
PDX-1: A Promising Therapeutic Target to Reverse Diabetes
 ,
,
Abstract
:1. Introduction
2. Gene Structure and Location, Protein Molecular Structure, Distribution, and Expression of PDX-1
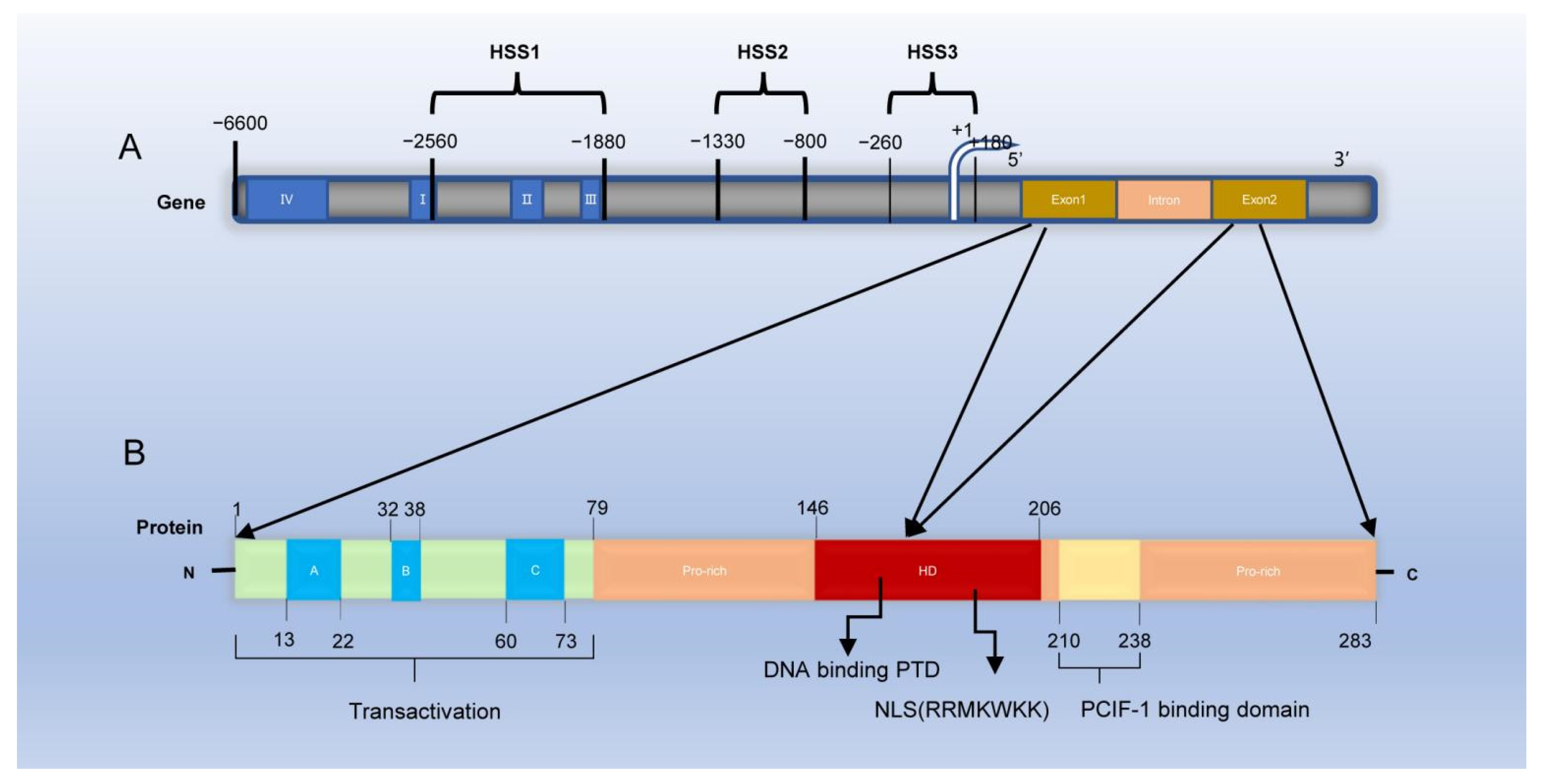
2.1. Localization and Molecular Structure of PDX-1
2.2. Tissue Distribution and Expression of PDX-1
3. Factors Regulating PDX-1 Expression
3.1. Nutrient Substances
3.2. Hormones
3.3. Oxidative Stress
3.4. Cytokines
4. PDX-1 and the Pancreas
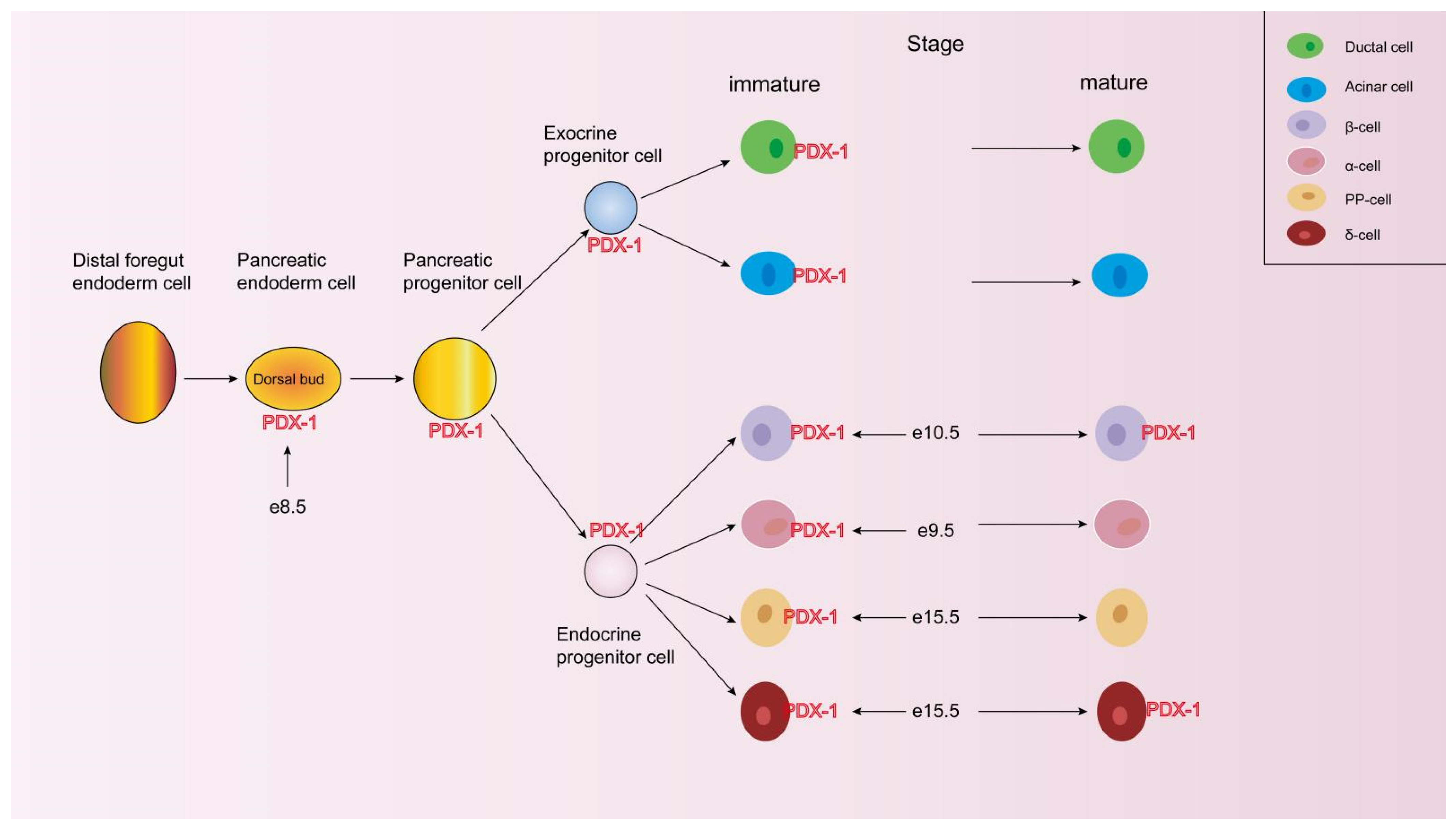
4.1. PDX-1 Promotes Pancreatic Development and Islet Cell Differentiation
4.2. PDX-1 Adjusts the Dedifferentiation, Redifferentiation, and Transdifferentiation of β-Cell
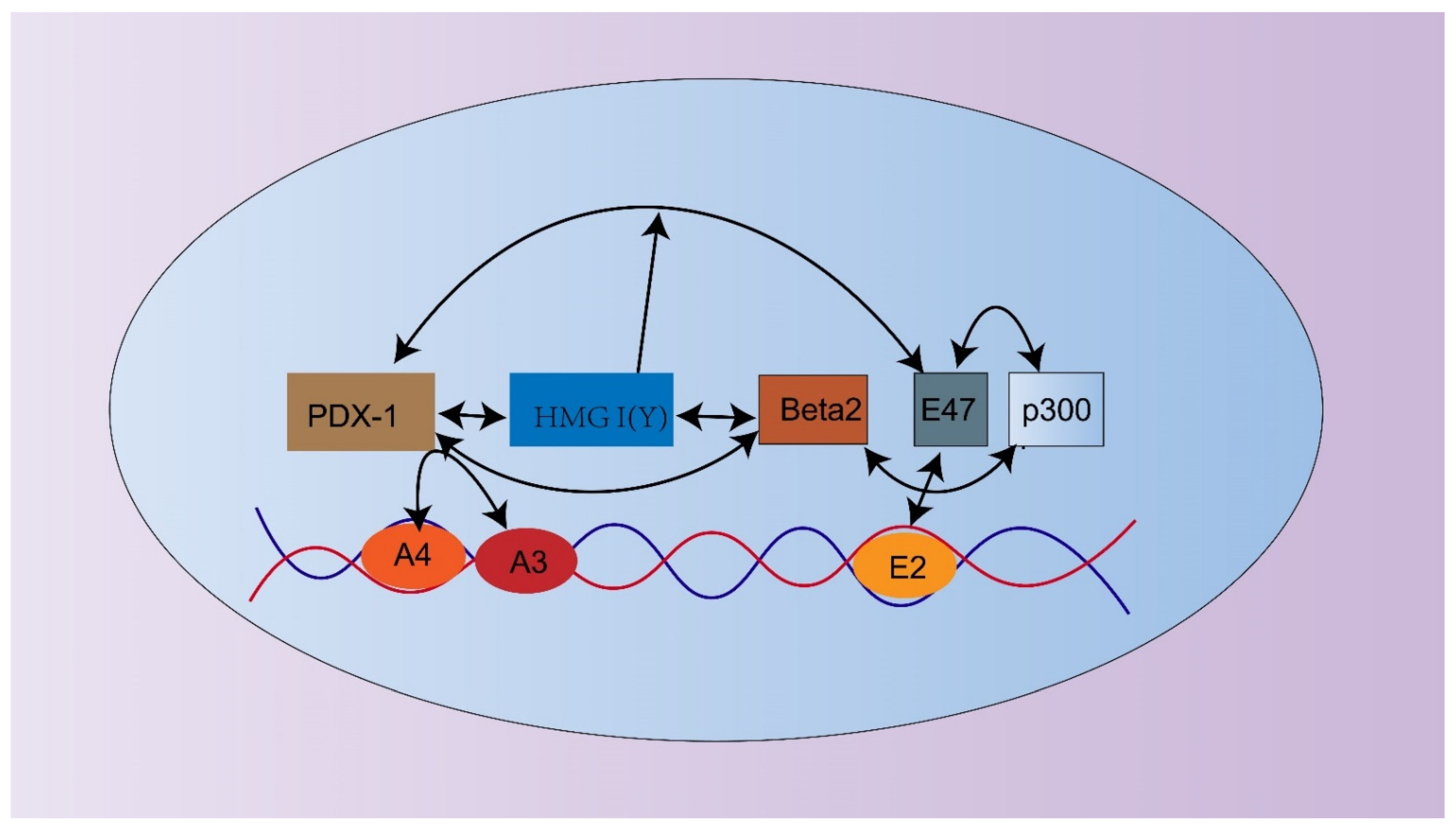
4.3. PDX-1 Regulates the Expression of the Insulin Gene and Insulin Secretion-Related Genes
4.3.1. Insulin
4.3.2. GCK
4.3.3. GLUT-2
4.3.4. IAPP
4.4. Mitochondrial Dysfunction
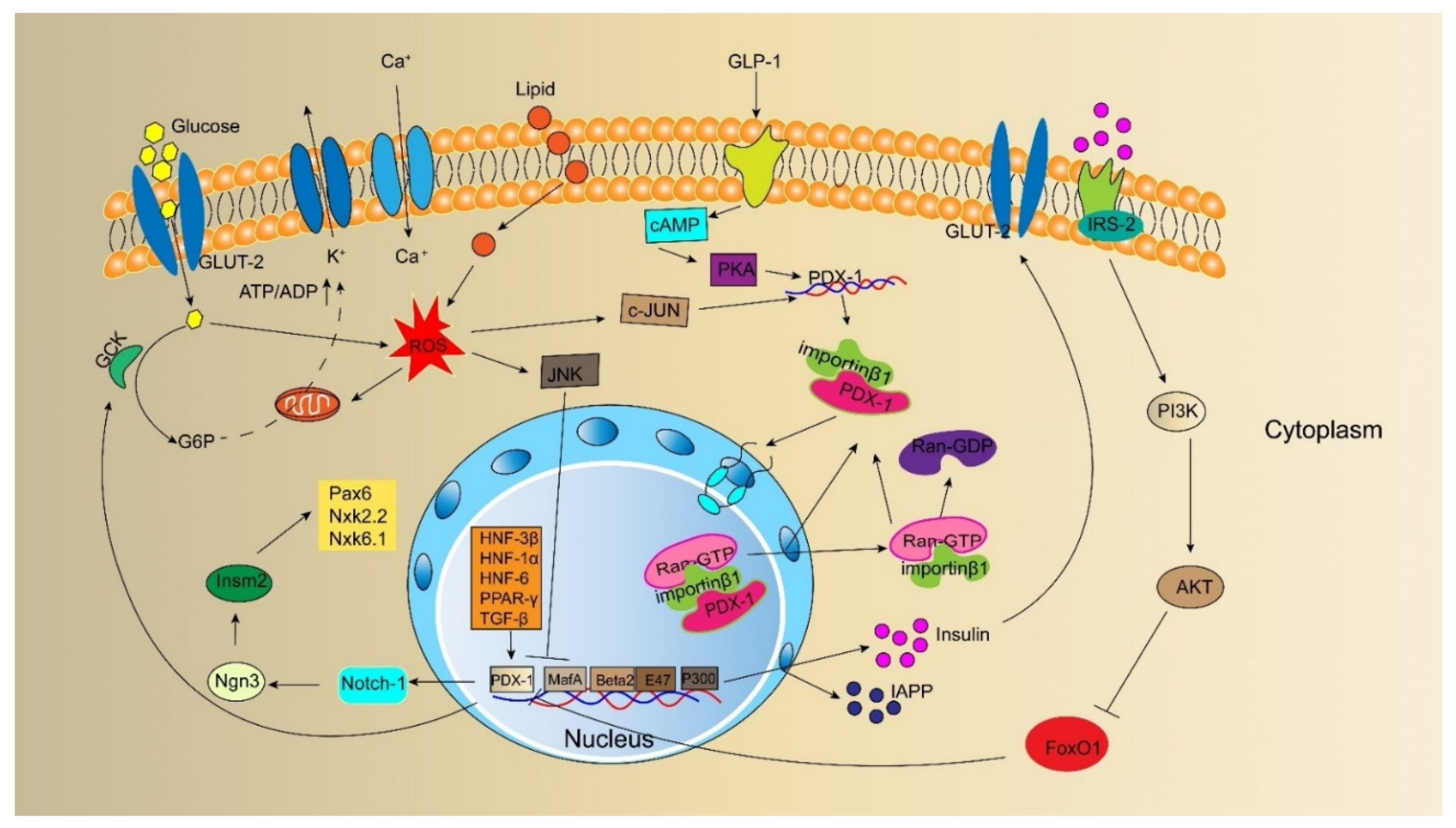
5. PDX-1 Is Involved in the Regulation of the Insulin Signaling Pathway
6. PDX-1 and the Reversal of T2DM
7. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vieira, R.; Souto, S.B.; Sánchez-López, E.; Machado, A.L.; Severino, P.; Jose, S.; Santini, A.; Fortuna, A.; García, M.L.; Silva, A.M.; et al. Sugar-Lowering Drugs for Type 2 Diabetes Mellitus and Metabolic Syndrome-Review of Classical and New Compounds: Part-I. Pharmaceuticals 2019, 12, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IDF Diabetes Atlas. Idf Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Bjornstad, P.; Drews, K.; Zeitler, P.S. Long-Term Complications in Youth-Onset Type 2 Diabetes. Reply. N. Engl. J. Med. 2021, 385, 2016. [Google Scholar] [CrossRef] [PubMed]
- Bolli, G.B.; Gerich, J.E. The “dawn phenomenon”—A common occurrence in both non-insulin-dependent and insulin-dependent diabetes mellitus. N. Engl. J. Med. 1984, 310, 746–750. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2022. Diabetes Care 2022, 45 (Suppl. 1), S17–S38. [Google Scholar] [CrossRef]
- Sperling, M.A.; Laffel, L.M. Current Management of Glycemia in Children with Type 1 Diabetes Mellitus. N. Engl. J. Med. 2022, 386, 1155–1164. [Google Scholar] [CrossRef]
- Syed, F.Z. Type 1 Diabetes Mellitus. Ann. Intern. Med. 2022, 175, Itc33–Itc48. [Google Scholar] [CrossRef]
- Demir, S.; Nawroth, P.P.; Herzig, S.; Ekim Üstünel, B. Emerging Targets in Type 2 Diabetes and Diabetic Complications. Adv. Sci. 2021, 8, e2100275. [Google Scholar] [CrossRef]
- Prasad, R.B.; Groop, L. Genetics of type 2 diabetes-pitfalls and possibilities. Genes 2015, 6, 87–123. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lang, G.; Shi, J. Epigenetic Regulation of PDX-1 in Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 431–442. [Google Scholar] [CrossRef]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Rines, A.K.; Sharabi, K.; Tavares, C.D.; Puigserver, P. Targeting hepatic glucose metabolism in the treatment of type 2 diabetes. Nat. Rev. Drug Discov. 2016, 15, 786–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, M.C.; Cefalu, W.T.; Evans, P.H.; Gerstein, H.C.; Nauck, M.A.; Oh, W.K.; Rothberg, A.E.; le Roux, C.W.; Rubino, F.; Schauer, P.; et al. Consensus Report: Definition and Interpretation of Remission in Type 2 Diabetes. Diabetes Care 2021, 44, 2438–2444. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Al-Mrabeh, A.; Sattar, N. Understanding the mechanisms of reversal of type 2 diabetes. Lancet Diabetes Endocrinol. 2019, 7, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Al-Mrabeh, A.; Zhyzhneuskaya, S.V.; Peters, C.; Barnes, A.C.; Melhem, S.; Jesuthasan, A.; Aribisala, B.; Hollingsworth, K.G.; Lietz, G.; Mathers, J.C.; et al. Hepatic Lipoprotein Export and Remission of Human Type 2 Diabetes after Weight Loss. Cell Metab. 2020, 31, 233–249.e234. [Google Scholar] [CrossRef]
- Wang, Z.; Xiong, H.; Ren, T.Y.S. Repair of Damaged Pancreatic β Cells: New Hope for a Type 2 Diabetes Reversal? J. Transl. Int. Med. 2021, 9, 150–151. [Google Scholar] [CrossRef]
- Singla, R.; Gupta, G.; Dutta, D.; Raizada, N.; Aggarwal, S. Diabetes reversal: Update on current knowledge and proposal of prediction score parameters for diabetes remission. Diabetes Metab. Syndr. 2022, 16, 102452. [Google Scholar] [CrossRef]
- Leonard, J.; Peers, B.; Johnson, T.; Ferreri, K.; Lee, S.; Montminy, M.R. Characterization of somatostatin transactivating factor-1, a novel homeobox factor that stimulates somatostatin expression in pancreatic islet cells. Mol. Endocrinol. 1993, 7, 1275–1283. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, J.; Carlsson, L.; Edlund, T.; Edlund, H. Insulin-promoter-factor 1 is required for pancreas development in mice. Nature 1994, 371, 606–609. [Google Scholar] [CrossRef]
- Dutta, S.; Bonner-Weir, S.; Montminy, M.; Wright, C. Regulatory factor linked to late-onset diabetes? Nature 1998, 392, 560. [Google Scholar] [CrossRef]
- Ferber, S.; Halkin, A.; Cohen, H.; Ber, I.; Einav, Y.; Goldberg, I.; Barshack, I.; Seijffers, R.; Kopolovic, J.; Kaiser, N.; et al. Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nat. Med. 2000, 6, 568–572. [Google Scholar] [CrossRef]
- Noguchi, H.; Kaneto, H.; Weir, G.C.; Bonner-Weir, S. PDX-1 protein containing its own antennapedia-like protein transduction domain can transduce pancreatic duct and islet cells. Diabetes 2003, 52, 1732–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, A.M.; Góñez, L.J.; Naselli, G.; Macdonald, R.J.; Harrison, L.C. Conditional expression demonstrates the role of the homeodomain transcription factor Pdx1 in maintenance and regeneration of beta-cells in the adult pancreas. Diabetes 2005, 54, 2586–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaeth, J.M.; Gupte, M.; Perelis, M.; Yang, Y.P.; Cyphert, H.; Guo, S.; Liu, J.H.; Guo, M.; Bass, J.; Magnuson, M.A.; et al. Defining a Novel Role for the Pdx1 Transcription Factor in Islet β-Cell Maturation and Proliferation During Weaning. Diabetes 2017, 66, 2830–2839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Guo, P.; Shiota, C.; Zhang, T.; Coudriet, G.M.; Fischbach, S.; Prasadan, K.; Fusco, J.; Ramachandran, S.; Witkowski, P.; et al. Endogenous Reprogramming of Alpha Cells into Beta Cells, Induced by Viral Gene Therapy, Reverses Autoimmune Diabetes. Cell Stem Cell 2018, 22, 78–90.e74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuyama, K.; Chera, S.; van Gurp, L.; Oropeza, D.; Ghila, L.; Damond, N.; Vethe, H.; Paulo, J.A.; Joosten, A.M.; Berney, T.; et al. Diabetes relief in mice by glucose-sensing insulin-secreting human α-cells. Nature 2019, 567, 43–48. [Google Scholar] [CrossRef]
