
2-Cell-like Cells: An Avenue for Improving SCNT Efficiency


Abstract
:1. Introduction
2. Characterization of 2CLCs
3. The Strategies Used for Inducing 2CLCs May Improve SCNT Efficiency
3.1. Effect of Regulating Histone-Modifying Enzymes on the Developmental Competence of Early Cloned Embryos
3.2. Effect of Regulating Methylation Levels on the Developmental Competence of Early Cloned Embryos
3.3. Effect of Double Homeobox (DUX, i.e., Pioneer Transcription Factors) on the Developmental Competence of Early Cloned Embryos
3.4. Effect of Regulating Reorganization of 3D Chromatin Structure on the Developmental Competence of Early Cloned Embryos
3.5. Effect of Regulating Chromatin Assembly on the Developmental Competence of Early Cloned Embryos
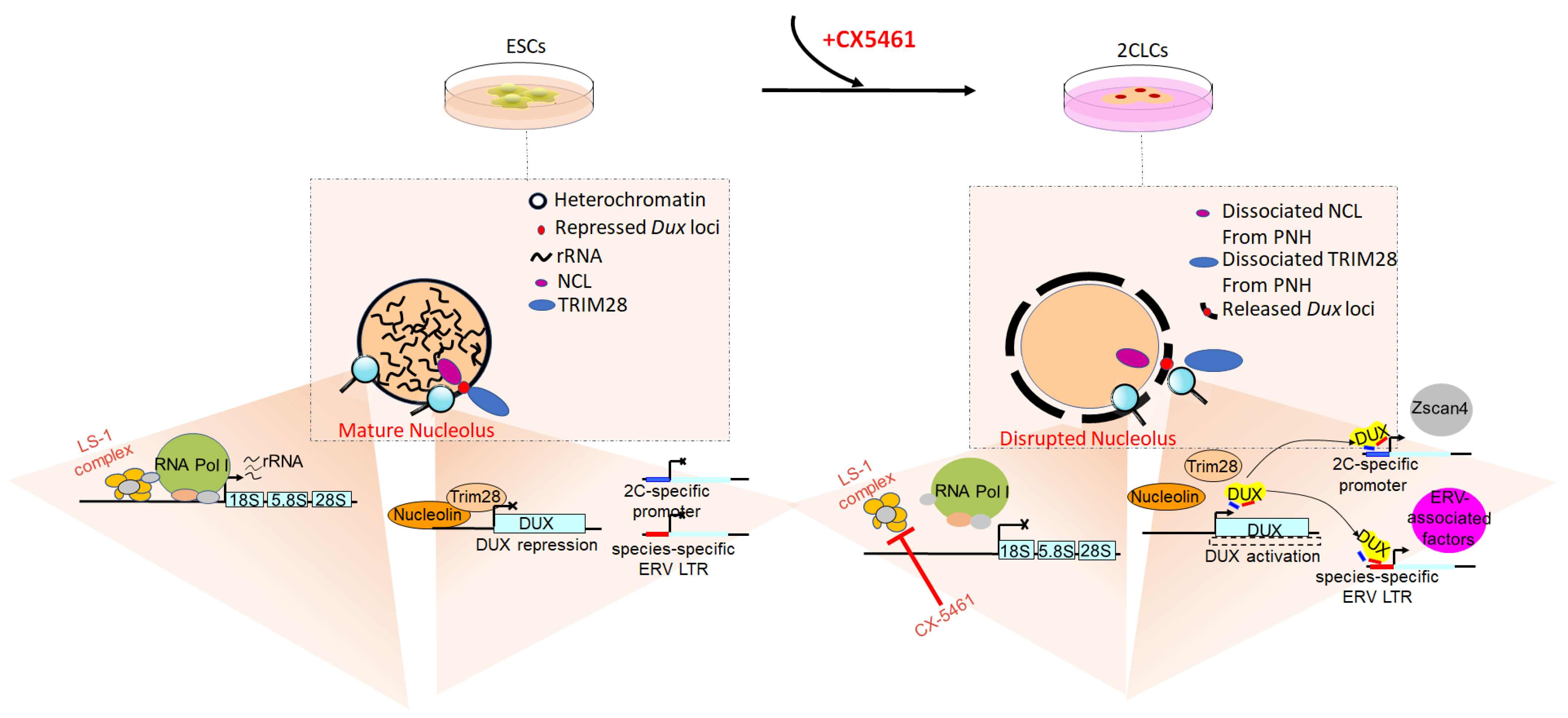
3.6. Effect of Regulating rRNA Synthesis and Nucleolar Maturation on the Developmental Competence of Early Cloned Embryos
3.7. Effect of Regulating Replication Features on the Developmental Competence of Early Cloned Embryos
3.8. Effect of Regulating Energy Substrates on the Developmental Competence of Early Cloned Embryos
3.9. Other Strategies That Have Not Been Applied to Improve SCNT Efficiency
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Macfarlan, T.S.; Gifford, W.D.; Agarwal, S.; Driscoll, S.; Lettieri, K.; Wang, J.; Andrews, S.E.; Franco, L.; Rosenfeld, M.G.; Ren, B.; et al. Endogenous retroviruses and neighboring genes are coordinately repressed by LSD1/KDM1A. Genes Dev. 2011, 25, 594–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlan, T.S.; Gifford, W.D.; Driscoll, S.; Lettieri, K.; Rowe, H.M.; Bonanomi, D.; Firth, A.; Singer, O.; Trono, D.; Pfaff, S.L. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 2012, 487, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peaston, A.E.; Evsikov, A.V.; Graber, J.H.; de Vries, W.N.; Holbrook, A.E.; Solter, D.; Knowles, B.B. Retrotransposons regulate host genes in mouse oocytes and preimplantation embryos. Dev. Cell 2004, 7, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, P.; Stein, P.; Anger, M.; Bernstein, E.; Hannon, G.J.; Schultz, R.M. RNAi and expression of retrotransposons MuERV-L and IAP in preimplantation mouse embryos. Dev. Biol. 2004, 269, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Tarkowski, A.K. Experiments on the Development of Isolated Blastomeres of Mouse Eggs. Nature 1959, 184, 1286–1287. [Google Scholar] [CrossRef]
- Togashi, M.; Suzuki, H.; Miyai, T.; Okamoto, M.T. Production of monozygotic twins by splitting of 2-cell stage embryos in mice. Jpn. J. Anim. Reprod. 1987, 33, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Sotomaru, Y.; Kato, Y.; Tsunoda, Y. Production of monozygotic twins after freezing and thawing of bisected mouse embryos. Cryobiology 1998, 37, 139–145. [Google Scholar] [CrossRef]
- Moore, N.W.; Adams, C.E.; Rowson, L.E. Developmental potential of single blastomeres of the rabbit egg. J. Reprod. Fertil. 1968, 17, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.H.; Loskutoff, N.M.; Plante, Y.; Betteridge, K.J. Production of four identical calves by the separation of blastomeres from an in vitro derived four-cell embryo. Vet. Rec. 1995, 137, 15–16. [Google Scholar] [CrossRef]
