.; et al. Regulatory Role of G Protein–Coupled Estrogen Receptor for Vascular Function and Obesity. Circ. Res. 2009, 104, 288–291. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wang, H.; Lin, M.; Groban, L. GPR30 decreases cardiac chymase/angiotensin II by inhibiting local mast cell number. Biochem. Biophys. Res. Commun. 2015, 459, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffari, S.; Nabi, F.N.; Sugiyama, M.G.; Lee, W.L. Estrogen Inhibits LDL (Low-Density Lipoprotein) Transcytosis by Human Coronary Artery Endothelial Cells via GPER (G-Protein–Coupled Estrogen Receptor) and SR-BI (Scavenger Receptor Class B Type 1). Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2283–2294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yu, X.; Szynkarski, C.K.; Meng, C.; Zhou, B.; Barhoumi, R.; White, R.E.; Heaps, C.L.; Stallone, J.N.; Han, G. Activation of GPER Induces Differentiation and Inhibition of Coronary Artery Smooth Muscle Cell Proliferation. PLoS ONE 2013, 8, e64771. [Google Scholar] [CrossRef]
- Deschamps, A.; Murphy, E. Activation of a novel estrogen receptor, GPER, is cardioprotective in male and female rats. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1806–H1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Sun, X.; Chou, J.; Lin, M.; Ferrario, C.M.; Zapata-Sudo, G.; Groban, L. Cardiomyocyte-specific deletion of the G protein-coupled estrogen receptor (GPER) leads to left ventricular dysfunction and adverse remodeling: A sex-specific gene profiling analysis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2016, 1863, 1870–1882. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-B.; Dong, J.; Pang, Y.; LaRocca, J.; Hixon, M.; Thomas, P.; Filardo, E.J. Anatomical location and redistribution of G protein-coupled estrogen receptor-1 during the estrus cycle in mouse kidney and specific binding to estrogens but not aldosterone. Mol. Cell. Endocrinol. 2013, 382, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, S.H.; Yamaleyeva, L.M.; Brosnihan, K.B.; Gallagher, P.E.; Chappell, M.C. Estrogen Receptor GPR30 Reduces Oxidative Stress and Proteinuria in the Salt-Sensitive Female mRen2.Lewis Rat. Hypertension 2011, 58, 665–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Han, Z.; Zhang, Y.; Zhou, Y.; Feng, Z.; Chen, L.; Li, X.; Li, L.; Si, J.-Q. G protein-coupled estrogen receptor activation improves contractile and diastolic functions in rat renal interlobular artery to protect against renal ischemia reperfusion injury. Biomed. Pharmacother. 2019, 112, 108666. [Google Scholar] [CrossRef]
- Li, Y.-C.; Ding, X.-S.; Li, H.-M.; Zhang, Y.; Bao, J. Role of G protein-coupled estrogen receptor 1 in modulating transforming growth factor-β stimulated mesangial cell extracellular matrix synthesis and migration. Mol. Cell. Endocrinol. 2014, 391, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Hutson, D.D.; Gurrala, R.; Ogola, B.O.; Zimmerman, M.A.; Mostany, R.; Satou, R.; Lindsey, S.H. Estrogen receptor profiles across tissues from male and female Rattus norvegicus. Biol. Sex Differ. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurrala, R.; Kilanowski-Doroh, I.M.; Hutson, D.D.; Ogola, B.O.; Zimmerman, M.A.; Katakam, P.V.G.; Satou, R.; Mostany, R.; Lindsey, S.H. Alterations in the estrogen receptor profile of cardiovascular tissues during aging. GeroScience 2021, 43, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Bologa, C.G.; Revankar, C.M.; Young, S.M.; Edwards, B.S.; Arterburn, J.B.; Kiselyov, A.S.; Parker, M.A.; Tkachenko, S.E.; Savchuck, N.P.; Sklar, L.A.; et al. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat. Chem. Biol. 2006, 2, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Le May, C.; Wong, W.P.; Ward, R.D.; Clegg, D.J.; Marcelli, M.; Korach, K.S.; Mauvais-Jarvis, F. Importance of extranuclear estrogen receptor-α and membrane g protein–coupled estrogen receptor in pancreatic islet survival. Diabetes 2009, 58, 2292–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Dehghani, B.; Li, Y.; Kaler, L.J.; Proctor, T.M.; Vandenbark, A.A.; Offner, H. Membrane estrogen receptor regulates experimental autoimmune encephalomyelitis through up-regulation of programmed death 1. J. Immunol. 2009, 182, 3294–3303. [Google Scholar] [CrossRef] [PubMed]
- Albanito, L.; Madeo, A.; Lappano, R.; Vivacqua, A.; Rago, V.; Carpino, A.; Oprea, T.; Prossnitz, E.; Musti, A.M.; Andò, S.; et al. G Protein–Coupled receptor 30 (gpr30) mediates gene expression changes and growth response to 17β-estradiol and selective gpr30 ligand g-1 in ovarian cancer cells. Cancer Res. 2007, 67, 1859–1866. [Google Scholar] [CrossRef] [Green Version]
- Di Mattia, R.A.; Mariángelo, J.I.; Blanco, P.G.; De Giusti, C.J.; Portiansky, E.L.; Mundiña-Weilenmann, C.; Aiello, E.A.; Orlowski, A. The activation of the G protein-coupled estrogen receptor (GPER) prevents and regresses cardiac hypertrophy. Life Sci. 2020, 242, 117211. [Google Scholar] [CrossRef] [PubMed]
- Groban, L.; Tran, Q.-K.; Ferrario, C.M.; Sun, X.; Cheng, C.P.; Kitzman, D.W.; Wang, H.; Lindsey, S. Female Heart Health: Is GPER the Missing Link? Front. Endocrinol. 2019, 10, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, M.; Burai, R.; Ramesh, C.; Petrie, W.K.; Alcon, S.N.; Nayak, T.; Bologa, C.; Leitao, A.; Brailoiu, E.; Deliu, E.; et al. In vivo effects of a GPR30 antagonist. Nat. Chem. Biol. 2009, 5, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, M.K.; Field, A.S.; Burai, R.; Ramesh, C.; Petrie, W.K.; Bologa, C.; Oprea, T.; Yamaguchi, Y.; Hayashi, S.-I.; Sklar, L.A.; et al. Identification of a GPER/GPR30 antagonist with improved estrogen receptor counterselectivity. J. Steroid Biochem. Mol. Biol. 2011, 127, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fountain, J.H.; Lappin, S.L. Physiology, Renin Angiotensin System; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Hsueh, W.A. Potential effects of renin activation on the regulation of renin production. Am. J. Physiol. Renal Physiol. 1984, 247, F205–F212. [Google Scholar] [CrossRef]
