

Next Generation Sequencing of Genotype Variants and Genetic Association between Heat Shock Proteins HSPA1B Single Nucleotide Polymorphism at the g.31829044 Locus and Heat Tolerance: A Pilot Quasi-Experimental Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Heat Tolerance Test Procedure
2.3. Blood Collection and Deoxyribonucleic Acid (DNA) Extraction
2.4. Primer Design
2.5. Polymerase Chain Reaction (PCR)
2.6. PCR Clean-Up
2.7. Library Preparation, Quantification and Sequencing
2.8. SNP Detection
2.9. Statistical Analyses
3. Results
3.1. Heat Tolerance Status and HSP SNP
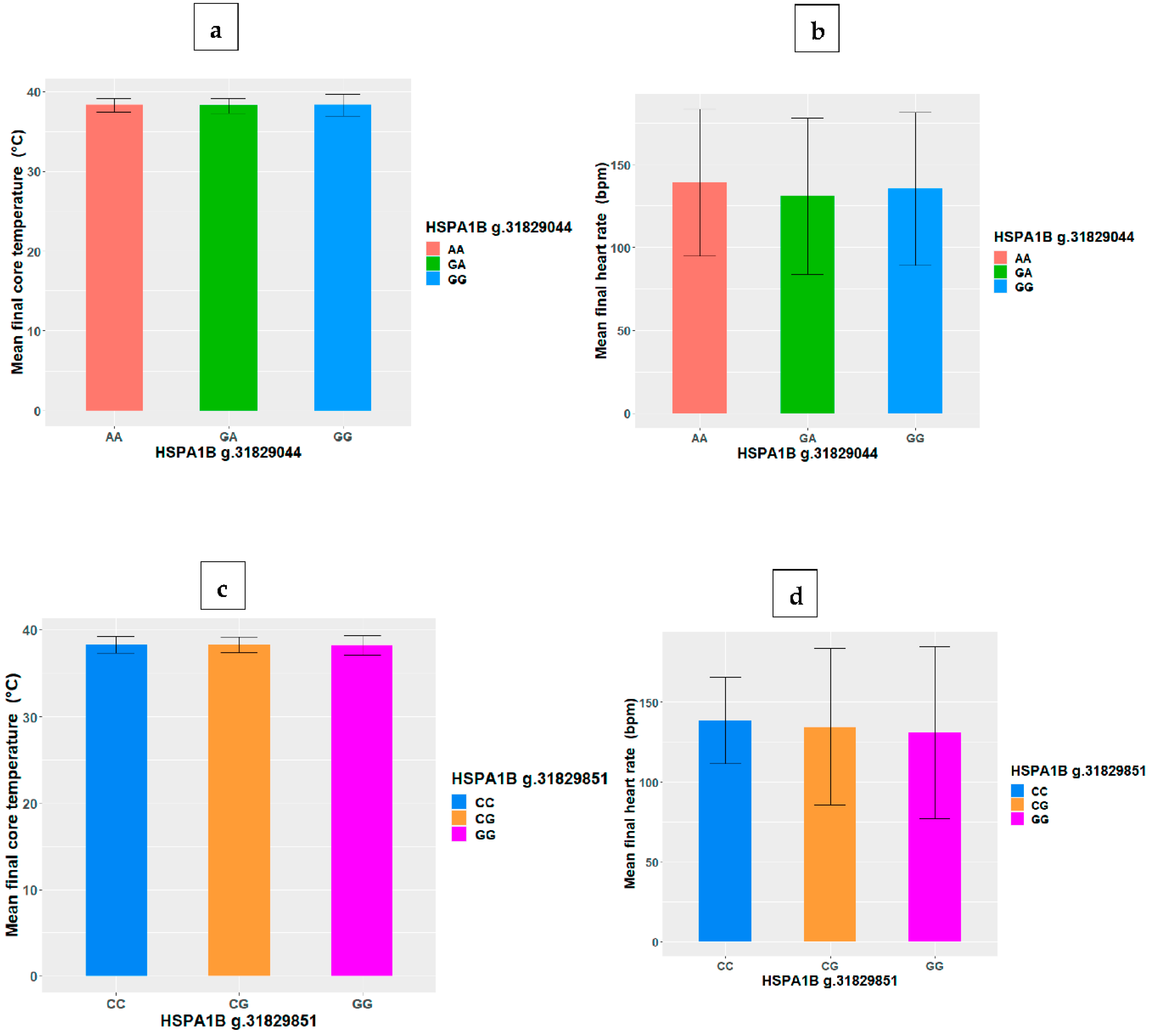
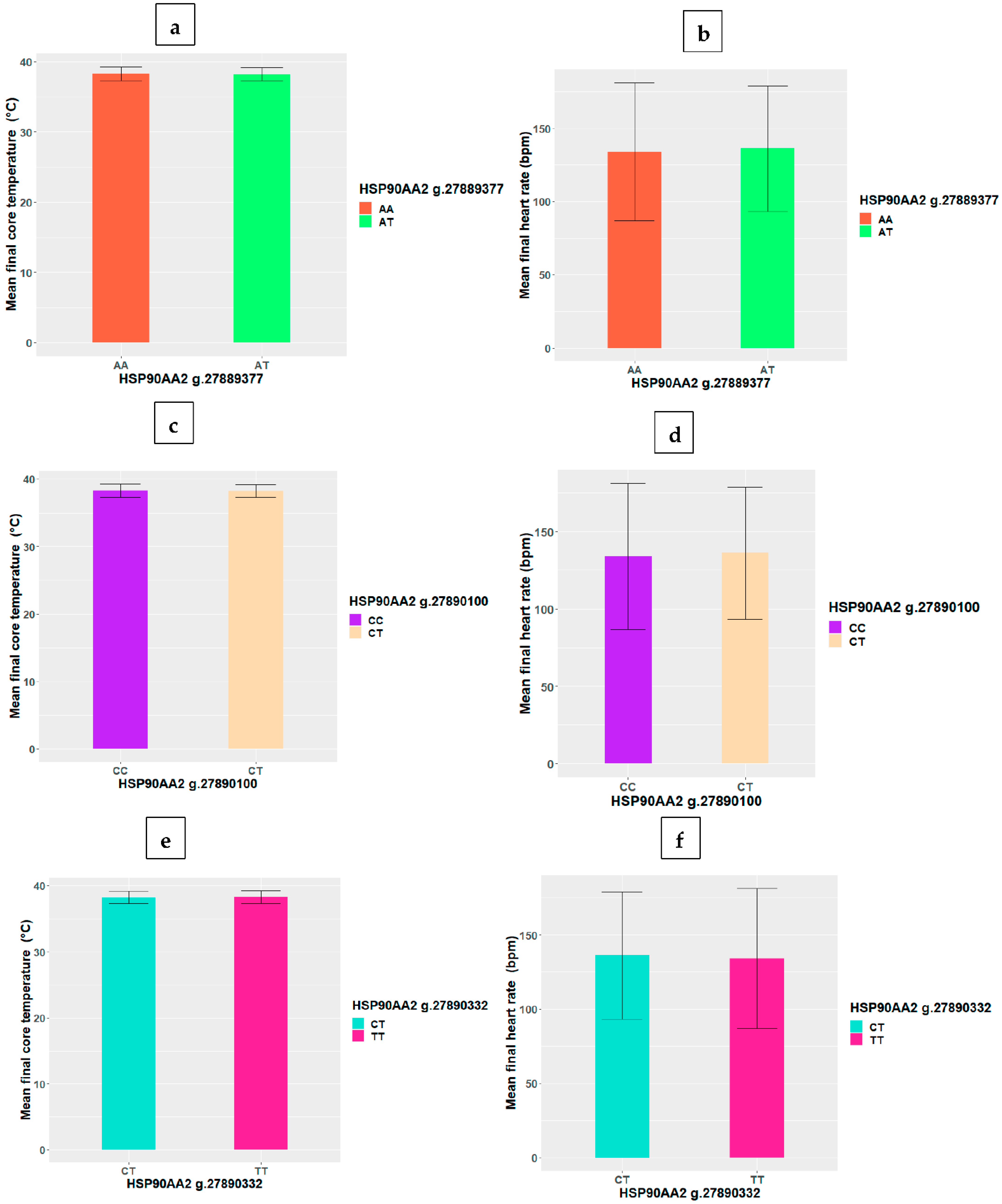
3.2. HSP SNP Differences in Final Core Temperature and Heart Rate
3.3. HSP SNP and EHS History
3.4. HSP SNP and Heat Tolerance Stratified by EHS History
4. Discussion
4.1. Strengths and Limitations
4.2. Implications for Policy and Future Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Miller, D.J.; Fort, P.E. Heat Shock Proteins Regulatory Role in Neurodevelopment. Front. Neurosci. 