

Polar Phenol Detection in Plasma and Serum: Insights on Sample Pre-Treatment for LC/MS Analysis and Application on the Serum of Corinthian Currant-Fed Rats
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Chemicals and Materials
2.2. Preparation of Stock and Working Solutions
2.3. Instrumentation Conditions
2.4. Sample Preparation
2.4.1. Deproteinization with ACN or MeOH/ACN 1:9
2.4.2. Deproteinization with ACN or MeOH/ACN 1:9, followed by SPE
2.4.3. Liquid–Liquid Extraction with EtOAc
2.4.4. Enzymatic Hydrolysis Followed by LLE with EtOAc
2.5. Method Validation
2.5.1. Specificity and Selectivity
2.5.2. Linearity, Carry-Over and Matrix Effect
2.5.3. Precision and Accuracy
2.5.4. Stability
2.5.5. Extraction Recovery
2.6. Polar Phenol Content of Corinthian Currant and Rat Chow
2.7. Animals and Diet
2.8. Statistical Analysis
3. Results and Discussion
3.1. Extraction Performance Comparison of Sample Preparation Methods
3.2. Extraction Performance of Enzymatic Hydrolysis, Followed by LLE
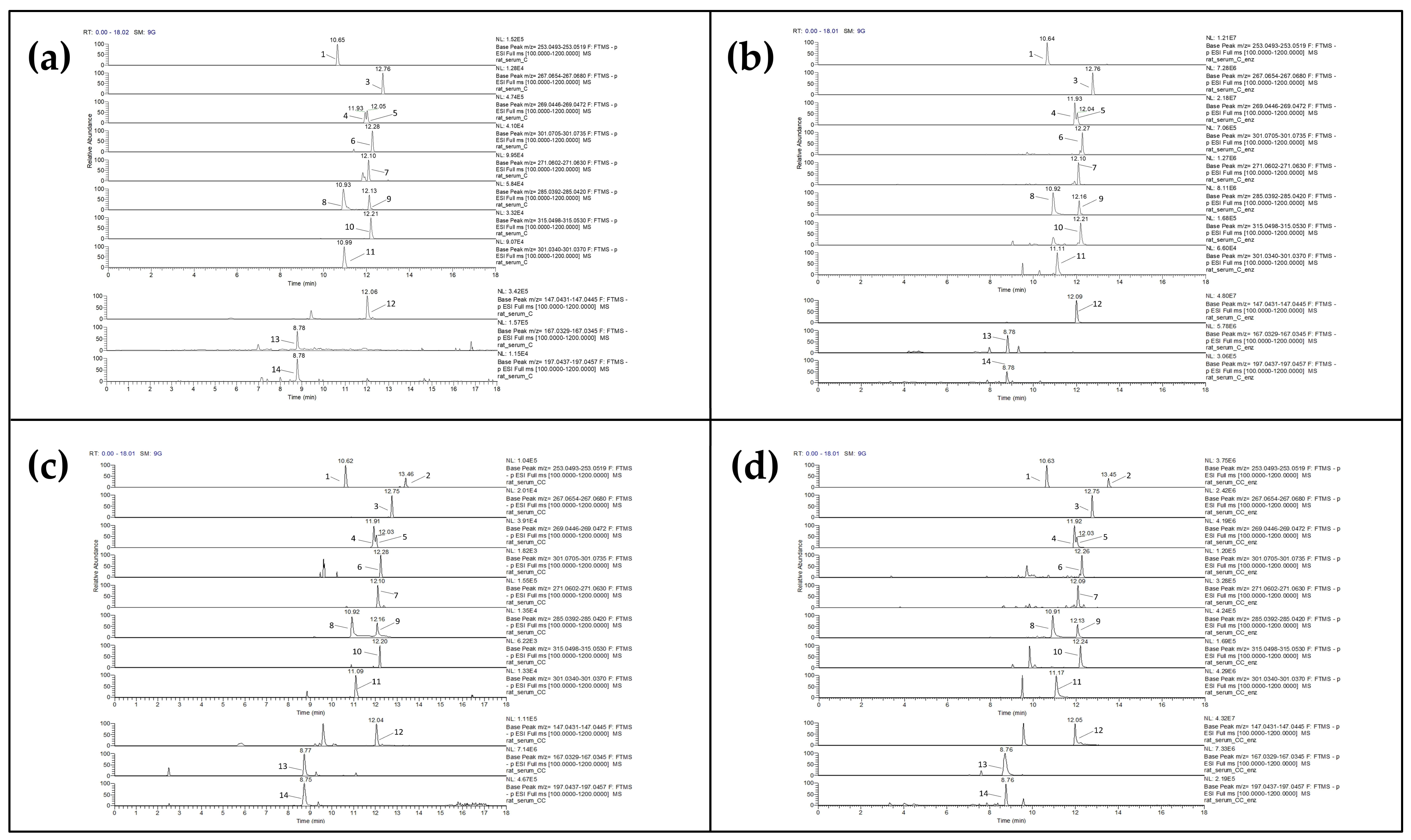
3.3. Polar Phenolics Detected in Serum of Corinthian Currant-Fed Rats
3.3.1. Comparisons of C vs. CC and Cenz vs. CCenz
3.3.2. Comparisons of C vs. Cenz and CC vs. CCenz
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiou, A.; Karathanos, V.; Mylona, A.; Salta, F.; Preventi, F.; Andrikopoulos, N. Currants (Vitis vinifera L.) Content of Simple Phenolics and Antioxidant Activity. Food Chem. 2007, 102, 516–522. [Google Scholar] [CrossRef]
- Chiou, A.; Panagopoulou, E.A.; Gatzali, F.; De Marchi, S.; Karathanos, V.T. Anthocyanins Content and Antioxidant Capacity of Corinthian Currants (Vitis vinifera L., Var. Apyrena). Food Chem. 2014, 146, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Kanellos, P.T.; Kaliora, A.C.; Gioxari, A.; Christopoulou, G.O.; Kalogeropoulos, N.; Karathanos, V.T. Absorption and Bioavailability of Antioxidant Phytochemicals and Increase of Serum Oxidation Resistance in Healthy Subjects Following Supplementation with Raisins. Plant Foods Hum. Nutr. 2013, 68, 411–415. [Google Scholar] [CrossRef]
- Vasilakopoulou, P.B.; Fanarioti, Ε.; Tsarouchi, M.; Kokotou, M.G.; Dermon, C.R.; Karathanos, V.T.; Chiou, A. Polar Phenol Detection in Rat Brain: Development and Validation of a Versatile UHPLC-MS Method and Application on the Brain Tissues of Corinthian Currant (Vitis vinifera L., Var. Apyrena) Fed Rats. Food Chem. 2022, 390, 133131. [Google Scholar] [CrossRef]
- Santos, A.C.; Costa, G.; Veiga, F.; Figueiredo, I.V.; Batista, M.T.; Ribeiro, A.J. Advance in Methods Studying the Pharmacokinetics of Polyphenols. Curr. Drug Metab. 2014, 15, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Serreli, G.; Deiana, M. In Vivo Formed Metabolites of Polyphenols and Their Biological Efficacy. Food Funct. 2019, 10, 6999–7021. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Naeem, A.; Ming, Y.; Pengyi, H.; Jie, K.Y.; Yali, L.; Haiyan, Z.; Shuai, X.; Wenjing, L.; Ling, W.; Xia, Z.M.; et al. The Fate of Flavonoids after Oral Administration: A Comprehensive Overview of Its Bioavailability. Crit. Rev. Food Sci. Nutr. 2021, 62, 1–18. [Google Scholar] [CrossRef]
- Passon, M. Dietary Phenolic Compounds in Biological Samples: Current Challenges in Analytical Chemistry. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1929–1958. ISBN 978-3-319-78030-6. [Google Scholar]
- Paglia, G.; Del Greco, F.M.; Sigurdsson, B.B.; Rainer, J.; Volani, C.; Hicks, A.A.; Pramstaller, P.P.; Smarason, S.V. Influence of Collection Tubes during Quantitative Targeted Metabolomics Studies in Human Blood Samples. Clin. Chim. Acta 2018, 486, 320–328. [Google Scholar] [CrossRef]