- Fischer, M.; Baessler, A.; Schunkert, H. Renin angiotensin system and gender differences in the cardiovascular system. Cardiovasc. Res. 2002, 53, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Komukai, K.; Mochizuki, S.; Yoshimura, M. Gender and the renin-angiotensin-aldosterone system. Fundam. Clin. Pharmacol. 2010, 24, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Pendergrass, K.D.; Pirro, N.T.; Westwood, B.M.; Ferrario, C.M.; Brosnihan, K.B.; Chappell, M.C. Sex differences in circulating and renal angiotensins of hypertensive mRen().Lewis but not normotensive Lewis rats. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H10–H20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapater, P.; Novalbos, J.; Gallego-Sandín, S.; Hernández, F.T.; Abad-Santos, F. Gender Differences in Angiotensin-Converting Enzyme (ACE) Activity and Inhibition by Enalaprilat in Healthy Volunteers. J. Cardiovasc. Pharmacol. 2004, 43, 737–744. [Google Scholar] [CrossRef]
- Miller, J.A.; Anacta, L.A.; Cattran, D.C. Impact of gender on the renal response to angiotensin II. Kidney Int. 1999, 55, 278–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichikawa, J.; Sumino, H.; Ichikawa, S.; Ozaki, M. Different effects of transdermal and oral hormone replacement therapy on the renin-angiotensin system, plasma bradykinin level, and blood pressure of normotensive postmenopausal women. Am. J. Hypertens. 2006, 19, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimian, T.; He, Y.; Schiffrin, E.L.; Touyz, R.M. Differential regulation of thioredoxin and NAD(P)H oxidase by angiotensin II in male and female mice. J. Hypertens. 2007, 25, 1263–1271. [Google Scholar] [CrossRef]
- Xue, B.; Pamidimukkala, J.; Hay, M. Sex differences in the development of angiotensin II-induced hypertension in conscious mice. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H2177–H2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartori-Valinotti, J.C.; Iliescu, R.; Yanes, L.L.; Dorsett-Martin, W.; Reckelhoff, J.F. Sex Differences in the Pressor Response to Angiotensin II When the Endogenous Renin-Angiotensin System Is Blocked. Hypertension 2008, 51, 1170–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Antonialli, M.M.; Tostes, R.; Fernandes, L.; Fior-Chadi, D.R.; Akamine, E.H.; Carvalho, M.H.C.; Fortes, Z.B.; Nigro, D. A lower ratio of AT1/AT2 receptors of angiotensin II is found in female than in male spontaneously hypertensive rats. Cardiovasc. Res. 2004, 62, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Takagi, G.; Asai, K.; Resuello, R.G.; Natividad, F.F.; Vatner, D.E.; Vatner, S.F.; Lakatta, E.G. Aging Increases Aortic MMP-2 Activity and Angiotensin II in Nonhuman Primates. Hypertension 2003, 41, 1308–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Jiang, L.; Monticone, R.E.; Lakatta, E.G. Proinflammation: The key to arterial aging. Trends Endocrinol. Metab. 2013, 25, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, J.; Walker, S.; Dworakowski, R.; Lakatta, E.; Shah, A. Involvement of NADPH oxidase in age-associated cardiac remodeling. J. Mol. Cell. Cardiol. 2010, 48, 765–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, J.; Spinetti, G.; Jiang, L.-Q.; Monticone, R.; Zhao, D.; Cheng, L.; Krawczyk, M.; Talan, M.; Pintus, G.; et al. Angiotensin II activates matrix metalloproteinase type ii and mimics age-associated carotid arterial remodeling in young rats. Am. J. Pathol. 2005, 167, 1429–1442. [Google Scholar] [CrossRef] [Green Version]
- Spinetti, G.; Wang, M.; Monticone, R.; Zhang, J.; Zhao, D.; Lakatta, E. Rat aortic MCP-1 and its receptor CCR2 increase with age and alter vascular smooth muscle cell function. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, J.; Jiang, L.-Q.; Spinetti, G.; Pintus, G.; Monticone, R.; Kolodgie, F.D.; Virmani, R.; Lakatta, E.G. Proinflammatory Profile Within the Grossly Normal Aged Human Aortic Wall. Hypertension 2007, 50, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, S.; Cassis, P.; Benigni, A. Aging and the renin-angiotensin system. Hypertension 2012, 60, 878–883. [Google Scholar] [CrossRef]
- Corman, B.; Michel, J.B. Renin-angiotensin system, converting-enzyme inhibition and kidney function in aging female rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1986, 251, R450–R455. [Google Scholar] [CrossRef]
- Belmin, J.; Lévy, B.I.; Michel, J.-B. Changes in the Renin-Angiotensin-Aldosterone Axis in Later Life. Drugs Aging 1994, 5, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, P.; Beretta-Piccoli, C.; Ziegler, W.H.; Keusch, G.; Glück, Z.; Reubi, F.C. Age versus urinary sodium for judging renin, aldosterone, and catecholamine levels: Studies in normal subjects and patients with essential hypertension. Kidney Int. 1978, 14, 619–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noth, R.H.; Lassman, M.N.; Tan, S.Y.; Fernandez-Cruz, A., Jr.; Mulrow, P.J. Age and the Renin-Aldosterone System. Arch. Intern. Med. 1977, 137, 1414–1417. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, S.H.; Cohen, J.A.; Brosnihan, K.B.; Gallagher, P.E.; Chappell, M.C. Chronic treatment with the g protein-coupled receptor 30 agonist g-1 decreases blood pressure in ovariectomized mren2.lewis rats. Endocrinology 2009, 150, 3753–3758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossmann, C.; Benesic, A.; Krug, A.W.; Freudinger, R.; Mildenberger, S.; Gassner, B.; Gekle, M. Human mineralocorticoid receptor expression renders cells responsive for nongenotropic aldosterone actions. Mol. Endocrinol. 2005, 19, 1697–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros, R.; Ding, Q.; Liu, B.; Chorazyczewski, J.; Feldman, R.D. Aldosterone mediates its rapid effects in vascular endothelial cells through GPER activation. Am. J. Physiol. Cell Physiol. 2013, 304, C532–C540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigiracciolo, D.C.; Scarpelli, A.; Lappano, R.; Pisano, A.; Santolla, M.F.; Avino, S.; De Marco, P.; Bussolati, B.; Maggiolini, M.; De Francesco, E.M. GPER is involved in the stimulatory effects of aldosterone in breast cancer cells and breast tumor-derived endothelial cells. Oncotarget 2016, 7, 94–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, N.S.; Cau, S.; Silva, M.; Manzato, C.P.; Mestriner, F.L.A.C.; Matsumoto, T.; Carneiro, F.; Tostes, R.C. Diabetes impairs the vascular effects of aldosterone mediated by G protein-coupled estrogen receptor activation. Front. Pharmacol. 2015, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehling, M. Rapid actions of aldosterone revisited: Receptors in the limelight. J. Steroid Biochem. Mol. Biol. 2018, 176, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Poulsen, S.B.; Wu, Q.; Esteva-Font, C.; Olesen, E.T.B.; Peng, L.; Olde, B.; Leeb-Lundberg, L.M.F.; Pisitkun, T.; Rieg, T.; et al. Rapid Aldosterone-Mediated Signaling in the DCT Increases Activity of the Thiazide-Sensitive NaCl Cotransporter. J. Am. Soc. Nephrol. 2019, 30, 1454–1470. [Google Scholar] [CrossRef]
