
The Disordered Dehydrin and Its Role in Plant Protection: A Biochemical Perspective

Abstract
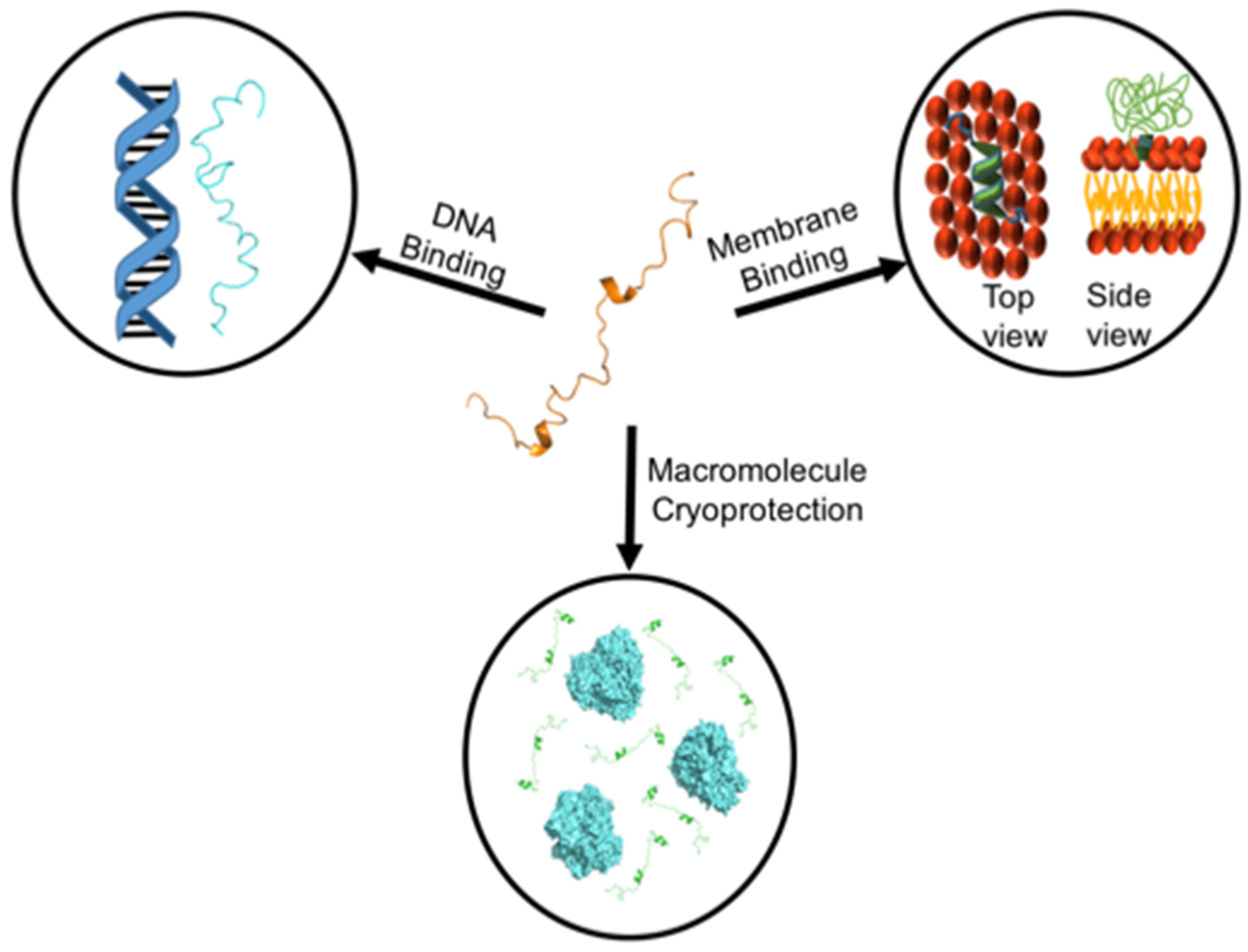
:1. Introduction
2. Dehydrin Sequence and Architecture
3. Evolution of Dehydrins
4. Intrinsic Disorder and Structure of Dehydrins
5. Membrane Binding
6. Protein Cryoprotection
7. DNA Binding and Protection
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Steponkus, P.L.; Uemura, M.; Joseph, R.A.; Gilmour, S.J.; Thomashow, M.F. Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1998, 95, 14570–14575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galau, G.A.; Hughes, D.W.; Dure, L. Abscisic acid induction of cloned cotton late embryogenesis-abundant (Lea) mRNAs. Plant Mol. Biol. 1986, 7, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Tunnacliffe, A.; Wise, M.J. The continuing conundrum of the LEA proteins. Naturwissenschaften 2007, 94, 791–812. [Google Scholar] [CrossRef] [PubMed]
- Hellwege, E.M.; Dietz, K.J.; Volk, O.H.; Hartung, W. Abscisic acid and the induction of desiccation tolerance in the extremely xerophilic liverwort Exormotheca holstii. Planta 1994, 194, 525–531. [Google Scholar] [CrossRef]
- Li, R.; Brawley, S.H.; Close, T.J. Proteins immunologically related to dehydrins in fucoid algae. J. Phycol. 1998, 34, 642–650. [Google Scholar] [CrossRef]
- Lan, T.; Gao, J.; Zeng, Q.Y. Genome-wide analysis of the LEA (late embryogenesis abundant) protein gene family in Populus trichocarpa. Tree Genet. Genomes 2013, 9, 253–264. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plantarum 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef] [Green Version]
- Nylander, M.; Svensson, J.; Palva, E.T.; Welin, B.V. Stress-induced accumulation and tissue-specific localization of dehydrins in Arabidopsis thaliana. Plant Mol. Biol. 2001, 45, 263–279. [Google Scholar] [CrossRef]
- Yang, Y.; He, M.; Zhu, Z.; Li, S.; Xu, Y.; Zhang, C.; Singer, S.D.; Wang, Y. Identification of the dehydrin gene family from grapevine species and analysis of their responsiveness to various forms of abiotic and biotic stress. BMC Plant Biol. 2012, 12, 140. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Hall, A.E.; Close, T.J. Chilling tolerance during emergence of cowpea associated with a dehydrin and slow electrolyte leakage. Crop Sci. 1997, 37, 1270–1277. [Google Scholar] [CrossRef]
- Cheng, Z.; Targolli, J.; Huang, X.; Wu, R. Wheat LEA genes, PMA80 and PMA1959, enhance dehydration tolerance of transgenic rice (Oryza sativa L.). Mol. Breed. 2002, 10, 71–82. [Google Scholar] [CrossRef]
- Hara, M.; Terashima, S.; Fukaya, T.; Kuboi, T. Enhancement of cold tolerance and inhibition of lipid peroxidation by citrus dehydrin in transgenic tobacco. Planta 2003, 217, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, Y.; Guan, Z.; Wei, W.; Han, L.; Chai, T. Expression and function of two dehydrins under environmental stresses in Brassica juncea L. Mol. Breed. 2008, 21, 431–438. [Google Scholar] [CrossRef]
- Xing, X.; Liu, Y.; Kong, X.; Liu, Y.; Li, D. Overexpression of a maize dehydrin gene, ZmDHN2b, in tobacco enhances tolerance to low temperature. Plant Growth Regul. 2011, 65, 109–118. [Google Scholar] [CrossRef]