- Mitalipov, S.M.; Yeoman, R.R.; Kuo, H.C.; Wolf, D.P. Monozygotic twinning in rhesus monkeys by manipulation of in vitro-derived embryos. Biol. Reprod. 2002, 66, 1449–1455. [Google Scholar] [CrossRef]
- Beddington, R.S.; Robertson, E.J. An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo. Development 1989, 105, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, C.; Huyser, C.; Egli, D.; Pepper, M.S. Lessons Learned from Somatic Cell Nuclear Transfer. Int. J. Mol. Sci. 2020, 21, 2314. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Minami, N.; Kono, T.; Imai, H. Zygotically activated genes are suppressed in mouse nuclear transferred embryos. Cloning Stem Cells 2006, 8, 295–304. [Google Scholar] [CrossRef]
- Matoba, S.; Liu, Y.; Lu, F.; Iwabuchi, K.A.; Shen, L.; Inoue, A.; Zhang, Y. Embryonic development following somatic cell nuclear transfer impeded by persisting histone methylation. Cell 2014, 159, 884–895. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Liu, X.; Wang, C.; Gao, Y.; Gao, R.; Kou, X.; Zhao, Y.; Li, J.; Wu, Y.; Xiu, W.; et al. Identification of key factors conquering developmental arrest of somatic cell cloned embryos by combining embryo biopsy and single-cell sequencing. Cell Discov. 2016, 2, 16010. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Song, L.; Liu, X.; Bai, L.; Li, G. KDM6A and KDM6B play contrasting roles in nuclear transfer embryos revealed by MERVL reporter system. EMBO Rep. 2018, 19, e46240. [Google Scholar] [CrossRef]
- Vasil, I.K.; Vasil, V. Totipotency and embryogenesis in plant cell and tissue cultures. In Vitro 1972, 8, 117–127. [Google Scholar] [CrossRef]
- Ishiuchi, T.; Torres-Padilla, M.E. Towards an understanding of the regulatory mechanisms of totipotency. Curr. Opin. Genet. Dev. 2013, 23, 512–518. [Google Scholar] [CrossRef]
- Edwards, R.G.; Beard, H.K. Oocyte polarity and cell determination in early mammalian embryos. Mol. Hum. Reprod. 1997, 3, 863–905. [Google Scholar] [CrossRef]
- Evsikov, A.V.; de Vries, W.N.; Peaston, A.E.; Radford, E.E.; Fancher, K.S.; Chen, F.H.; Blake, J.A.; Bult, C.J.; Latham, K.E.; Solter, D.; et al. Systems biology of the 2-cell mouse embryo. Cytogenet. Genome Res. 2004, 105, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Kigami, D.; Minami, N.; Takayama, H.; Imai, H. MuERV-L is one of the earliest transcribed genes in mouse one-cell embryos. Biol. Reprod. 2003, 68, 651–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooga, M.; Fulka, H.; Hashimoto, S.; Suzuki, M.G.; Aoki, F. Analysis of chromatin structure in mouse preimplantation embryos by fluorescent recovery after photobleaching. Epigenetics 2016, 11, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Bošković, A.; Eid, A.; Pontabry, J.; Ishiuchi, T.; Spiegelhalter, C.; Raghu Ram, E.V.; Meshorer, E.; Torres-Padilla, M.E. Higher chromatin mobility supports totipotency and precedes pluripotency in vivo. Genes Dev. 2014, 28, 1042–1047. [Google Scholar] [CrossRef] [Green Version]
