
Spreading of Aggregated α-Synuclein in Sagittal Organotypic Mouse Brain Slices


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Organotypic Brain Slices
2.3. Lactate Dehydrogenase (LDH) Assay
2.4. Preparation of Collagen Hydrogels and α-Syn Proteins
2.5. Immunohistochemistry
2.6. Release Experiments
2.7. Western Blotting
2.8. Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
2.9. Data Analysis and Statistics
3. Results
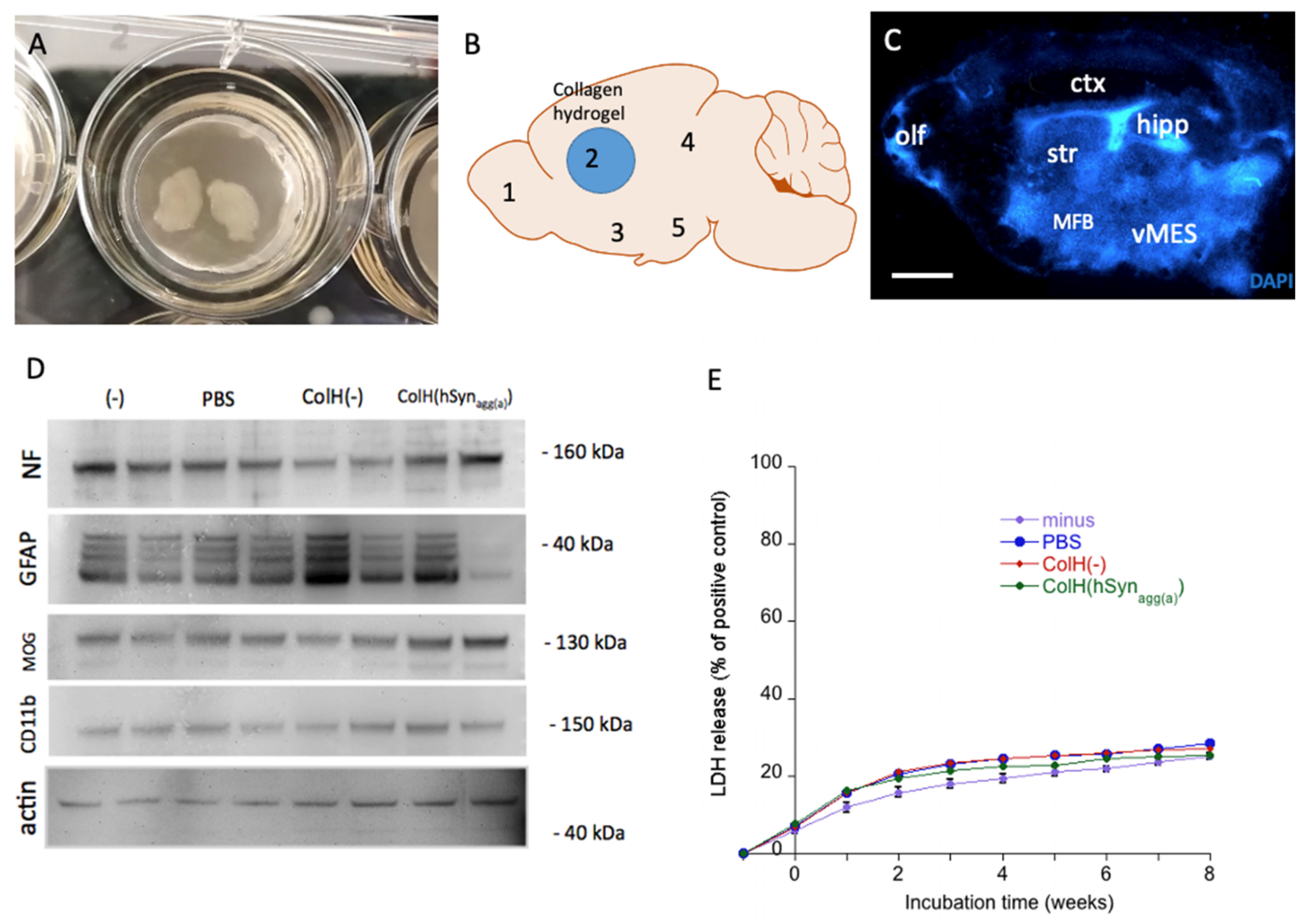
3.1. Viability of Organotypic Brain Slices
3.2. α-Syn Loaded in Collagen Hydrogels
3.3. Characterization of α-Syn Proteins by Western Blotting
3.4. Release of α-Syn from Collagen Hydrogels
3.5. qRT-PCR of Endogenous α-Syn Expression
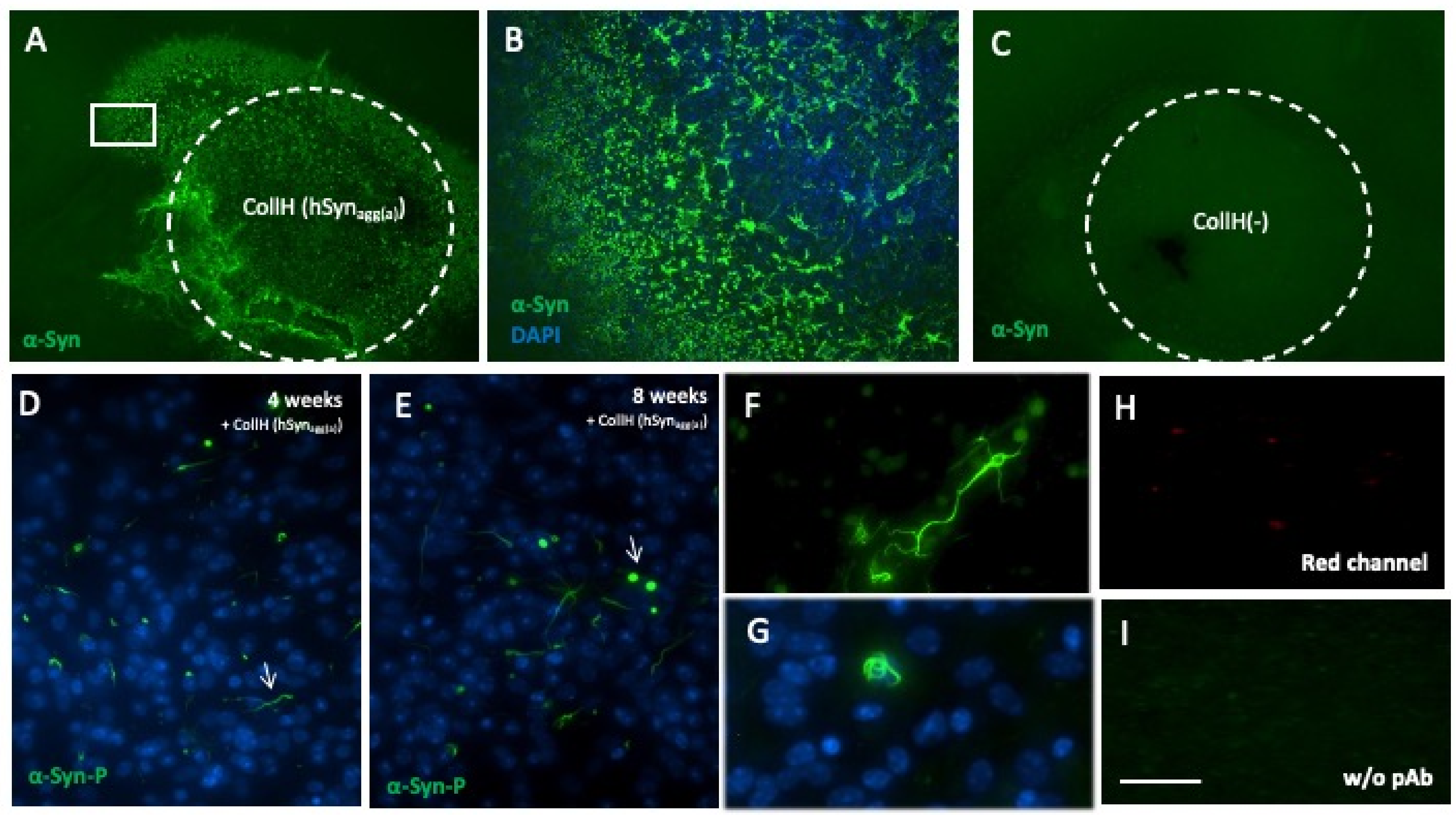
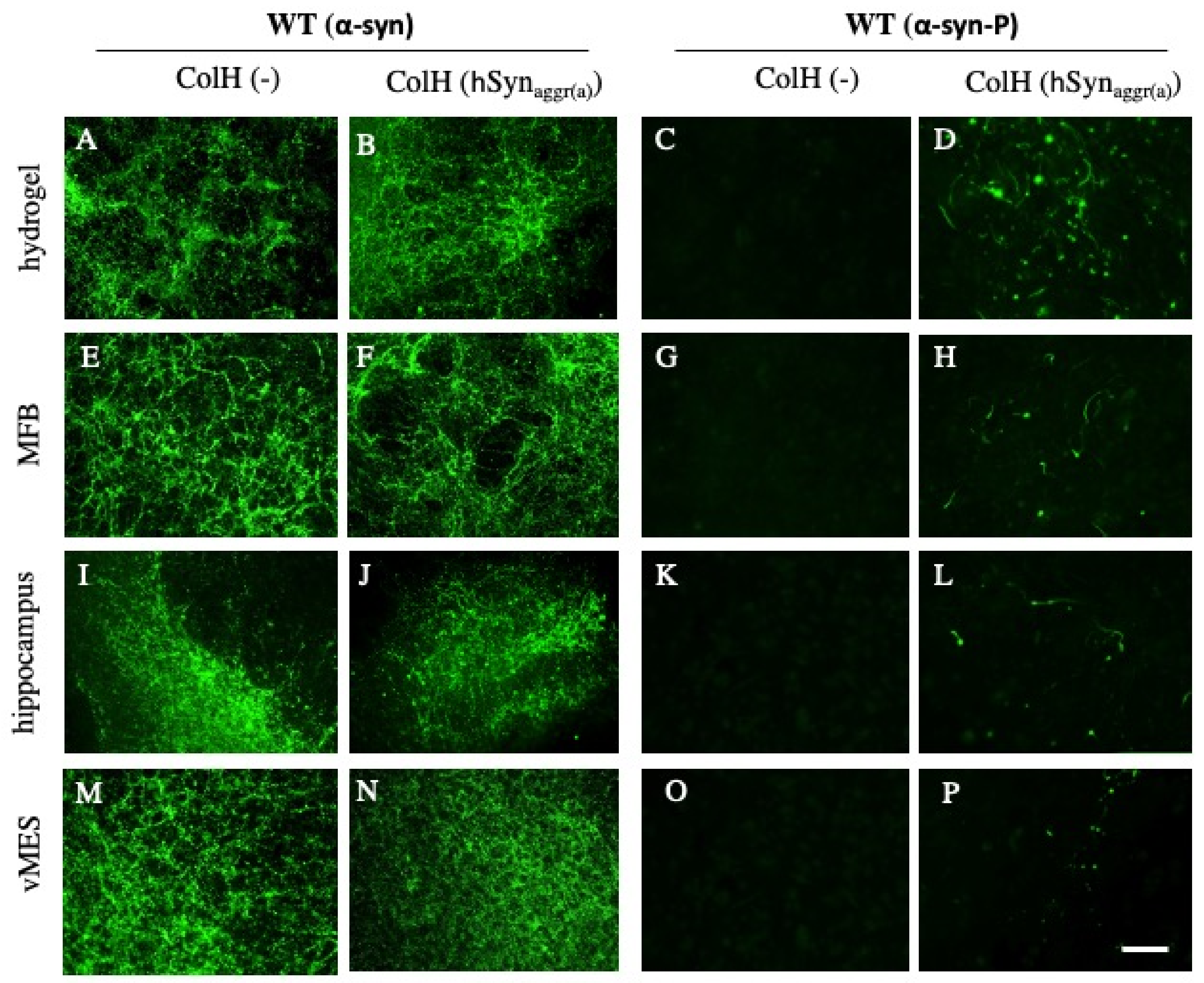
3.6. Spreading of α-Syn in Slices Taken from Wild-Type Mice
3.7. Spreading of α-Syn in Slices Taken from Transgenic PLP Mice
3.8. Quantification of α-Syn Aggregation
3.9. Localization of the α-Syn Aggregates
3.10. Effect of α-Syn Aggregates on Dopaminergic Neuron Survival
4. Discussion
- (a)
- Organotypic brain slices to study α-syn spreading
- (b)
- Collagen hydrogels for application of α-syn PFFs
- (c)
- α-syn proteins tested in this study
- (d)
- Characterization of α-syn immunostainings
- (e)
- Spreading of α-syn in slices taken from wild type mice
- (f)
- Spreading of α-syn in TG PLP organotypic brain slices
- (g)
- Cellular localization of α-syn aggregates
- (h)
- Effect of α-syn aggregation on dopaminergic neurons
- (i)
- Mechanism of α-syn aggregation and spreading
- (j)
- Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bendor, J.T.; Logan, T.P.; Edwards, R.H. The function of α-synuclein. Neuron 2013, 79, 1044–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillantini, M.; Schmidt, M.; Lee, V.; Trojanowski, J.; Jakes, R.; Goedert, M. α-Synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. α-Synuclein