- Eiam-Ong, S.; Chaipipat, M.; Manotham, K. Aldosterone rapidly activates p-PKC delta and GPR30 but suppresses p-PKC epsilon protein levels in rat kidney. Endocr. Regul. 2019, 53, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros, R.; Ding, Q.; Sklar, L.A.; Prossnitz, E.; Arterburn, J.B.; Chorazyczewski, J.; Feldman, R.D. GPR30 expression is required for the mineralocorticoid receptor–independent rapid vascular effects of aldosterone. Hypertension 2011, 57, 442–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, R.D.; Gros, R. Unraveling the mechanisms underlying the rapid vascular effects of steroids: Sorting out the receptors and the pathways. Br. J. Pharmacol. 2011, 163, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frommer, K.W.; Muller-Ladner, U. Expression and function of ETA and ETB receptors in SSc. Rheumatology 2008, 47, v27–v28. [Google Scholar] [CrossRef] [Green Version]
- Barton, M.; Yanagisawa, M. Endothelin: 20 years from discovery to therapyThis article is one of a selection of papers published in the special issue (part 2 of 2) on Forefronts in Endothelin. Can. J. Physiol. Pharmacol. 2008, 86, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Wynne, B.M.; Chiao, C.-W.; Webb, R.C. Vascular smooth muscle cell signaling mechanisms for contraction to angiotensin II and endothelin-1. J. Am. Soc. Hypertens. 2009, 3, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, J.; Goresky, C.A.; Fournier, A. Pulmonary clearance of circulating endothelin-1 in dogs in vivo: Exclusive role of ETBreceptors. J. Appl. Physiol. 1996, 81, 1510–1515. [Google Scholar] [CrossRef]
- Fukuroda, T.; Fujikawa, T.; Ozaki, S.; Ishikawa, K.; Yano, M.; Nishikibe, M. Clearance of circulating endothelin-1 by etb receptors in rats. Biochem. Biophys. Res. Commun. 1994, 199, 1461–1465. [Google Scholar] [CrossRef]
- Barton, M.; Cosentino, F.; Brandes, R.P.; Moreau, P.; Shaw, S.; Lüscher, T.F. Anatomic heterogeneity of vascular aging: Role of nitric oxide and endothelin. Hypertension 1997, 30, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Kumazaki, T.; Wadhwa, R.; Kaul, S.C.; Mitsui, Y. Expression of endothelin, fibronectin, and mortalin as aging and mortality markers. Exp. Gerontol. 1997, 32, 95–103. [Google Scholar] [CrossRef]
- Goettsch, W.; Lattmann, T.; Amann, K.; Szibor, M.; Morawietz, H.; Münter, K.; Müller, S.P.; Shaw, S.; Barton, M. Increased expression of endothelin-1 and inducible nitric oxide synthase isoform II in aging arteries in vivo: Implications for atherosclerosis. Biochem. Biophys. Res. Commun. 2001, 280, 908–913. [Google Scholar] [CrossRef]
- Lekontseva, O.; Chakrabarti, S.; Davidge, S.T. Endothelin in the female vasculature: A role in aging? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R509–R516. [Google Scholar] [CrossRef] [PubMed]
- Moreau, K.L.; Hildreth, K.L.; Meditz, A.L.; Deane, K.D.; Kohrt, W.M. endothelial function is impaired across the stages of the menopause transition in healthy women. J. Clin. Endocrinol. Metab. 2012, 97, 4692–4700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polderman, K.H.; Stehouwer, C.D.A.; Van Kamp, G.J.; Dekker, G.A.; Verheugt, F.W.A.; Gooren, L.J.G. influence of sex hormones on plasma endothelin levels. Ann. Intern. Med. 1993, 118, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, T.; Yanagisawa, M.; Iida, K.; Ajisaka, R.; Suzuki, N.; Fujino, M.; Goto, K.; Masaki, T.; Sugishita, Y. Age- and sex-related variation of plasma endothelin-1 concentration in normal and hypertensive subjects. Am. Heart J. 1992, 123, 1092–1093. [Google Scholar] [CrossRef]
- Carbonne, B.; Mignot, T.; Tsatsaris, V.; Ferré, F. Changes in plasma and amniotic fluid endothelin levels during pregnancy: Facts or artefacts? Eur. J. Obstet. Gynecol. Reprod. Biol. 1998, 76, 15–19. [Google Scholar] [CrossRef]
- Lygnos, M.C.; Pappa, K.I.; Papadaki, H.A.; Relakis, C.; Koumantakis, E.; Anagnou, N.P.; Eliopoulos, G.D. Changes in maternal plasma levels of VEGF, bFGF, TGF-beta1, ET-1 and sKL during uncomplicated pregnancy, hypertensive pregnancy and gestational diabetes. In Vivo 2006, 20, 157–163. [Google Scholar] [PubMed]
- Pappa, K.I.; Anagnou, N.P.; Daskalakis, G.; Ioannides, I.; Angelidou, K.; Antsaklis, A. Maternal and fetal circulating sKL and ET-1 levels as function of normal labor at term. J. Matern. Fetal Neonatal Med. 2010, 24, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Polderman, K.H.; Stehouwer, C.D.; van Kamp, G.J.; Schalkwijk, C.G.; Gooren, L.J. Modulation of plasma endothelin levels by the menstrual cycle. Metabolism 2000, 49, 648–650. [Google Scholar] [CrossRef]
- Sebzda, K.N.; Kuczmarski, A.V.; Pohlig, R.T.; Lennon, S.L.; Edwards, D.G.; Wenner, M.M. Ovarian hormones modulate endothelin-1 receptor responses in young women. Microcirculation 2018, 25, e12490. [Google Scholar] [CrossRef] [Green Version]
- Wenner, M.M.; Sebzda, K.N.; Kuczmarski, A.V.; Pohlig, R.T.; Edwards, D.G. ETBreceptor contribution to vascular dysfunction in postmenopausal women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R51–R57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Ding, T.; Liu, H.; Jian, L. GPER was associated with hypertension in post-menopausal women. Open Med. 2018, 13, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Baretella, O.; Prossnitz, E.; Barton, M. Dilation of epicardial coronary arteries by the g protein-coupled estrogen receptor agonists g-1 and ici 182,780. Pharmacology 2010, 86, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.; Field, A.S.; Kanagy, N.L.; Barton, M.; Prossnitz, E.R. GPER regulates endothelin-dependent vascular tone and intracellular calcium. Life Sci. 2012, 91, 623–627. [Google Scholar] [CrossRef] [Green Version]