- Ishiuchi, T.; Enriquez-Gasca, R.; Mizutani, E.; Bošković, A.; Ziegler-Birling, C.; Rodriguez-Terrones, D.; Wakayama, T.; Vaquerizas, J.M.; Torres-Padilla, M.E. Early embryonic-like cells are induced by downregulating replication-dependent chromatin assembly. Nat. Struct. Mol. Biol. 2015, 22, 662–671. [Google Scholar] [CrossRef]
- Xie, S.Q.; Leeke, B.J.; Whilding, C.; Wagner, R.T.; Garcia-Llagostera, F.; Low, Y.; Chammas, P.; Cheung, N.T.; Dormann, D.; McManus, M.T.; et al. Nucleolar-based Dux repression is essential for embryonic two-cell stage exit. Genes Dev. 2022, 36, 331–347. [Google Scholar] [CrossRef]
- Hendrickson, P.G.; Doráis, J.A.; Grow, E.J.; Whiddon, J.L.; Lim, J.W.; Wike, C.L.; Weaver, B.D.; Pflueger, C.; Emery, B.R.; Wilcox, A.L.; et al. Conserved roles of mouse DUX and human DUX4 in activating cleavage-stage genes and MERVL/HERVL retrotransposons. Nat. Genet. 2017, 49, 925–934. [Google Scholar] [CrossRef]
- Du, Z.; Zheng, H.; Huang, B.; Ma, R.; Wu, J.; Zhang, X.; He, J.; Xiang, Y.; Wang, Q.; Li, Y.; et al. Allelic reprogramming of 3D chromatin architecture during early mammalian development. Nature 2017, 547, 232–235. [Google Scholar] [CrossRef]
- Kai, K.; Diaz, N.; Enriquez-Gasca, R.; Gaume, X.; Vaquerizas, J.M. Transposable elements drive reorganisation of 3D chromatin during early embryogenesis. BioRxiv 2019. [Google Scholar] [CrossRef]
- Eckersley-Maslin, M.A.; Svensson, V.; Krueger, C.; Stubbs, T.M.; Giehr, P.; Krueger, F.; Miragaia, R.J.; Kyriakopoulos, C.; Berrens, R.V.; Milagre, I.; et al. MERVL/Zscan4 Network Activation Results in Transient Genome-wide DNA Demethylation of mESCs. Cell Rep. 2016, 17, 179–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messerschmidt, D.M.; Knowles, B.B.; Solter, D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev. 2014, 28, 812–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthmann, M.; Iturbide, A.; Torres-Padilla, M.E.; Rodriguez-Terrones, D.; Hartleben, G.; Gaume, X.; Eid, A. A distinct metabolic state arises during the emergence of 2-cell-like cells. EMBO Rep. 2020, 21, e48354. [Google Scholar] [CrossRef]
- Dumollard, R.; Carroll, J.; Duchen, M.R.; Campbell, K.; Swann, K. Mitochondrial function and redox state in mammalian embryos. Semin. Cell Dev. Biol. 2009, 20, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tan, D.E.K.; Chia, G.; Tan, H.; Leong, H.F.; Chen, B.J.; Lau, M.S.; Tan, K.Y.S.; Bi, X.; Yang, D.; et al. Maternal factor NELFA drives a 2C-like state in mouse embryonic stem cells. Nat. Cell Biol. 2020, 22, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.J. Metabolism of Preimplantation Embryo Development: A Bystander or an Active Participant? Curr. Top. Dev. Biol. 2016, 120, 259–310. [Google Scholar] [PubMed]