- Plebani, M.; Banfi, G.; Bernardini, S.; Bondanini, F.; Conti, L.; Dorizzi, R.; Ferrara, F.E.; Mancini, R.; Trenti, T. Serum or Plasma? An Old Question Looking for New Answers. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 178–187. [Google Scholar] [CrossRef]
- López-Yerena, A.; Domínguez-López, I.; Vallverdú-Queralt, A.; Pérez, M.; Jáuregui, O.; Escribano-Ferrer, E.; Lamuela-Raventós, R.M. Metabolomics Technologies for the Identification and Quantification of Dietary Phenolic Compound Metabolites: An Overview. Antioxidants 2021, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Moein, M.M.; El Beqqali, A.; Abdel-Rehim, M. Bioanalytical Method Development and Validation: Critical Concepts and Strategies. J. Chromatogr. B 2017, 1043, 3–11. [Google Scholar] [CrossRef]
- Wu, W.; Li, K.; Zhao, C.; Ran, X.; Zhang, Y.; Zhang, T. A Rapid HPLC–MS/MS Method for the Simultaneous Determination of Luteolin, Resveratrol and Their Metabolites in Rat Plasma and Its Application to Pharmacokinetic Interaction Studies. J. Chromatogr. B 2022, 1191, 123118. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Zhang, Y.-B.; Yang, X.-W.; Xu, W.; Liu, L.; Zhang, P.; Gong, Y.; Liu, N.-F.; Peng, K.-F. Simultaneous Determination of Twenty-Five Compounds with Anti-Inflammatory Activity in Spatholobi Caulis by Using an Optimized UFLC-MS/MS Method: An Application to Pharmacokinetic Study. J. Pharm. Biomed. Anal. 2021, 204, 114267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cui, W.; Liu, X.; Wang, N.; Kong, W.; Sui, J.; Li, H.; Wang, S. Simultaneous Determination of Seven Bioactive Constituents from Salvia Miltiorrhiza in Rat Plasma by HPLC-MS/MS: Application to a Comparative Pharmacokinetic Study. Separations 2021, 8, 93. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, Z.; Cai, W.; Tian, X.; Liu, S. A Validated LC–MS/MS Method for the Simultaneous Determination of Ticagrelor, Its Two Metabolites and Major Constituents of Tea Polyphenols in Rat Plasma and Its Application in a Pharmacokinetic Study. J. Chromatogr. Sci. 2021, 59, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Eure, W.D.; Grossman, R.G.; Horner, P.J.; Chow, D.S.-L. LC-MS/MS Assay of Riluzole and Etoricoxib in Rat Plasma and Brain Tissue with Applications for Sampling and Evaluation in Pre-Clinical Rat Model of Traumatic Brain Injury. Talanta Open 2021, 4, 100052. [Google Scholar] [CrossRef]
- Chavari, M.; de Góes, P.R.N.; Lachi-Silva, L.; Barth, A.B.; da Silva, A.O.F.; Longhini, R.; de Mello, J.C.P.; Kimura, E.; Diniz, A. Simultaneous Liquid Chromatography- Tandem Mass Spectrometry Method to Quantify Epicatechin and Procyanidin B2 in Rat Plasma after Oral Administration of Trichilia Catigua (Catuaba) Extract and Its Application to a Pharmacokinetic Study. Rev. Bras. Farmacogn. 2019, 29, 457–463. [Google Scholar] [CrossRef]
- Tao, Y.; Xu, X.; Yan, J.; Cai, B. A Sensitive UPLC–MS/MS Method for Simultaneous Determination of Polyphenols in Rat Plasma: Application to a Pharmacokinetic Study of Dispensing Granules and Standard Decoction of Cinnamomum Cassia Twigs. Biomed. Chromatogr. 2019, 33, e4534. [Google Scholar] [CrossRef] [PubMed]
- Castello, F.; Costabile, G.; Bresciani, L.; Tassotti, M.; Naviglio, D.; Luongo, D.; Ciciola, P.; Vitale, M.; Vetrani, C.; Galaverna, G.; et al. Bioavailability and Pharmacokinetic Profile of Grape Pomace Phenolic Compounds in Humans. Arch. Biochem. Biophys. 2018, 646, 1–9. [Google Scholar] [CrossRef]