- Holm, A.; Hellstrand, P.; Olde, B.; Svensson, D.; Leeb-Lundberg, L.F.; Nilsson, B.-O. The G Protein-Coupled Estrogen Receptor 1 (GPER1/GPR30) Agonist G-1 Regulates Vascular Smooth Muscle Cell Ca2+ Handling. J. Vasc. Res. 2013, 50, 421–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Fredette, N.C.; Daniel, C.; Sharma, G.; Amann, K.; Arterburn, J.B.; Barton, M.; Prossnitz, E.R. Obligatory role for GPER in cardiovascular aging and disease. Sci. Signal. 2016, 9, ra105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Fredette, N.C.; Sharma, G.; Barton, M.; Prossnitz, E.R. GPER is required for the age-dependent upregulation of the myocardial endothelin system. Life Sci. 2016, 159, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, D.; Pollock, D.M. Contribution of endothelin a receptors in endothelin 1–dependent natriuresis in female rats. Hypertension 2009, 53, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Bagnall, A.; Stricklett, P.K.; Webb, D.; Kotelevtsev, Y.; Kohan, D.E. Combined knockout of collecting duct endothelin A and B receptors causes hypertension and sodium retention. Am. J. Physiol. Renal Physiol. 2008, 295, F1635–F1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boesen, E.I.; Pollock, D.M. Cooperative role of ETAand ETBreceptors in mediating the diuretic response to intramedullary hyperosmotic NaCl infusion. Am. J. Physiol. Renal Physiol. 2010, 299, F1424–F1432. [Google Scholar] [CrossRef] [Green Version]
- Hyndman, K.; Pollock, J.S. Nitric oxide and the A and B of endothelin of sodium homeostasis. Curr. Opin. Nephrol. Hypertens. 2013, 22, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Speed, J.S.; Hyndman, K.A.; O’Connor, P.M.; Pollock, D.M. Sex differences in ET-1 receptor expression and Ca2+ signaling in the IMCD. Am. J. Physiol. Renal Physiol. 2013, 305, F1099–F1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohar, E.Y.; Pollock, D.M. Functional Interaction of Endothelin Receptors in Mediating Natriuresis Evoked by G Protein–Coupled Estrogen Receptor 1. J. Pharmacol. Exp. Ther. 2020, 376, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, J.; Martasek, P.; McMillan, K.; Salerno, J.; Liu, Q.; Gross, S.; Masters, B. Modular Structure of Neuronal Nitric Oxide Synthase: Localization of the Arginine Binding Site and Modulation by Pterin. Biochem. Biophys. Res. Commun. 1995, 210, 288–294. [Google Scholar] [CrossRef]
- Crane, B.R.; Arvai, A.S.; Ghosh, D.K.; Wu, C.; Getzoff, E.D.; Stuehr, D.J.; Tainer, J.A. Structure of Nitric Oxide Synthase Oxygenase Dimer with Pterin and Substrate. Science 1998, 279, 2121–2126. [Google Scholar] [CrossRef] [PubMed]
- Farah, C.; Michel, L.Y.; Balligand, J.-L. Nitric oxide signalling in cardiovascular health and disease. Nat. Rev. Cardiol. 2018, 15, 292–316. [Google Scholar] [CrossRef] [PubMed]
- Carlström, M. Nitric oxide signalling in kidney regulation and cardiometabolic health. Nat. Rev. Nephrol. 2021, 17, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.L.N.S.; Sivakumar, V.; Bitla, A.; Reddy, Y.; Kiranmayi, V.; Krishna, G. Nitric oxide status in patients with chronic kidney disease. Indian J. Nephrol. 2015, 25, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.-L.; Ritter, J.K. Role of Nitric Oxide in the Cardiovascular and Renal Systems. Int. J. Mol. Sci. 2018, 19, 2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauser, K.; Rubanyi, G.M. Gender difference in bioassayable endothelium-derived nitric oxide from isolated rat aortae. Am. J. Physiol. Heart Circ. Physiol. 1994, 267, H2311–H2317. [Google Scholar] [CrossRef] [PubMed]
- Kauser, K.; Rubanyi, G.M. Gender Difference in Endothelial Dysfunction in the Aorta of Spontaneously Hypertensive Rats. Hypertension 1995, 25, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F.; Hennington, B.S.; Moore, A.G.; Blanchard, E.J.; Cameron, J. Gender Differences in the Renal Nitric Oxide (NO) System. Am. J. Hypertens. 1998, 11, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Neugarten, J.; Ding, Q.; Friedman, A.; Lei, J.; Silbiger, S. Sex hormones and renal nitric oxide synthases. J. Am. Soc. Nephrol. 1997, 8, 1240–1246. [Google Scholar] [CrossRef]
- Hill, C.; Lateef, A.M.; Engels, K.; Samsell, L.; Baylis, C. Basal and stimulated nitric oxide in control of kidney function in the aging rat. Am. J. Physiol. Content 1997, 272, R1747–R1753. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F.; Kellum, J.A.; Blanchard, E.; Bacon, E.E.; Wesley, A.J.; Kruckeberg, W.C. Changes in nitric oxide precursor, L-arginine, and metabolites, nitrate and nitrite, with aging. Life Sci. 1994, 55, 1895–1902. [Google Scholar] [CrossRef]
- Thomas, S.E.; Anderson, S.; Gordon, K.L.; Oyama, T.T.; Shankland, S.J.; Johnson, R.J. Tubulointerstitial disease in aging: Evidence for underlying peritubular capillary damage, a potential role for renal ischemia. J. Am. Soc. Nephrol. 1998, 9, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Erdely, A.; Greenfeld, Z.; Wagner, L.; Baylis, C. Sexual dimorphism in the aging kidney: Effects on injury and nitric oxide system. Kidney Int. 2003, 63, 1021–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielstein, J.T.; Bode-Böger, S.; Frölich, J.C.; Ritz, E.; Haller, H.; Fliser, D. Asymmetric Dimethylarginine, Blood Pressure, and Renal Perfusion in Elderly Subjects. Circulation 2003, 107, 1891–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torregrossa, A.C.; Aranke, M.; Bryan, N.S. Nitric oxide and geriatrics: Implications in diagnostics and treatment of the elderly. J. Geriatr. Cardiol. 2011, 8, 230–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taddei, S.; Virdis, A.; Ghiadoni, L.; Salvetti, G.; Bernini, G.; Magagna, A.; Salvetti, A. Age-Related Reduction of NO Availability and Oxidative Stress in Humans. Hypertension 2001, 38, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, S.H.; Liu, L.; Chappell, M.C. Vasodilation by GPER in mesenteric arteries involves both endothelial nitric oxide and smooth muscle cAMP signaling. Steroids 2013, 81, 99–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, N.T.B.; Rouver, W.D.N.; Freitas-Lima, L.C.; Vieira-Alves, I.; Lemos, V.S.; dos Santos, R.L. Sex Differences in the Vasodilation Mediated by G Protein-Coupled Estrogen Receptor (GPER) in Hypertensive Rats. Front. Physiol. 2021, 12, 1078. [Google Scholar] [CrossRef]
- Fredette, N.C.; Meyer, M.R.; Prossnitz, E.R. Role of GPER in estrogen-dependent nitric oxide formation and vasodilation. J. Steroid Biochem. Mol. Biol. 2018, 176, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, P.; Aires, R.D.; Lemos, V.S.; Bissoli, N.S.; dos Santos, R.L. GPER agonist dilates mesenteric arteries via PI3K-Akt-eNOS and potassium channels in both sexes. Life Sci. 2017, 183, 21–27. [Google Scholar] [CrossRef]