- Ma, J.; Svoboda, P.; Schultz, R.M.; Stein, P. Regulation of zygotic gene activation in the preimplantation mouse embryo: Global activation and repression of gene expression. Biol. Reprod. 2001, 64, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- Wiekowski, M.; Miranda, M.; Nothias, J.Y.; DePamphilis, M.L. Changes in histone synthesis and modification at the beginning of mouse development correlate with the establishment of chromatin mediated repression of transcription. J. Cell Sci. 1997, 110 (Pt 10), 1147–1158. [Google Scholar] [CrossRef]
- Mao, T.; Han, C.; Deng, R.; Wei, B.; Meng, P.; Luo, Y.; Zhang, Y. Treating donor cells with 2-PCPA corrects aberrant histone H3K4 dimethylation and improves cloned goat embryo development. Syst. Biol. Reprod. Med. 2018, 64, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, H.; Yao, J.; Qin, G.; Wang, F.; Wang, X.; Luo, A.; Zheng, Q.; Cao, C.; Zhao, J. BIX-01294 increases pig cloning efficiency by improving epigenetic reprogramming of somatic cell nuclei. Reproduction 2016, 151, 39–49. [Google Scholar] [CrossRef]
- Kishigami, S.; Mizutani, E.; Ohta, H.; Hikichi, T.; Thuan, N.V.; Wakayama, S.; Bui, H.T.; Wakayama, T. Significant improvement of mouse cloning technique by treatment with trichostatin A after somatic nuclear transfer. Biochem. Biophys. Res. Commun. 2006, 340, 183–189. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Villemoes, K.; Pedersen, A.M.; Purup, S.; Vajta, G. An epigenetic modifier results in improved in vitro blastocyst production after somatic cell nuclear transfer. Cloning Stem Cells 2007, 9, 357–363. [Google Scholar] [CrossRef]
- Fu, X.; Wu, X.; Djekidel, M.N.; Zhang, Y. Myc and Dnmt1 impede the pluripotent to totipotent state transition in embryonic stem cells. Nat. Cell Biol. 2019, 21, 835–844. [Google Scholar] [CrossRef]
- Dan, J.; Rousseau, P.; Hardikar, S.; Veland, N.; Wong, J.; Autexier, C.; Chen, T. Zscan4 Inhibits Maintenance DNA Methylation to Facilitate Telomere Elongation in Mouse Embryonic Stem Cells. Cell Rep. 2017, 20, 1936–1949. [Google Scholar] [CrossRef] [Green Version]
- Peat, J.R.; Reik, W. Incomplete methylation reprogramming in SCNT embryos. Nat. Genet. 2012, 44, 965–966. [Google Scholar] [CrossRef]
- Xu, W.H.; Li, Z.C.; Wu, Z.F.; Shi, J.S. Effect of DNMT1 on development, gene transcription and DNA methylation of porcine (Sus scrofa) cloned embryos. J. Agric. Biotechnol. 2018, 26, 2075–2083. [Google Scholar]
- Song, X.; Liu, Z.; He, H.; Wang, J.; Li, H.; Li, J.; Li, F.; Jiang, Z.; Huan, Y. Dnmt1s in donor cells is a barrier to SCNT-mediated DNA methylation reprogramming in pigs. Oncotarget 2017, 8, 34980–34991. [Google Scholar] [CrossRef] [Green Version]
- Whiddon, J.L.; Langford, A.T.; Wong, C.J.; Zhong, J.W.; Tapscott, S.J. Conservation and innovation in the DUX4-family gene network. Nat. Genet. 2017, 49, 935–940. [Google Scholar] [CrossRef] [Green Version]
- De Iaco, A.; Planet, E.; Coluccio, A.; Verp, S.; Duc, J.; Trono, D. DUX-family transcription factors regulate zygotic genome activation in placental mammals. Nat. Genet. 2017, 49, 941–945. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, L.; Liu, W.; Qiao, Z.; Shen, S.; Zhu, Q.; Gao, R.; Wang, M.; Wang, M.; Li, C.; et al. Dux-Mediated Corrections of Aberrant H3K9ac during 2-Cell Genome Activation Optimize Efficiency of Somatic Cell Nuclear Transfer. Cell Stem Cell 2021, 28, 150–163. [Google Scholar] [CrossRef]