- Xu, Z.-L.; Xu, M.-Y.; Wang, H.-T.; Xu, Q.-X.; Liu, M.-Y.; Jia, C.-P.; Geng, F.; Zhang, N. Pharmacokinetics of Eight Flavonoids in Rats Assayed by UPLC-MS/MS after Oral Administration of Drynariae Rhizoma Extract. J. Anal. Methods Chem. 2018, 2018, 4789196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, M.; Romero, M.-P.; Macià, A.; Valls, R.M.; Fernández, S.; Solà, R.; Motilva, M.-J. Improved Method for Identifying and Quantifying Olive Oil Phenolic Compounds and Their Metabolites in Human Plasma by Microelution Solid-Phase Extraction Plate and Liquid Chromatography–Tandem Mass Spectrometry. J. Chromatogr. B 2009, 877, 4097–4106. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Chen, C.Y.O.; Preuss, H.G.; Ray, S.D.; Bucci, L.R.; Ji, J.; Ruff, K.J. The Fallacy of Enzymatic Hydrolysis for the Determination of Bioactive Curcumin in Plasma Samples as an Indication of Bioavailability: A Comparative Study. BMC Complement. Altern. Med. 2019, 19, 293. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Lanzi, C.; Perdicaro, D.J.; Antoniolli, A.; Piccoli, P.; Vazquez Prieto, M.A.; Fontana, A. Phenolic Metabolites in Plasma and Tissues of Rats Fed with a Grape Pomace Extract as Assessed by Liquid Chromatography-Tandem Mass Spectrometry. Arch. Biochem. Biophys. 2018, 651, 28–33. [Google Scholar] [CrossRef]
- Matsumoto, T.; Matsubara, Y.; Mizuhara, Y.; Sekiguchi, K.; Koseki, J.; Tsuchiya, K.; Nishimura, H.; Watanabe, J.; Kaneko, A.; Maemura, K.; et al. Plasma Pharmacokinetics of Polyphenols in a Traditional Japanese Medicine, Jumihaidokuto, Which Suppresses Propionibacterium Acnes-Induced Dermatitis in Rats. Molecules 2015, 20, 18031–18046. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, D.M.; Pinto, C.B.; Sampaio, G.R.; Yonekura, L.; Catharino, R.R.; Bastos, D.H.M. Development and Validation of Methods for the Extraction of Phenolic Acids from Plasma, Urine, and Liver and Analysis by UPLC-MS. J. Agric. Food Chem. 2013, 61, 6113–6121. [Google Scholar] [CrossRef]
- Huo, Y.; Zhang, Q.; Li, Q.; Geng, B.; Bi, K. Development of a UFLC-MS/MS Method for the Simultaneous Determination of Seven Tea Catechins in Rat Plasma and Its Application to a Pharmacokinetic Study after Administration of Green Tea Extract. J. Pharm. Biomed. Anal. 2016, 125, 229–235. [Google Scholar] [CrossRef]
- Quifer-Rada, P.; Martínez-Huélamo, M.; Lamuela-Raventos, R.M. Is Enzymatic Hydrolysis a Reliable Analytical Strategy to Quantify Glucuronidated and Sulfated Polyphenol Metabolites in Human Fluids? Food Funct. 2017, 8, 2419–2424. [Google Scholar] [CrossRef]
- Taylor, J.I.; Grace, P.B.; Bingham, S.A. Optimization of Conditions for the Enzymatic Hydrolysis of Phytoestrogen Conjugates in Urine and Plasma. Anal. Biochem. 2005, 341, 220–229. [Google Scholar] [CrossRef]
- Yanni, A.E.; Efthymiou, V.; Lelovas, P.; Agrogiannis, G.; Kostomitsopoulos, N.; Karathanos, V.T. Effects of Dietary Corinthian Currants (Vitis vinifera L., Var. Apyrena) on Atherosclerosis and Plasma Phenolic Compounds during Prolonged Hypercholesterolemia in New Zealand White Rabbits. Food Funct. 2015, 6, 963–971. [Google Scholar] [CrossRef]
- Ding, Y.; Peng, M.; Zhang, T.; Tao, J.-S.; Cai, Z.-Z.; Zhang, Y. Quantification of Conjugated Metabolites of Drugs in Biological Matrices after the Hydrolysis with β-Glucuronidase and Sufatase: A Review of Bio-Analytical Methods. Biomed. Chromatogr. 2013, 27, 1280–1295. [Google Scholar] [CrossRef]
- Fernández-Ochoa, Á.; de la Luz Cádiz-Gurrea, M.; Fernández-Moreno, P.; Rojas-García, A.; Arráez-Román, D.; Segura-Carretero, A. Recent Analytical Approaches for the Study of Bioavailability and Metabolism of Bioactive Phenolic Compounds. Molecules 2022, 27, 777. [Google Scholar] [CrossRef]
- EMEA/CHMP/EWP/192217/2009 Guideline on Bioanalytical Method Validation; Committee for Medicinal Products for Human Use: Amsterdam, The Netherlands, 2011.
