
Abnormal RasGRP1 Expression in the Post-Mortem Brain and Blood Serum of Schizophrenia Patients

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Post-Mortem Tissue Collection
2.2. RNA Extraction and Quantitative RT-PCR Analysis
2.3. Western Blotting
2.4. Human Patient and Control Serum Collection
2.5. Enzyme Linked Immunosorbent Assay (ELISA)
2.6. Statistical Analysis
3. Results
3.1. RasGRP1 Protein Levels Negatively Correlate with Age in the DLPFC of Schizophrenia Patients
3.2. Increased RasGRP1 mRNA Levels in the DLPFC of Patients with SCZ
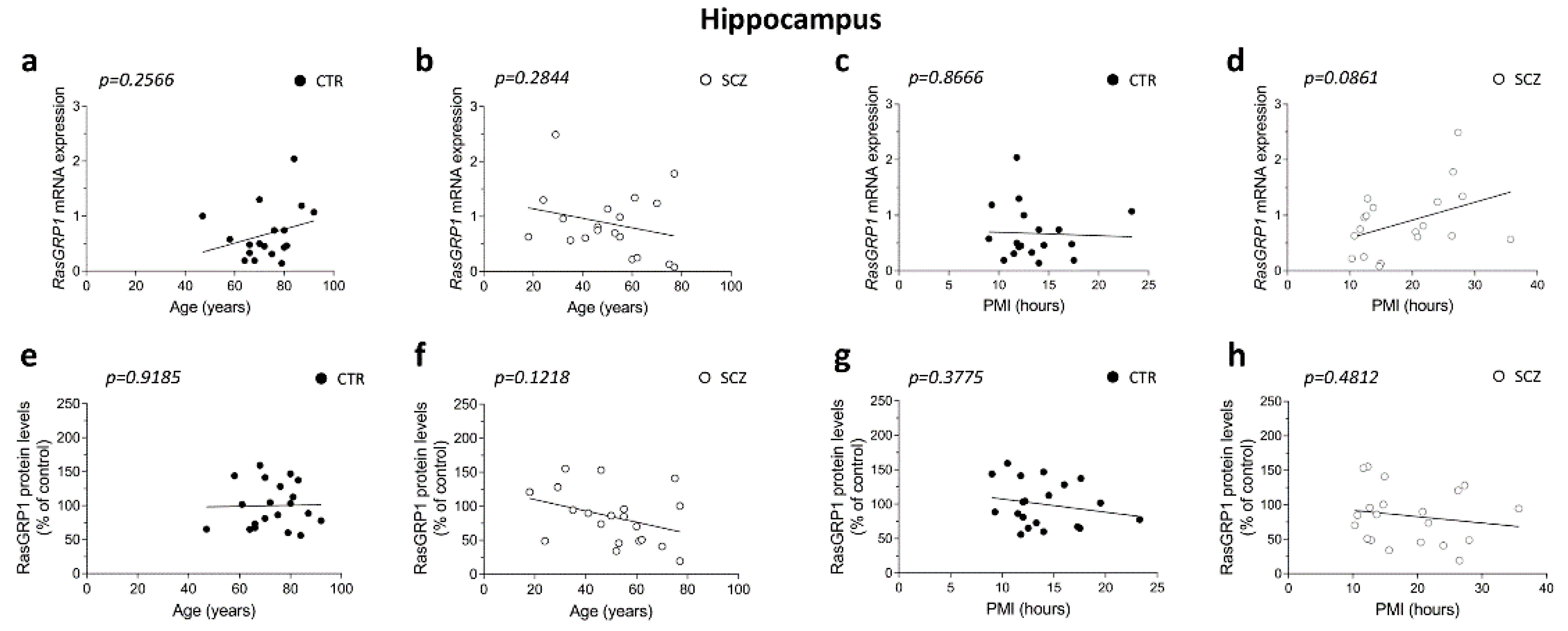
3.3. Lack of Correlation between RasGRP1 mRNA and Protein Levels with Age and PMI in the Hippocampus of Schizophrenia Patients
3.4. Unaltered RasGRP1 mRNA and Protein Levels in the Hippocampus of Schizophrenia Patients
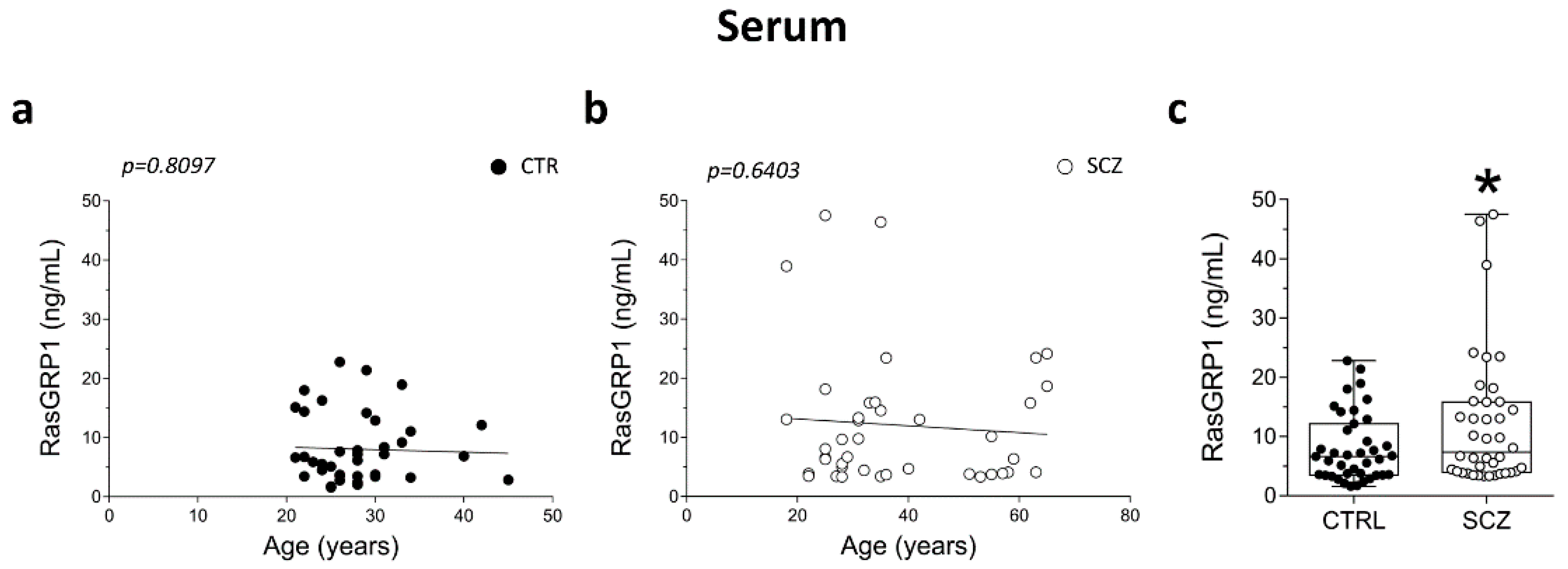
3.5. Detection of RasGRP1 Protein Concentration in the Serum of Healthy Controls and Schizophrenia Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marder, S.R.; Galderisi, S. The current conceptualization of negative symptoms in schizophrenia. World Psychiatry 2017, 16, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Howes, O.D.; McCutcheon, R.; Owen, M.J.; Murray, R. The Role of Genes, Stress, and Dopamine in the Development of Schizophrenia. Biol. Psychiatry 2017, 81, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, K.