in filamentous inclusions of Lewy bodies from Parkinson’s disease and dementia with Lewy bodies. Proc. Natl. Acad. Sci. USA 1998, 95, 6469–6473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundin, P.; Melki, R.; Kopito, R. Prion-like transmission of protein aggregates in neurodegenerative diseases. Nat. Rev. Mol. Cell Biol. 2010, 11, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Kordower, J.H.; Chu, Y.; Hauser, R.A.; Freeman, T.B.; Olanow, C.W. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat. Med. 2008, 14, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Englund, E.; Holton, J.L.; Soulet, D.; Hagell, P.; Lees, A.J.; Lashley, T.; Quinn, N.P.; Rehncrona, S.; Björklund, A.; et al. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest host-to-graft disease propagation. Nat. Med. 2008, 14, 501–503. [Google Scholar] [CrossRef] [PubMed]
- Danzer, K.M.; Krebs, S.K.; Wolff, M.; Birk, G.; Hengerer, B. Seeding induced by α-synuclein oligomers provides evidence for spreading of α-synuclein pathology. J. Neurochem. 2009, 111, 192–203. [Google Scholar] [CrossRef]
- Freundt, E.C.; Maynard, N.; Clancy, E.; Roy, S.; Bousset, L.; Sourigues, Y.; Covert, M.; Melki, R.; Kirkegaard, K.; Brahic, M. Neuron-to-neuron transmission of α-synuclein fibrils through axonal transport. Ann. Neurol. 2012, 72, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Mougenot, A.-L.; Nicot, S.; Bencsik, A.; Morignat, E.; Verchère, J.; Lakhdar, L.; Legastelois, S.; Baron, T. Prion-like acceleration of a synucleinopathy in a transgenic mouse model. Neurobiol. Aging 2012, 33, 2225–2228. [Google Scholar] [CrossRef]
- Ellemberg, D.; Henry, L.C.; Macciocchi, S.N.; Guskiewicz, K.M.; Broglio, S.P. Advances in Sport Concussion Assessment: From Behavioral to Brain Imaging Measures. J. Neurotrauma 2009, 26, 2365–2382. [Google Scholar] [CrossRef] [PubMed]