- Tropea, T.; De Francesco, E.M.; Rigiracciolo, D.C.; Maggiolini, M.; Wareing, M.; Osol, G.; Mandalà, M. Pregnancy Augments G Protein Estrogen Receptor (GPER) Induced Vasodilation in Rat Uterine Arteries via the Nitric Oxide-cGMP Signaling Pathway. PLoS ONE 2015, 10, e0141997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsey, S.H.; da Silva, A.S.; Silva, M.S.; Chappell, M.C. Reduced vasorelaxation to estradiol and G-1 in aged female and adult male rats is associated with GPR30 downregulation. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E113–E118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Fredette, N.C.; Howard, T.A.; Hu, C.; Ramesh, C.; Daniel, C.; Amann, K.; Arterburn, J.B.; Barton, M.; Prossnitz, E.R. G Protein-coupled Estrogen Receptor Protects from Atherosclerosis. Sci. Rep. 2014, 4, 7564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Prossnitz, E.R.; Barton, M. GPER/GPR30 and Regulation of Vascular Tone and Blood Pressure. Immunol. Endocr. Metab. Agents Med. Chem. 2011, 11, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsey, S.H.; Carver, K.A.; Prossnitz, E.R.; Chappell, M.C. Vasodilation in Response to the GPR30 Agonist G-1 is Not Different from Estradiol in the mRen2.Lewis Female Rat. J. Cardiovasc. Pharmacol. 2011, 57, 598–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, B.R.S.; Miller, A.A.; Sobey, C.G. Endothelium-dependent relaxation by G protein-coupled receptor 30 agonists in rat carotid arteries. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1055–H1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Rosemann, T.; Barton, M.; Prossnitz, E.R. GPER Mediates Functional Endothelial Aging in Renal Arteries. Pharmacology 2017, 100, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, N.S.; Temsah, R.M.; Netticadan, T. Role of oxidative stress in cardiovascular diseases. J. Hypertens. 2000, 18, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Tomsa, A.M.; Alexa, A.L.; Junie, M.L.; Rachisan, A.L.; Ciumarnean, L. Oxidative stress as a potential target in acute kidney injury. PeerJ 2019, 7, e8046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamboa, J.L.; Billings, F.T.; Bojanowski, M.T.; Gilliam, L.A.; Yu, C.; Roshanravan, B.; Roberts, L.J., 2nd; Himmelfarb, J.; Ikizler, T.A.; Brown, N.J. Mitochondrial dysfunction and oxidative stress in patients with chronic kidney disease. Physiol. Rep. 2016, 4, e12780. [Google Scholar] [CrossRef] [PubMed]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender difference in oxidative stress: A new look at the mechanisms for cardiovascular diseases. J. Cell. Mol. Med. 2016, 21, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Barp, J.; Araújo, A.; Fernandes, T.; Rigatto, K.; Llesuy, S.; Belló-Klein, A.; Singal, P. Myocardial antioxidant and oxidative stress changes due to sex hormones. Braz. J. Med. Biol. Res. 2002, 35, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Ide, T.; Tsutsui, H.; Ohashi, N.; Hayashidani, S.; Suematsu, N.; Tsuchihashi, M.; Tamai, H.; Takeshita, A. Greater Oxidative Stress in Healthy Young Men Compared with Premenopausal Women. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matarrese, P.; Colasanti, T.; Ascione, B.; Margutti, P.; Franconi, F.; Alessandri, C.; Conti, F.; Riccieri, V.; Rosano, G.; Ortona, E.; et al. Gender Disparity in Susceptibility to Oxidative Stress and Apoptosis Induced by Autoantibodies Specific to RLIP76 in Vascular Cells. Antioxid. Redox Signal. 2011, 15, 2825–2836. [Google Scholar] [CrossRef] [PubMed]
- Dantas, A.P.V.; Franco, M.D.C.P.; Silva-Antonialli, M.M.; Tostes, R.C.A.; Fortes, Z.B.; Nigro, D.; Carvalho, M.H.C. Gender differences in superoxide generation in microvessels of hypertensive rats: Role of NAD(P)H-oxidase. Cardiovasc. Res. 2004, 61, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, K.; Elmarakby, A.A.; El-Remessy, A.B.; Sullivan, J.C. Oxidative stress contributes to sex differences in angiotensin II-mediated hypertension in spontaneously hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R274–R282. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; DELLA-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Mehra, N.; Swarnakar, N. Role of Antioxidants for the Treatment of Cardiovascular Diseases: Challenges and Opportunities. Curr. Pharm. Des. 2015, 21, 4441–4455. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Kashyap, S.; Murphy, B.; Hutson, D.D.; Budish, R.A.; Trimmer, E.H.; Zimmerman, M.A.; Trask, A.J.; Miller, K.S.; Chappell, M.C.; et al. GPER activation ameliorates aortic remodeling induced by salt-sensitive hypertension. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H953–H961. [Google Scholar] [CrossRef] [PubMed]
- Bopassa, J.C.; Eghbali, M.; Toro, L.; Stefani, E. A novel estrogen receptor GPER inhibits mitochondria permeability transition pore opening and protects the heart against ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H16–H23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Jessup, J.A.; Lin, M.S.; Chagas, C.; Lindsey, S.H.; Groban, L. Activation of GPR30 attenuates diastolic dysfunction and left ventricle remodelling in oophorectomized mRen2.Lewis rats. Cardiovasc. Res. 2012, 94, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Sun, X.; Lin, M.S.; Ferrario, C.M.; Van Remmen, H.; Groban, L. G protein-coupled estrogen receptor (GPER) deficiency induces cardiac remodeling through oxidative stress. Transl. Res. 2018, 199, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Kurt, A.H.; Bozkus, F.; Uremis, N.; Uremis, M.M. The protective role of G protein-coupled estrogen receptor 1 (GPER-1) on methotrexate-induced nephrotoxicity in human renal epithelium cells. Ren. Fail. 2016, 38, 686–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-B.; Han, J.; Zhang, N.; Tian, Z.; Li, X.-B.; Zhao, M.-G. Neuroprotective effects of oestrogen against oxidative toxicity through activation of G-protein-coupled receptor 30 receptor. Clin. Exp. Pharmacol. Physiol. 2011, 38, 577–585. [Google Scholar] [CrossRef]