-A.; Burne, T.; Kesby, J.P. Subcortical Dopamine and Cognition in Schizophrenia: Looking Beyond Psychosis in Preclinical Models. Front. Neurosci. 2020, 14, 542. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, W.T.; O’Shea, S.D. Clozapine and GABA transmission in schizophrenia disease models: Establishing principles to guide treatments. Pharmacol. Ther. 2015, 150, 47–80. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Coyle, J.T. Glutamate hypothesis in schizophrenia. Psychiatry Clin. Neurosci. 2019, 73, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, N.; van Praag, H.M. The role of serotonin in schizophrenia. Eur. Neuropsychopharmacol. 1995, 5, 11–23. [Google Scholar] [CrossRef]
- Di Maio, A.; De Rosa, A.; Pelucchi, S.; Garofalo, M.; Marciano, B.; Nuzzo, T.; Gardoni, F.; Isidori, A.M.; Di Luca, M.; Errico, F.; et al. Analysis of mRNA and Protein Levels of CAP2, DLG1 and ADAM10 Genes in Post-Mortem Brain of Schizophrenia, Parkinson’s and Alzheimer’s Disease Patients. Int. J. Mol. Sci. 2022, 23, 1539. [Google Scholar] [CrossRef]
- De Rosa, A.; Fontana, A.; Nuzzo, T.; Garofalo, M.; Di Maio, A.; Punzo, D.; Copetti, M.; Bertolino, A.; Errico, F.; Rampino, A.; et al. Machine Learning algorithm unveils glutamatergic alterations in the post-mortem schizophrenia brain. NPJ Schizophrenia. in press.