- Luk, K.C.; Kehm, V.M.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M. Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice. J. Exp. Med. 2012, 209, 975–988. [Google Scholar] [CrossRef] [Green Version]
- Luk, K.C.; Kehm, V.; Carroll, J.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M.-Y. Pathological α-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science 2012, 338, 949–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humpel, C. Neuroscience forefront review Organotypic brain slice cultures: A review. Neuroscience 2015, 305, 86–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfarrash, S.; Jensen, P. Organotypic hippocampal slices, an emerging tool to model synucleinopathies. Neural Regen. Res. 2021, 16, 999–1000. [Google Scholar]
- Elfarrash, S.; Jensen, N.M.; Ferreira, N.; Betzer, C.; Thevathasan, J.V.; Diekmann, R.; Adel, M.; Omar, N.M.; Boraie, M.Z.; Gad, S.; et al. Organotypic slice culture model demonstrates inter-neuronal spreading of alphα-synuclein aggregates. Acta Neuropathol. Commun. 2019, 7, 1–16. [Google Scholar] [CrossRef]
- Roux, A.; Wang, X.; Becker, K.; Ma, J. Modeling α-Synucleinopathy in Organotypic Brain Slice Culture with Preformed α-Synuclein Amyloid Fibrils. J. Parkinsons Dis. 2020, 10, 1397–1410. [Google Scholar] [CrossRef]
- Shrivastava, A.N.; Bousset, L.; Renner, M.; Redeker, V.; Savistchenko, J.; Triller, A.; Melki, R. Differential Membrane Binding and Seeding of Distinct α-Synuclein Fibrillar Polymorphs. Biophys. J. 2020, 118, 1301–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Shaikh, M.A.; Meymand, E.S.; Zhang, B.; Luk, K.C.; Trojanowski, J.Q.; Lee VM, Y. Neuronal activity modulates alphα-synuclein aggregation and spreading in organotypic brain slice cultures and in vivo. Acta Neuropathol. 2020, 140, 831–849. [Google Scholar] [CrossRef]