- Yang, L.; Liu, X.; Song, L.; Di, A.; Su, G.; Bai, C.; Wei, Z.; Li, G. Transient Dux expression facilitates nuclear transfer and induced pluripotent stem cell reprogramming. EMBO Rep. 2020, 21, e50054. [Google Scholar] [CrossRef]
- Olbrich, T.; Vega-Sendino, M.; Tillo, D.; Wu, W.; Zolnerowich, N.; Pavani, R.; Tran, A.D.; Domingo, C.N.; Franco, M.; Markiewicz-Potoczny, M.; et al. CTCF is a barrier for 2C-like reprogramming. Nat. Commun. 2021, 12, 4856. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, Y.; Zhou, J.; Bi, Y.; Xu, J.; Xu, C.; Kou, X.; Zhao, Y.; Li, Y.; Tu, Z.; et al. Precise temporal regulation of Dux is important for embryo development. Cell Res. 2019, 29, 956–959. [Google Scholar] [CrossRef]
- Haarhuis, J.H.I.; van der Weide, R.H.; Blomen, V.A.; Yáñez-Cuna, J.O.; Amendola, M.; van Ruiten, M.S.; Krijger, P.H.L.; Teunissen, H.; Medema, R.H.; van Steensel, B.; et al. The Cohesin Release Factor WAPL Restricts Chromatin Loop Extension. Cell 2017, 169, 693–707. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, W.; Abdennur, N.; Goloborodko, A.; Pekowska, A.; Fudenberg, G.; Loe-Mie, Y.; Fonseca, N.A.; Huber, W.; Haering, C.H.; Mirny, L.; et al. Two independent modes of chromatin organization revealed by cohesin removal. Nature 2017, 551, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.D.P.; Feldmann, A.; Hernández-Rodríguez, B.; Díaz, N.; Brown, J.M.; Fursova, N.A.; Blackledge, N.P.; Prathapan, P.; Dobrinic, P.; Huseyin, M.K.; et al. Cohesin Disrupts Polycomb-Dependent Chromosome Interactions in Embryonic Stem Cells. Cell Rep. 2020, 30, 820–835. [Google Scholar] [CrossRef] [Green Version]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Wu, D.Y.; Zheng, H.; Wang, Y.; Sun, Q.R.; Liu, X.; Wang, L.Y.; Xiong, W.J.; Wang, Q.; Rhodes, J.D.P.; et al. Analysis of Genome Architecture during SCNT Reveals a Role of Cohesin in Impeding Minor ZGA. Mol. Cell 2020, 79, 234–250. [Google Scholar] [CrossRef]
- Smith, S.; Stillman, B. Purification and characterization of CAF-I, a human cell factor required for chromatin assembly during DNA replication in vitro. Cell 1989, 58, 15–25. [Google Scholar] [CrossRef]
- Verreault, A.; Kaufman, P.D.; Kobayashi, R.; Stillman, B. Nucleosome assembly by a complex of CAF-1 and acetylated histones H3/H4. Cell 1996, 87, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Yu, Z.; Zhang, S.; Liang, X.; Chen, J.; Li, C.; Ma, J.; Jiao, R. Drosophila CAF-1 regulates HP1-mediated epigenetic silencing and pericentric heterochromatin stability. J. Cell Sci. 2010, 123, 2853–2861. [Google Scholar] [CrossRef]
- McGrath, J.; Solter, D. Inability of mouse blastomere nuclei transferred to enucleated zygotes to support development in vitro. Science 1984, 226, 1317–1319. [Google Scholar] [CrossRef]