- Bylda, C.; Thiele, R.; Kobold, U.; Volmer, D.A. Recent Advances in Sample Preparation Techniques to Overcome Difficulties Encountered during Quantitative Analysis of Small Molecules from Biofluids Using LC-MS/MS. Analyst 2014, 139, 2265–2276. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, H.; Huan, X.; Cao, N.; Guan, H.; Zhang, H.; Cheng, X.; Wang, C. Simultaneous LC-MS/MS Bioanalysis of Alkaloids, Terpenoids, and Flavonoids in Rat Plasma through Salting-out-Assisted Liquid-Liquid Extraction after Oral Administration of Extract from Tetradium Ruticarpum and Glycyrrhiza Uralensis: A Sample Preparation Strategy to Broaden Analyte Coverage of Herbal Medicines. Anal. Bioanal. Chem. 2021, 413, 5871–5884. [Google Scholar] [CrossRef]
- Cortese, M.; Gigliobianco, M.R.; Magnoni, F.; Censi, R.; Di Martino, P. Compensate for or Minimize Matrix Effects? Strategies for Overcoming Matrix Effects in Liquid Chromatography-Mass Spectrometry Technique: A Tutorial Review. Molecules 2020, 25, 3047. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef] [PubMed]
- Luis, P.B.; Kunihiro, A.G.; Funk, J.L.; Schneider, C. Incomplete Hydrolysis of Curcumin Conjugates by β-Glucuronidase: Detection of Complex Conjugates in Plasma. Mol. Nutr. Food Res. 2020, 64, 1901037. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, L.; Pastene, E.; Duran-Sandoval, D.; Vergara, C.; Von Baer, D.; Mardones, C. Pharmacokinetics of Low Molecular Weight Phenolic Compounds in Gerbil Plasma after the Consumption of Calafate Berry (Berberis Microphylla) Extract. Food Chem. 2018, 268, 347–354. [Google Scholar] [CrossRef]


{kind=link}
| Analyte | PPT_1 | PPT_2 | PPT_3 | PPT_4 | PPT + SPE_1 | PPT + SPE_2 | PPT + SPE_3 | PPT + SPE_4 | LLE_1 | LLE_2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Isoflavones | ||||||||||
| Daidzein | 55 ± 3 ab | 48 ± 0 ab | 38 ± 2 a | 41 ± 2 a | 47 ± 3 ab | 116 ± 6 d | 115 ± 9 d | 61 ± 18 b | 90 ± 8 c | 56 ± 4 ab |
| Formononetin | 54 ± 2 bc | 45 ± 0 ab | 32 ± 1 a | 33 ± 1 a | 42 ± 4 ab | 118 ± 9 e | 110 ± 6 e | 69 ± 20 c | 84 ± 7 d | 50 ± 2 b |
| Genistein | 52 ± 2 b | 47 ± 1 ab | 34 ± 1 a | 39 ± 2 ab | 36 ± 3 ab | 82 ± 3 c | 86 ± 5 c | 74 ± 22 c | 84 ± 11 c | 52 ± 3 b |
| Flavanones | ||||||||||
| Hesperetin | 45 ± 2 b | 44 ± 0 ab | 27 ± 1 a | 35 ± 2 ab | 38 ± 3 ab | 109 ± 7 d | 80 ± 6 c | 75 ± 22 c | 87 ± 6 cd | 51 ± 3 b |
| Naringenin | 47 ± 2 bc | 47 ± 1 bc | 26 ± 1 a | 37 ± 1 abc | 33 ± 3 ab | 106 ± 7 ef | 78 ± 6 de | 107 ± 8 f | 83 ± 9 d | 53 ± 3 c |
| Flavones | ||||||||||
| Apigenin | 33 ± 2 a | 40 ± 0 ab | 26 ± 1 a | 28 ± 1 a | 98 ± 7 d | 86 ± 12 cd | 74 ± 8 c | 87 ± 9 c | 80 ± 12 c | 50 ± 3 b |
| Chrysin | 47 ± 1 b | 45 ± 1 b | 20 ± 1 a | 21 ± 0 a | 98 ± 10 d | 84 ± 11 c | 77 ± 7 c | 77 ± 6 c | 78 ± 8 c | 51 ± 3 b |
| Luteolin | 28 ± 2 a | 34 ± 1 ab | 26 ± 1 a | 27 ± 0 a | 66 ± 4 c | 76 ± 11 cd | 75 ± 5 cd | 95 ± 7 d | 76 ± 13 cd | 46 ± 8 b |
| Flavonols | ||||||||||
| Isorhamnetin | n.d. | 27 ± 15 b | 25 ± 0 b | 25 ± 1 b | 42 ± 3 c | 56 ± 11 d | 77 ± 5 e | 84 ± 6 e | 80 ± 8 e | 59 ± 3 d |
| Kaempferol | 5 ± 1 a | n.d. | 22 ± 0 b | 24 ± 1 b | 92 ± 7 f | 58 ± 13 cd | 67 ± 6 de | 80 ± 7 e | 77 ± 10 e | 57 ± 3 c |
| Quercetin | n.d. | 7 ± 7 a | 22 ± 0 b | 21 ± 0 b | 6 ± 6 a | 55 ± 14 c | 69 ± 5 de | 75 ± 4 de | 80 ± 10 e | 63 ± 10 cd |