- Barth, M.; Bacioglu, M.; Schwarz, N.; Novotny, R.; Brandes, J.; Welzer, M.; Mazzitelli, S.; Häsler, L.M.; Schweighauser, M.; Wuttke, T.V.; et al. Microglial inclusions and neurofilament light chain release follow neuronal α-synuclein lesions in long-term brain slice cultures. Mol. Neurodegener. 2021, 16, 1–17. [Google Scholar] [CrossRef]
- Refolo, V.; Bez, F.; Polissidis, A.; Kuzdas-Wood, D.; Sturm, E.; Kamaratou, M.; Poewe, W.; Stefanis, L.; Cenci, M.A.; Romero-Ramos, M.; et al. Progressive striatonigral degeneration in a transgenic mouse model of multiple system atrophy: Translational implications for interventional therapies. Acta Neuropathol. Commun. 2018, 6, 2. [Google Scholar] [CrossRef]
- Ullrich, C.; Daschil, N.; Humpel, C. Organotypic vibrosections: Novel whole sagittal brain cultures. J. Neurosci. Methods 2011, 201, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Ucar, B.; Humpel, C. Therapeutic efficacy of glial cell-derived neurotrophic factor loaded collagen scaffolds in ex vivo organotypic brain slice Parkinson’s disease models. Brain Res. Bull. 2019, 149, 86–95. [Google Scholar] [CrossRef]
- Foidl, B.M.; Ucar, B.; Schwarz, A.; Rebelo, A.L.; Pandit, A.; Humpel, C. Nerve growth factor released from collagen scaffolds protects axotomized cholinergic neurons of the basal nucleus of Meynert in organotypic brain slices. J. Neurosci. Methods 2018, 295, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Moelgg, K.; Jummun, F.; Humpel, C. Spreading of Beta-Amyloid in Organotypic Mouse Brain Slices and Microglial Elimination and Effects on Cholinergic Neurons. Biomolecules 2021, 11, 434. [Google Scholar] [CrossRef] [PubMed]
- Pineau, H.; Sim, V. POSCAbilities: The application of the prion organotypic slice culture assay to neurodegenerative disease research. Biomolecules 2020, 10, 1079. [Google Scholar] [CrossRef]
- Ucar, B.; Yusufogullari, S.; Humpel, C. Collagen hydrogels loaded with fibroblast growth factor—2 as a bridge to repair brain vessels in organotypic brain slices. Exp. Brain Res. 2020, 238, 2521–2529. [Google Scholar] [CrossRef]
