
Broccoli Myrosinase cDNA Expression in Escherichia coli and Saccharomyces cerevisiae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining Recombinant Clones
2.2. Expression of the cDNA Encoding Myrosinase in E. coli and S. cerevisiae
2.3. Purification of Myrosinase Enzyme
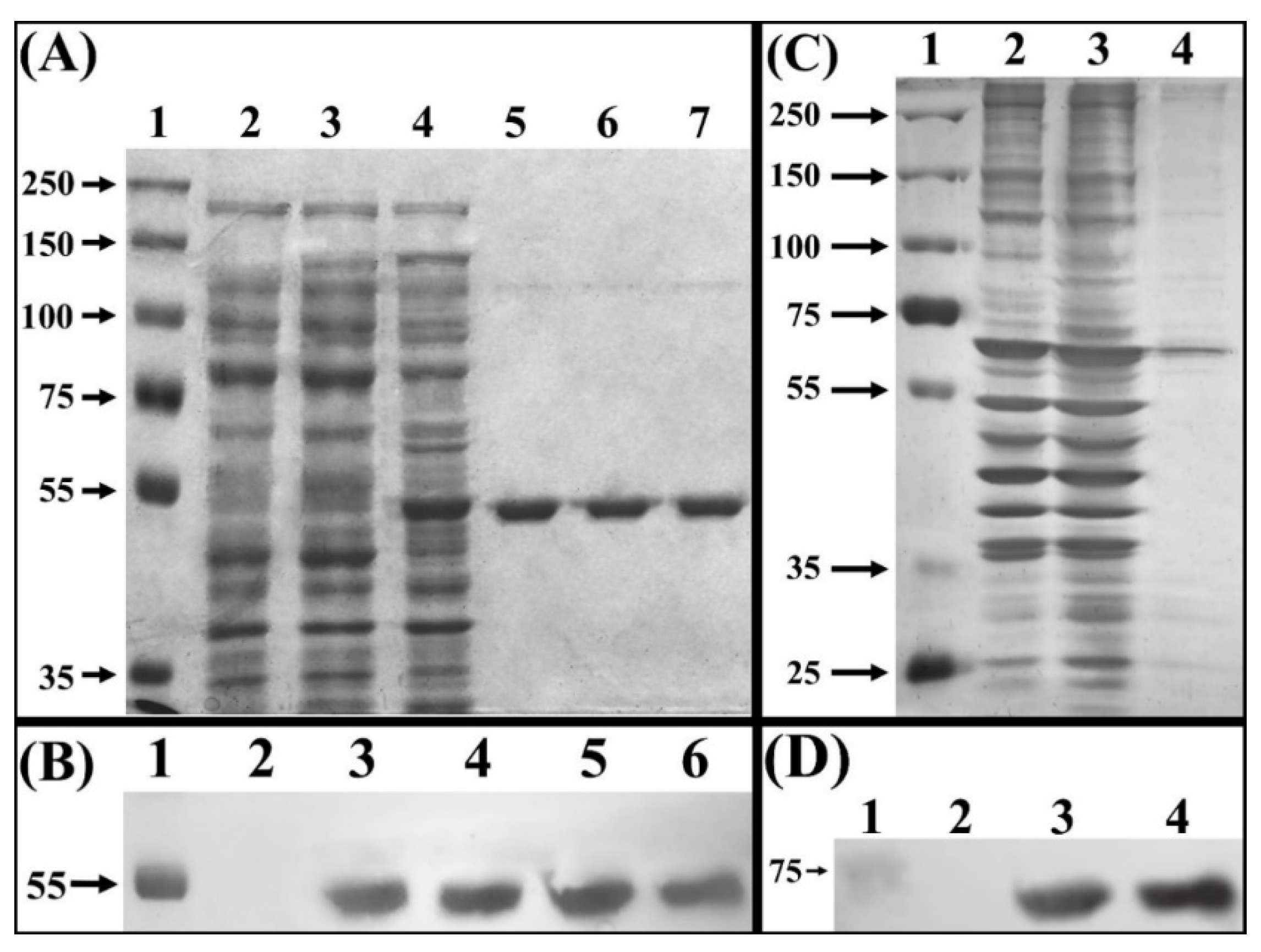
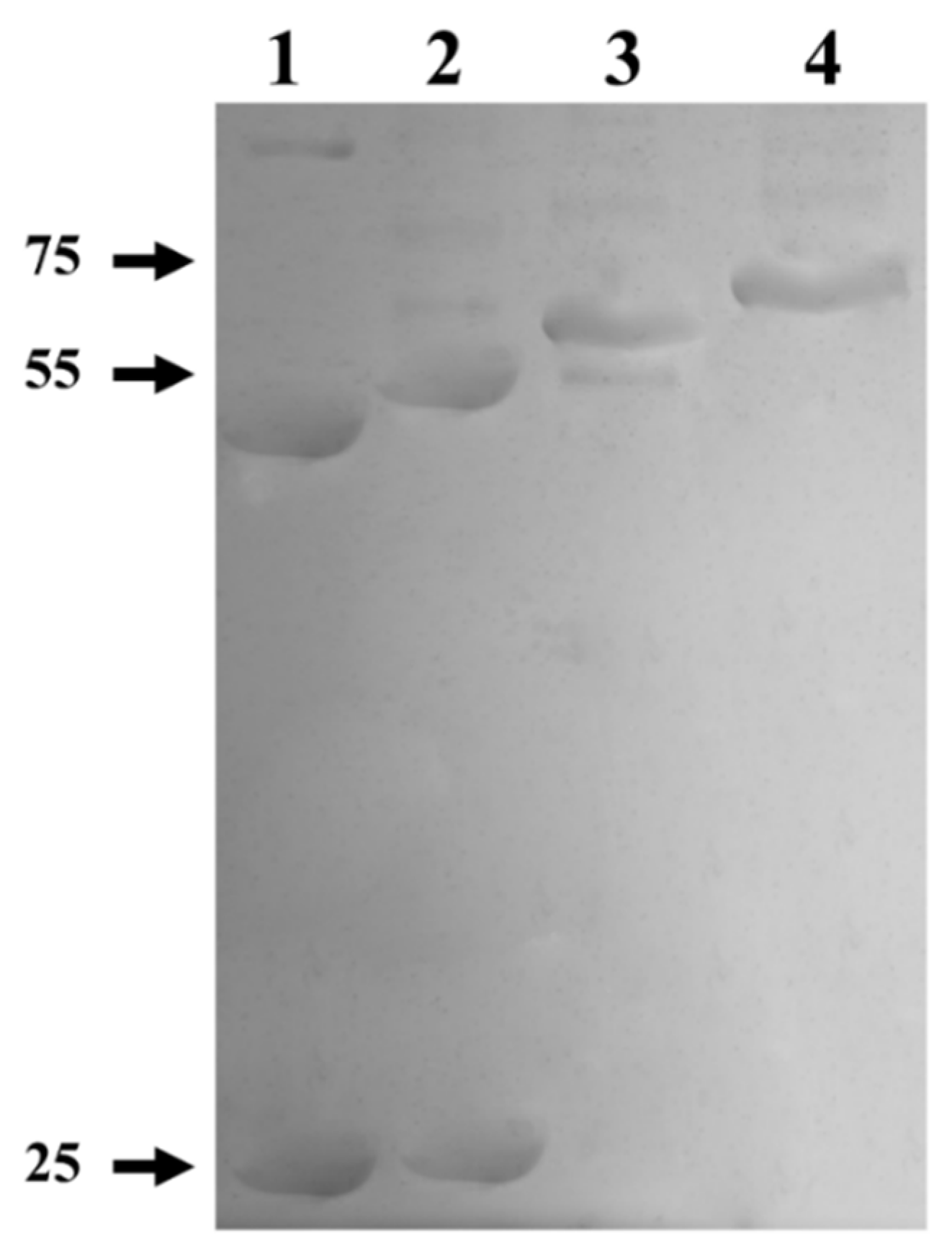
2.4. SDS-PAGE and Western Blot
2.5. Determination of the Molecular Mass of Native Myrosinase
2.6. Enzymatic Activity of Recombinant Myrosinase
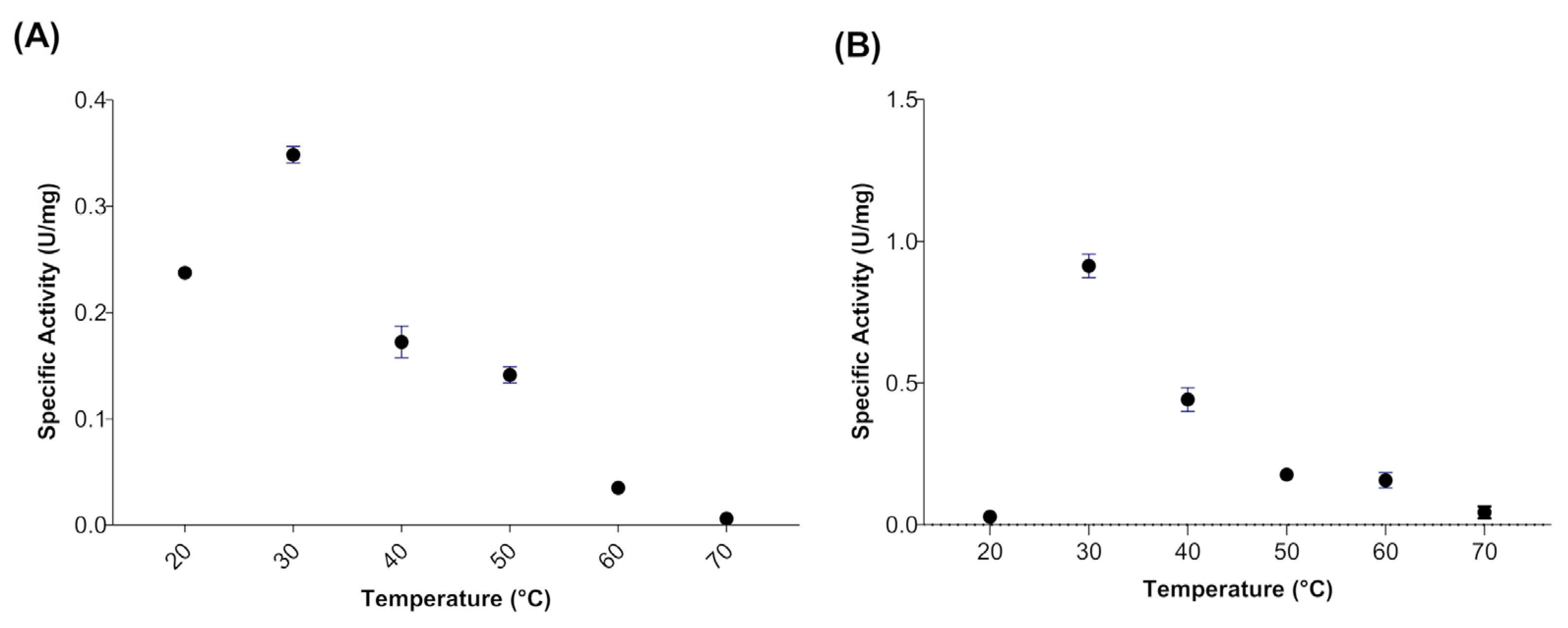
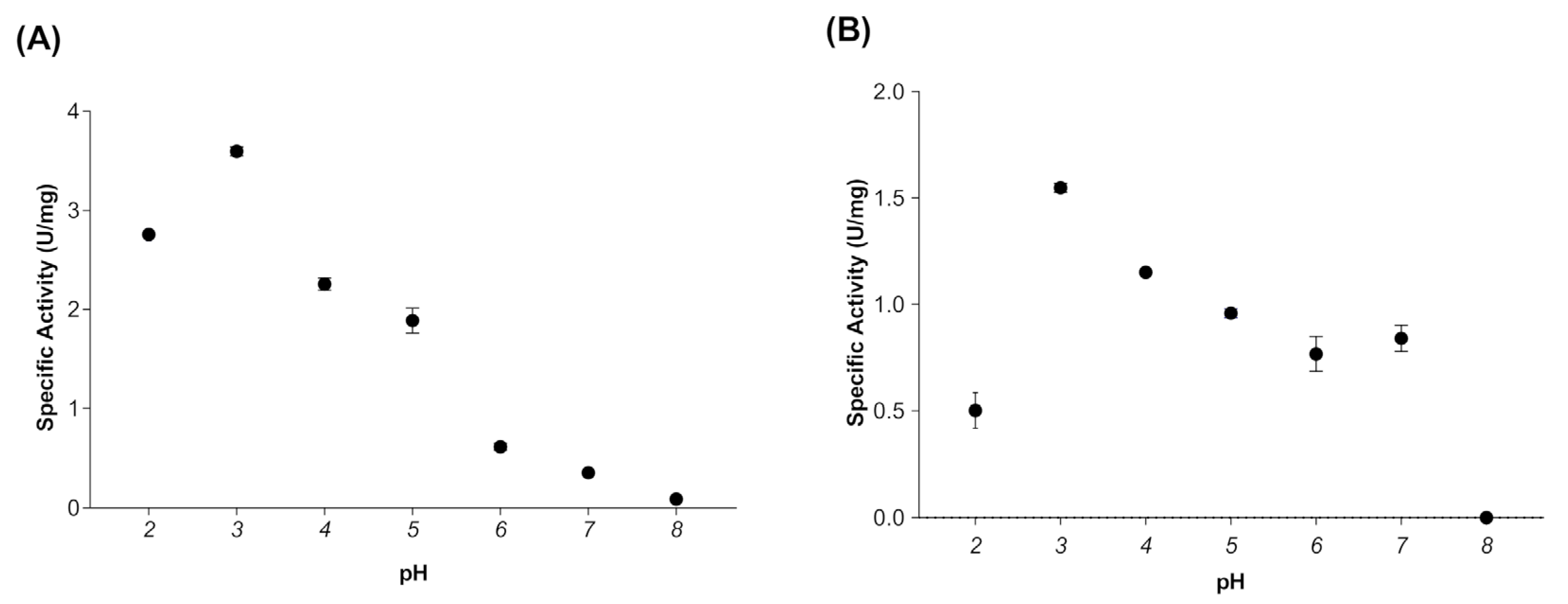
2.7. Effect of Temperature and pH on Myrosinase Activity
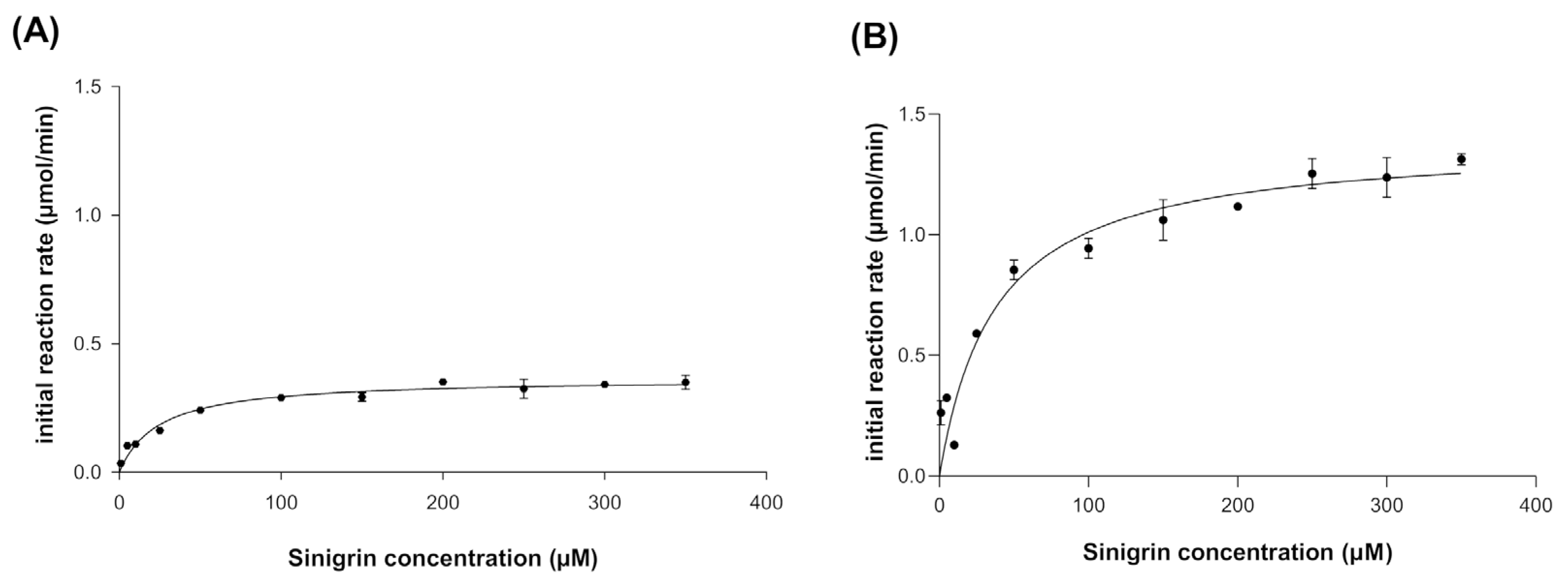
2.8. Kinetic Characterization of Recombinant Myrosinase
2.9. Deglycosylation and Proteolytic Digestion of Recombinant Myrosinase
2.10. Statistical Analysis
3. Results
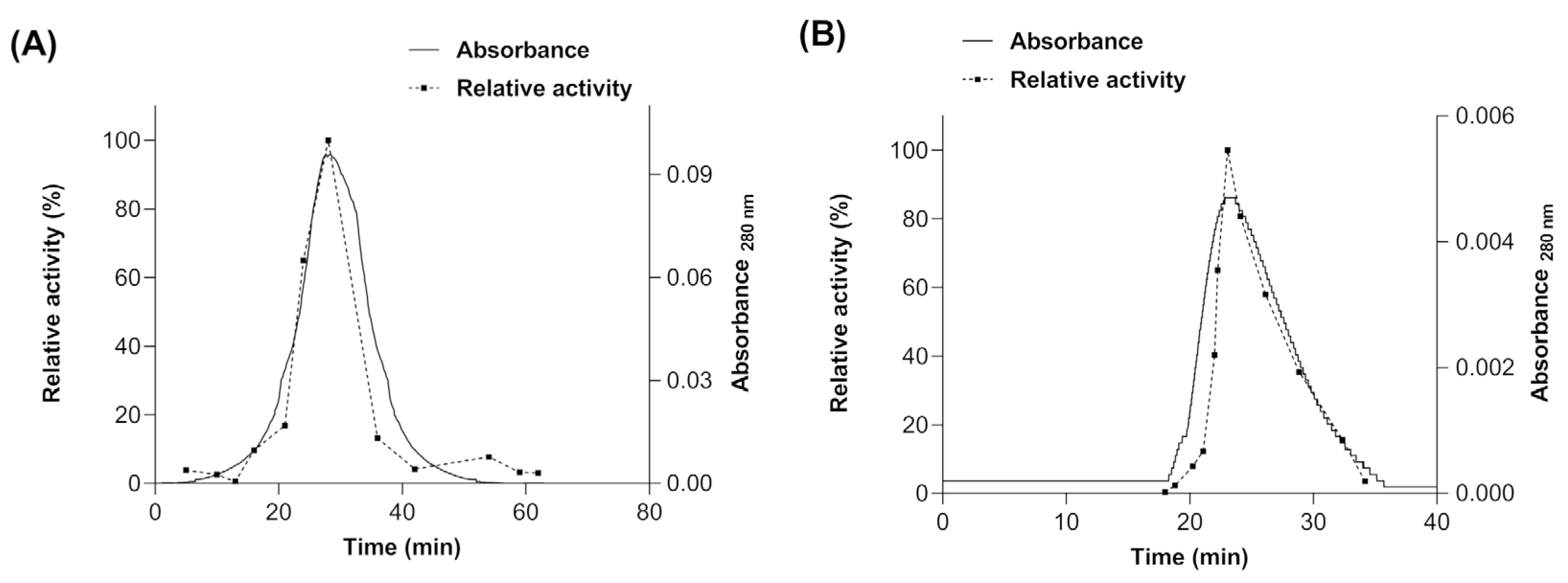
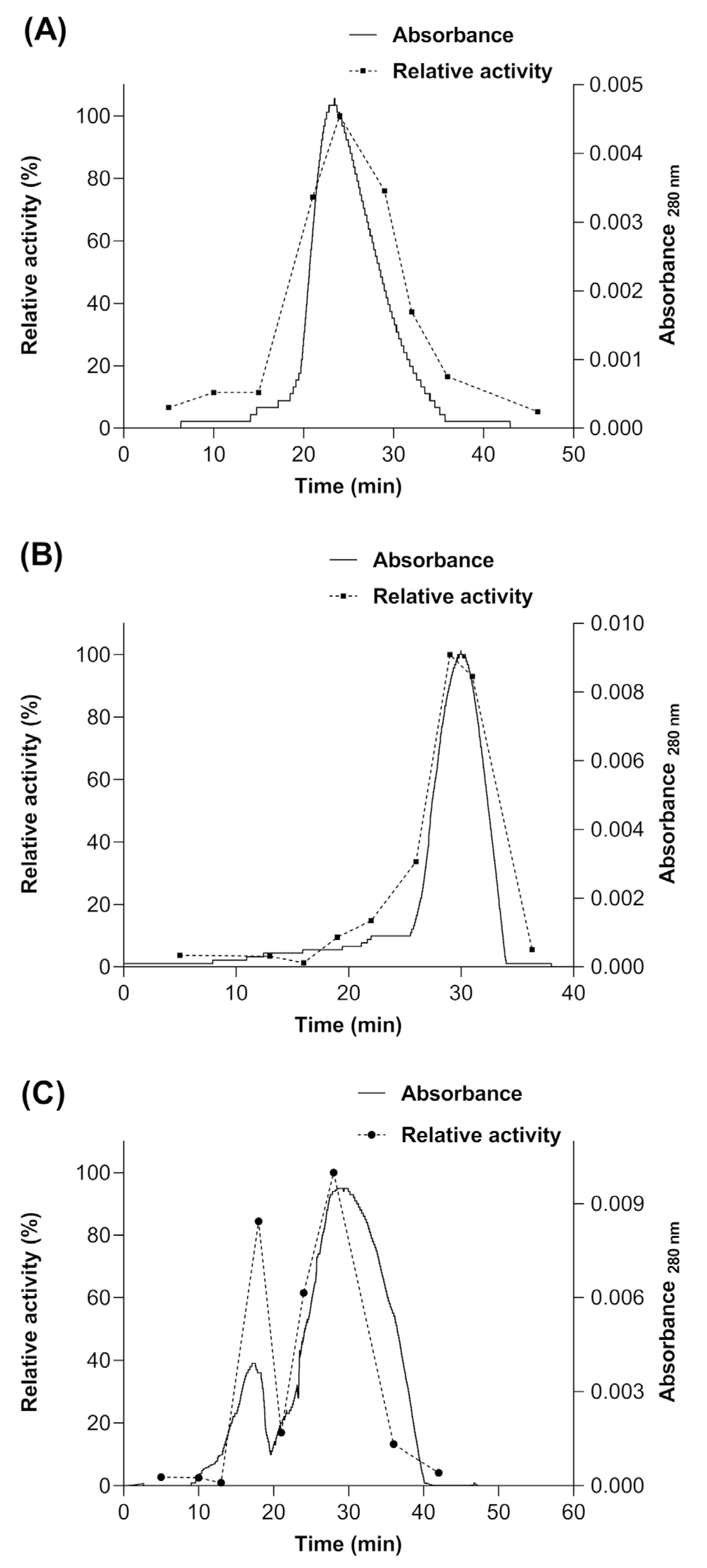
3.1. Myrosinase cDNA Expression and Recombinant Protein Purification
3.2. Effect of Temperature and pH on Myrosinase Activity
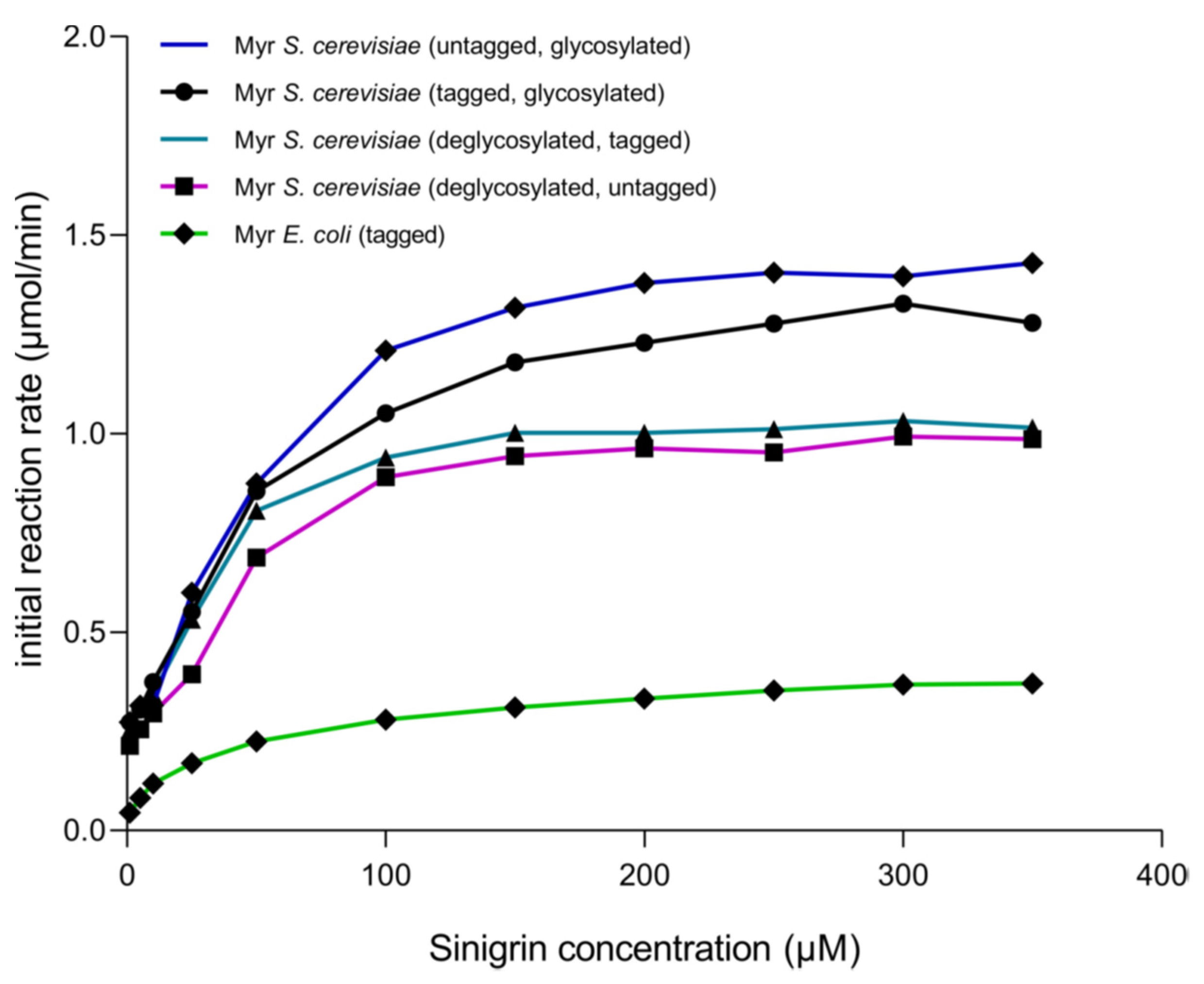
3.3. Kinetic Behavior of Recombinant Myrosinase
3.4. Determination of the Molecular Mass of Native Myrosinases
3.5. Proteolytic Digestion and Deglycosylation of Myrosinase Produced in S. cerevisiae MGY70-myr
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhat, R.; Vyas, D. Myrosinase: Insights on structural, catalytic, regulatory, and environmental interactions. Crit. Rev. Biotechnol. 2019, 39, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef] [PubMed]