| Flavan-3-ols | ||||||||||
| Catechin | n.d. | n.d. | n.d. | n.d. | 32 ± 2 b | 55 ± 10 c | 62 ± 6 cd | 59 ± 2 cd | 66 ± 9 a | 4 ± 2 d |
| Epicatechin | n.d. | n.d. | n.d. | n.d. | 67 ± 4 cd | 56 ± 11 b | 62 ± 5 bc | 60 ± 1 bc | 73 ± 7 d | 1 ± 1 a |
| Epicatechin gallate | n.d. | n.d. | n.d. | n.d. | n.d. | 61 ± 15 c | 72 ± 5 c | 59 ± 3 c | 91 ± 16 d | 24 ± 1 b |
| Epigallocatechin | n.d. | n.d. | n.d. | n.d. | n.d. | 50 ± 11 b | 57 ± 5 bc | 61 ± 2 c | 71 ± 15 c | 2 ± 1 a |
| Epigallocatechin gallate | n.d. | n.d. | n.d. | n.d. | n.d. | 55 ± 16 c | 65 ± 5 c | 55 ± 3 c | 92 ± 9 d | 18 ± 4 b |
| Procyanidin B2 | n.d. | n.d. | n.d. | n.d. | n.d. | 52 ± 14 c | 59 ± 4 c | 44 ± 2 b | n.d. | n.d. |
| Flavonol glycosides | ||||||||||
| Isoquercetin | 30 ± 4 ab | 28 ± 2 ab | 31 ± 2 ab | 34 ± 2 b | 18 ± 2 a | 86 ± 6 d | 68 ± 11 d | 72 ± 6 d | 105 ± 12 e | 51 ± 2 c |
| Rutin | 32 ± 2 ab | 29 ± 3 ab | 29 ± 1 ab | 36 ± 2 b | 21 ± 3 a | 93 ± 5 d | 73 ± 4 c | 65 ± 8 c | 99 ± 8 d | 66 ± 3 c |
| Stilbenes | ||||||||||
| trans-Resveratrol | 37 ± 3 c | n.d. | 4 ± 1 a | 15 ± 2 b | 28 ± 2 c | 60 ± 6 de | 48 ± 5 d | 62 ± 4 ef | 69 ± 9 f | 61 ± 2 ef |
| E-Viniferin | 37 ± 2 c | 23 ± 1 bc | 12 ± 1 ab | 23 ± 1 bc | n.d.a | 67 ± 20 d | 63 ± 13 d | 69 ± 10 d | 66 ± 10 d | 98 ± 6 e |
| Piceid | 51 ± 2 de | 43 ± 1 cd | 22 ± 4 ab | 32 ± 2 bc | 12 ± 2 a | 79 ± 5 g | 62 ± 5 fg | 95 ± 11 h | 99 ± 8 h | 59 ± 4 ef |
| Phenylethanoids | ||||||||||
| 3-Hydroxytyrosol | 14 ± 2 a | 6 ± 0 a | 36 ± 2 bc | 39 ± 2 bc | 34 ± 7 b | 99 ± 8 g | 68 ± 3 ef | 59 ± 16 de | 76 ± 6 f | 49 ± 2 cd |
| Oleuropein | n.d. | n.d. | n.d. | 39 ± 1 bc | 33 ± 11 b | 56 ± 11 de | 74 ± 4 f | 67 ± 2 ef | 95 ± 12 g | 50 ± 4 cd |
| Cinnamic acid and derivatives | ||||||||||
| Caffeic acid | n.d. | n.d. | 30 ± 3 b | 39 ± 1 bc | 51 ± 9 c | 53 ± 9 c | 65 ± 10 d | 109 ± 2 e | 87 ± 9 d | 50 ± 2 c |
| trans-Cinnamic acid | 64 ± 5 def | 45 ± 3 bc | 20 ± 2 a | 29 ± 2 ab | 45 ± 16 bc | 105 ± 11 g | 75 ± 5 ef | 60 ± 15 cde | 82 ± 6 fg | 46 ± 2 bcd |
| Chlorogenic acid | 24 ± 1 ab | 3 ± 1 a | 33 ± 6 b | 55 ± 4 cd | 68 ± 7 d | 100 ± 8 f | 65 ± 5 de | 103 ± 3 ef | 41 ± 3 bc | 26 ± 2 b |
| o-Coumaric acid | 49 ± 1 ab | 42 ± 1 a | 36 ± 2 a | 39 ± 1 a | 42 ± 8 a | 80 ± 4 c | 108 ± 8 d | 64 ± 17 b | 93 ± 7 cd | 47 ± 2 a |
| p-Coumaric acid | 48 ± 2 bc | 43 ± 1 abc | 33 ± 3 a | 39 ± 2 ab | 51 ± 4 c | 77 ± 4 d | 69 ± 11 de | 84 ± 6 ef | 88 ± 5 f | 51 ± 2 c |
| Ferulic acid | 48 ± 2 c | 42 ± 0 bc | 28 ± 4 ab | 39 ± 1 abc | 23 ± 3 a | 80 ± 5 d | 64 ± 9 d | 117 ± 2 e | 88 ± 5 d | 51 ± 3 c |
| Neochlorogenic acid | 20 ± 1 ab | 3 ± 1 a | 27 ± 6 b | 52 ± 3 c | 17 ± 5 ab | 79 ± 9 d | 72 ± 7 d | 107 ± 11 d | 25 ± 7 ab | 33 ± 3 bc |
| Sinapic acid | 47 ± 1 bc | 42 ± 1 bc | 29 ± 4 ab | 37 ± 2 ab | 20 ± 2 a | 87 ± 6 e | 62 ± 8 de | 120 ± 11 f | 85 ± 5 e | 56 ± 3 cd |
| Benzoic acid derivatives | ||||||||||
| Gallic acid | n.d. | n.d. | 32 ± 2 b | 33 ± 1 b | 6 ± 1 a | 40 ± 7 bc | 62 ± 12 d | 58 ± 17 c | 38 ± 7 bc | 78 ± 7 d |
| Syringic acid | n.d. | n.d. | 33 ± 3 bc | 23 ± 1 ab | n.d. | 22 ± 7 b | 58 ± 12 cd | 11 ± 8 ab | 76 ± 9 d | 24 ± 16 b |
| Vanillic acid | 56 ± 3 bc | 49 ± 1 abc | 39 ± 2 a | 37 ± 1 a | 44 ± 15 ab | 81 ± 5 d | 77 ± 6 d | 59 ± 11 c | 86 ± 11 d | 59 ± 3 c |