- Ucar, B. Natural biomaterials in brain repair: A focus on collagen. Neurochem. Int. 2021, 146, 105033. [Google Scholar] [CrossRef]
- Hoffmann, A.C.; Minakaki, G.; Menges, S.; Salvi, R.; Savitskiy, S.; Kazman, A.; Miranda, H.V.; Mielenz, D.; Klucken, J.; Winkler, J.; et al. Extracellular aggregated alpha synuclein primarily triggers lysosomal dysfunction in neural cells prevented by trehalose. Sci. Rep. 2019, 9, 1–18. [Google Scholar]
- Masuda-Suzukake, M.; Nonaka, T.; Hosokawa, M.; Oikawa, T.; Arai, T.; Akiyama, H.; Mann, D.M.A.; Hasegawa, M. Prion-like spreading of pathological α-synuclein in brain. Brain 2013, 136, 1128–1138. [Google Scholar] [CrossRef]
- Howe, J.W.; Sortwell, C.E.; Duffy, M.F.; Kemp, C.J.; Russell, C.P.; Kubik, M.; Patel, P.; Luk, K.C.; El-Agnaf, O.M.A.; Patterson, J.R. Preformed fibrils generated from mouse alphα-synuclein produce more inclusion pathology in rats than fibrils generated from rat alphα-synuclein. Parkinsonism Relat. Disord. 2021, 89, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Rochet, J.C.; Conway, K.A.; Lansbury, P.T. Inhibition of fibrillization and accumulation of prefibrillar oligomers in mixtures of human and mouse α-synuclein. Biochemistry 2000, 39, 10619–10626. [Google Scholar] [CrossRef]
- Gracia, P.; Camino, J.D.; Volpicelli-Daley, L.; Cremades, N. Multiplicity of α-synuclein aggregated species and their possible roles in disease. Int. J. Mol. Sci. 2020, 21, 8043. [Google Scholar] [CrossRef]
- Fujiwara, H.; Hasegawa, M.; Dohmae, N.; Kawashima, A.; Masliah, E.; Goldberg, M.S.; Shen, J.; Takio, K.; Iwatsubo, T. α-Synuclein Is Phosphorylated in Synucleinopathy Lesions. Nat. Cell Biol. 2002, 4, 160–164. [Google Scholar] [CrossRef]