- Latté, K.P.; Appel, K.E.; Lampen, A. Health benefits and possible risks of broccoli—An overview. Food Chem. Toxicol. 2011, 49, 3287–3309. [Google Scholar] [CrossRef] [PubMed]
- Backenköhler, A.; Eisenschmidt, D.; Schneegans, N.; Strieker, M.; Brandt, W.; Wittstock, U. Iron is a centrally bound cofactor of specifier proteins involved in glucosinolate breakdown. PLoS ONE 2018, 13, e0205755. [Google Scholar] [CrossRef] [PubMed]
- Nintemann, S.J.; Hunziker, P.; Andersen, T.G.; Schulz, A.; Burow, M.; Halkier, B.A. Localization of the glucosinolate biosynthetic enzymes reveals distinct spatial patterns for the biosynthesis of indole and aliphatic glucosinolates. Physiol. Plant. 2017, 163, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ballesta, M.D.C.; Carvajal, M. Myrosinase in Brassicaceae: The most important issue for glucosinolate turnover and food quality. Phytochem. Rev. 2015, 14, 1045–1051. [Google Scholar] [CrossRef]
- Liang, H.; Yuan, Q.; Xiao, Q. Effects of metal ions on myrosinase activity and the formation of sulforaphane in broccoli seed. J. Mol. Catal. B Enzym. 2006, 43, 19–22. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Mahn, A.; Castillo, A. Potential of sulforaphane as a natural immune system enhancer: A review. Molecules 2021, 26, 752. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Isothiocyanates: An overview of their antimicrobial activity against human infections. Molecules 2018, 23, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, R.; Finiguerra, M.G.; Rossi, A.A.A.; Palmieri, S. Isolation