- Bao, F.; Du, D.; An, Y.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Overexpression of Prunus mume dehydrin genes in tobacco enhances tolerance to cold and drought. Front. Plant Sci. 2017, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Puhakainen, T.; Hess, M.W.; Mäkelä, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol. 2004, 54, 743–753. [Google Scholar] [CrossRef]
- Dure, L.; Crouch, M.; Harada, J.; Ho, T.H.D.; Mundy, J.; Quatrano, R.; Thomas, T.; Sung, Z.R. Common amino acid sequence domains among the LEA proteins of higher plants. Plant Mol. Biol. 1989, 12, 475–486. [Google Scholar] [CrossRef]
- Strimbeck, G.R. Hiding in plain sight: The F-segment and other conserved features of seed plant SKn dehydrins. Planta 2017, 245, 1061–1066. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.A.; Veltri, M.; Boddington, K.F.; Singh, K.K.; Graether, S.P. Genome analysis of conserved dehydrin motifs in vascular plants. Front. Plant Sci. 2017, 8, 709. [Google Scholar] [CrossRef] [Green Version]
- Riley, A.C.; Ashlock, D.A.; Graether, S.P. Evolution of the modular, disordered stress proteins known as dehydrins. PLoS ONE 2019, 14, e0211813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boddington, K.F.; Graether, S.P. Binding of a Vitis riparia Dehydrin to DNA. Plant Sci. 2019, 287, 110172. [Google Scholar] [CrossRef] [PubMed]
- Maszkowska, J.; Dębski, J.; Kulik, A.; Kistowski, M.; Bucholc, M.; Lichocka, M.; Klimecka, M.; Sztatelman, O.; Szymańska, K.P.; Dadlez, M.; et al. Phosphoproteomic analysis reveals that dehydrins ERD10 and ERD14 are phosphorylated by SNF1-related protein kinase 2.10 in response to osmotic stress. Plant Cell Environ. 2019, 42, 931–946. [Google Scholar] [CrossRef]
- Alsheikh, M.K.; Heyen, B.J.; Randall, S.K. Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. J. Biol. Chem. 2003, 278, 40882–40889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goday, A.; Jensen, A.B.; Culiáñez-Macià, F.A.; Mar Albà, M.; Figueras, M.; Serratosa, J.; Torrent, M.; Pagès, M. The maize abscisic acid-responsive protein Rab17 is located in the nucleus and interacts with nuclear localization signals. Plant Cell 1994, 6, 351–360. [Google Scholar] [PubMed]
- Saavedra, L.; Svensson, J.; Carballo, V.; Izmendi, D.; Welin, B.; Vidal, S. A dehydrin gene in Physcomitrella patens is required for salt and osmotic stress tolerance. Plant J. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Wei, H.; Yang, Y.; Himmel, M.E.; Tucker, M.P.; Ding, S.Y.; Yang, S.; Arora, R. Identification and characterization of five cold stress-related rhododendron dehydrin genes: Spotlight on a FSK-type dehydrin with multiple F-segments. Front. Bioeng. Biotech. 2019, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Li, C.M.; Liu, B.G.; Ge, S.J.; Dong, X.M.; Li, W.; Zhu, H.Y.; Wang, B.C.; Yang, C.P. Genome-wide identification and characterization of a dehydrin gene family in poplar (Populus trichocarpa). Plant Mol. Biol. Rep. 2012, 30, 848–859. [Google Scholar] [CrossRef]