- Tenreiro, S.; Reimão-Pinto, M.M.; Antas, P.; Rino, J.; Wawrzycka, D.; Macedo, D.; Rosado-Ramos, R.; Amen, T.; Waiss, M.; Magalhães, F.; et al. Phosphorylation Modulates Clearance of Alpha-Synuclein Inclusions in a Yeast Model of Parkinson’s Disease. PLoS Genet. 2014, 10, e1004302. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, M.; Svenningsson, P. Update on alpha-synuclein-based biomarker approaches in the skin, submandibular gland, gastrointestinal tract, and biofluids. Curr. Opin. Neurol. 2021, 34, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Alam, P.; Bousset, L.; Melki, R.; Otzen, D.E. α-synuclein oligomers and fibrils: A spectrum of species, a spectrum of toxicities. J. Neurochem. 2019, 150, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Kahle, P.J.; Neumann, M.; Ozmen, L.; Müller, V.; Jacobsen, H.; Spooren, W.; Fuss, B.; Mallon, B.; Macklin, W.B.; Fujiwara, H.; et al. Hyperphosphorylation and insolubility of α-synuclein in transgenic mouse oligodendrocytes. EMBO Rep. 2002, 3, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Laferrière, F.; He, X.; Zinghirino, F.; Doudnikoff, E.; Faggiani, E.; Meissner, W.G.; Bezard, E.; De Giorgi, F.; Ichas, F. Overexpression of α-Synuclein by Oligodendrocytes in Transgenic Mice Does Not Recapitulate the Fibrillar Aggregation Seen in Multiple System Atrophy. Cells 2020, 9, 2371. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Li, X.; Liu, G.; Han, J.; Zhang, C.; Li, Y.; Xu, S.; Liu, C.; Gao, Y.; Yang, H.; et al. Extensive nuclear localization of α-synuclein in normal rat brain neurons revealed by a novel monoclonal antibody. Neuroscience 2007, 145, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, H.; Stefanovic, A.N.D.; Subramaniam, V.; Claessens, M.M.A.E. Membrane interactions and fibrillization of α-synuclein play an essential role in membrane disruption. FEBS Lett. 2014, 588, 4457–4463. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.; Ramachandran, M.; Moharil, P.; Harumalani, M.; Jaiswal, A.K. Biomaterials and cells for cardiac tissue engineering: Current choices. Mater. Sci. Eng. C 2017, 79, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Valdinocci, D.; Radford, R.A.W.; Siow, S.M.; Chung, R.S.; Pountney, D.L. Potential modes of intercellular α-synuclein transmission. Int. J. Mol. Sci. 2017, 18, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, D.M.; Selkoe, D.J. A critical appraisal of the pathogenic protein spread hypothesis of neurodegeneration. Nat. Rev. Neurosci. 2016, 17, 251–260. [Google Scholar] [CrossRef]
- Giráldez-Pérez, R.M.; Antolín-Vallespín, M.; Muñoz, M.D.; Sánchez-Capelo, A. Models of α-synuclein aggregation in Parkinson’s disease. Acta Neuropathol. Commun. 2014, 2, 176. [Google Scholar] [CrossRef] [Green Version]
- Recasens, A.; Ulusoy, A.; Kahle, P.J.; Di Monte, D.; Dehay, B. In vivo models of alphα-synuclein transmission and propagation. Cell Tissue Res. 2018, 373, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Tarutani, A.; Suzuki, G.; Shimozawa, A.; Nonaka, T.; Akiyama, H.; Hisanaga, S.-I.; Hasegawa, M. The effect of fragmented pathogenic α-synuclein seeds on prion-like propagation. J. Biol. Chem. 2016, 291, 18675–18688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humpel, C. Organotypic Brain Slices of ADULT Transgenic Mice: A Tool to Study Alzheimer’s Disease. Curr. Alzheimer Res. 2019, 16, 172–181. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uçar, B.; Stefanova, N.; Humpel, C. Spreading of Aggregated α-Synuclein in Sagittal Organotypic Mouse Brain Slices. Biomolecules 2022, 12, 163. https://doi.org/10.3390/biom12020163
Uçar B, Stefanova N, Humpel C. Spreading of Aggregated α-Synuclein in Sagittal Organotypic Mouse Brain Slices. Biomolecules. 2022; 12(2):163. https://doi.org/10.3390/biom12020163
Chicago/Turabian StyleUçar, Buket, Nadia Stefanova, and Christian Humpel. 2022. "Spreading of Aggregated α-Synuclein in Sagittal Organotypic Mouse Brain Slices" Biomolecules 12, no. 2: 163. https://doi.org/10.3390/biom12020163