- Irish Schizophrenia Genomics Consortium; Wellcome Trust Case Control Consortium. Genome-wide association study implicates HLA-C*01:02 as a risk factor at the major histocompatibility complex locus in schizophrenia. Biol. Psychiatry 2012, 72, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Müller, N.; Schwarz, M.J. Immune System and Schizophrenia. Curr. Immunol. Rev. 2010, 6, 213–220. [Google Scholar] [CrossRef]
- Davies, C.; Segre, G.; Estradé, A.; Radua, J.; De Micheli, A.; Provenzani, U.; Oliver, D.; de Pablo, G.S.; Ramella-Cravaro, V.; Besozzi, M.; et al. Prenatal and perinatal risk and protective factors for psychosis: A systematic review and meta-analysis. Lancet Psychiatry 2020, 7, 399–410. [Google Scholar] [CrossRef]
- Kinney, D.K.; Hintz, K.; Shearer, E.M.; Barch, D.H.; Riffin, C.; Whitley, K.; Butler, R. A unifying hypothesis of schizophrenia: Abnormal immune system development may help explain roles of prenatal hazards, post-pubertal onset, stress, genes, climate, infections, and brain dysfunction. Med. Hypotheses 2010, 74, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Pouget, J.G.; Zai, C.C.; Kennedy, J.L. The complement system in schizophrenia: Where are we now and what’s next? Mol. Psychiatry 2020, 25, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, C.; Gelernter, J.; Zhao, H. Pervasive pleiotropy between psychiatric disorders and immune disorders revealed by integrative analysis of multiple GWAS. Qual. Life Res. 2015, 134, 1195–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, J.L.; Rehmann, H.; Wittinghofer, A. GEFs and GAPs: Critical Elements in the Control of Small G Proteins. Cell 2007, 129, 865–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanpoor, M.; Ghaedi, H.; Omrani, M.D. The genetic basis for the inverse relationship between rheumatoid arthritis and schizophrenia. Mol. Genet. Genomic Med. 2020, 8, 1483. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Yun, G.; Hebert, A.; Kong, G.; Ranheim, E.A.; Finn, R.; Rajagoplan, A.; Li, S.; Zhou, Y.; Yu, M.; et al. NrasQ61R/+ and Kras−/− cooperate to downregulate Rasgrp1 and promote lympho-myeloid leukemia in early T-cell precursors. Blood 2021, 137, 3259–3271. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Kurita, T.; Yasuda, S.; Fujieda, Y.; Bohgaki, T.; Katsuyama, T.; Tsokos, G.C.; Moulton, V.R.; Atsumi, T. Decreased Expression of Serine/Arginine-Rich Splicing Factor 1 in T Cells From Patients With Active Systemic Lupus Erythematosus Accounts for Reduced Expression of RasGRP1 and DNA Methyltransferase 1. Arthritis Rheumatol. 2018, 70, 2046–2056. [Google Scholar] [CrossRef] [Green Version]
- Hartzell, C.; Ksionda, O.; Lemmens, E.; Coakley, K.; Yang, M.; Dail, M.; Harvey, R.C.; Govern, C.; Bakker, J.; Lenstra, T.L.; et al. Dysregulated RasGRP1 Responds to Cytokine Receptor Input in T Cell Leukemogenesis. Sci. Signal. 2013, 6, ra21. [Google Scholar] [CrossRef] [Green Version]