- De Francesco, E.M.; Rocca, C.; Scavello, F.; Amelio, D.; Pasqua, T.; Rigiracciolo, D.C.; Scarpelli, A.; Avino, S.; Cirillo, F.; Amodio, N.; et al. Protective Role of GPER Agonist G-1 on Cardiotoxicity Induced by Doxorubicin. J. Cell. Physiol. 2017, 232, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Oberg, B.P.; McMenamin, E.; Lucas, F.L.; McMonagle, E.; Morrow, J.; Ikizler, T.A.; Himmelfarb, J. Increased prevalence of oxidant stress and inflammation in patients with moderate to severe chronic kidney disease. Kidney Int. 2004, 65, 1009–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dounousi, E.; Papavasiliou, E.; Makedou, A.; Ioannou, K.; Katopodis, K.P.; Tselepis, A.; Siamopoulos, K.C.; Tsakiris, D. Oxidative Stress Is Progressively Enhanced with Advancing Stages of CKD. Am. J. Kidney Dis. 2006, 48, 752–760. [Google Scholar] [CrossRef] [Green Version]
- Himmelfarb, J.; McMonagle, E.; McMenamin, E. Plasma protein thiol oxidation and carbonyl formation in chronic renal failure. Kidney Int. 2000, 58, 2571–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geovanini, G.R.; Libby, P. Atherosclerosis and inflammation: Overview and updates. Clin. Sci. 2018, 132, 1243–1252. [Google Scholar] [CrossRef]
- Sorriento, D.; Iaccarino, G. Inflammation and Cardiovascular Diseases: The Most Recent Findings. Int. J. Mol. Sci. 2019, 20, 3879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikizler, T.A.; Wingard, R.L.; Harvell, J.; Shyr, Y.; Hakim, R.M. Association of morbidity with markers of nutrition and inflammation in chronic hemodialysis patients: A prospective study. Kidney Int. 1999, 55, 1945–1951. [Google Scholar] [CrossRef] [Green Version]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Liberale, L.; Montecucco, F.; Tardif, J.-C.; Libby, P.; Camici, G.G. Inflamm-ageing: The role of inflammation in age-dependent cardiovascular disease. Eur. Heart J. 2020, 41, 2974–2982. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Davidge, S.T. G-Protein Coupled Receptor 30 (GPR30): A Novel Regulator of Endothelial Inflammation. PLoS ONE 2012, 7, e52357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Fredette, N.C.; Barton, M.; Prossnitz, E.R. G protein-coupled estrogen receptor inhibits vascular prostanoid production and activity. J. Endocrinol. 2015, 227, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, G.; Hu, C.; Brigman, J.L.; Zhu, G.; Hathaway, H.J.; Prossnitz, E.R. GPER Deficiency in Male Mice Results in Insulin Resistance, Dyslipidemia, and a Proinflammatory State. Endocrinology 2013, 154, 4136–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Włodarczyk, M.; Sobolewska-Włodarczyk, A.; Cygankiewicz, A.I.; Jacenik, D.; Piechota-Polańczyk, A.; Stec-Michalska, K.; Krajewska, W.M.; Fichna, J.; Wiśniewska-Jarosińska, M. G Protein-Coupled Receptor 30 (GPR30) Expression Pattern in Inflammatory Bowel Disease Patients Suggests its Key Role in the Inflammatory Process. A Preliminary Study. J. Gastrointest. Liver Dis. 2017, 26, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Jacenik, D.; Zielińska, M.; Mokrowiecka, A.; Michlewska, S.; Małecka-Panas, E.; Kordek, R.; Fichna, J.; Krajewska, W.M. G protein-coupled estrogen receptor mediates anti-inflammatory action in Crohn’s disease. Sci. Rep. 2019, 9, 6749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caillon, A.; Mian, M.O.R.; Fraulob-Aquino, J.C.; Huo, K.-G.; Barhoumi, T.; Ouerd, S.; Sinnaeve, P.R.; Paradis, P.; Schiffrin, E.L. γδ T Cells Mediate Angiotensin II-Induced Hypertension and Vascular Injury. Circulation 2017, 135, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.D.; Limbird, L.E. GPER (GPR30): A Nongenomic Receptor (GPCR) for Steroid Hormones with Implications for Cardiovascular Disease and Cancer. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Notas, G.; Kampa, M.; Castanas, E. G Protein-Coupled Estrogen Receptor in Immune Cells and Its Role in Immune-Related Diseases. Front. Endocrinol. 2020, 11, 579420. [Google Scholar] [CrossRef] [PubMed]
- Brunsing, R.L.; Owens, K.S.; Prossnitz, E.R. The G Protein-coupled Estrogen Receptor (GPER) Agonist G-1 Expands the Regulatory T-cell Population Under TH17-polarizing Conditions. J. Immunother. 2013, 36, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunsing, R.L.; Prossnitz, E.R. Induction of interleukin-10 in the T helper type 17 effector population by the G protein coupled estrogen receptor (GPER) agonist G-1. Immunology 2011, 134, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Bowling, M.R.; Xing, D.; Kapadia, A.; Chen, Y.-F.; Szalai, A.J.; Oparil, S.; Hage, F.G. Estrogen Effects on Vascular Inflammation Are Age Dependent: Role of estrogen receptors. Arteriosclerosis, thrombosis, and vascular biology. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1477–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Sun, X.; Hodge, H.S.; Ferrario, C.M.; Groban, L. NLRP3 inhibition improves heart function in GPER knockout mice. Biochem. Biophys. Res. Commun. 2019, 514, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Pelekanou, V.; Kampa, M.; Kiagiadaki, F.; Deli, A.; Theodoropoulos, P.; Agrogiannis, G.; Patsouris, E.; Tsapis, A.; Castanas, E.; Notas, G. Estrogen anti-inflammatory activity on human monocytes is mediated through cross-talk between estrogen receptor ERα36 and GPR30/GPER1. J. Leukoc. Biol. 2015, 99, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Gohar, E.Y.; Almutlaq, R.N.; Daugherty, E.M.; Butt, M.K.; Jin, C.; Pollock, J.S.; Pollock, D.M.; De Miguel, C. Activation of G protein-coupled estrogen receptor 1 ameliorates proximal tubular injury and proteinuria in Dahl salt-sensitive female rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R297–R306. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Blood Pressure/Hypertension. Available online: https://www.who.int/data/gho/indicator-metadata-registry/imr-details/3155 (accessed on 16 January 2022).
- Arima, H.; Barzi, F.; Chalmers, J. Mortality patterns in hypertension. J. Hypertens. 2011, 29 (Suppl. 1), S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Ostchega, Y.; Fryar, C.D.; Nwankwo, T.; Nguyen, D.T. Hypertension Prevalence Among Adults Aged 18 and Over: United States, 2017–2018; NCHS Data Brief, No. 364; National Center for Health Statistics: Hyattsville, MD, USA, 2020. Available online: https://www.cdc.gov/nchs/products/databriefs/db364.htm (accessed on 10 January 2022).
- World Health Organization. Hypertension. Available online: https://www.who.int/news-room/fact-sheets/detail/hypertension (accessed on 20 January 2022).