and biochemical characterization of a basic myrosinase from ripe Crambe abyssinica seeds, highly specific for epi-progoitrin. J. Agric. Food Chem. 2003, 51, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kushad, M.M. Purification and characterization of myrosinase from horseradish (Armoracia rusticana) roots. Plant Physiol. Biochem. 2005, 43, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Van Eylen, D.; Indrawati; Hendrickx, M.E.; Van Loey, A. Temperature and pressure stability of mustard seed (Sinapis alba L.) myrosinase. Food Chem. 2006, 97, 263–271. [Google Scholar] [CrossRef]
- Zhou, C.; Tokuhisa, J.; Bevan, D.; Esen, A. Properties of b-thioglucoside hydrolases (TGG1 and TGG2) from leaves of Arabidopsis thaliana. Plant Sci. 2012, 191, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Kaur, T.; Khajuria, M.; Vyas, R.; Vyas, D. Purification and characterization of a novel redox-regulated isoform of myrosinase (β-Thioglucoside Glucohydrolase) from Lepidium latifolium L. J. Agric. Food Chem. 2015, 63, 10218–10226. [Google Scholar] [CrossRef] [PubMed]
- Ghawi, S.K.; Methven, L.; Rastall, R.A.; Niranjan, K. Thermal and high hydrostatic pressure inactivation of myrosinase from green cabbage: A kinetic study. Food Chem. 2012, 131, 1240–1247. [Google Scholar] [CrossRef]
- El-Shora, H.; El-Shobaky, A.; El-Atrozy, M. Activity of purified bacterial myrosinase and its essential residues. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Francis, F.; Lognay, G.; Wathelet, J.