- Somekh, I.; Marquardt, B.; Liu, Y.; Rohlfs, M.; Hollizeck, S.; Karakukcu, M.; Unal, E.; Yilmaz, E.; Patiroglu, T.; Cansever, M.; et al. Correction to: Novel Mutations in RASGRP1 Are Associated with Immunodeficiency, Immune Dysregulation, and EBV-Induced Lymphoma. J. Clin. Immunol. 2018, 38, 711. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Takuma, K.; Kamei, H.; Ito, Y.; Nakamichi, N.; Ibi, D.; Nakanishi, Y.; Murai, M.; Mizoguchi, H.; Nabeshima, T.; et al. Dopamine D1 receptors regulate protein synthesis-dependent long-term recognition memory via extracellular signal-regulated kinase 1/2 in the prefrontal cortex. Learn. Mem. 2007, 14, 117–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshraghi, M.; Ramírez-Jarquín, U.N.; Shahani, N.; Nuzzo, T.; De Rosa, A.; Swarnkar, S.; Galli, N.; Rivera, O.; Tsaprailis, G.; Scharager-Tapia, C.; et al. RasGRP1 is a causal factor in the development of l-DOPA–induced dyskinesia in Parkinson’s disease. Sci. Adv. 2020, 6, eaaz7001. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, E.; Jaffe, A.E.; Straub, R.E.; Chen, Q.; Shin, J.H.; Hyde, T.M.; Kleinman, J.E.; Weinberger, D.R. Identification and prioritization of gene sets associated with schizophrenia risk by co-expression network analysis in human brain. Mol. Psychiatry 2018, 25, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, J.R.; Zhai, S.; Sauvage, M.; Kitsukawa, T.; Burguiere, E.; Thomsen, M.; Zhang, H.; Costa, C.; Martella, G.; Ghiglieri, V.; et al. CalDAG-GEFI mediates striatal cholinergic modulation of dendritic excitability, synaptic plasticity and psychomotor behaviors. Neurobiol. Dis. 2021, 158, 105473. [Google Scholar] [CrossRef]
- Querques, G.; Souied, E.H. Vascularized Drusen: Slowly Progressive Type 1 Neovascularization Mimicking Drusenoid Retinal Pigment Epithelium Elevation. Retina 2015, 35, 2433–2439. [Google Scholar] [CrossRef]
- Agilent, version 2100; Agilent Expert Bioanalyzer Software: Santa Clara, CA, USA, 2019.
- Bio-Rad, version 3.20; Experion Software: Hercules, CA, USA, 2017.
- Pierret, P.; Vallée, A.; Mechawar, N.; Dower, N.; Stone, J.; Richardson, P.; Dunn, R. Cellular and subcellular localization of Ras guanyl nucleotide-releasing protein in the rat hippocampus. Neuroscience 2001, 108, 381–390. [Google Scholar] [CrossRef]
- Daley, S.R.; Coakley, K.M.; Hu, D.Y.; Randall, K.L.; Jenne, C.N.; Limnander, A.; Myers, D.R.; Polakos, N.K.; Enders, A.; Roots, C.; et al. Rasgrp1 mutation increases naive T-cell CD44 expression and drives mTOR-dependent accumulation of Helios+ T cells and autoantibodies. Elife 2013, 2, e01020. [Google Scholar] [CrossRef]
- Khandaker, G.M.; Cousins, L.; Deakin, J.; Lennox, B.; Yolken, R.; Jones, P. Inflammation and immunity in schizophrenia: Implications for pathophysiology and treatment. Lancet Psychiatry 2015, 2, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Dower, N.A.; Stang, S.L.; Bottorff, D.A.; Ebinu, J.O.; Dickie, P.; Ostergaard, H.L.; Stone, J.C. RasGRP is essential for mouse thymocyte differentiation and TCR signaling. Nat. Immunol. 2000, 1, 317–321. [Google Scholar] [CrossRef]