| Analyte | PPT_1 | PPT_2 | PPT_3 | PPT_4 | PPT + SPE_1 | PPT + SPE_2 | PPT + SPE_3 | PPT + SPE_4 | LLE_1 | LLE_2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Isoflavones | ||||||||||
| Daidzein | 42 ± 10 ab | 51 ± 17 abcd | 24 ± 4 a | 49 ± 4 abc | 80 ± 12 def | 70 ± 17 bcd | 72 ± 7 cde | 108 ± 11 f | 100 ± 9 ef | 66 ± 1 bcd |
| Formononetin | 39 ± 7 ab | 47 ± 14 abc | 31 ± 2 a | 44 ± 3 abc | 55 ± 14 bc | 107 ± 5 e | 66 ± 1 cd | 93 ± 6 e | 87 ± 6 de | 57 ± 2 bc |
| Genistein | 17 ± 24 ab | 1 ± 1 a | 10 ± 3 a | 37 ± 3 abc | 80 ± 31 d | 92 ± 4 d | 58 ± 20 bcd | 81 ± 3 d | 75 ± 8 cd | 60 ± 4 cd |
| Flavanones | ||||||||||
| Hesperetin | 10 ± 17 a | n.d. | 3 ± 2 a | 18 ± 3 ab | 62 ± 12 cd | 78 ± 8 d | 46 ± 17 bc | 70 ± 12 cd | 68 ± 6 cd | 57 ± 3 cd |
| Naringenin | 10 ± 16 a | n.d. | 4 ± 2 a | 20 ± 3 a | 61 ± 11 bc | 78 ± 9 c | 49 ± 14 b | 65 ± 3 bc | 73 ± 8 bc | 59 ± 2 bc |
| Flavones | ||||||||||
| Apigenin | 2 ± 1 a | 1 ± 1 a | 13 ± 2 ab | 32 ± 2 bc | 55 ± 14 de | 68 ± 6 def | 48 ± 12 cd | 69 ± 9 ef | 82 ± 5 f | 53 ± 1 cde |
| Chrysin | 10 ± 13 a | 2 ± 3 a | 20 ± 1 ab | 31 ± 1 abc | 65 ± 21 de | 71 ± 10 de | 51 ± 15 cd | 62 ± 11 de | 84 ± 3 e | 46 ± 4 bcd |
| Luteolin | 8 ± 13 a | 1 ± 1 a | 8 ± 2 a | 30 ± 1 ab | 74 ± 24 c | 63 ± 7 bc | 53 ± 21 bc | 64 ± 7 bc | 71 ± 14 c | 48 ± 6 bc |
| Flavonols | ||||||||||
| Isorhamnetin | 5 ± 9 ab | n.d. | 18 ± 3 abc | 32 ± 1 bcd | 74 ± 27 e | 57 ± 5 de | 46 ± 12 cde | 51 ± 8 de | 65 ± 4 e | 7 ± 5 ab |
| Kaempferol | 12 ± 21 a | n.d. | 18 ± 2 ab | 34 ± 1 abc | 74 ± 24 d | 57 ± 5 cd | 52 ± 14 bcd | 54 ± 8 cd | 67 ± 6 cd | 8 ± 6 a |
| Quercetin | 4 ± 6 a | n.d. | 10 ± 2 ab | 29 ± 1 abc | 79 ± 28 e | 50 ± 5 cde | 44 ± 16 cd | 37 ± 7 bcd | 68 ± 4 de | 7 ± 5 ab |
| Flavan-3-ols | ||||||||||
| Catechin | n.d. | n.d. | n.d. | n.d. | 64 ± 7 bc | 81 ± 3 c | 47 ± 20 b | 53 ± 4 b | 81 ± 9 c | 6 ± 4 a |
| Epicatechin | n.d. | n.d. | n.d. | n.d. | 73 ± 27 cd | 78 ± 5 d | 37 ± 4 b | 51 ± 4 bc | 69 ± 6 cd | 7 ± 2 a |
| Epicatechin gallate | n.d. | n.d. | n.d. | n.d. | 47 ± 12 b | 53 ± 8 b | 35 ± 14 b | 40 ± 1 b | 98 ± 19 d | 3 ± 3 a |
| Epigallocatechin | n.d. | n.d. | n.d. | n.d. | 39 ± 14 b | 60 ± 2 bc | 42 ± 18 b | 38 ± 9 b | 73 ± 12 c | 5 ± 4 a |
| Epigallocatechin gallate | n.d. | n.d. | n.d. | n.d. | 37 ± 14 bc | 40 ± 7 bc | 33 ± 13 bc | 29 ± 4 b | 55 ± 15 c | n.d. |
| Procyanidin B2 | n.d. | n.d. | n.d. | n.d. | 51 ± 30 bc | 65 ± 11 c | 27 ± 16 ab | 37 ± 8 bc | n.d. | n.d. |
| Flavonol glycosides | ||||||||||
| Isoquercetin | 11 ± 18 ab | n.d. | n.d. | 30 ± 13 abc | 70 ± 16 cd | 80 ± 14 d | 59 ± 24 cd | 83 ± 9 d | 86 ± 18 d | 49 ± 4 bcd |
| Rutin | n.d. | n.d. | n.d. | 29 ± 13 abc | 79 ± 34 d | 77 ± 11 d | 67 ± 27 bcd | 89 ± 8 d | 71 ± 7 cd | 25 ± 4 ab |
| Stilbenes | ||||||||||
| trans-Resveratrol | 14 ± 24 ab | n.d. | n.d. | n.d. | 76 ± 39 c | 80 ± 8 c | 40 ± 15 abc | 47 ± 0 bc | 58 ± 5 bc | 46 ± 4 bc |
| E-Viniferin | 19 ± 17 ab | 4 ± 4 a | n.d. | 4 ± 4 a | 11 ± 14 ab | 24 ± 2 ab | 25 ± 13 ab | 35 ± 4 b | 96 ± 7 c | 6 ± 4 a |
| Piceid | 31 ± 17 abc | 28 ± 8 ab | n.d. | n.d. | 80 ± 17 de | 67 ± 18 cde | 45 ± 17 bcd | 53 ± 4 bcde | 87 ± 15 e | 26 ± 9 ab |
| Phenylethanoids | ||||||||||