-P.; Haubruge, E. Characterisation of aphid myrosinase and degradation studies of glucosinolates. Arch. Insect Biochem. Physiol. 2002, 50, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, W.P.; Cottaz, S.; Driguez, H.; Iori, R.; Palmieri, S.; Henrissat, B. The crystal structures of Sinapis alba myrosinase and a covalent glycosyl–Enzyme intermediate provide insights into the substrate recognition and active-site machinery of an S-glycosidase. Structure 1997, 5, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okunade, O.A.; Ghawi, S.K.; Methven, L.; Niranjan, K. Thermal and pressure stability of myrosinase enzymes from black mustard (Brassica nigra L. W.D.J. Koch. var. nigra), brown mustard (Brassica juncea L. Czern. var. juncea) and yellow mustard (Sinapsis alba L. subsp. maire) seeds. Food Chem. 2015, 187, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Ludikhuyze, L.; Rodrigo, L.; Hendrickx, M. The activity of myrosinase from broccoli (Brassica oleracea L. cv. italica): Influence of intrinsic and extrinsic factors. J. Food Prot. 2000, 63, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; Dekker, M. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219. [Google Scholar] [CrossRef] [PubMed]
- Mahn, A.; Angulo, A.; Cabañas, F. Purification and Characterization of Broccoli (Brassica oleracea var oleracea) Myrosinase (β-Thioglucosidase Glucohydrolase). J. Agric. Food Chem. 2014, 62, 11666–11671. [Google Scholar] [CrossRef] [PubMed]