- Percharde, M.; Lin, C.J.; Yin, Y.; Guan, J.; Peixoto, G.A.; Bulut-Karslioglu, A.; Biechele, S.; Huang, B.; Shen, X.; Ramalho-Santos, M. A LINE1-Nucleolin Partnership Regulates Early Development and ESC Identity. Cell 2018, 174, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Kresoja-Rakic, J.; Santoro, R. Nucleolus and rRNA Gene Chromatin in Early Embryo Development. Trends Genet. TIG 2019, 35, 868–879. [Google Scholar] [CrossRef] [Green Version]
- Fléchon, J.E.; Kopecný, V. The nature of the ‘nucleolus precursor body’ in early preimplantation embryos: A review of fine-structure cytochemical, immunocytochemical and autoradiographic data related to nucleolar function. Zygote 1998, 6, 183–191. [Google Scholar] [CrossRef]
- Lavrentyeva, E.; Shishova, K.; Kagarlitsky, G.; Zatsepina, O. Localisation of RNAs and proteins in nucleolar precursor bodies of early mouse embryos. Reprod. Fertil. Dev. 2017, 29, 509–520. [Google Scholar] [CrossRef]
- Yao, R.W.; Xu, G.; Wang, Y.; Shan, L.; Luan, P.F.; Wang, Y.; Wu, M.; Yang, L.Z.; Xing, Y.H.; Yang, L.; et al. Nascent Pre-rRNA Sorting via Phase Separation Drives the Assembly of Dense Fibrillar Components in the Human Nucleolus. Mol. Cell 2019, 76, 767–783. [Google Scholar] [CrossRef]
- Lafontaine, D.L.J.; Riback, J.A.; Brangwynne, C.P.; Bascetin, R. The nucleolus as a multiphase liquid condensate. Nat. Rev. Mol. Cell Biol. 2021, 22, 165–182. [Google Scholar] [CrossRef]
- Yu, H.; Sun, Z.; Tan, T.; Pan, H.; Zhao, J.; Zhang, L.; Chen, J.; Lei, A.; Zhu, Y.; Chen, L.; et al. rRNA biogenesis regulates mouse 2C-like state by 3D structure reorganization of peri-nucleolar heterochromatin. Nat. Commun. 2021, 12, 6365. [Google Scholar] [CrossRef]
- Strom, A.R.; Emelyanov, A.V.; Mir, M.; Fyodorov, D.V.; Darzacq, X.; Karpen, G.H. Phase separation drives heterochromatin domain formation. Nature 2017, 547, 241–245. [Google Scholar] [CrossRef]
- Sanulli, S.; Trnka, M.J.; Dharmarajan, V.; Tibble, R.W.; Pascal, B.D.; Burlingame, A.L.; Griffin, P.R.; Gross, J.D.; Narlikar, G.J. HP1 reshapes nucleosome core to promote phase separation of heterochromatin. Nature 2019, 575, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Hnisz, D.; Shrinivas, K.; Young, R.A.; Chakraborty, A.K.; Sharp, P.A. A Phase Separation Model for Transcriptional Control. Cell 2017, 169, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakayama, T.; Perry, A.C.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Kyogoku, H.; Wakayama, T.; Kitajima, T.S.; Miyano, T. Single nucleolus precursor body formation in the pronucleus of mouse zygotes and SCNT embryos. PLoS ONE 2018, 13, e0202663. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Pang, N.; Liu, Z.; Lei, L. Transient inhibition of rDNA transcription in donor cells improves ribosome biogenesis and preimplantation development of embryos derived from somatic cell nuclear transfer. FASEB J. 2020, 34, 8283–8295. [Google Scholar] [CrossRef] [Green Version]
- Németh, A.; Conesa, A.; Santoyo-Lopez, J.; Medina, I.; Montaner, D.; Péterfia, B.; Solovei, I.; Cremer, T.; Dopazo, J.; Längst, G. Initial genomics of the human nucleolus. PLoS Genet. 2010, 6, e1000889. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.Y.; Shao, W.; Chang, L.; Yin, Y.; Li, T.; Zhang, H.; Hong, Y.; Percharde, M.; Guo, L.; Wu, Z.; et al. Genomic Repeats Categorize Genes with Distinct Functions for Orchestrated Regulation. Cell Rep. 2020, 30, 3296–3311. [Google Scholar] [CrossRef] [Green Version]