- Zhang, X.H.; Moloney, M.M.; Chinnappa, C.C. Nucleotide sequence of a cDNA clone encoding a dehydrin-like protein from Stellaria longipes. Plant Physiol. 1993, 103, 1029. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.S.; Kim, H.Y.; Kim, Y.S.; Choi, H.G.; Kang, S.H.; Yoon, H.S. Expression of dehydrin gene from Arctic Cerastium arcticum increases abiotic stress tolerance and enhances the fermentation capacity of a genetically engineered Saccharomyces cerevisiae laboratory strain. Appl. Microbiol. Biotechnol. 2013, 97, 8997–9009. [Google Scholar] [CrossRef]
- Verma, G.; Dhar, Y.V.; Srivastava, D.; Kidwai, M.; Chauhan, P.S.; Bag, S.K.; Asif, M.H.; Chakrabarty, D. Genome-wide analysis of rice dehydrin gene family: Its evolutionary conservedness and expression pattern in response to PEG induced dehydration stress. PLoS ONE 2017, 12, e0176399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zan, T.; Li, L.; Li, J.; Zhang, L.; Li, X. Genome-wide identification and characterization of late embryogenesis abundant protein-encoding gene family in wheat: Evolution and expression profiles during development and stress. Gene 2020, 736, 144422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, H.; Liang, D.; Lin, L.; Deng, H.; Lv, X.; Wang, Z.; Zhang, X.; Wang, J.; Xiong, B. Genome-wide identification and expression profiling of the dehydrin gene family in Actinidia chinensis. Sci. Hortic. 2021, 280, 109930. [Google Scholar] [CrossRef]
- Archambault, A.; Strömvik, M.V. The Y-segment of novel cold dehydrin genes is conserved and codons in the PR-10 genes are under positive selection in Oxytropis (Fabaceae) from contrasting climates. Mol. Genet. Genom. 2012, 287, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Charfeddine, S.; Saïdi, M.N.; Charfeddine, M.; Gargouri-Bouzid, R. Genome-wide identification and expression profiling of the late embryogenesis abundant genes in potato with emphasis on dehydrins. Mol. Biol. Rep. 2015, 42, 1163–1174. [Google Scholar] [CrossRef]
- Stival Sena, J.; Giguère, I.; Rigault, P.; Bousquet, J.; Mackay, J. Expansion of the dehydrin gene family in the Pinaceae is associated with considerable structural diversity and drought-responsive expression. Tree Physiol. 2018, 38, 442–456. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Zhao, T.; Ligterink, W.; Schranz, E.; Hilhorst, H.W. Dissecting the genomic diversification of late embryogenesis abundant (LEA) protein gene families in plants. Genome Biol. Evol. 2019, 11, 459–471. [Google Scholar] [CrossRef]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Coupling of folding and binding for unstructured proteins. Curr. Opin. Struct. Biol. 2002, 12, 54–60. [Google Scholar] [CrossRef]
- Lisse, T.; Bartels, D.; Kalbitzer, H.R.; Jaenicke, R. The recombinant dehydrin-like desiccation stress protein from the resurrection plant Craterostigma plantagineum displays no defined three-dimensional structure in its native state. Biol. Chem. 1996, 377, 555–562. [Google Scholar]