- Román, J.; Castillo, A.; Cottet, L.; Mahn, A. Kinetic and structural study of broccoli myrosinase and its interaction with different glucosinolates. Food Chem. 2018, 254, 87–94. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkmen, M. Production of disulfide-bonded proteins in Escherichia coli. Protein Expr. Purif. 2012, 82, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosano, G.L.; Morales, E.S.; Ceccarelli, E.A. New tools for recombinant protein production in Escherichia coli: A 5-year update. Protein Sci. 2019, 28, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Geymonat, M.; Spanos, A.; Sedgwick, S.G. Saccharomyces cerevisiae autoselection system for optimised recombinant protein expression. Gene 2007, 399, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Husebye, H.; Arzt, S.; Burmeister, W.; Hartel, F.; Brandt, A.; Rossiter, J.; Bones, A. Crystal structure at 1.1 Å resolution of an insect myrosinase from Brevicoryne brassicae shows its close relationship to b-glucosidases. Insect Biochem. Mol. Biol. 2005, 35, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Halkier, B.A. Functional Expression and Characterization of the Myrosinase MYR1 from Brassica napus in Saccharomyces cerevisiae. Protein Expr. Purif. 1999, 17, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Nong, H.; Zhang, J.-M.; Li, D.-Q.; Wang, M.; Sun, X.-P.; Zhu, Y.J.; Meijer, J.; Wang, Q.-H. Characterization of a Novel β-thioglucosidase CpTGG1 in Carica papaya and its Substrate-dependent and Ascorbic Acid-independent O-β-glucosidase Activity. J. Integr. Plant Biol. 2010, 52, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D. Plasmids and their