- Shahani, N.; Swarnkar, S.; Giovinazzo, V.; Morgenweck, J.; Bohn, L.M.; Scharager-Tapia, C.; Pascal, B.; Martinez-Acedo, P.; Khare, K.; Subramaniam, S. RasGRP1 promotes amphetamine-induced motor behavior through a Rhes interaction network (“Rhesactome”) in the striatum. Sci. Signal. 2016, 9, ra111. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-L.; Fann, C.S.-J.; Liu, C.-M.; Chen, W.J.; Wu, J.-Y.; Hung, S.-I.; Chen, C.-H.; Jou, Y.-S.; Liu, S.-K.; Hwang, T.-J.; et al. RASD2, MYH9, and CACNG2 Genes at Chromosome 22q12 Associated with the Subgroup of Schizophrenia with Non-Deficit in Sustained Attention and Executive Function. Biol. Psychiatry 2008, 64, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, F.; D’Angelo, L.; de Girolamo, P.; Avallone, L.; de Lange, P.; Usiello, A. The Thyroid Hormone-target Gene Rhes a Novel Crossroad for Neurological and Psychiatric Disorders: New Insights from Animal Models. Neurosci. 2018, 384, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Vitucci, D.; Di Giorgio, A.; Napolitano, F.; Pelosi, B.; Blasi, G.; Errico, F.; Attrotto, M.T.; Gelao, B.; Fazio, L.; Taurisano, P.; et al. Rasd2 Modulates Prefronto-Striatal Phenotypes in Humans and ‘Schizophrenia-Like Behaviors’ in Mice. Neuropsychopharmacology 2015, 41, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Hayashi-Takagi, A.; Toyota, T.; Yoshikawa, T.; Iwamoto, K. Gene expression analysis in lymphoblastoid cells as a potential biomarker of bipolar disorder. J. Hum. Genet. 2011, 56, 779–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Identification of risk loci with shared effects on five major psychiatric disorders: A genome-wide analysis. Lancet 2013, 381, 1371–1379. [Google Scholar] [CrossRef]
- Golinski, M.L.; Vandhuick, T.; Derambure, C.; Freret, M.; Lecuyer, M.; Guillou, C.; Hiron, M.; Boyer, O.; le Loet, X.; Vittecoq, O.; et al. Dysregulation of RasGRP1 in rheumatoid arthritis and modulation of RasGRP3 as a biomarker of TNFalpha inhibitors. Arthritis Res. Ther. 2015, 17, 382. [Google Scholar] [CrossRef] [Green Version]
- Torretta, S.; Rampino, A.; Basso, M.; Pergola, G.; Di Carlo, P.; Shin, J.H.; Kleinman, J.E.; Hyde, T.M.; Weinberger, D.R.; Masellis, R.; et al. NURR1 and ERR1 Modulate the Expression of Genes of a DRD2 Coexpression Network Enriched for Schizophrenia Risk. J. Neurosci. 2019, 40, 932–941. [Google Scholar] [CrossRef]
- Dadalko, O.I.; Siuta, M.; Poe, A.; Erreger, K.; Matthies, H.J.; Niswender, K.; Galli, A. mTORC2/rictor signaling disrupts dopamine-dependent behaviors via defects in striatal dopamine neurotransmission. J. Neurosci. 2015, 35, 8843–8854. [Google Scholar] [CrossRef]
- Kawasaki, H.; Springett, G.M.; Toki, S.; Canales, J.J.; Harlan, P.; Blumenstiel, J.; Chen, E.J.; Bany, I.A.; Mochizuki, N.; Ashbacher, A.; et al. A Rap guanine nucleotide exchange factor enriched highly in the basal ganglia. Proc. Natl. Acad. Sci. USA 1998, 95, 13278–13283. [Google Scholar] [CrossRef] [Green Version]
- Ebinu, J.O.; Bottorff, D.A.; Chan, E.Y.W.; Stang, S.L.; Dunn, R.J.; Stone, J.C. RasGRP, a Ras Guanyl Nucleotide- Releasing Protein with Calcium- and Diacylglycerol-Binding Motifs. Science 1998, 280, 1082–1086. [Google Scholar] [CrossRef]