- Hughes, S.; Graether, S.P. Cryoprotective mechanism of a small intrinsically disordered dehydrin protein. Protein Sci. 2011, 20, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, J.A.; Singh, V.K.; Jia, Z.; Forman-Kay, J.D. Sensitivity of secondary structure propensities to sequence differences between α-and γ-synuclein: Implications for fibrillation. Protein Sci. 2006, 15, 2795–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouillon, J.M.; Eriksson, S.K.; Harryson, P. Mimicking the plant cell interior under water stress by macromolecular crowding: Disordered dehydrin proteins are highly resistant to structural collapse. Plant Physiol. 2008, 148, 1925–1937. [Google Scholar] [CrossRef] [Green Version]
- Receveur-Bréchot, V.; Bourhis, J.M.; Uversky, V.N.; Canard, B.; Longhi, S. Assessing protein disorder and induced folding. Proteins 2006, 62, 24–45. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Sánchez, I.E.; Martynowicz, D.M.; Rodríguez-Hernández, A.A.; Pérez-Morales, M.B.; Graether, S.P.; Jiménez-Bremont, J.F. A dehydrin-dehydrin interaction: The case of SK3 from Opuntia streptacantha. Front. Plant Sci. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Sánchez, I.E.; Maruri, I.; Ferrando, A.; Carbonell, J.; Graether, S.P.; Jimenez Bremont, J.F. Nuclear localization of the dehydrin OpsDHN1 is determined by histidine-rich motif. Front. Plant Sci. 2015, 6, 702. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Graether, S.P.; Jiménez-Bremont, J.F. In vivo evidence for homo-and heterodimeric interactions of Arabidopsis thaliana dehydrins AtCOR47, AtERD10, and AtRAB18. Sci. Rep. 2017, 7, 17036. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Kamm, W.J.; Steponkus, P.L. Lamellar-to-hexagonalII phase transitions in the plasma membrane of isolated protoplasts after freeze-induced dehydration. Proc. Natl. Acad. Sci. USA 1984, 81, 6373–6377. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Hall, A.E.; Close, T.J. Purification and partial characterization of a dehydrin involved in chilling tolerance during seedling emergence of cowpea. Plant Physiol. 1999, 120, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Terashima, S.; Kuboi, T. Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu. J. Plant Physiol. 2001, 158, 1333–1339. [Google Scholar] [CrossRef]
- Soulages, J.L.; Kim, K.; Arrese, E.L.; Walters, C.; Cushman, J.C. Conformation of a group 2 late embryogenesis abundant protein from soybean. Evidence of poly (L-proline)-type II structure. Plant Physiol. 2003, 131, 963–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koag, M.C.; Fenton, R.D.; Wilkens, S.; Close, T.J. The binding of maize DHN1 to lipid vesicles. Gain of structure and lipid specificity. Plant Physiol. 2003, 131, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koag, M.-C.; Wilkens, S.; Fenton, R.D.; Resnik, J.; Vo, E.; Close, T.J. The K-Segment of Maize DHN1 Mediates Binding to Anionic Phospholipid Vesicles and Concomitant Structural Changes. Plant Physiol. 2009, 150, 1503–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Nassuth, A. Stress-and development-induced expression of spliced and unspliced transcripts from two highly similar dehydrin 1 genes in V. riparia and V. vinifera. Plant Cell Rep. 2006, 25, 968–977. [Google Scholar] [CrossRef]
- Clarke, M.W.; Boddington, K.F.; Warnica, J.M.; Atkinson, J.; McKenna, S.; Madge, J.; Barker, C.H.; Graether, S.P. Structural and functional insights into the cryoprotection of membranes by the intrinsically disordered dehydrins. J. Biol. Chem. 2015, 290, 26900–26913. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.; Eremina, N.; Barth, A.; Danielsson, J.; Harryson, P. Membrane-induced folding of the plant stress dehydrin Lti30. Plant Physiol. 2016, 171, 932–943. [Google Scholar]
- Strandberg, E.; Killian, J.A. Snorkeling of lysine side chains in transmembrane helices: How easy can it get? FEBS Lett. 2003, 544, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Marzinek, J.K.; Jefferies, D.; Bond, P.J.; Harryson, P.; Wohland, T. The disordered plant dehydrin Lti30 protects the membrane during water-related stress by cross-linking lipids. J. Biol. Chem. 2019, 294, 6468–6482. [Google Scholar] [CrossRef] [Green Version]
- Andersson, J.M.; Pham, Q.D.; Mateos, H.; Eriksson, S.; Harryson, P.; Sparr, E. The plant dehydrin Lti30 stabilizes lipid lamellar structures in varying hydration conditions. J. Lipid Res. 2020, 61, 1014–1024. [Google Scholar] [CrossRef]
- Kovacs, D.; Kalmar, E.; Torok, Z.; Tompa, P. Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol. 2008, 147, 381–390. [Google Scholar] [CrossRef]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, S.K.; Kutzer, M.; Procek, J.; Gröbner, G.; Harryson, P. Tunable membrane binding of the intrinsically disordered dehydrin Lti30, a cold-induced plant stress protein. Plant Cell 2011, 23, 2391–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibowski, E.; Szcześ, A. Zeta potential and surface charge of DPPC and DOPC liposomes in the presence of PLC enzyme. Adsorption 2016, 22, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Chakrabortee, S.; Tripathi, R.; Watson, M.; Schierle, G.S.K.; Kurniawan, D.P.; Kaminski, C.F.; Wise, M.J.; Tunnacliffe, A. Intrinsically disordered proteins as molecular shields. Mol. BioSyst. 2012, 8, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Tompa, P.; Bánki, P.; Bokor, M.; Kamasa, P.; Kovács, D.; Lasanda, G.; Tompa, K. Protein-water and protein-buffer interactions in the aqueous solution of an intrinsically unstructured plant dehydrin: NMR intensity and DSC aspects. Biophys. J. 2006, 91, 2243–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Thomashow, M.F. A cold-regulated Arabidopsis gene encodes a polypeptide having potent cryoprotective activity. Biochem. Biophys. Res. Commun. 1992, 183, 1103–1108. [Google Scholar] [CrossRef]
- Kazuoka, T.; Oeda, K. Purification and characterization of COR85-oligomeric complex from cold-acclimated spinach. Plant Cell Physiol. 1994, 35, 601–611. [Google Scholar] [CrossRef]
- Momma, M.; Kaneko, S.; Haraguchi, K.; Matsukura, U. Peptide mapping and assessment of cryoprotective activity of 26/27-kDa dehydrin from soybean seeds. Biosci. Biotechnol. Biochem. 2003, 67, 1832–1835. [Google Scholar] [CrossRef] [Green Version]
- Goyal, K.; Walton, L.J.; Tunnacliffe, A. LEA proteins prevent protein aggregation due to water stress. Biochem. J. 2005, 388, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Drira, M.; Saibi, W.; Brini, F.; Gargouri, A.; Masmoudi, K.; Hanin, M. The K-segments of the wheat dehydrin DHN-5 are essential for the protection of lactate dehydrogenase and β-glucosidase activities in vitro. Mol. Biotechnol. 2013, 54, 643–650. [Google Scholar] [CrossRef]