usefulness in molecular cloning. In Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 1, pp. 51–52. [Google Scholar]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Geymonat, M.; Spanos, A.; Sedgwick, S. Production of mitotic regulators using an autoselection system for protein expression in budding yeast. In Mitosis: Methods and Protocols; Humana Press: Edinburg, UK, 2009; Volume 545, pp. 63–80. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chang, F.; Zhang, X.; Pan, Y.; Lu, Y.; Fang, W.; Fang, Z.; Xiao, Y. Light induced expression of β-glucosidase in Escherichia coli with autolysis of cell. BMC Biotechnol. 2017, 17, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero, M.E.; Barriuso, J.; Medrano, F.J.; Prieto, A.; Martínez, M.J. Heterologous expression of a fungal sterol esterase/lipase in different hosts: Effect on solubility, glycosylation and production. J. Biosci. Bioeng. 2015, 120, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.; Surolia, A.; Mukhopadhyay, C. Dynamics simulation of soybean agglutinin (SBA) dimer reveals the impact of glycosylation on its enhanced structural stability. Carbohydr. Res. 2016, 428, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Qi, Y.; Im, W. Effects of N-glycosylation on protein conformation and dynamics: Protein Data Bank analysis and molecular dynamics simulation study. Sci. Rep. 2015, 5, 8926. [Google Scholar] [CrossRef] [PubMed]