- Michalet, X.; Ekong, R.; Fougerousse, F.; Rousseaux, S.; Schurra, C.; Hornigold, N.; van Slegtenhorst, M.; Wolfe, J.; Povey, S.; Beckmann, J.S.; et al. Dynamic molecular combing: Stretching the whole human genome for high-resolution studies. Science 1997, 277, 1518–1523. [Google Scholar] [CrossRef] [Green Version]
- Técher, H.; Koundrioukoff, S.; Azar, D.; Wilhelm, T.; Carignon, S.; Brison, O.; Debatisse, M.; Le Tallec, B. Replication dynamics: Biases and robustness of DNA fiber analysis. J. Mol. Biol. 2013, 425, 4845–4855. [Google Scholar] [CrossRef]
- Nakatani, T.; Lin, J.; Ji, F.; Ettinger, A.; Pontabry, J.; Tokoro, M.; Altamirano-Pacheco, L.; Fiorentino, J.; Mahammadov, E.; Hatano, Y.; et al. DNA replication fork speed underlies cell fate changes and promotes reprogramming. Nat. Genet. 2022, 54, 318–327. [Google Scholar] [CrossRef]
- Poli, J.; Tsaponina, O.; Crabbé, L.; Keszthelyi, A.; Pantesco, V.; Chabes, A.; Lengronne, A.; Pasero, P. dNTP pools determine fork progression and origin usage under replication stress. EMBO J. 2012, 31, 883–894. [Google Scholar] [CrossRef]
- Atashpaz, S.; Shams, S.S.; Gonzalez, J.M.; Sebestyén, E.; Costanzo, V. ATR expands embryonic stem cell fate potential in response to replication stress. Elife Sci. 2020, 9, e54756. [Google Scholar] [CrossRef]
- Aze, A.; Sannino, V.; Soffientini, P.; Bachi, A.; Costanzo, V. Centromeric DNA replication reconstitution reveals DNA loops and ATR checkpoint suppression. Nat. Cell Biol. 2016, 18, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.Y.; Dai, J.J.; Wu, C.F.; Gu, X.L.; Liu, L.; Wu, Z.Q.; Xie, Y.N.; Wu, B.; Chen, H.L.; Li, Y.; et al. Positive effects of treatment of donor cells with aphidicolin on the preimplantation development of somatic cell nuclear transfer embryos in Chinese Bama mini-pig (Sus Scrofa). Anim. Sci. J. Nihon Chikusan Gakkaiho 2012, 83, 103–110. [Google Scholar] [CrossRef]
- Koo, O.J.; Hossein, M.S.; Hong, S.G.; Martinez-Conejero, J.A.; Lee, B.C. Cell cycle synchronization of canine ear fibroblasts for somatic cell nuclear transfer. Zygote 2009, 17, 37–43. [Google Scholar] [CrossRef]
- Tian, Q.; Zhou, L.Q. Lactate Activates Germline and Cleavage Embryo Genes in Mouse Embryonic Stem Cells. Cells 2022, 11, 548. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Lee, G.S.; Kim, H.S.; Hyun, S.H.; Kim, D.Y.; Lee, S.H.; Nam, D.H.; Jeong, Y.W.; Kim, S.; Kang, S.K.; Lee, B.C.; et al. Improved developmental competence of cloned porcine embryos with different energy supplements and chemical activation. Mol. Reprod. Dev. 2003, 66, 17–23. [Google Scholar] [CrossRef]
- Wilding, M.; Fiorentino, A.; De Simone, M.L.; Infante, V.; De Matteo, L.; Marino, M.; Dale, B. Energy substrates, mitochondrial membrane potential and human preimplantation embryo division. Reprod. Biomed. Online 2002, 5, 39–42. [Google Scholar] [CrossRef]