- Hughes, S.L.; Schart, V.; Malcolmson, J.; Hogarth, K.A.; Martynowicz, D.M.; Tralman-Baker, E.; Patel, S.N.; Graether, S.P. The importance of size and disorder in the cryoprotective effects of dehydrins. Plant Physiol. 2013, 163, 1376–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Liu, M.; Chen, Y.; Wang, J.; Lu, C. Heterologous expression of the dehydrin-like protein gene AmCIP from Ammopiptanthus mongolicus enhances viability of Escherichia coli and tobacco under cold stress. Plant Growth Regul. 2016, 79, 71–80. [Google Scholar] [CrossRef]
- Hara, M.; Endo, T.; Kamiya, K.; Kameyama, A. The role of hydrophobic amino acids of K-segments in the cryoprotection of lactate dehydrogenase by dehydrins. J. Plant Physiol. 2017, 210, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Momma, M. Detection and Cryoprotective Activity of Dehydrin Proteins from Rice Bran and Soybean Whey. J. Nutr. Sci. Vitaminol. 2019, 65, S129–S133. [Google Scholar] [CrossRef]
- Palmer, S.R.; De Villa, R.; Graether, S.P. Sequence composition versus sequence order in the cryoprotective function of an intrinsically disordered stress-response protein. Protein Sci. 2019, 28, 1448–1459. [Google Scholar] [CrossRef]
- Ohkubo, T.; Kameyama, A.; Kamiya, K.; Kondo, M.; Hara, M. F-segments of Arabidopsis dehydrins show cryoprotective activities for lactate dehydrogenase depending on the hydrophobic residues. Phytochemistry 2020, 173, 112300. [Google Scholar] [CrossRef]
- Houde, M.; Daniel, C.; Lachapelle, M.; Allard, F.; Laliberté, S.; Sarhan, F. Immunolocalization of freezing-tolerance-associated proteins in the cytoplasm and nucleoplasm of wheat crown tissues. Plant J. 1995, 8, 583–593. [Google Scholar] [CrossRef]
- Wisniewski, M.; Webb, R.; Balsamo, R.; Close, T.J.; Yu, X.M.; Griffith, M. Purification, immunolocalization, cryoprotective, and antifreeze activity of PCA60: A dehydrin from peach (Prunus persica). Physiol. Plantarum 1999, 105, 600–608. [Google Scholar] [CrossRef]
- Bravo, L.A.; Gallardo, J.; Navarrete, A.; Olave, N.; Martínez, J.; Alberdi, M.; Close, T.J.; Corcuera, L.J. Cryoprotective activity of a cold-induced dehydrin purified from barley. Physiol. Plantarum 2003, 118, 262–269. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Rodrigo, M.J.; Lafuente, M.T.; Granell, A.; Zacarias, L. Dehydrin from citrus, which confers in vitro dehydration and freezing protection activity, is constitutive and highly expressed in the flavedo of fruit but responsive to cold and water stress in leaves. J. Agric. Food Chem. 2004, 52, 1950–1957. [Google Scholar] [CrossRef]
- Reyes, J.L.; Campos, F.; Wei, H.U.I.; Arora, R.; Yang, Y.; Karlson, D.T.; Covarrubias, A.A. Functional dissection of hydrophilins during in vitro freeze protection. Plant Cell Environ. 2008, 31, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.X. Effect of mixed macromolecular crowding agents on protein folding. Proteins 2008, 72, 1109–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, L.A.; Mooradally, A.W.; Zaslavsky, B.; Uversky, V.N.; Graether, S.P. Effect of an intrinsically disordered plant stress protein on the properties of water. Biophys. J. 2018, 115, 1696–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.W.; Kamlet, M.J. The solvatochromic comparison method. 2. The. alpha.-scale of solvent hydrogen-bond donor (HBD) acidities. J. Am. Chem. Soc. 1976, 98, 2886–2894. [Google Scholar] [CrossRef]