- Feldman, R.D.; Gros, R.; Ding, Q.; Hussain, Y.; Ban, M.R.; McIntyre, A.D.; Hegele, R.A. A common hypofunctional genetic variant of GPER is associated with increased blood pressure in women. Br. J. Clin. Pharmacol. 2014, 78, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Debortoli, A.R.; Rouver, W.; Delgado, N.T.B.; Mengal, V.; Claudio, E.R.G.; Pernomian, L.; Bendhack, L.M.; Moysés, M.R.; Dos Santos, R.L. GPER modulates tone and coronary vascular reactivity in male and female rats. J. Mol. Endocrinol. 2017, 59, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Arefin, S.; Simoncini, T.; Wieland, R.; Hammarqvist, F.; Spina, S.; Goglia, L.; Kublickiene, K. Vasodilatory effects of the selective GPER agonist G-1 is maximal in arteries of postmenopausal women. Maturitas 2014, 78, 123–130. [Google Scholar] [CrossRef]
- Peixoto, P.; da Silva, J.F.; Aires, R.D.; Costa, E.D.; Lemos, V.S.; Bissoli, N.S.; dos Santos, R.L. Sex difference in GPER expression does not change vascular relaxation or reactive oxygen species generation in rat mesenteric resistance arteries. Life Sci. 2018, 211, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Dietrich, H.H.; Xiang, C.; Dacey, R.G., Jr. G Protein–Coupled Estrogen Receptor Agonist Improves Cerebral Microvascular Function After Hypoxia/Reoxygenation Injury in Male and Female Rats. Stroke 2013, 44, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Rojas, L.; Castro, L.A.R.; Ramírez, V.; Bautista-Pérez, R.; Rafael, C.; Castañeda-Bueno, M.; Meade, P.; Heros, P.D.L.; Arroyo-Garza, I.; Bernard, V.; et al. Ovarian hormones and prolactin increase renal NaCl cotransporter phosphorylation. Am. J. Physiol. Renal Physiol. 2015, 308, F799–F808. [Google Scholar] [CrossRef]
- Hofmeister, M.V.; Damkier, H.H.; Christensen, B.M.; Olde, B.; Leeb-Lundberg, L.M.F.; Fenton, R.A.; Praetorius, H.A.; Praetorius, J. 17β-Estradiol induces nongenomic effects in renal intercalated cells through G protein-coupled estrogen receptor 1. Am. J. Physiol. Renal Physiol. 2012, 302, F358–F368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, Y.; Ding, Q.; Connelly, P.; Brunt, J.H.; Ban, M.R.; McIntyre, A.D.; Huff, M.W.; Gros, R.; Hegele, R.A.; Feldman, R.D. G-Protein Estrogen Receptor as a Regulator of Low-Density Lipoprotein Cholesterol Metabolism: Cellular and population genetic studies. Arterioscler. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Murphy, S.L.; Kochanek, K.D.; Arias, E. Deaths: Final Data for 2019. Natl. Vital Stat. Rep. 2021, 70, 8. [Google Scholar]
- Center for Disease Control and Prevention. Know Your Risk for Heart Disease. Available online: https://www.cdc.gov/heartdisease/risk_factors.htm (accessed on 10 January 2022).
- Zhang, X.; Li, T.; Cheng, H.-J.; Wang, H.; Ferrario, C.M.; Groban, L.; Cheng, C.P. Chronic GPR30 agonist therapy causes restoration of normal cardiac functional performance in a male mouse model of progressive heart failure: Insights into cellular mechanisms. Life Sci. 2021, 285, 119955. [Google Scholar] [CrossRef]
- Li, W.-L.; Xiang, W.; Ping, Y. Activation of novel estrogen receptor GPER results in inhibition of cardiocyte apoptosis and cardioprotection. Mol. Med. Rep. 2015, 12, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Madungwe, N.; Junho, C.V.C.; Bopassa, J.C. Activation of G protein-coupled oestrogen receptor 1 at the onset of reperfusion protects the myocardium against ischemia/reperfusion injury by reducing mitochondrial dysfunction and mitophagy. Br. J. Pharmacol. 2017, 174, 4329–4344. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.E.; Singh, H.; Lu, R.; Olde, B.; Leeb-Lundberg, L.M.F.; Bopassa, J.C. G Protein-Coupled Estrogen Receptor 1 Mediates Acute Estrogen-Induced Cardioprotection via MEK/ERK/GSK-3β Pathway after Ischemia/Reperfusion. PLoS ONE 2015, 10, e0135988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weil, B.R.; Manukyan, M.C.; Herrmann, J.L.; Wang, Y.; Abarbanell, A.M.; Poynter, J.A.; Meldrum, D.R. Signaling via GPR30 protects the myocardium from ischemia/reperfusion injury. Surgery 2010, 148, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Jessup, J.A.; Lindsey, S.; Wang, H.; Chappell, M.C.; Groban, L. Attenuation of Salt-Induced Cardiac Remodeling and Diastolic Dysfunction by the GPER Agonist G-1 in Female mRen2.Lewis Rats. PLoS ONE 2010, 5, e15433. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.C.; Beyger, L.; Guchardi, J.; Holdway, D.A. The Effects of 17α-Ethinylestradiol on the Heart Rate of Embryonic Japanese Medaka (Oryzias latipes). Environ. Toxicol. Chem. 2020, 39, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Edwards, H.E.; Souder, J.P.; Ryan, K.J.; Cui, X.; Gorelick, D.A. G protein-coupled estrogen receptor regulates embryonic heart rate in zebrafish. PLoS Genet. 2017, 13, e1007069. [Google Scholar] [CrossRef] [PubMed]
- Palevsky, P.M.; Liu, K.; Brophy, P.D.; Chawla, L.; Parikh, C.R.; Thakar, C.V.; Tolwani, A.J.; Waikar, S.S.; Weisbord, S.D. KDOQI US Commentary on the 2012 KDIGO Clinical Practice Guideline for Acute Kidney Injury. Am. J. Kidney Dis. 2013, 61, 649–672. [Google Scholar] [CrossRef] [PubMed]
- International Society of Nephrology. Acute Kidney Injury. Available online: https://www.theisn.org/commitment-to-kidney-health/focus-areas/acute-kidney-injury/ (accessed on 10 January 2022).