| 3-Hydroxytyrosol | 27 ± 9 a | 30 ± 8 ab | 36 ± 2 ab | 47 ± 4 bc | 76 ± 5 d | 80 ± 15 d | 74 ± 1 d | 115 ± 7 e | 63 ± 4 cd | 62 ± 4 cd |
| Oleuropein | 45 ± 78 ab | n.d. | n.d. | n.d. | 62 ± 2 ab | 84 ± 9 b | 46 ± 19 ab | 52 ± 1 ab | 91 ± 9 b | 7 ± 6 a |
| Cinnamic acid and derivatives | ||||||||||
| Caffeic acid | 16 ± 25 abc | n.d. | 10 ± 1 ab | 22 ± 5 abc | 81 ± 31 d | 92 ± 22 d | 52 ± 20 bcd | 61 ± 2 cd | 82 ± 6 d | 24 ± 8 abc |
| trans-Cinnamic acid | 31 ± 10 a | 47 ± 23 ab | 28 ± 2 a | 39 ± 4 a | 72 ± 7 ab | 74 ± 24 ab | 72 ± 39 ab | 98 ± 16 b | 61 ± 5 ab | 59 ± 2 ab |
| Chlorogenic acid | 44 ± 17 bcd | 33 ± 16 abc | n.d. | 10 ± 4 ab | 71 ± 9 d | 73 ± 14 d | 67 ± 29 cd | 56 ± 6 cd | 38 ± 6 abcd | 36 ± 5 abcd |
| o-Coumaric acid | 36 ± 11 ab | 47 ± 17 abc | 17 ± 1 a | 45 ± 6 abc | 73 ± 10 cde | 66 ± 18 bcd | 65 ± 26 bcd | 93 ± 10 cd | 106 ± 4 d | 68 ± 2 bcd |
| p-Coumaric acid | 35 ± 10 abc | 44 ± 15 bcd | 13 ± 1 a | 23 ± 7 ab | 69 ± 10 de | 65 ± 16 de | 51 ± 18 bcd | 63 ± 5 cde | 88 ± 6 e | 67 ± 0 de |
| Ferulic acid | 37 ± 13 abc | 44 ± 17 abcd | 11 ± 1 a | 21 ± 6 ab | 76 ± 11 de | 64 ± 16 cde | 52 ± 20 bcde | 60 ± 4 cde | 83 ± 3 e | 59 ± 4 cde |
| Neochlorogenic acid | 28 ± 13 abc | 34 ± 16 abc | n.d. | 11 ± 5 ab | 83 ± 32 de | 95 ± 9 e | 46 ± 19 bcd | 51 ± 3 cd | n.d. | 9 ± 4 ab |
| Sinapic acid | 28 ± 24 | 28 ± 10 | 7 ± 1 | 18 ± 4 | 81 ± 32 | 93 ± 16 | 48 ± 17 | 57 ± 3 | 90 ± 8 | 37 ± 5 |
| Benzoic acid derivatives | ||||||||||
| Gallic acid | 19 ± 0 a | n.d. | n.d. | 41 ± 10 abc | 82 ± 32 cd | 65 ± 2 bcd | 67 ± 0 bcd | 98 ± 11 d | 34 ± 2 ab | 11 ± 6 a |
| Syringic acid | 10 ± 9 a | 7 ± 12 a | 32 ± 1 ab | 46 ± 3 abc | 38 ± 0 ab | 61 ± 22 bc | 70 ± 5 bc | 80 ± 9 c | 68 ± 11 bc | 17 ± 3 a |
| Vanillic acid | 41 ± 8 ab | 54 ± 22 ab | 33 ± 1 a | 55 ± 7 ab | 63 ± 2 ab | 59 ± 21 ab | 74 ± 15 b | 68 ± 15 ab | 72 ± 7 b | 76 ± 7 b |
| Analyte | Plasma | Serum | ||
|---|---|---|---|---|
| (−) | (+) | (−) | (+) | |
| Isoflavones | ||||
| Daidzein | 80 ± 7 a | 69 ± 6 b | 73 ± 7 a | 73 ± 4 a |
| Formononetin | 57 ± 7 a | 27 ± 2 b | 66 ± 6 a | 24 ± 2 b |
| Genistein | 53 ± 2 a | 51 ± 6 a | 55 ± 9 a | 56 ± 4 a |
| Flavanones | ||||
| Hesperetin | 70 ± 4 a | 59 ± 6 b | 78 ± 8 a | 68 ± 4 b |
| Naringenin | 46 ± 6 a | 22 ± 3 b | 46 ± 2 a | 35 ± 2 b |
| Flavones | ||||
| Apigenin | 76 ± 2 a | 47 ± 4 b | 68 ± 2 a | 56 ± 1 b |
| Chrysin | 85 ± 4 a | 56 ± 2 b | 63 ± 1 a | 35 ± 1 b |
| Luteolin | 34 ± 4 a | 49 ± 8 b | 41 ± 9 a | 58 ± 6 b |
| Flavonols | ||||
| Isorhamnetin | 62 ± 9 a | 13 ± 2 b | 66 ± 5 a | 33 ± 1 b |
| Kaempferol | 51 ± 4 a | 23 ± 3 b | 48 ± 6 a | 20 ± 2 b |
| Quercetin | 42 ± 3 a | 2.2 ± 1 b | 49 ± 1 a | 6 ± 1 b |
| Flavan-3-ols | ||||
| Catechin | 67 ± 1 a | 50 ± 5 b | 59 ± 3 a | 55 ± 2 a |
| Epicatechin | 63 ± 4 a | 57 ± 5 a | 59 ± 8 a | 60 ± 2 a |
| Epicatechin gallate | 106 ± 5 a | 42 ± 8 b | 81 ± 6 a | 31 ± 0.3 b |
| Epigallocatechin | 27 ± 2 a | 11 ± 5 b | 15 ± 2 a | 22 ± 3 a |
| Epigallocatechin gallate | 62 ± 5 a | 26 ± 1 b | 53 ± 3 a | 20 ± 1 b |
| Procyanidin B2 | 6 ± 0 a | 7 ± 0 b | 7 ± 1 a | 5 ± 0 b |
| Flavonol glycosides | ||||
| Isoquercetin | 59 ± 2 a | n.d. | 52 ± 3 a | n.d. |
| Rutin | 27 ± 2 a | n.d. | 42 ± 4 a | n.d. |
| Stilbenes | ||||
| trans-Resveratrol | 67 ± 4 a | 54 ± 6 b | 79 ± 11 a | 57 ± 3 b |