- Kamlet, M.J.; Taft, R.W. The solvatochromic comparison method. I. The. beta.-scale of solvent hydrogen-bond acceptor (HBA) basicities. J. Am. Chem. Soc. 1976, 98, 377–383. [Google Scholar] [CrossRef]
- Kamlet, M.J.; Abboud, J.L.; Taft, R.W. The solvatochromic comparison method. 6. The. pi.* scale of solvent polarities. J. Am. Chem. Soc. 1977, 99, 6027–6038. [Google Scholar] [CrossRef]
- Godoy, J.A.; Lunar, R.; Torres-Schumann, S.; Moreno, J.; Rodrigo, R.M.; Pintor-Toro, J.A. Expression, tissue distribution and subcellular localization of dehydrin TAS14 in salt-stressed tomato plants. Plant Mol. Biol. 1994, 26, 1921–1934. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.A.; Rebala, K.C.; Venkataraman, G.; Parida, A. A diurnally regulated dehydrin from Avicennia marina that shows nucleo-cytoplasmic localization and is phosphorylated by Casein kinase II in vitro. Plant Physiol. Biochem. 2009, 47, 701–709. [Google Scholar] [CrossRef]
- Lin, C.H.; Peng, P.H.; Ko, C.Y.; Markhart, A.H.; Lin, T.Y. Characterization of a novel Y2K-type dehydrin VrDhn1 from Vigna radiata. Plant Cell Physiol. 2012, 53, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Szabala, B.M.; Fudali, S.; Rorat, T. Accumulation of acidic SK 3 dehydrins in phloem cells of cold-and drought-stressed plants of the Solanaceae. Planta 2014, 239, 847–863. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Zhang, T.; Yang, X.; Li, D. Functional characterization of KS-type dehydrin ZmDHN13 and its related conserved domains under oxidative stress. Sci. Rep. 2017, 7, 7361. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Shinoda, Y.; Tanaka, Y.; Kuboi, T. DNA binding of citrus dehydrin promoted by zinc ion. Plant Cell Environ. 2009, 32, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Fujinaga, M.; Kuboi, T. Metal binding by citrus dehydrin with histidine-rich domains. J. Exp. Bot. 2005, 56, 2695–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Plant/Author | LEA Protein | Expression System | Applied Stress | Observed Phenotype | Post-Stress Change without Dehydrin | Post-Stress Change with Dehydrin |
|---|---|---|---|---|---|---|
| Vigna Unguiculata (L.) Walp. [11] | Unnamed Dehydrin: Present in line 1393-2-11 | Naturally occurring | Seeds grown for 27 days at 15 °C | Percent emergence | 13% | 50% |
| Seeds incubated for 12 h at 15 °C | Electrolyte leakage | 41.3 mS/m·g | 27.1 mS/m·g | |||
| Oryza sativa L. [12] | PMA80, wheat dehydrin | pGHNC5, Act1 promoter | Six-week old plants grown with 200 mM NaCl for nine days | Electrolyte leakage from leaves | +342% from day 3–7 | +155% from day 3–7 |
| Height of plant after recovery | 48.2 ± 2.8 cm | 63.1 cm | ||||
| Fresh/dry shoot weight after recovery | Fresh: 3.37 g Dry: 0.67 g | Fresh: 4.8 g Dry: 1.025 g | ||||
| Six-week old plants denied water for eight days | Height of plant, after eight days of recovery | 47.8 ± 2.4 cm | 63.6 cm | |||