- Stoffers, D.A.; Zinkin, N.T.; Stanojevic, V.; Clarke, W.L.; Habener, J.F. Pancreatic agenesis attributable to a single nucleotide deletion in the human IPF1 gene coding sequence. Nat. Genet. 1997, 15, 106–110. [Google Scholar] [CrossRef]
- Gao, T.; McKenna, B.; Li, C.; Reichert, M.; Nguyen, J.; Singh, T.; Yang, C.; Pannikar, A.; Doliba, N.; Zhang, T.; et al. Pdx1 maintains β cell identity and function by repressing an α cell program. Cell Metab. 2014, 19, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Scott, V.; Clark, A.R.; Hutton, J.C.; Docherty, K. Two proteins act as the IUF1 insulin gene enhancer binding factor. FEBS Lett. 1991, 290, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Ohlsson, H.; Karlsson, K.; Edlund, T. IPF1, a homeodomain-containing transactivator of the insulin gene. EMBO J. 1993, 12, 4251–4259. [Google Scholar] [CrossRef]
- Miller, C.P.; McGehee, R.E., Jr.; Habener, J.F. IDX-1: A new homeodomain transcription factor expressed in rat pancreatic islets and duodenum that transactivates the somatostatin gene. EMBO J. 1994, 13, 1145–1156. [Google Scholar] [CrossRef]
- Wright, C.V.; Schnegelsberg, P.; De Robertis, E.M. XlHbox 8: A novel Xenopus homeo protein restricted to a narrow band of endoderm. Development 1989, 105, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, N.; Serikawa, T.; Walther, R. Pdx1, a homeodomain transcription factor required for pancreas development, maps to rat chromosome 12. Exp. Anim. 1997, 46, 323–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Leonard, J.; Lee, S.; Chapman, H.D.; Leiter, E.H.; Montminy, M.R. Pancreatic islet expression of the homeobox factor STF-1 relies on an E-box motif that binds USF. J. Biol. Chem. 1996, 271, 2294–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffel, M.; Stein, R.; Wright, C.V.; Espinosa, R., 3rd; Le Beau, M.M.; Bell, G.I. Localization of human homeodomain transcription factor insulin promoter factor 1 (IPF1) to chromosome band 13q12.1. Genomics 1995, 28, 125–126. [Google Scholar] [CrossRef]
- Inoue, H.; Riggs, A.C.; Tanizawa, Y.; Ueda, K.; Kuwano, A.; Liu, L.; Donis-Keller, H.; Permutt, M.A. Isolation, characterization, and chromosomal mapping of the human insulin promoter factor 1 (IPF-1) gene. Diabetes 1996, 45, 789–794. [Google Scholar] [CrossRef]
- Melloul, D.; Marshak, S.; Cerasi, E. Regulation of pdx-1 gene expression. Diabetes 2002, 51 (Suppl. 3), S320–S325. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.L.; Gannon, M.; Peshavaria, M.; Offield, M.F.; Henderson, E.; Ray, M.; Marks, A.; Gamer, L.W.; Wright, C.V.; Stein, R. Hepatocyte nuclear factor 3beta is involved in pancreatic beta-cell-specific transcription of the pdx-1 gene. Mol. Cell. Biol. 1997, 17, 6002–6013. [Google Scholar] [CrossRef] [Green Version]
- Gerrish, K.; Gannon, M.; Shih, D.; Henderson, E.; Stoffel, M.; Wright, C.V.E.; Stein, R. Pancreatic beta cell-specific transcription of the pdx-1 gene. The role of conserved upstream control regions and their hepatic nuclear factor 3beta sites. J. Biol. Chem. 2000, 275, 3485–3492. [Google Scholar] [CrossRef] [Green Version]
- Gerrish, K.; Van Velkinburgh, J.C.; Stein, R. Conserved transcriptional regulatory domains of the pdx-1 gene. Mol. Endocrinol. 2004, 18, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Samaras, S.E.; Cissell, M.A.; Gerrish, K.; Wright, C.V.E.; Gannon, M.; Stein, R. Conserved sequences in a tissue-specific regulatory region of the pdx-1 gene mediate transcription in Pancreatic beta cells: Role for hepatocyte nuclear factor 3 beta and Pax6. Mol. Cell. Biol. 2002, 22, 4702–4713. [Google Scholar] [CrossRef]
- Wiebe, P.O.; Kormish, J.D.; Roper, V.T.; Fujitani, Y.; Alston, N.I.; Zaret, K.S.; Wright, C.V.E.; Stein, R.W.; Gannon, M. Ptf1a binds to and activates area III, a highly conserved region of the Pdx1 promoter that mediates early pancreas-wide Pdx1 expression. Mol. Cell. Biol. 2007, 27, 4093–4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffers, D.; Stanojevic, V.; Habener, J.F. Insulin promoter factor-1 gene mutation linked to early-onset type 2 diabetes mellitus directs expression of a dominant negative isoprotein. J. Clin. Investig. 1998, 102, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, W.M.; Frayling, T.; Ellard, S.; Evans, J.C.; Allen, L.I.; Bulman, M.P.; Ayres, S.; Shepherd, M.; Clark, P.; Millward, A.; et al. Missense mutations in the insulin promoter factor-1 gene predispose to type 2 diabetes. J. Clin. Investig. 1999, 104, R33–R39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, J.; Macfarlane, W.M.; Lehto, M.; Gu, H.F.; Shepherd, L.M.; Ivarsson, S.A.; Wibell, L.; Smith, T.; Groop, L.C. Functional consequences of mutations in the MODY4 gene (IPF1) and coexistence with MODY3 mutations. Diabetologia 2001, 44, 249–258. [Google Scholar] [CrossRef]
- McKinnon, C.M.; Docherty, K. Pancreatic duodenal homeobox-1, PDX-1, a major regulator of beta cell identity and function. Diabetologia 2001, 44, 1203–1214. [Google Scholar] [CrossRef]
- Qiu, Y.; Guo, M.; Huang, S.; Stein, R. Insulin gene transcription is mediated by interactions between the p300 coactivator and PDX-1, BETA2, and E47. Mol. Cell Biol. 2002, 22, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Hui, H.; Perfetti, R. Pancreas duodenum homeobox-1 regulates pancreas development during embryogenesis and islet cell function in adulthood. Eur. J. Endocrinol. 2002, 146, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moede, T.; Leibiger, B.; Pour, H.G.; Berggren, P.; Leibiger, I.B. Identification of a nuclear localization signal, RRMKWKK, in the homeodomain transcription factor PDX-1. FEBS Lett. 1999, 461, 229–234. [Google Scholar] [CrossRef]
- Macfarlane, W.M.; McKinnon, C.M.; Felton-Edkins, Z.A.; Cragg, H.; James, R.F.; Docherty, K. Glucose stimulates translocation of the homeodomain transcription factor PDX1 from the cytoplasm to the nucleus in pancreatic beta-cells. J. Biol. Chem. 1999, 274, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Elrick, L.J.; Docherty, K. Phosphorylation-dependent nucleocytoplasmic shuttling of pancreatic duodenal homeobox-1. Diabetes 2001, 50, 2244–2252. [Google Scholar] [CrossRef]
- Kishi, A.; Nakamura, T.; Nishio, Y.; Maegawa, H.; Kashiwagi, A. Sumoylation of Pdx1 is associated with its nuclear localization and insulin gene activation. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E830–E840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessabi, B.; Ziegler, P.; Schmidt, I.; Hessabi, C.; Walther, R. The nuclear localization signal (NLS) of PDX-1 is part of the homeodomain and represents a novel type of NLS. Eur. J. Biochem. 1999, 263, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Guillemain, G.; Da Silva Xavier, G.; Rafiq, I.; Leturque, A.; Rutter, G.A. Importin beta1 mediates the glucose-stimulated nuclear import of pancreatic and duodenal homeobox-1 in pancreatic islet beta-cells (MIN6). Biochem. J. 2004, 378, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, H.; Matsushita, M.; Matsumoto, S.; Lu, Y.-F.; Matsui, H.; Bonner-Weir, S. Mechanism of PDX-1 protein transduction. Biochem. Biophys. Res. Commun. 2005, 332, 68–74. [Google Scholar] [CrossRef]
- Liu, A.; Desai, B.M.; Stoffers, D.A. Identification of PCIF1, a POZ domain protein that inhibits PDX-1 (MODY4) transcriptional activity. Mol. Cell Biol. 2004, 24, 4372–4383. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, R.K.; Yu, S.M.; Flores, L.E.; Jhala, U.S. Glucose regulates steady-state levels of PDX1 via the reciprocal actions of GSK3 and AKT kinases. J. Biol. Chem. 2010, 285, 3406–3416. [Google Scholar] [CrossRef] [Green Version]
- Guz, Y.; Montminy, M.R.; Stein, R.; Leonard, J.; Gamer, L.W.; Wright, C.V.; Teitelman, G. Expression of murine STF-1, a putative insulin gene transcription factor, in beta cells of pancreas, duodenal epithelium and pancreatic exocrine and endocrine progenitors during ontogeny. Development 1995, 121, 11–18. [Google Scholar] [CrossRef]
- Sander, M.; German, M.S. The beta cell transcription factors and development of the pancreas. J. Mol. Med. 1997, 75, 327–340. [Google Scholar] [CrossRef]
- Stoffers, D.A.; Heller, R.S.; Miller, C.P.; Habener, J.F. Developmental expression of the homeodomain protein IDX-1 in mice transgenic for an IDX-1 promoter/lacZ transcriptional reporter. Endocrinology 1999, 140, 5374–5381. [Google Scholar] [CrossRef]
- Perez-Villamil, B.; Schwartz, P.T.; Vallejo, M. The pancreatic homeodomain transcription factor IDX1/IPF1 is expressed in neural cells during brain development. Endocrinology 1999, 140, 3857–3860. [Google Scholar] [CrossRef]
- Sakai, H.; Eishi, Y.; Li, X.L.; Akiyama, Y.; Miyake, S.; Takizawa, T.; Konishi, N.; Tatematsu, M.; Koike, M.; Yuasa, Y. PDX1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut 2004, 53, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballian, N.; Liu, S.H.; Brunicardi, F.C. Transcription factor PDX-1 in human colorectal adenocarcinoma: A potential tumor marker? World J. Gastroenterol. 2008, 14, 5823–5826. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Gou, S.M.; Wang, C.Y.; Wu, H.S.; Xiong, J.X.; Zhou, F. Pancreas duodenal homeobox-1 expression and significance in pancreatic cancer. World J. Gastroenterol. 2007, 13, 2615–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.P.; Li, Z.J.; Magnusson, J.; Brunicardi, F.C. Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J. Surg. 2005, 29, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Evans-Molina, C.; Garmey, J.C.; Ketchum, R.; Brayman, K.L.; Deng, S.; Mirmira, R.G. Glucose regulation of insulin gene transcription and pre-mRNA processing in human islets. Diabetes 2007, 56, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Ahlgren, U.; Jonsson, J.; Jonsson, L.; Simu, K.; Edlund, H. beta-cell-specific inactivation of the mouse Ipf1/Pdx1 gene results in loss of the beta-cell phenotype and maturity onset diabetes. Genes Dev. 1998, 12, 1763–1768. [Google Scholar] [CrossRef] [Green Version]
- Gremlich, S.; Bonny, C.; Waeber, G.; Thorens, B. Fatty acids decrease IDX-1 expression in rat pancreatic islets and reduce GLUT2, glucokinase, insulin, and somatostatin levels. J. Biol. Chem. 1997, 272, 30261–30269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagman, D.K.; Hays, L.B.; Parazzoli, S.D.; Poitout, V. Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J. Biol. Chem. 2005, 280, 32413–32418. [Google Scholar] [CrossRef] [Green Version]
- Shimo, N.; Matsuoka, T.A.; Miyatsuka, T.; Takebe, S.; Tochino, Y.; Takahara, M.; Kaneto, H.; Shimomura, I. Short-term selective alleviation of glucotoxicity and lipotoxicity ameliorates the suppressed expression of key β-cell factors under diabetic conditions. Biochem. Biophys. Res. Commun. 2015, 467, 948–954. [Google Scholar] [CrossRef]
- McLean, B.A.; Wong, C.K.; Campbell, J.E.; Hodson, D.J.; Trapp, S.; Drucker, D.J. Revisiting the Complexity of GLP-1 Action from Sites of Synthesis to Receptor Activation. Endocr. Rev. 2021, 42, 101–132. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, J.; Doyle, M.E.; Egan, J.M. Glucagon-like peptide-1 causes pancreatic duodenal homeobox-1 protein translocation from the cytoplasm to the nucleus of pancreatic beta-cells by a cyclic adenosine monophosphate/protein kinase A-dependent mechanism. Endocrinology 2001, 142, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cahill, C.M.; Piñeyro, M.A.; Zhou, J.; Doyle, M.E.; Egan, J.M. Glucagon-like peptide-1 regulates the beta cell transcription factor, PDX-1, in insulinoma cells. Endocrinology 1999, 140, 4904–4907. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.L.; Kwon, O.; Kim, S.G.; Lee, I.K.; Kim, Y.D. B-cell translocation gene 2 positively regulates GLP-1-stimulated insulin secretion via induction of PDX-1 in pancreatic β-cells. Exp. Mol. Med. 2013, 45, e25. [Google Scholar] [CrossRef] [Green Version]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Dohi, T.; Martin, N.M.; Raskett, C.M.; Liu, Q.; Altieri, D.C. Essential role of the small GTPase Ran in postnatal pancreatic islet development. PLoS ONE 2011, 6, e27879. [Google Scholar] [CrossRef] [Green Version]
- Perfetti, R.; Zhou, J.; Doyle, M.E.; Egan, J.M. Glucagon-like peptide-1 induces cell proliferation and pancreatic-duodenum homeobox-1 expression and increases endocrine cell mass in the pancreas of old, glucose-intolerant rats. Endocrinology 2000, 141, 4600–4605. [Google Scholar] [CrossRef]
- Baeeri, M.; Rahimifard, M.; Daghighi, S.M.; Khan, F.; Salami, S.A.; Moini-Nodeh, S.; Haghi-Aminjan, H.; Bayrami, Z.; Rezaee, F.; Abdollahi, M. Cannabinoids as anti-ROS in aged pancreatic islet cells. Life Sci. 2020, 256, 117969. [Google Scholar] [CrossRef]
- Leenders, F.; Groen, N.; de Graaf, N.; Engelse, M.A.; Rabelink, T.J.; de Koning, E.J.P.; Carlotti, F. Oxidative Stress Leads to β-Cell Dysfunction Through Loss of β-Cell Identity. Front. Immunol. 2021, 12, 690379. [Google Scholar] [CrossRef]
- Matsuoka, T.; Kajimoto, Y.; Watada, H.; Kaneto, H.; Kishimoto, M.; Umayahara, Y.; Fujitani, Y.; Kamada, T.; Kawamori, R.; Yamasaki, Y. Glycation-dependent, reactive oxygen species-mediated suppression of the insulin gene promoter activity in HIT cells. J. Clin. Investig. 1997, 99, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Kawamori, D.; Kajimoto, Y.; Kaneto, H.; Umayahara, Y.; Fujitani, Y.; Miyatsuka, T.; Watada, H.; Leibiger, I.B.; Yamasaki, Y.; Hori, M. Oxidative stress induces nucleo-cytoplasmic translocation of pancreatic transcription factor PDX-1 through activation of c-Jun NH(2)-terminal kinase. Diabetes 2003, 52, 2896–2904. [Google Scholar] [CrossRef]
- Liang, H.; Pan, Y.; Teng, Y.; Yuan, S.; Wu, X.; Yang, H.; Zhou, P. A proteoglycan extract from Ganoderma Lucidum protects pancreatic beta-cells against STZ-induced apoptosis. Biosci. Biotechnol. Biochem. 2020, 84, 2491–2498. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Kajimoto, Y.; Miyagawa, J.; Matsuoka, T.; Fujitani, Y.; Umayahara, Y.; Hanafusa, T.; Matsuzawa, Y.; Yamasaki, Y.; Hori, M. Beneficial effects of antioxidants in diabetes: Possible protection of pancreatic beta-cells against glucose toxicity. Diabetes 1999, 48, 2398–2406. [Google Scholar] [CrossRef] [PubMed]
- Baumel-Alterzon, S.; Scott, D.K. Regulation of Pdx1 by oxidative stress and Nrf2 in pancreatic beta-cells. Front. Endocrinol. 2022, 13, 1011187. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; LeLay, J.; Vatamaniuk, M.Z.; Rieck, S.; Friedman, J.R.; Kaestner, K.H. Dynamic regulation of Pdx1 enhancers by Foxa1 and Foxa2 is essential for pancreas development. Genes Dev. 2008, 22, 3435–3448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshak, S.; Benshushan, E.; Shoshkes, M.; Havin, L.; Cerasi, E.; Melloul, D. Functional conservation of regulatory elements in the pdx-1 gene: PDX-1 and hepatocyte nuclear factor 3beta transcription factors mediate beta-cell-specific expression. Mol. Cell Biol. 2000, 20, 7583–7590. [Google Scholar] [CrossRef] [Green Version]
- Jacquemin, P.; Lemaigre, F.P.; Rousseau, G.G. The Onecut transcription factor HNF-6 (OC-1) is required for timely specification of the pancreas and acts upstream of Pdx-1 in the specification cascade. Dev. Biol. 2003, 258, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Sayo, Y.; Hosokawa, H.; Imachi, H.; Murao, K.; Sato, M.; Wong, N.C.; Ishida, T.; Takahara, J. Transforming growth factor beta induction of insulin gene expression is mediated by pancreatic and duodenal homeobox gene-1 in rat insulinoma cells. Eur. J. Biochem. 2000, 267, 971–978. [Google Scholar] [CrossRef]
- Moibi, J.A.; Gupta, D.; Jetton, T.L.; Peshavaria, M.; Desai, R.; Leahy, J.L. Peroxisome proliferator-activated receptor-gamma regulates expression of PDX-1 and NKX6.1 in INS-1 cells. Diabetes 2007, 56, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Honoré, C.; Rescan, C.; Hald, J.; McGrath, P.S.; Petersen, M.B.; Hansson, M.; Klein, T.; Østergaard, S.; Wells, J.M.; Madsen, O.D. Revisiting the immunocytochemical detection of Neurogenin 3 expression in mouse and man. Diabetes Obes. Metab. 2016, 18 (Suppl. 1), 10–22. [Google Scholar] [CrossRef]
- Oliver-Krasinski, J.M.; Kasner, M.T.; Yang, J.; Crutchlow, M.F.; Rustgi, A.K.; Kaestner, K.H.; Stoffers, D.A. The diabetes gene Pdx1 regulates the transcriptional network of pancreatic endocrine progenitor cells in mice. J. Clin. Investig. 2009, 119, 1888–1898. [Google Scholar] [CrossRef]
- Edlund, H. Developmental biology of the pancreas. Diabetes 2001, 50 (Suppl. 1), S5–S9. [Google Scholar] [CrossRef] [Green Version]
- Yoshiji, S.; Horikawa, Y.; Kubota, S.; Enya, M.; Iwasaki, Y.; Keidai, Y.; Aizawa-Abe, M.; Iwasaki, K.; Honjo, S.; Hosomichi, K.; et al. First Japanese Family With PDX1-MODY (MODY4): A Novel PDX1 Frameshift Mutation, Clinical Characteristics, and Implications. J. Endocr. Soc. 2022, 6, bvab159. [Google Scholar] [CrossRef] [PubMed]
- Wiggenhauser, L.M.; Metzger, L.; Bennewitz, K.; Soleymani, S.; Boger, M.; Tabler, C.T.; Hausser, I.; Sticht, C.; Wohlfart, P.; Volk, N.; et al. pdx1 Knockout Leads to a Diabetic Nephropathy- Like Phenotype in Zebrafish and Identifies Phosphatidylethanolamine as Metabolite Promoting Early Diabetic Kidney Damage. Diabetes 2022, 71, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Kamisako, T.; Kagawa, T.; Haraguchi, S.; Yagoto, M.; Takahashi, R.; Kawai, K.; Suemizu, H. Expression of pancreatic and duodenal homeobox1 (PDX1) protein in the interior and exterior regions of the intestine, revealed by development and analysis of Pdx1 knockout mice. Lab. Anim. Res. 2015, 31, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilarino, M.; Rashid, S.T.; Suchy, F.P.; McNabb, B.R.; van der Meulen, T.; Fine, E.J.; Ahsan, S.D.; Mursaliyev, N.; Sebastiano, V.; Diab, S.S.; et al. CRISPR/Cas9 microinjection in oocytes disables pancreas development in sheep. Sci. Rep. 2017, 7, 17472. [Google Scholar] [CrossRef] [Green Version]
- Spaeth, J.M.; Liu, J.H.; Peters, D.; Guo, M.; Osipovich, A.B.; Mohammadi, F.; Roy, N.; Bhushan, A.; Magnuson, M.A.; Hebrok, M.; et al. The Pdx1-Bound Swi/Snf Chromatin Remodeling Complex Regulates Pancreatic Progenitor Cell Proliferation and Mature Islet β-Cell Function. Diabetes 2019, 68, 1806–1818. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.D.; Ahmed, N.T.; Luciani, D.S.; Han, Z.; Tran, H.; Fujita, J.; Misler, S.; Edlund, H.; Polonsky, K.S. Increased islet apoptosis in Pdx1+/- mice. J. Clin. Investig. 2003, 111, 1147–1160. [Google Scholar] [CrossRef]
- Talchai, C.; Xuan, S.; Lin, H.V.; Sussel, L.; Accili, D. Pancreatic β cell dedifferentiation as a mechanism of diabetic β cell failure. Cell 2012, 150, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Cim, A.; Sawyer, G.J.; Zhang, X.; Su, H.; Collins, L.; Jones, P.; Antoniou, M.; Reynes, J.P.; Lipps, H.J.; Fabre, J.W. In vivo studies on non-viral transdifferentiation of liver cells towards pancreatic β cells. J. Endocrinol. 2012, 214, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Kajimoto, Y.; Yasuda, T.; Watada, H.; Fujitani, Y.; Kosaka, H.; Gotow, T.; Miyatsuka, T.; Umayahara, Y.; Yamasaki, Y.; et al. PDX-1 induces differentiation of intestinal epithelioid IEC-6 into insulin-producing cells. Diabetes 2002, 51, 2505–2513. [Google Scholar] [CrossRef]
- Van Pham, P.; Thi-My Nguyen, P.; Thai-Quynh Nguyen, A.; Minh Pham, V.; Nguyen-Tu Bui, A.; Thi-Tung Dang, L.; Gia Nguyen, K.; Kim Phan, N. Improved differentiation of umbilical cord blood-derived mesenchymal stem cells into insulin-producing cells by PDX-1 mRNA transfection. Differentiation 2014, 87, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, Y.; Wang, X.; Song, J.; Jia, Y. Expression of Pdx-1 in bone marrow mesenchymal stem cells promotes differentiation of islet-like cells in vitro. Sci. China C Life Sci. 2006, 49, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yuan, Y.; Chen, Y. PDX1 transfection induces human adipose derived stem cells differentiation into islet-like cells: What is the benefit for diabetic rats? Pharmazie 2018, 73, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Ramzy, A.; Thompson, D.M.; Ward-Hartstonge, K.A.; Ivison, S.; Cook, L.; Garcia, R.V.; Loyal, J.; Kim, P.T.W.; Warnock, G.L.; Levings, M.K.; et al. Implanted pluripotent stem-cell-derived pancreatic endoderm cells secrete glucose-responsive C-peptide in patients with type 1 diabetes. Cell Stem Cell 2021, 28, 2047–2061.e5. [Google Scholar] [CrossRef]
- Petersen, H.V.; Serup, P.; Leonard, J.; Michelsen, B.K.; Madsen, O.D. Transcriptional regulation of the human insulin gene is dependent on the homeodomain protein STF1/IPF1 acting through the CT boxes. Proc. Natl. Acad. Sci. USA 1994, 91, 10465–10469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmon, J.S.; Stein, R.; Robertson, R.P. Oxidative stress-mediated, post-translational loss of MafA protein as a contributing mechanism to loss of insulin gene expression in glucotoxic beta cells. J. Biol. Chem. 2005, 280, 11107–11113. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Guo, M.; Matsuoka, T.A.; Hagman, D.K.; Parazzoli, S.D.; Poitout, V.; Stein, R. The islet beta cell-enriched MafA activator is a key regulator of insulin gene transcription. J. Biol. Chem. 2005, 280, 11887–11894. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, O.; Edlund, T.; Moss, J.B.; Rutter, W.J.; Walker, M.D. A mutational analysis of the insulin gene transcription control region: Expression in beta cells is dependent on two related sequences within the enhancer. Proc. Natl. Acad. Sci. USA 1987, 84, 8819–8823. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Miyatsuka, T.; Sasaki, S.; Miyashita, K.; Kubo, F.; Shimo, N.; Takebe, S.; Watada, H.; Kaneto, H.; Matsuoka, T.A.; et al. Preserving expression of Pdx1 improves β-cell failure in diabetic mice. Biochem. Biophys. Res. Commun. 2017, 483, 418–424. [Google Scholar] [CrossRef]
- Ohneda, K.; Mirmira, R.G.; Wang, J.; Johnson, J.D.; German, M.S. The homeodomain of PDX-1 mediates multiple protein-protein interactions in the formation of a transcriptional activation complex on the insulin promoter. Mol. Cell Biol. 2000, 20, 900–911. [Google Scholar] [CrossRef]
- Arcidiacono, B.; Iiritano, S.; Chiefari, E.; Brunetti, F.S.; Gu, G.; Foti, D.P.; Brunetti, A. Cooperation between HMGA1, PDX-1, and MafA is Essential for Glucose-Induced Insulin Transcription in Pancreatic Beta Cells. Front. Endocrinol. 2014, 5, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Li, L.; Wan, L.; Huang, Y.; Cao, S. Glucokinase as an emerging anti-diabetes target and recent progress in the development of its agonists. J. Enzym. Inhib. Med. Chem. 2022, 37, 606–615. [Google Scholar] [CrossRef]
- Watada, H.; Kajimoto, Y.; Umayahara, Y.; Matsuoka, T.; Kaneto, H.; Fujitani, Y.; Kamada, T.; Kawamori, R.; Yamasaki, Y. The human glucokinase gene beta-cell-type promoter: An essential role of insulin promoter factor 1/PDX-1 in its activation in HIT-T15 cells. Diabetes 1996, 45, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, R.; Qiao, H.; Zhang, H.; Wang, Y.; Yuan, H.; Liu, Q.; Liu, D.; Chen, L.; Pei, X. Generation of insulin-producing cells from PDX-1 gene-modified human mesenchymal stem cells. J. Cell Physiol. 2007, 211, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Watada, H.; Kajimoto, Y.; Miyagawa, J.; Hanafusa, T.; Hamaguchi, K.; Matsuoka, T.; Yamamoto, K.; Matsuzawa, Y.; Kawamori, R.; Yamasaki, Y. PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes 1996, 45, 1826–1831. [Google Scholar] [CrossRef] [Green Version]
- Thorens, B. GLUT2, glucose sensing and glucose homeostasis. Diabetologia 2015, 58, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waeber, G.; Thompson, N.; Nicod, P.; Bonny, C. Transcriptional activation of the GLUT2 gene by the IPF-1/STF-1/IDX-1 homeobox factor. Mol. Endocrinol. 1996, 10, 1327–1334. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, N.; Wang, L.; Guo, H.; Guan, Q.; Cui, B.; Tian, L.; Gao, L.; Zhao, J. AMP-activated protein kinase and pancreatic/duodenal homeobox-1 involved in insulin secretion under high leucine exposure in rat insulinoma beta-cells. J. Cell Mol. Med. 2009, 13, 758–770. [Google Scholar] [CrossRef]
- Watada, H.; Kajimoto, Y.; Kaneto, H.; Matsuoka, T.; Fujitani, Y.; Miyazaki, J.; Yamasaki, Y. Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem. Biophys. Res. Commun. 1996, 229, 746–751. [Google Scholar] [CrossRef]
- Carty, M.D.; Lillquist, J.S.; Peshavaria, M.; Stein, R.; Soeller, W.C. Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J. Biol. Chem. 1997, 272, 11986–11993. [Google Scholar] [CrossRef]
- Akesson, B.; Panagiotidis, G.; Westermark, P.; Lundquist, I. Islet amyloid polypeptide inhibits glucagon release and exerts a dual action on insulin release from isolated islets. Regul. Pept. 2003, 111, 55–60. [Google Scholar] [CrossRef]
- Smith, S.B.; Watada, H.; Scheel, D.W.; Mrejen, C.; German, M.S. Autoregulation and maturity onset diabetes of the young transcription factors control the human PAX4 promoter. J. Biol. Chem. 2000, 275, 36910–36919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, R.N.; Jhala, U.S.; Winnay, J.N.; Krajewski, S.; Montminy, M.; Kahn, C.R. PDX-1 haploinsufficiency limits the compensatory islet hyperplasia that occurs in response to insulin resistance. J. Clin. Investig. 2004, 114, 828–836. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Kim, W.H.; Lim, J.H.; Song, E.H.; Song, J.; Choi, K.Y.; Jung, M.H. Mitochondrial dysfunction: Glucokinase downregulation lowers interaction of glucokinase with mitochondria, resulting in apoptosis of pancreatic beta-cells. Cell. Signal. 2009, 21, 69–78. [Google Scholar] [CrossRef]
- Soleimanpour, S.A.; Ferrari, A.M.; Raum, J.C.; Groff, D.N.; Yang, J.; Kaufman, B.A.; Stoffers, D.A. Diabetes Susceptibility Genes Pdx1 and Clec16a Function in a Pathway Regulating Mitophagy in β-Cells. Diabetes 2015, 64, 3475–3484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, R.; Jo, S.; Lockridge, A.; Ferrington, D.A.; Murray, K.; Eschenlauer, A.; Bernal-Mizrachi, E.; Fujitani, Y.; Alejandro, E.U. OGT Regulates Mitochondrial Biogenesis and Function via Diabetes Susceptibility Gene Pdx1. Diabetes 2021, 70, 2608–2625. [Google Scholar] [CrossRef]
- Gauthier, B.R.; Wiederkehr, A.; Baquié, M.; Dai, C.; Powers, A.C.; Kerr-Conte, J.; Pattou, F.; MacDonald, R.J.; Ferrer, J.; Wollheim, C.B. PDX1 deficiency causes mitochondrial dysfunction and defective insulin secretion through TFAM suppression. Cell Metab. 2009, 10, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, T.; Zhang, H.; Li, S.; Tian, H. Glucagon-like peptide 1 receptor agonist ameliorates the insulin resistance function of islet β cells via the activation of PDX-1/JAK signaling transduction in C57/BL6 mice with high-fat diet-induced diabetes. Int. J. Mol. Med. 2017, 39, 1029–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, T.; Nakae, J.; Kitamura, Y.; Kido, Y.; Biggs, W.H., 3rd; Wright, C.V.; White, M.F.; Arden, K.C.; Accili, D. The forkhead transcription factor Foxo1 links insulin signaling to Pdx1 regulation of pancreatic beta cell growth. J. Clin. Investig. 2002, 110, 1839–1847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Sun, P.; Shen, C.; Liu, X.; Sun, J.; Li, D.; Liu, Z.; Zhang, W.; Zhang, K.; Niu, Y. Role and mechanism of PI3K/AKT/FoxO1/PDX-1 signaling pathway in functional changes of pancreatic islets in rats after severe burns. Life Sci. 2020, 258, 118145. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, L.; Zhang, Y.; Yan, S.; Huang, L. Improvement of Lipotoxicity-Induced Islet β Cellular Insulin Secretion Disorder by Osteocalcin. J. Diabetes Res. 2022, 2022, 3025538. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, G.; Kaiser, N.; Cerasi, E. β-Cell failure in type 2 diabetes. J. Diabetes Investig. 2011, 2, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.P.; Thorel, F.; Boyer, D.F.; Herrera, P.L.; Wright, C.V. Context-specific α- to-β-cell reprogramming by forced Pdx1 expression. Genes Dev. 2011, 25, 1680–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Fu, J.; Zhang, Z.; Zhang, Y.; Liang, Y.; Ge, C.; Qin, X. Pancreatic islet regeneration through PDX-1/Notch-1/Ngn3 signaling after gastric bypass surgery in db/db mice. Exp. Ther. Med. 2017, 14, 2831–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, H.; Fujimiya, M.; Matsumura, K.; Younan, P.; Imaeda, H.; Maeda, M.; Chan, L. NeuroD-betacellulin gene therapy induces islet neogenesis in the liver and reverses diabetes in mice. Nat. Med. 2003, 9, 596–603. [Google Scholar] [CrossRef]
- Bahrebar, M.; Soleimani, M.; Karimi, M.H.; Vahdati, A.; Yaghobi, R. Generation of Islet-like Cell Aggregates from Human Adipose Tissue-derived Stem Cells by Lentiviral Overexpression of PDX-1. Int. J. Organ. Transpl. Med. 2015, 6, 61–76. [Google Scholar]
- Lima, M.J.; Muir, K.R.; Docherty, H.M.; McGowan, N.W.; Forbes, S.; Heremans, Y.; Heimberg, H.; Casey, J.; Docherty, K. Generation of Functional Beta-Like Cells from Human Exocrine Pancreas. PLoS ONE 2016, 11, e0156204. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, T.A.; Kawashima, S.; Miyatsuka, T.; Sasaki, S.; Shimo, N.; Katakami, N.; Kawamori, D.; Takebe, S.; Herrera, P.L.; Kaneto, H.; et al. Mafa Enables Pdx1 to Effectively Convert Pancreatic Islet Progenitors and Committed Islet α-Cells Into β-Cells In Vivo. Diabetes 2017, 66, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, M.; Kechele, D.O.; Broda, T.; Zhang, X.; Enriquez, J.R.; McCauley, H.A.; Sanchez, J.G.; McCracken, K.; Palermo, J.; Bernieh, A.; et al. Using Human Induced Pluripotent Stem Cell-Derived Organoids to Identify New Pathologies in Patients With PDX1 Mutations. Gastroenterology 2022, 163, 1053–1063.e1057. [Google Scholar] [CrossRef]
- Tu, P.; Huang, B.; Li, M.; Zhang, Y.; Bao, S.; Tu, N.; Yang, Y.; Lu, J. Exendin-4 may improve type 2 diabetes by modulating the epigenetic modifications of pancreatic histone H3 in STZ-induced diabetic C57BL/6 J mice. J. Physiol. Biochem. 2022, 78, 51–59. [Google Scholar] [CrossRef]
- Kim, M.K.; Cheong, Y.H.; Lee, S.H.; Kim, T.H.; Jung, I.H.; Chae, Y.; Lee, J.H.; Yang, E.K.; Park, H.; Yang, J.S.; et al. A novel GPR119 agonist DA-1241 preserves pancreatic function via the suppression of ER stress and increased PDX1 expression. Biomed. Pharmacother. 2021, 144, 112324. [Google Scholar] [CrossRef]
- Kornelius, E.; Li, H.H.; Peng, C.H.; Yang, Y.S.; Chen, W.J.; Chang, Y.Z.; Bai, Y.C.; Liu, S.; Huang, C.N.; Lin, C.L. Liraglutide protects against glucolipotoxicity-induced RIN-m5F β-cell apoptosis through restoration of PDX1 expression. J. Cell Mol. Med. 2019, 23, 619–629. [Google Scholar] [CrossRef]
- Zhou, D.; Chen, L.; Mou, X. Acarbose ameliorates spontaneous type-2 diabetes in db/db mice by inhibiting PDX-1 methylation. Mol. Med. Rep. 2021, 23, 72. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Li, K.; Liang, C.; Zhou, Z.; Wang, J.; Wang, S.; Liu, L.; Yu, C.L.; Song, Z.B.; Bao, Y.L.; et al. Tectorigenin enhances PDX1 expression and protects pancreatic β-cells by activating ERK and reducing ER stress. J. Biol. Chem. 2020, 295, 12975–12992. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Raya, L.; Juárez, J.; Pérez, J.; González, I. Effect of Silymarin in Pdx-1 expression and the proliferation of pancreatic β-cells in a pancreatectomy model. Phytomedicine 2014, 21, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lee, J.S.; Sezirahiga, J.; Kwon, H.C.; Jang, D.S.; Kang, K.S. Bioactive Phytochemicals Isolated from Akebia quinata Enhances Glucose-Stimulated Insulin Secretion by Inducing PDX-1. Plants 2020, 9, 1087. [Google Scholar] [CrossRef]
- Subramanian, M.; Thotakura, B.; Chandra Sekaran, S.P.; Jyothi, A.K.; Sundaramurthi, I. Naringin (4′,5,7-Trihydroxyflavanone 7-Rhamnoglucoside) Attenuates β-Cell Dysfunction in Diabetic Rats through Upregulation of PDX-1. Cells Tissues Organs 2018, 206, 133–143. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, F.; Tian, W.; Lai, J.; Qian, L.; Hong, W.; Chen, H.; Li, L.C. Andrographolide promotes pancreatic duct cells differentiation into insulin-producing cells by targeting PDX-1. Biochem. Pharmacol. 2020, 174, 113785. [Google Scholar] [CrossRef]
- Duan, J.; Zhang, F.; Lu, M.; Deng, W.; Zhai, Y.; Zhao, Y.; He, L.; Bai, Z.; Wang, Y.; Zhang, C. Swietenine and swietenolide from Swietenia macrophylla king improve insulin secretion and attenuate apoptosis in H2O2 induced INS-1 cells. Environ. Toxicol. 2022, 37, 2780–2792. [Google Scholar] [CrossRef]
- Fang, H.; Ding, Y.; Xia, S.; Chen, Q.; Niu, B. Loureirin B promotes insulin secretion through GLP-1R and AKT/PDX1 pathways. Eur. J. Pharmacol. 2022, 936, 175377. [Google Scholar] [CrossRef]
- Mansourzadeh, S.; Esmaeili, F.; Shabani, L.; Gharibi, S. Trans-differentiation of mouse mesenchymal stem cells into pancreatic β-like cells by a traditional anti-diabetic medicinal herb Medicago sativa L. J. Tradit. Complement. Med. 2022, 12, 466–476. [Google Scholar] [CrossRef]
- Soleimani-Dodran, M.; Alipanah-Moghadam, R.; Jeddi, F.; Babaei, M.; Salimnejad, R.; Bahreini, E. Effect of hydroalcoholic seed extract of Nigella sativa on hepatic and pancreatic factors of Nrf2 and FGF21 in the regulation of insulin transcription factors of MafA and PDX-1 in streptozotocin-treated diabetic rats. Nutr. Metab. 2022, 19, 64. [Google Scholar] [CrossRef]
- Yang, B.; Luo, Y.; Wei, X.; Kan, J. Polysaccharide from Hovenia dulcis (Guaizao) improves pancreatic injury and regulates liver glycometabolism to alleviate STZ-induced type 1 diabetes mellitus in rats. Int. J. Biol. Macromol. 2022, 214, 655–663. [Google Scholar] [CrossRef]
- Rad, M.G.; Sharifi, M.; Meamar, R.; Soltani, N. The role of pancreas to improve hyperglycemia in STZ-induced diabetic rats by thiamine disulfide. Nutr. Diabetes 2022, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Kannan, P.; Raghunathan, M.; Mohan, T.; Palanivelu, S.; Periandavan, K. Gymnemic Acid Ameliorates Pancreatic β-Cell Dysfunction by Modulating Pdx1 Expression: A Possible Strategy for β-Cell Regeneration. Tissue Eng. Regen. Med. 2022, 19, 603–616. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, A.; Gong, Y.; Sun, W.; Yan, B.; Lei, S.; Yao, L.-H. Improving effect of cordycepin on insulin synthesis and secretion in normal and oxidative-damaged INS-1 cells. Eur. J. Pharmacol. 2022, 920, 174843. [Google Scholar] [CrossRef]
- El-Huneidi, W.; Anjum, S.; Saleh, M.A.; Bustanji, Y.; Abu-Gharbieh, E.; Taneera, J. Carnosic Acid Protects INS-1 β-Cells against Streptozotocin-Induced Damage by Inhibiting Apoptosis and Improving Insulin Secretion and Glucose Uptake. Molecules 2022, 27, 2102. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, E.M.; Moawed, F.S.M.; Abdel-Hamid, G.R. Icariin Promote Stem Cells Regeneration and Repair Acinar Cells in L-arginine / Radiation -Inducing Chronic Pancreatitis in Rats. Dose-Response A Publ. Int. Hormesis Soc. 2020, 18, 1559325820970810. [Google Scholar] [CrossRef] [PubMed]
- Hanchang, W.; Khamchan, A.; Wongmanee, N.; Seedadee, C. Hesperidin ameliorates pancreatic β-cell dysfunction and apoptosis in streptozotocin-induced diabetic rat model. Life Sci. 2019, 235, 116858. [Google Scholar] [CrossRef] [PubMed]
- Golestaneh, E.; Aslani, A.; Aghaei, M.; Hashemnia, M.; Aarabi, M.H. Preparation and characterisation of a new form of silymarin as a potential antidiabetic agent in the adult male rat. Arch. Physiol. Biochem. 2021, 1–11. [Google Scholar] [CrossRef]
- Bahrami, G.; Sajadimajd, S.; Mohammadi, B.; Hatami, R.; Miraghaee, S.; Keshavarzi, S.; Khazaei, M.; Madani, S.H. Anti-diabetic effect of a novel oligosaccharide isolated from Rosa canina via modulation of DNA methylation in Streptozotocin-diabetic rats. Daru J. Fac. Pharm. Tehran Univ. Med. Sci. 2020, 28, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhen, W.; Maechler, P.; Liu, D. Small molecule kaempferol modulates PDX-1 protein expression and subsequently promotes pancreatic β-cell survival and function via CREB. J. Nutr. Biochem. 2013, 24, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Sinha, R.-A.; Singh, B.-K.; Li, G.-D.; Han, W.; Yen, P.-M. Resveratrol induces insulin gene expression in mouse pancreatic α-cells. Cell. Biosci. 2013, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-N.; Liu, Q.; Lei, L.; Sun, S.-J.; Li, C.-N.; Huan, Y.; Hou, S.-C.; Shen, Z.-F. The Chinese patent medicine, Jin-tang-ning, ameliorates hyperglycemia through improving β cell function in pre-diabetic KKAy mice. Chin. J. Nat. Med. 2020, 18, 827–836. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upstream | Downstream | |
|---|---|---|
| 1 | glucose | insulin |
| 2 | lipid | GCK |
| 3 | GLP-1 | GLUT-2 |
| 4 | ROS | IAPP |
| 5 | HNF-3β | somatostatin |
| 6 | HNF-6 | Ngn3 |
| 7 | TGF-β | Insm2 |
| 8 | PPAR-γ | synaptophysin |
| 9 | HNF-1α | Pax4 |
| 10 | Sp1 | MafA |
| 11 | HMGA1 | Nkx6.1 |
| 12 | FoxO1 | |
| 13 | Foxa1 | |
| 14 | Foxa2 | |
| 15 | PKA | |
| 16 | c-JUN | |
| 17 | JNK |
| Number | Name | Experimental Model | Administration Dose | Drug Effect | Reference |
|---|---|---|---|---|---|
| 1 | Swietenine (Stn) and swietenolide (Std) | INS-1 cells (Procell CL-0368) | 2 μM, 5 μM, 8 μM, 10 μM, 15 μM, 20 μM, 30 μM, 40 μM, 50 μM | It up-regulates the expression of PDX-1 protein, improves the insulin secretion function, protects oxidative stress injury, and reduces apoptosis. | [150] |
| 2 | Loureirin B | 3-week-old male C57BL/6J mice (14–15 g) | 45 mg/kg i.g. | It activates the AKT/PDX-1 signaling pathway. | [151] |
| 3 | Medicago sativa L. | Bone marrow mesenchymal stem cells (MSCs) | 50 μg/mL | It has the potential of differentiation induction of MSCs into IPCs with the characteristics of pancreatic β-like cells. | [152] |
| 4 | Nigella sativa seed | Male diabetic Wistar rats | 200 mg/kg, 400 mg/kg p.o. | It reduces oxidative stress and tissue damage, modifies the expression levels of PDX-1 and MafA genes, and regulates insulin secretion and blood glucose levels. | [153] |
| 5 | HDPs-2A (a polysaccharide purified from Hovenia dulcis) | 7-week-old male Sprague Dawley (SD) rats (170 ± 10 g) | 300 mg/kg, 200 mg/kg, 100 mg/kg, p.o. | It up-regulates PDX-1, activates and up-regulates IRS2 expression, and regulates apoptosis and regeneration of islet β cells to recover islet β-cell function injury in TIDM rats. | [154] |
| 6 | Thiamine disulfide (TD) | 4-week-old male Wistar rats (180–250 g) | 40 mg/kg i.p. | It increases serum insulin levels, IIR, and expression of PDX-1 and GLUT-2 genes. | [155] |
| 7 | Gymnemic acid (GA) | 2-month-old male albino Wistar rats (130–150 g) | 150 mg/kg p.o. | It ameliorates pancreatic β-cell dysfunction by modulating PDX-1 expression. | [156] |
| 8 | Cordycepin | INS-1 cells | 0.5~20 μM | It upregulates the mRNA level and protein expression of insulin, PDX-1, and GLUT-1. | [157] |
| 9 | Carnosic acid (CA) | INS-1 cells | 2.5 μM, 5 μM, 10 μM | It can protect β-cells through the PI3K/AKT/PDX-1/insulin pathway and mitochondria-mediated apoptosis. | [158] |
| 10 | Icariin | 6-week-old male albino rats (170–200 g) | 100 mg/kg p.o. | Icariin, and/or MSCs promoted the regeneration of pancreatic tissues by releasing PDX-1 and MafA involved in the recruitment of stem/progenitor cells in the tissue. | [159] |
| 11 | Hesperidin | Male Sprague Dawley rats | 100 mg·kg−1 p.o. | It enhances β-cell proliferation and repair and raises serum insulin levels. | [160] |
| 12 | A new form of silymarin solution (NFSM) | Male Wistar rats (220–250 g) | 100 mg/kg p.o. | It increases the expression of PDX-1 and insulin genes. | [161] |
| 13 | Oligosaccharide fraction isolated from Rosa canina | 8-week-old male Wistar rats (200–250 g) | 10, 20 and 30 mg/kg i.g. | It increases the expression of PDX-1 and may contribute to the modulation of DNA methylation. | [162] |
| 14 | Andrographolide named C1037 | 8-week-old male Kunming mice (18–22 g) | 50 mg/kg i.g. | It promotes pancreatic duct cell differentiation into insulin-producing cells by targeting PDX-1. | [149] |
| 15 | Tectorigenin | INS-1 cells;Diet-induced obese C57BL/6J mice | 40 μg/mL; 10, 20, 40 mg/kg i.p. | It enhances PDX-1 expression and protects pancreatic β-cells by activating ERK and reducing ER stress. | [145] |
| 16 | Stigmasterol-3-O-β-d-glucoside | INS-1 cells | 5 μM,10 μM | It enhances the PI3K-dependent phosphorylation of Akt at Ser473. The PI3K-dependent phosphorylation of Akt induces the movement of PDX-1 from the nucleus to the cytoplasm and regulates the proliferation of pancreatic β-cells. | [147] |
| 17 | Naringin (4′,5,7-Trihydroxyflavanone 7-Rhamnoglucoside) | Male adult Wistar rats (250–300 g) | 100 mg/kg p.o. | It increases insulin gene expression and insulin secretion by upregulating the PDX-1 gene and protein expression. | [148] |
| 18 | Small molecule kaempferol | INS-1E cells | 0.1 μM, 1 μM, 10 μM | It protects islet cells through PDX-1/cAMP/PKA/CREB signaling pathway. | [163] |
| 19 | Resveratrol | αTC9 cells | 25 μM | It inhibits histone deacetylase and promotes insulin expression synthesis by increasing PDX-1 expression levels. | [164] |
| 20 | Jin-tang-ning (JTN) | 8-week-old female KKAy mice and gender-matched C57BL/6J mice | 8 g JTN powder/kg | It upregulates expression levels of GCK and PDX-1. | [165] |
| 21 | DA-1241 | 7-week-old male ICR mice and Sprague-Dawley (SD) rats | 100 mg/kg i.p. | It can preserve pancreatic functions by suppressing ER stress and increasing PDX-1 expression. | [142] |
| 22 | Acarbose | 8-week-old db/db mice and male -+/db mice | 9 g/kg | It prevents the nuclear export of PDX-1 and blocks the increase in methylated PDX-1 in T2DM mouse β-cells. | [144] |
| 23 | Liraglutide | Rat RINm5F β-cell | 0.1 μmol/L | It can restore the expression of PDX-1 and upregulate mitophagy to restore mitochondrial function and ameliorate β-cell impairment. | [143] |
| 24 | Exendin-4 | 3-week-old C57BL/6J mice | 10 μg/kg | It can improve T2DM progression by reversing global pancreatic histone H3K9 and H3K23 acetylation, H3K4 mono-methylation, and H3K9 di-methylation and also reverse the inhibitory state of PDX-1. | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Fang, X.; Wei, J.; Miao, R.; Wu, H.; Ma, K.; Tian, J. PDX-1: A Promising Therapeutic Target to Reverse Diabetes. Biomolecules 2022, 12, 1785. https://doi.org/10.3390/biom12121785
Zhang Y, Fang X, Wei J, Miao R, Wu H, Ma K, Tian J. PDX-1: A Promising Therapeutic Target to Reverse Diabetes. Biomolecules. 2022; 12(12):1785. https://doi.org/10.3390/biom12121785
Chicago/Turabian StyleZhang, Yanjiao, Xinyi Fang, Jiahua Wei, Runyu Miao, Haoran Wu, Kaile Ma, and Jiaxing Tian. 2022. "PDX-1: A Promising Therapeutic Target to Reverse Diabetes" Biomolecules 12, no. 12: 1785. https://doi.org/10.3390/biom12121785