- Neugarten, J.; Golestaneh, L.; Kolhe, N.V. Sex differences in acute kidney injury requiring dialysis. BMC Nephrol. 2018, 19, 131. [Google Scholar] [CrossRef] [PubMed]
- Hansrivijit, P.; Qian, C.; Boonpheng, B.; Thongprayoon, C.; Vallabhajosyula, S.; Cheungpasitporn, W.; Ghahramani, N. Incidence of acute kidney injury and its association with mortality in patients with COVID-19: A meta-analysis. J. Investig. Med. 2020, 68, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kader, K.; Palevsky, P.M. Acute Kidney Injury in the Elderly. Clin. Geriatr. Med. 2009, 25, 331–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coca, S.G. Acute Kidney Injury in Elderly Persons. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 2010, 56, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, E.; Alegre, L.; Blanco-Sanchez, I.; Sáenz-Morales, D.; Aguado-Fraile, E.; Ponte, B.; Ramos, E.; Saiz, A.; Jiménez, C.; Ordoñez, A.; et al. Hypoxia Inducible Factor 1-Alpha (HIF-1 Alpha) Is Induced during Reperfusion after Renal Ischemia and Is Critical for Proximal Tubule Cell Survival. PLoS ONE 2012, 7, e33258. [Google Scholar] [CrossRef] [PubMed]
- Neusser, M.A.; Lindenmeyer, M.T.; Moll, A.G.; Segerer, S.; Edenhofer, I.; Sen, K.; Stiehl, D.P.; Kretzler, M.; Gröne, H.-J.; Schlöndorff, D.; et al. Human Nephrosclerosis Triggers a Hypoxia-Related Glomerulopathy. Am. J. Pathol. 2010, 176, 594–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.K.; Anp, M.C.C. Acute Kidney Injury: Advances in Definition, Pathophysiology, and Diagnosis. AACN Adv. Crit. Care 2010, 21, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Singh, N.; Pathak, D.; Bedi, P.M.S. Estradiol attenuates ischemia reperfusion-induced acute kidney injury through PPAR-γ stimulated eNOS activation in rats. Mol. Cell. Biochem. 2018, 453, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Chang, C.-Y.; Chang, S.-T.; Chen, S.-H. 17β-Estradiol Accelerated Renal Tubule Regeneration in Male Rats After Ischemia/Reperfusion-Induced Acute Kidney Injury. Shock 2016, 46, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Randeva, H.S.; Patel, V.H.; Chen, J.; Ramanjaneya, M.; Karteris, E.; Zachariades, E.; Thomas, P.; Been, M. G-protein coupled estrogen receptor 1 expression in rat and human heart: Protective role during ischaemic stress. Int. J. Mol. Med. 2010, 26, 193–199. [Google Scholar] [CrossRef]
- Sartori-Valinotti, J.C.; Venegas-Pont, M.R.; LaMarca, B.B.; Romero, D.G.; Yanes, L.L.; Racusen, L.C.; Jones, A.V.; Ryan, M.J.; Reckelhoff, J.F. Rosiglitazone reduces blood pressure in female Dahl salt-sensitive rats. Steroids 2010, 75, 794–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banke, A.B.S.; Schou, M.; Videbaek, L.; Møller, J.E.; Torp-Pedersen, C.; Gustafsson, F.; Dahl, J.S.; Køber, L.; Hildebrandt, P.R.; Gislason, G.H. Incidence of cancer in patients with chronic heart failure: A long-term follow-up study. Eur. J. Heart Fail. 2016, 18, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Hasin, T.; Gerber, Y.; McNallan, S.M.; Weston, S.A.; Kushwaha, S.S.; Nelson, T.J.; Cerhan, J.R.; Roger, V.L. Patients with Heart Failure Have an Increased Risk of Incident Cancer. J. Am. Coll. Cardiol. 2013, 62, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Hasin, T.; Gerber, Y.; Weston, S.A.; Jiang, R.; Killian, J.M.; Manemann, S.M.; Cerhan, J.R.; Roger, V.L. Heart Failure after Myocardial Infarction Is Associated with Increased Risk of Cancer. J. Am. Coll. Cardiol. 2016, 68, 265–271. [Google Scholar] [CrossRef]
- Hasin, T.; Iakobishvili, Z.; Weisz, G. Associated Risk of Malignancy in Patients with Cardiovascular Disease: Evidence and Possible Mechanism. Am. J. Med. 2017, 130, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Adami, J.; Gabel, H.W.; Lindelof, B.; Ekstrom, K.; Rydh, B.; Glimelius, B.; Ekbom, A.; Adami, H.-O.; Granath, F. Cancer risk following organ transplantation: A nationwide cohort study in Sweden. Br. J. Cancer 2003, 89, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Buell, J.F.; Gross, T.G.; Woodle, E.S. Malignancy after Transplantation. Transplantation 2005, 80, S254–S264. [Google Scholar] [CrossRef] [PubMed]
- Vajdic, C.M.; McDonald, S.P.; McCredie, M.R.E.; van Leeuwen, M.; Stewart, J.H.; Law, M.; Chapman, J.R.; Webster, A.; Kaldor, J.M.; Grulich, A.E. Cancer Incidence Before and After Kidney Transplantation. JAMA 2006, 296, 2823–2831. [Google Scholar] [CrossRef] [Green Version]
- Maisonneuve, P.; Agodoa, L.; Gellert, R.; Stewart, J.H.; Buccianti, G.; Lowenfels, A.B.; Wolf, R.A.; Jones, E.; Dsiney, A.P.; Briggs, D.; et al. Cancer in patients on dialysis for end-stage renal disease: An international collaborative study. Lancet 1999, 354, 93–99. [Google Scholar] [CrossRef]
- Lowrance, W.T.; Ordoñez, J.; Udaltsova, N.; Russo, P.; Go, A.S. CKD and the Risk of Incident Cancer. J. Am. Soc. Nephrol. 2014, 25, 2327–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleman, B.M.; Moser, E.C.; Nuver, J.; Suter, T.M.; Maraldo, M.V.; Specht, L.; Vrieling, C.; Darby, S.C. Cardiovascular disease after cancer therapy. Eur. J. Cancer Suppl. 2014, 12, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, G.T.; Kawashima, T.; Leisenring, W.; Stratton, K.; Stovall, M.; Hudson, M.M.; Sklar, C.A.; Robison, L.L.; Oeffinger, K.C. Aging and Risk of Severe, Disabling, Life-Threatening, and Fatal Events in the Childhood Cancer Survivor Study. J. Clin. Oncol. 2014, 32, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Rosner, M.H.; Jhaveri, K.D.; McMahon, B.A.; Perazella, M.A. Onconephrology: The intersections between the kidney and cancer. CA Cancer J. Clin. 2021, 71, 47–77. [Google Scholar] [CrossRef] [PubMed]
- Malyszko, J.; Tesarova, P.; Capasso, G.; Capasso, A. The link between kidney disease and cancer: Complications and treatment. Lancet 2020, 396, 277–287. [Google Scholar] [CrossRef]
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of Chemotherapy plus a Monoclonal Antibody against HER2 for Metastatic Breast Cancer That Overexpresses HER2. N. Engl. J. Med. 2001, 344, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Hu, C.; Staquicini Daniela, I.; Brigman Jonathan, L.; Liu, M.; Mauvais-Jarvis, F.; Pasqualini, R.; Arap, W.; Arterburn Jeffrey, B.; Hathaway Helen, J.; et al. Preclinical efficacy of the GPER-selective agonist G-1 in mouse models of obesity and diabetes. Sci. Transl. Med. 2020, 12, eaau5956. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.; Nasci, V.L.; Guthrie, G.; Ertuglu, L.A.; Butt, M.K.; Kirabo, A.; Gohar, E.Y. Emerging Roles for G Protein-Coupled Estrogen Receptor 1 in Cardio-Renal Health: Implications for Aging. Biomolecules 2022, 12, 412. https://doi.org/10.3390/biom12030412
Singh R, Nasci VL, Guthrie G, Ertuglu LA, Butt MK, Kirabo A, Gohar EY. Emerging Roles for G Protein-Coupled Estrogen Receptor 1 in Cardio-Renal Health: Implications for Aging. Biomolecules. 2022; 12(3):412. https://doi.org/10.3390/biom12030412
Chicago/Turabian StyleSingh, Ravneet, Victoria L. Nasci, Ginger Guthrie, Lale A. Ertuglu, Maryam K. Butt, Annet Kirabo, and Eman Y. Gohar. 2022. "Emerging Roles for G Protein-Coupled Estrogen Receptor 1 in Cardio-Renal Health: Implications for Aging" Biomolecules 12, no. 3: 412. https://doi.org/10.3390/biom12030412