| Fresh/dry shoot weight, after eight days of recovery | Fresh 3.23 ± 0.06 g Dry: 0.7 ± 0.01 g | Fresh: 4.8 g Dry: 1.053 g | ||||
| Nicotiana tabacum L. [13] | Citrus unshiu dehydrin (CuCOR19) | pBI121, CaMV 35S promoter | Seedlings incubated at −4 °C for three hours | Electrolyte leakage expressed as percent of total | 19.2 ± 2.6% | 12.3 ± 2.6% |
| MDA production | 4.6 nmol/g fresh weight | 2.7 nmol/g fresh weight | ||||
| Grown at 15 °C | Germination | 72% (GD50 = 11.9 days) | 88% (GD50 = 8.1 days) | |||
| Increase in seedling fresh weight over 15 days (day 6 to 21 after sowing) | 3.9 mg/plant | 6.7 mg/plant | ||||
| Nicotiana tabacum NC89 [14] | Brassica juncea dehydrins (BjDHN2/BjDHN3) | pBI121, CaMV 35S promoter | Leaf disks from four-week old plants floated on MS (⅛) solution at 4 °C for seven days | Chlorophyll content | 43% mg/g of dry weight | BjDHN2: 60% mg/g·DW BjDHN3: 69% mg/g·DW |
| Grown with 150 mM NaCl for 10 days followed by two weeks of recovery | Relative water content | 75% | BjDHN2: 92% BjDHN3: 90% | |||
| Electrolyte leakage | 16% | BjDHN2: 3% BjDHN3: 7% | ||||
| MDA content | 4.75 mmol/g of fresh weight | BjDHN2 3.5 mmol/g·FW BjDHN3: 3.5 mmol/g·FW | ||||
| Nicotiana tabacum L. cv NC89 [15] | Maize dehydrin, ZmDHN2b | pBI121, CaMV 35S promoter | Germinated at 15 °C | Germination | Began germinating on day 11. By day 30 <60% had germinated. | Began germinating on day 7. By day 30, 80% had germinated. |
| Germinated and grown at 25 °C for four days. Incubated at 15 °C for two weeks. | Root length | <1.0 cm | >1.8 cm | |||
| Six-week old plants incubated at 4 °C for 24 h | MDA content | 2.5-fold increase over 24 h | Two-fold increase over 24 h | |||
| Electrolyte leakage | 3.14-fold increase over 24 h | 2.3-fold increase over 24 h | ||||
| Nicotiana tabacum [16] | Prunus mume dehydrins (PmLEA10, PmLEA19, PmLEA20, PmLEA29) | pEarleyGate203, CaMV 35S promoter | Two-month old plants incubated at 4 °C for 24 h | Increase in electrolyte leakage relative to untreated plants | 1.9% | PmLEA10: 1.3% PmLEA19: 1.15% PmLEA20: 0.75% PmLEA29: 1.2% |
| Increase in MDA content relative to untreated plants | 3.3% | PmLEA10: 1.6% PmLEA19: 1.8% PmLEA20: 0.75% PmLEA29: 1.9% | ||||
| Two-month old plants denied water for 15 days | Increase in electrolyte leakage relative to untreated plants | 1.5% | PmLEA10: 1.3% PmLEA19: 1.15% PmLEA20: 0.8% PmLEA29: 1.2% | |||
| Increase in MDA content relative to untreated plants | 2% | PmLEA10: 1.5% PmLEA19: 1.8% PmLEA20: 0.75% PmLEA29: 1.25% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, M.A.; Graether, S.P. The Disordered Dehydrin and Its Role in Plant Protection: A Biochemical Perspective. Biomolecules 2022, 12, 294. https://doi.org/10.3390/biom12020294
Smith MA, Graether SP. The Disordered Dehydrin and Its Role in Plant Protection: A Biochemical Perspective. Biomolecules. 2022; 12(2):294. https://doi.org/10.3390/biom12020294
Chicago/Turabian StyleSmith, Margaret A., and Steffen P. Graether. 2022. "The Disordered Dehydrin and Its Role in Plant Protection: A Biochemical Perspective" Biomolecules 12, no. 2: 294. https://doi.org/10.3390/biom12020294