- Pontoppidan, B.; Ekbom, B.; Eriksson, S.; Meijer, J. Purification and characterization of myrosinase from the cabbage aphid (Brevicoryne brassicae), a brassica herbivore. Eur. J. Biochem. 2001, 268, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.; Bridges, M.; Bones, A.; Cole, R.; Rossiter, J. Purification and characterization of a non-plant myrosinase from the cabbage aphid Brevicoryne brassicae (L.). Insect Biochem. Mol. Biol. 2000, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lenman, M.; Rodin, J.; Josefsson, L.-G.; Rask, L. Immunological characterization of rapeseed myrosinase. Eur. J. Biochem. 1990, 194, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.; Chakrabarty, R.; Bejai, S.; Zhang, J.; Rask, L.; Meijer, J. Myrosinases from root and leaves of Arabidopsis thaliana have different catalytic properties. Phytochemistry 2009, 70, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S.; Thamilarasan, S.K.; Park, J.-I.; Chung, M.-Y.; Nou, I.-S. Molecular modeling of myrosinase from Brassica oleracea: A structural investigation of sinigrin interaction. Genes 2015, 6, 1315–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Härtel, F.V.; Brandt, A. Characterization of a Brassica napus myrosinase expressed and secreted by Pichia pastoris. Protein Expr. Purif. 2002, 24, 221–226. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Total Protein Concentration (mg/mL) | Total Activity * (U) | Specific Activity (U/mg) | Accumulated Yield (%) |
|---|---|---|---|---|
| Myrosinase produced in E. coli | ||||
| Crude extract | 3.526 | 5.121 | 1.452 | 100 |
| Total proteins eluted with wash buffer | 0.963 | 4.922 | 5.112 | 97.5 |