- Lane, M.; Gardner, D.K. Lactate regulates pyruvate uptake and metabolism in the preimplantation mouse embryo. Biol. Reprod. 2000, 62, 16–22. [Google Scholar] [CrossRef]
- Ghirlando, R.; Felsenfeld, G. CTCF: Making the right connections. Genes Dev. 2016, 30, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Xu, Y.; Chen, X.; Feng, S.; Liu, Z.; Sun, Y.; Yao, X.; Li, F.; Zhu, W.; Gao, L.; et al. 3D Chromatin Structures of Mature Gametes and Structural Reprogramming during Mammalian Embryogenesis. Cell 2017, 170, 367–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ke, Y.; Wu, K.; Zhao, H.; Sun, Y.; Gao, L.; Liu, Z.; Zhang, J.; Tao, W.; Hou, Z.; et al. Key role for CTCF in establishing chromatin structure in human embryos. Nature 2019, 576, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, X.; Zang, R.; Chen, J.; Fidalgo, M.; Sanchez-Priego, C.; Yang, J.; Caichen, A.; Ma, F.; Macfarlan, T.; et al. DUX-miR-344-ZMYM2-Mediated Activation of MERVL LTRs Induces a Totipotent 2C-like State. Cell Stem Cell 2020, 26, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Sagane, K.; Owa, T.; Mimori-Kiyosue, Y.; Shimizu, H.; Uesugi, M.; Ishihama, Y.; Iwata, M.; Mizui, Y. Splicing factor SF3b as a target of the antitumor natural product pladienolide. Nat. Chem. Biol. 2007, 3, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yang, M.; Li, S.; Zhang, J.; Peng, B.; Wang, C.; Chang, Z.; Ong, J.; Du, P. Mouse totipotent stem cells captured and maintained through spliceosomal repression. Cell 2021, 184, 2843–2859.e20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Features | 2-Cell-Stage Embryo | 2CLCs | Reference |
|---|---|---|---|
| Developmental competence | totipotent | expanded cell fate potential | [2,5] |
| 2-cell-specific transcripts | high level | high level | [2] |
| Pluripotency genes | downregulation | downregulation | [2] |
| Histones mobility | high core-histone mobility (H3.1, H3.2 and H2A) | high core-histone mobility (histones H3.1 and H2A) | [24,25] |
| Chromocenters | no defined chromocenters | no defined chromocenters | [26] |
| Nucleolus formation | NPBs | ring-like immature nucleoli | [27] |
| Chromatin accessibility | open chromatin landscape | open chromatin landscape | [26,28] |
| Higher-order chromatin organization | TADs and loops are weak | TADs and loops are weak | [29,30] |
| Global DNA methylation | global hypomethylation | global hypomethylation | [31,32] |
| Oxygen consumption | low | low | [33,34] |
| Glycolytic activity | low | low | [35,36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, B.; Ma, H.; Liu, D. 2-Cell-like Cells: An Avenue for Improving SCNT Efficiency. Biomolecules 2022, 12, 1611. https://doi.org/10.3390/biom12111611
Fu B, Ma H, Liu D. 2-Cell-like Cells: An Avenue for Improving SCNT Efficiency. Biomolecules. 2022; 12(11):1611. https://doi.org/10.3390/biom12111611
Chicago/Turabian StyleFu, Bo, Hong Ma, and Di Liu. 2022. "2-Cell-like Cells: An Avenue for Improving SCNT Efficiency" Biomolecules 12, no. 11: 1611. https://doi.org/10.3390/biom12111611