| E-Viniferin | 33 ± 3 a | 22 ± 2 b | 28 ± 4 a | 33 ± 2 a |
| Piceid | 67 ± 3 a | 2 ± 1 b | 70 ± 4 a | n.d. |
| Phenylethanoids | ||||
| 3-Hydroxytyrosol | 74 ± 5 b | 58 ± 5 a | 80 ± 5 a | 71 ± 3 a |
| Oleuropein | 66 ± 1 a | n.d. | 55 ± 4 a | n.d. |
| Cinnamic acid and derivatives | ||||
| Caffeic acid | 29 ± 4 a | 30 ± 1.2 a | 35 ± 4 a | 26 ± 1 a |
| trans-Cinnamic acid | 32 ± 4 a | 27 ± 1 a | 35 ± 3 a | 31 ± 2 a |
| Chlorogenic acid | 4 ± 1 a | 1 ± 1 a | 3 ± 1 a | n.d. |
| o-Coumaric acid | 61 ± 6 a | 123 ± 8 b | 62 ± 5 a | 109 ± 5 b |
| p-Coumaric acid | 66 ± 3 a | 50 ± 1 a | 52 ± 3 a | 66 ± 1 ab |
| Ferulic acid | 97 ± 6 a | 93 ± 5 a | 83 ± 7 a | 78 ± 2 a |
| Neochlorogenic acid | 6 ± 1 a | 2 ± 1 a | 3 ± 1 a | 1 ± 1 a |
| Sinapic acid | 57 ± 2 a | 16 ± 1 b | 49 ± 5 a | 10 ± 2 b |
| Benzoic acid derivatives | ||||
| Gallic acid | 38 ± 2 a | 24 ± 5 b | 66 ± 2 a | 68 ± 0 a |
| Syringic acid | n.d. | n.d. | n.d. | n.d. |
| Vanillic acid | 55 ± 4 a | 62 ± 5 ab | 44 ± 2 a | 43 ± 1 a |
| Analyte | C | Cenz | CC | CCenz |
|---|---|---|---|---|
| Isoflavones | ||||
| Daidzein | <LOQ | 5.9 ± 2.21 c | 4.35 ± 1.62 ad | 16.89 ± 6.59 d |
| Formononetin | 0.57 ± 0.08 ac | 34.11 ± 7.39 c | 1.85 ± 0.54 ad | 85.45 ± 31.19 d |
| Genistein | <LOQ | 17.83 ± 6.12 bc | <LOQ | 40.4 ± 16.28 bd |
| Flavanones | ||||
| Hesperetin | <LOQ | 2.44 ± 0.79 c | <LOQ | 9.8 ± 3.82 d |
| Naringenin | <LOQ | 3.77 ± 0.83 c | <LOQ | 8.61 ± 2.97 d |
| Flavones | ||||
| Apigenin | <LOQ | 45.75 ± 15.08 bc | 1.53 ± 0.36 ad | 181.66 ± 48.95 bd |
| Chrysin | n.d. | n.d. | <LOQ | 2.54 ± 1.06 bd |
| Luteolin | 0.71 ± 0.44 c | 6.99 ± 3.63 b | 1.07 ± 0.62 cd | 113.74 ± 66.08 bd |
| Flavonols | ||||
| Isorhamnetin | <LOQ | <LOQ | <LOQ | 5.93 ± 2.22 bd |
| Kaempferol | <LOQ | 4.43 ± 3.34 c | <LOQ | 7.52 ± 2.3 d |
| Quercetin | <LOQ | <LOQ | <LOQ | 8.54 ± 4.83 d |
| Cinnamic acid and derivatives | ||||
| trans-Cinnamic acid | 76.8 ± 32.92 c | 188.37 ± 89.04 c | 99.35 ± 41.79 d | 255.57 ± 116.85 d |
| Benzoic acid derivatives | ||||
| Vanillic acid | 47.92 ± 17.64 ac | 705.75 ± 44.9 c | 438.92 ± 162.4 ad | 897.81 ± 173.96 d |
| Syringic acid | 3.45 ± 1.20 c | 17.65 ± 8.33 c | 28.5 ± 8.29 d | 43.2 ± 10.85 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilakopoulou, P.B.; Gousgouni, A.-T.; Yanni, A.E.; Kostomitsopoulos, N.; Karathanos, V.T.; Chiou, A. Polar Phenol Detection in Plasma and Serum: Insights on Sample Pre-Treatment for LC/MS Analysis and Application on the Serum of Corinthian Currant-Fed Rats. Biomolecules 2022, 12, 1838. https://doi.org/10.3390/biom12121838
Vasilakopoulou PB, Gousgouni A-T, Yanni AE, Kostomitsopoulos N, Karathanos VT, Chiou A. Polar Phenol Detection in Plasma and Serum: Insights on Sample Pre-Treatment for LC/MS Analysis and Application on the Serum of Corinthian Currant-Fed Rats. Biomolecules. 2022; 12(12):1838. https://doi.org/10.3390/biom12121838
Chicago/Turabian StyleVasilakopoulou, Paraskevi B., Aimilia-Tatiana Gousgouni, Amalia E. Yanni, Nikolaos Kostomitsopoulos, Vaios T. Karathanos, and Antonia Chiou. 2022. "Polar Phenol Detection in Plasma and Serum: Insights on Sample Pre-Treatment for LC/MS Analysis and Application on the Serum of Corinthian Currant-Fed Rats" Biomolecules 12, no. 12: 1838. https://doi.org/10.3390/biom12121838