| Elution of proteins retained in the column | 0.721 | 4.619 | 6.407 | 90.2 |
| Myrosinase produced in S. cerevisiae | ||||
| Crude extract | 5.271 | 5.639 | 1.070 | 100 |
| Total proteins eluted with wash buffer | 3.210 | 3.848 | 1.199 | 68.2 |
| Elution of proteins retained in the column | 0.512 | 2.706 | 5.287 | 48.0 |
| Myrosinase From: | Km (μM) | Vmax (μmol/min) | kcat (s−1) | kcat/Km (s−1 μM−1) |
|---|---|---|---|---|
| S. cerevisiae (glycosylated, with GST tag) | 37.37 ± 7.51 | 1.388 ± 0.066 | 27.77 ± 1.323 | 0.743 |
| S. cerevisiae (glycosylated, without GST tag) | 25.92 ± 4.563 | 1.079 ± 0.047 | 21.58 ± 0.801 | 0.833 |
| S. cerevisiae (deglycosylated, with GST tag) | 17.44 ± 2.828 | 1.106 ± 0.034 | 22.11 ± 0.6796 | 1.268 |
| S. cerevisiae (deglycosylated, without GST tag) | 35.82 ± 5.767 | 1.595 ± 0.060 | 31.9 ± 1.2 | 0. 891 |
| E. coli | 24.32 ± 3.47 | 0.364 ± 0.011 | 7.288 ± 0.2152 | 0.300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curiqueo, C.; Mahn, A.; Castillo, A. Broccoli Myrosinase cDNA Expression in Escherichia coli and Saccharomyces cerevisiae. Biomolecules 2022, 12, 233. https://doi.org/10.3390/biom12020233
Curiqueo C, Mahn A, Castillo A. Broccoli Myrosinase cDNA Expression in Escherichia coli and Saccharomyces cerevisiae. Biomolecules. 2022; 12(2):233. https://doi.org/10.3390/biom12020233
Chicago/Turabian StyleCuriqueo, Carolina, Andrea Mahn, and Antonio Castillo. 2022. "Broccoli Myrosinase cDNA Expression in Escherichia coli and Saccharomyces cerevisiae" Biomolecules 12, no. 2: 233. https://doi.org/10.3390/biom12020233