- Haberman, R.P.; Lee, H.J.; Colantuoni, C.; Koh, M.T.; Gallagher, M. Rapid encoding of new information alters the profile of plasticity-related mRNA transcripts in the hippocampal CA3 region. Proc. Natl. Acad. Sci. USA 2008, 105, 10601–10606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierret, P.; Dunn, R.; Djordjevic, B.; Stone, J.; Richardson, P. Distribution of ras guanyl releasing protein (RasGRP) mRNA in the adult rat central nervous system. J. Neurocytol. 2000, 29, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Lisowski, P.; Wieczorek, M.; Goscik, J.; Juszczak, G.; Stankiewicz, A.; Zwierzchowski, L.; Swiergiel, A.H. Effects of Chronic Stress on Prefrontal Cortex Transcriptome in Mice Displaying Different Genetic Backgrounds. J. Mol. Neurosci. 2013, 50, 33–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pries, L.-K.; Van Os, J.; Have, M.T.; De Graaf, R.; Van Dorsselaer, S.; Bak, M.; Lin, B.D.; Van Eijk, K.R.; Kenis, G.; Richards, A.; et al. Association of Recent Stressful Life Events with Mental and Physical Health in the Context of Genomic and Exposomic Liability for Schizophrenia. JAMA Psychiatry 2020, 77, 1296. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control | Schizophrenia | Statistic | p-Value |
|---|---|---|---|---|
| Subjects (total number) | 20 | 20 | − | − |
| Gender (M/F) | 16/4 | 12/8 | χ2 = 1.071 (df = 1) | 0.301 a |
| Age (years, median [IQR]) | 73.50 [66.00–80.25] | 52.50 [39.50–61.25] | t = 4.819 (df = 38) | <0.001 b |
| PMI (hours, median [IQR]) | 12.90 [11.80–16.32] | 15.25 [12.52–24.58] | t = −2.426 (df = 38) | 0.020 c |
| pH (median, [IQR]) | 6.54 [6.49–6.63] | 6.50 [6.42–6.56] | t = 0.708 (df = 26) | 0.485 c |
| RIN (median, [IQR]) | 6.05 [5.50–7.12] | 6.75 [6.05–7.08] | t = −0.036 (df = 38) | 0.971 b |
| Characteristics | Control | Schizophrenia | Statistic | p-Value |
|---|---|---|---|---|
| Subjects (total number) | 39 | 40 | − | − |
| Gender (M/F) | 15/24 | 22/18 | χ2 = 2.17 (df = 1) | 0.141 |
| Age (years, median [IQR]) | 28 [24,25,28–32] | 33.50 [27.25–54.5] | t = 4.021 (df = 78) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Rosa, A.; Di Maio, A.; Torretta, S.; Garofalo, M.; Giorgelli, V.; Masellis, R.; Nuzzo, T.; Errico, F.; Bertolino, A.; Subramaniam, S.; et al. Abnormal RasGRP1 Expression in the Post-Mortem Brain and Blood Serum of Schizophrenia Patients. Biomolecules 2022, 12, 328. https://doi.org/10.3390/biom12020328
De Rosa A, Di Maio A, Torretta S, Garofalo M, Giorgelli V, Masellis R, Nuzzo T, Errico F, Bertolino A, Subramaniam S, et al. Abnormal RasGRP1 Expression in the Post-Mortem Brain and Blood Serum of Schizophrenia Patients. Biomolecules. 2022; 12(2):328. https://doi.org/10.3390/biom12020328
Chicago/Turabian StyleDe Rosa, Arianna, Anna Di Maio, Silvia Torretta, Martina Garofalo, Valentina Giorgelli, Rita Masellis, Tommaso Nuzzo, Francesco Errico, Alessandro Bertolino, Srinivasa Subramaniam, and et al. 2022. "Abnormal RasGRP1 Expression in the Post-Mortem Brain and Blood Serum of Schizophrenia Patients" Biomolecules 12, no. 2: 328. https://doi.org/10.3390/biom12020328