2018, 12, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solarz, A.; Majcher-Maślanka, I.; Kryst, J.; Chocyk, A. A Search for Biomarkers of Early-life Stress-related Psychopathology: Focus on 70-kDa Heat Shock Proteins. Neuroscience 2021, 463, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Szyller, J.; Bil-Lula, I. Heat Shock Proteins in Oxidative Stress and Ischemia/Reperfusion Injury and Benefits from Physical Exercises: A Review to the Current Knowledge. Oxid. Med. Cell. Longev. 2021, 2021, 6678457. [Google Scholar] [CrossRef]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jee, H. Size dependent classification of heat shock proteins: A mini-review. J. Exerc. Rehabil. 2016, 12, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Chang, N.-C.; Yang, H.-L.; Dai, C.-Y.; Lin, W.-Y.; Hsieh, M.-H.; Chien, C.-Y.; Ho, K.-Y. The association of heat shock protein genetic polymorphisms with age-related hearing impairment in Taiwan. J. Otolaryngol. Head Neck Surg. 2021, 50, 31. [Google Scholar] [CrossRef]
- Wang, H.; Ba, Y.; Han, W.; Zhang, H.; Zhu, L.; Jiang, P. Association of heat shock protein polymorphisms with patient susceptibility to coronary artery disease comorbid depression and anxiety in a Chinese population. PeerJ 2021, 9, e11636. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Q.; Yu, H.; Du, H.; Li, L.; He, Y.; Zhu, S.; Li, C.; Zhang, S.; Luo, B.; et al. Genetic association study of a novel indel polymorphism in HSPA1B with the risk of sudden cardiac death in the Chinese populations. Forensic Sci. Int. 2021, 318, 110637. [Google Scholar] [CrossRef]
- Kohan, L.; Tabiee, O.; Sepahi, N. HSPA1L and HSPA1B gene polymorphisms and haplotypes are associated with idiopathic male infertility in Iranian population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 240, 57–61. [Google Scholar] [CrossRef]
- Salehi, Z.; Gholaminia, M.; Gholaminia, Z.; Panjtanpanah, M.; Qazvini, M.G. The GG genotype of the HSPA1B gene is associated with increased risk of glaucoma in northern Iran. Mol. Biol. 2017, 51, 31–36. [Google Scholar] [CrossRef]
- Vardar, S.A.; Doğanlar, Z.B.; Kaya, O.; Tayfur, P.; Sut, N.; Doğanlar, O. Different responses of apoptotic, inflammatory and heat shock protein gene expression to a single bout of high-intensity interval exercise between physically active and inactive men. Appl. Physiol. Nutr. Metab. 2021, 46, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Epstein, Y. Heat intolerance: Predisposing factor or residual injury? Med. Sci. Sport. Exerc. 1990, 22, 29–35. [Google Scholar] [CrossRef]
- Moore, A.C.; