
FGF9 Promotes Expression of HAS2 in Palatal Elevation via the Wnt/β-Catenin/TCF7L2 Pathway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Transgenic Mice
2.2. Micro-CT Analysis, Hematoxylin and Eosin (HE) Staining and Masson’s Trichrome (MT) Staining
2.3. Immunohistochemistry (IHC)
2.4. Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′-3′) |
|---|---|
| Fgf9 | F:GACAATAATTTCCACCTCC F:CACTGGGCTCTAACTCTTC R:ATTTGCTATGCACGGACAC |
| Has1 | F:CCACTAGTGCAATCACAGAAAG R:GTGTGGCTTTGTGTCTCAATAG |
| Has2 | F:TGTGAGAGGTTTCTATGTGTCCT R:ACCGTACAGTCCAAATGAGAAGT |
| Has3 | F:CCGATCTCATTTCTAAGCAAGC R:CCTCTCCTATTTGTAGCTCTGG |
| CTNNB1 | F:CCATGATTCCTTCATATTTGC R:GTAATACGGTTATCCACGCG |
| TCF7L2 | F:CCCCTGACTTGAACCCACCC R:CCCTCGTCGTCGGATTTGAT |
| GAPDH | F:GGAGCGAGATCCCTCCAAAAT R:GGCTGTTGTCATACTTCTCATGG |
2.5. Western Blotting
2.6. Quantification of Hyaluronic Acid
2.7. GeneMANIA and Jaspar Database
2.8. Isolation, Culture, and Treatment of Mouse Embryonic Palatal Mesenchymal(MEPM) Cells
2.9. Statistical Analysis
3. Results
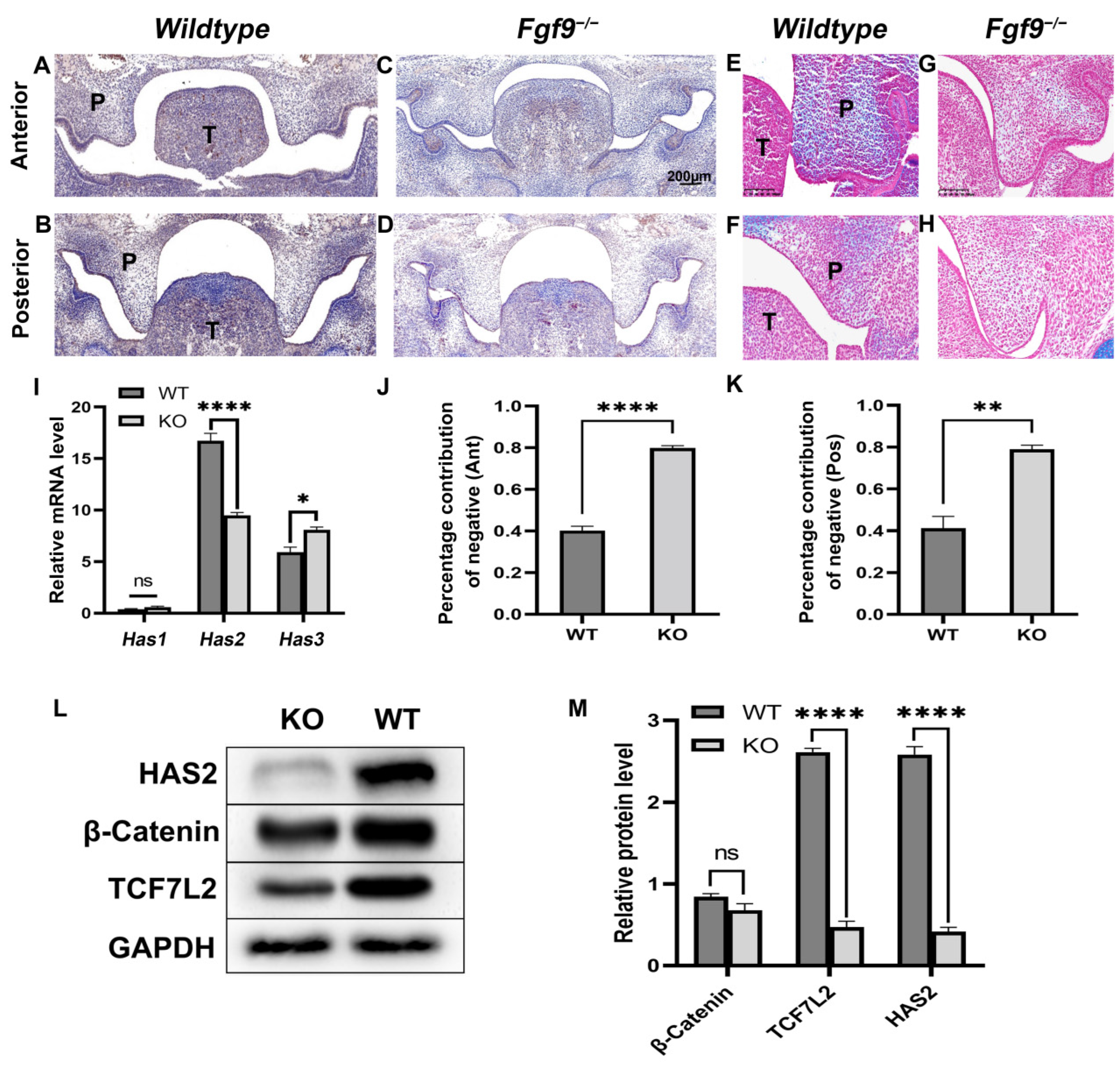
3.1. Fgf9−/− Embryos Exhibited Cleft Palate
3.2. Abnormal HA Accumulation and HAS2 Expression in the Fgf9−/− Embryos
3.3. FGF9 Promotes HAS2 and Tcf7L2 Expression in Palate
Fgf9 Promotes HAS2 Expression via the Tcf7L2-Activated Wnt Pathway In Vitro
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burg, M.L.; Chai, Y.; Yao, C.A.; Magee, W., III; Figueiredo, J.C. Epidemiology, etiology, and treatment of isolated cleft palate. Front. Physiol. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, B.M.; Murray, J.C. Sequence evaluation of FGF and FGFR gene conserved non-coding elements in non-syndromic cleft lip and palate cases. Am. J. Med. Genet. A 2007, 143A, 3228–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, D.; Zimmerman, N.; Bedford, M.; Avivi, A.; Yayon, A. Identification of fibroblast growth factor 9 (FGF9) as a high affinity, heparin dependent ligand for FGF receptors 3 and 2 but not for FGF receptors 1 and 4. Growth Factors 1995, 12, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Murakami, H.; Okawa, A.; Okimoto, N.; Hiraoka, S.; Nakahara, T.; Akasaka, R.; Shiraishi, Y.I.; Futatsugi, N.; Mizutani-Koseki, Y.; et al. FGF9 monomer–dimer equilibrium regulates extracellular matrix affiffiffinity and tissue diffffusion. Nat. Genet. 2009, 41, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, J.S.; Feldman, B.; Nadeau, J.H.; Goldfarb, M.; Ornitz, D.M. Genomic organization and embryonic expression of the mouse fibroblast growth factor 9 gene. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1999, 216, 72–88. [Google Scholar] [CrossRef]
- Hammond, N.L.; Brookes, K.J.; Dixon, M.J. Ectopic Hedgehog Signaling Causes Cleft Palate and Defective Osteogenesis. J. Dent. Res. 2018, 97, 1485–1493. [Google Scholar] [CrossRef]
- Colvin, J.S.; White, A.C.; Pratt, S.J.; Ornitz, D.M. Lung hypoplasia and neonatal death in Fgf9–null mice identify this gene as an essential regulator of lung mesenchyme. Development 2001, 128, 2095–2106. [Google Scholar] [CrossRef]
- Li, R.; Sun, Y.; Chen, Z.; Zheng, M.; Shan, Y.; Ying, X.; Weng, M.; Chen, Z. The Fibroblast Growth Factor 9 (Fgf9) Participates in Palatogenesis by Promoting Palatal Growth and Elevation. Front. Physiol. 2021, 12, 653040. [Google Scholar] [CrossRef]
- Li, C.; Lan, Y.; Jiang, R. Molecular and Cellular Mechanisms of Palate Development. J. Dent. Res. 2017, 96, 1184–1191. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X.; Wang, Y.; Wang, Y.; Zhang, J.; Shi, B.; Li, C. Three-dimensional reconstruction of systematic histological sections: Application to observations on palatal shelf elevation. Int. J. Oral Sci. 2021, 13, 17. [Google Scholar] [CrossRef]
- Yonemitsu, M.A.; Lin, T.Y.; Yu, K. Hyaluronic acid is required for palatal shelf movement and its interaction with the tongue during palatal shelf elevation. Dev. Biol. 2020, 457, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Albeiroti, S.; Ayasoufi, K.; Hill, D.R.; Shen, B.; de la Motte, C.A. Platelet hyaluronidase-2: An enzyme that translocates to the surface upon activation to function in extracellular matrix degradation. Blood 2015, 125, 1460–1469. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, C.; Zhu, Z.; Yuan, L.; Chan, W.Y.; Sha, O. Extracellular Matrix Remodeling During Palate Development. Organogenesis 2020, 16, 43–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway, J.L.; Jones, S.J.; Mossey, P.A.; Ellis, I.R. The control and importance of hyaluronan synthase expression in palatogenesis. Front. Physiol. 2013, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, S.M.; Ruest, L.B.; Benson, M.D.; Svoboda, K.K.H. Extracellular Matrix in Secondary Palate Development. Anat. Rec. 2020, 303, 1543–1556. [Google Scholar] [CrossRef]
- Paiva, K.B.S.; Maas, C.S.; Dos Santos, P.M.; Granjeiro, J.M.; Letra, A. Extracellular Matrix Composition and Remodeling: Current Perspectives on Secondary Palate Formation, Cleft Lip/Palate, and Palatal Reconstruction. Front. Cell Dev. Biol. 2019, 7, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Qin, C.; Jiang, R. Requirement of Hyaluronan Synthase-2 in Craniofacial and Palate Development. J. Dent. Res. 2019, 98, 1367–1375. [Google Scholar] [CrossRef]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and Function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef]
- Fouladi-Nashta, A.A.; Raheem, K.A.; Marei, W.F.; Ghafari, F.; Hartshorne, G.M. Regulation and roles of the hyaluronan system in mammalian reproduction. Reproduction 2017, 153, R43–R58. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, L.; Ouyang, X.; He, P. Transcription Factor-7-Like-2 (TCF7L2) in Atherosclerosis: A Potential Biomarker and Therapeutic Target. Front. Cardiovasc. Med. 2021, 8, 701279. [Google Scholar] [CrossRef]
- Zhang, X.; Weng, M.; Chen, Z. Fibroblast Growth Factor 9 (FGF9) negatively regulates the early stage of chondrogenic differentiation. PLoS ONE 2021, 16, e0241281. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Chen, Z.; Xiao, Q.; Li, R.; Chen, Z. A review of FGF signaling in palate development. Biomed. Pharmacother. 2018, 103, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Greene, R.M.; Pisano, M.M. Palate morphogenesis: Current understanding and future directions. Birth Defects Res. 2010, 90, 133–154. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Zhang, N.; Liu, H.; Xu, J.; Jiang, R. Golgb1 regulates protein glycosylation and is crucial for mammalian palate development. Development 2016, 143, 2344–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, A.S.; Goudy, S.L. Cellular and molecular mechanisms of cleft palate development. Laryngoscope Investig. Otolaryngol. 2018, 4, 160–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolk, A.; Hatipoglu, D.; Cutler, A.; Ali, M.; Bell, L.; Qi, J.H.; Singh, R.; Batoki, J.; Karle, L.; Bonilha, V.L.; et al. Role of FGF and hyaluronan in choroidal neovascularization in sorsby fundus dystrophy. Cells 2020, 9, 608. [Google Scholar] [CrossRef] [Green Version]
- Virakul, S.; Heutz, J.W.; Dalm, V.A.; Peeters, R.P.; Paridaens, D.; van den Bosch, W.A.; Hirankarn, N.; van Hagen, P.M.; Dik, W.A. Basic FGF and PDGF–BB synergistically stimulate hyaluronan and IL–6 production by orbital fifibroblasts. Mol. Cell Endocrinol. 2016, 433, 94–104. [Google Scholar] [CrossRef]
- McCarthy, N.; Sidik, A.; Bertrand, J.Y.; Eberhart, J.K. An Fgf–Shh signaling hierarchy regulates early specifification of the zebrafifish skull. Dev. Biol. 2016, 415, 261–277. [Google Scholar] [CrossRef]
- Ferguson, M.W. The mechanism of palatal shelf elevation and the pathogenesis of cleft palate. Virchows Archiv A Pathol. Anat. Histol. 1977, 375, 97–113. [Google Scholar] [CrossRef]
- Hung, I.H.; Schoenwolf, G.C.; Lewandoski, M.; Ornitz, D.M. A combined series of Fgf9 and Fgf18 mutant alleles identififies unique and redundant roles in skeletal development. Dev. Biol. 2016, 411, 72–84. [Google Scholar] [CrossRef]
- Jacobson, A.; Brinck, J.; Briskin, M.J.; Spicer, A.P.; Heldin, P. Expression of human hyaluronan synthases in response to external stimuli. Biochem. J. 2000, 348, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Sawai, T.; Yoshida, M.; Lenas, P.; Yamada, Y.; Imagawa, M.; Shinomura, T.; Hamaguchi, M.; Yoshida, Y.; Ohnuki, Y.; et al. Three isoforms of mammalian hyaluronan synthases have distinct enzymatic properties. J. Biol. Chem. 1999, 274, 25085–25092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.R.; Han, X.H.; Nishimori, K.; Ben-Avraham, D.; Oh, Y.J.; Shim, J.W.; Yoon, J.K. Canonical WNT/β-catenin Signaling Activated by WNT9b and RSPO2 Cooperation Regulates Facial Morphogenesis in Mice. Front. Cell Dev. Biol. 2020, 8, 264. [Google Scholar] [CrossRef]
- Reynolds, K.; Kumari, P.; Rincon, L.S.; Gu, R.; Ji, Y.; Kumar, S.; Zhou, C.J. Wnt signaling in orofacial clefts: Crosstalk, pathogenesis and models. Dis. Model. Mech. 2019, 12, dmm037051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvinen, E.; Shimomura-Kuroki, J.; Balic, A.; Jussila, M.; Thesleff, I. Mesenchymal Wnt/beta-catenin signaling limits tooth number. Development 2018, 145, dev158048. [Google Scholar] [CrossRef] [Green Version]
- Eubelen, M.; Bostaille, N.; Cabochette, P.; Gauquier, A.; Tebabi, P.; Dumitru, A.C.; Koehler, M.; Gut, P.; Alsteens, D.; Stainier, D.Y.R.; et al. A molecular mechanism for Wnt ligand-specific signaling. Science 2018, 361, eaat1178. [Google Scholar] [CrossRef] [Green Version]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–11205. [Google Scholar] [CrossRef] [Green Version]
- Voloshanenko, O.; Schwartz, U.; Kranz, D.; Rauscher, B.; Linnebacher, M.; Augustin, I.; Boutros, M. β-catenin-independent regulation of Wnt target genes by RoR2 and ATF2/ATF4 in colon cancer cells. Sci. Rep. 2018, 8, 3178. [Google Scholar] [CrossRef] [Green Version]
- Sada, R.; Kimura, H.; Fukata, Y.; Fukata, M.; Yamamoto, H.; Kikuchi, A. Dynamic palmitoylation controls the microdomain localization of the DKK1 receptors CKAP4 and LRP6. Sci. Signal. 2019, 12, eaat9519. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sakane, H.; Yamamoto, H.; Michiue, T.; Kikuchi, A. Wnt3a and Dkk1 regulate distinct internalization pathways of LRP6 to tune the activation of beta-catenin signaling. Dev. Cell 2008, 15, 37–48. [Google Scholar] [CrossRef]
- Tortelote, G.G.; Reis, R.R.; de Almeida Mendes, F.; Abreu, J.G. Complexity of the Wnt/β-catenin pathway: Searching for an activation model. Cell. Signal. 2017, 40, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Doumpas, N.; Lampart, F.; Robinson, M.D.; Lentini, A.; Nestor, C.E.; Cantù, C.; Basler, K. TCF/LEF dependent and independent transcriptional regulation of Wnt/β-catenin target genes. EMBO J. 2019, 38, e98873. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.H.; Li, Y.L.; Liu, N.J.; Yang, Z.; Tao, X.M.; Du, Y.P.; Wang, X.C.; Lu, B.; Zhang, Z.Y.; Hu, R.M.; et al. TCF7L2 regulates pancreatic β-cell function through PI3K/AKT signal pathway. Diabetol. Metab. Syndr. 2019, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Brinkmeier, M.L.; Potok, M.A.; Cha, K.B.; Gridley, T.; Stifani, S.; Meeldijk, J.; Clevers, H.; Camper, S.A. TCF and Groucho-related genes influence pituitary growth and development. Mol. Endocrinol. 2003, 17, 2152–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Rooker, S.M.; Helms, J.A. Molecular control of facial morphology. Semin. Cell Dev. Biol. 2010, 21, 309–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Wang, W.; Wu, X.; Shi, J. Pizotifen inhibits the proliferation and invasion of gastric cancer cells. Exp. Med. 2020, 19, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Zhong, K.; Xiao, J.; Huang, Y.; Wei, Y.; Tang, L.; Chen, S.; Wu, J.; Ma, J.; Cao, Z.; et al. Exposure to pyrimethanil induces developmental toxicity and cardiotoxicity in zebrafish. Chemosphere 2020, 255, 126889. [Google Scholar] [CrossRef]
- Liu, J.; Wu, X.; Mitchell, B.; Kintner, C.; Ding, S.; Schultz, P.G. A small-molecule agonist of the Wnt signaling pathway. Angew. Chem. Int. Ed. Engl. 2005, 44, 1987–1990. [Google Scholar] [CrossRef]




| Name | Score | Relative Score | Sequence ID | Start | End | Strand | Predicted Sequence |
|---|---|---|---|---|---|---|---|
| TCF7L2 | 13.432302 | 0.92807441 | NC_000008.11 | 1370 | 1383 | − | ACTGATCAAAGGGC |
| TCF7L2 | 9.703565 | 0.87654208 | NC_000008.11 | 719 | 732 | + | CGTGATGAAAGCAT |
| TCF7L2 | 9.428534 | 0.87274107 | NC_000008.11 | 448 | 461 | + | GGAGTTCAAACAAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Ying, X.; Li, R.; Weng, M.; Shi, J.; Chen, Z. FGF9 Promotes Expression of HAS2 in Palatal Elevation via the Wnt/β-Catenin/TCF7L2 Pathway. Biomolecules 2022, 12, 1639. https://doi.org/10.3390/biom12111639
Sun Y, Ying X, Li R, Weng M, Shi J, Chen Z. FGF9 Promotes Expression of HAS2 in Palatal Elevation via the Wnt/β-Catenin/TCF7L2 Pathway. Biomolecules. 2022; 12(11):1639. https://doi.org/10.3390/biom12111639
Chicago/Turabian StyleSun, Yidan, Xiyu Ying, Ruomei Li, Mengjia Weng, Jiajun Shi, and Zhenqi Chen. 2022. "FGF9 Promotes Expression of HAS2 in Palatal Elevation via the Wnt/β-Catenin/TCF7L2 Pathway" Biomolecules 12, no. 11: 1639. https://doi.org/10.3390/biom12111639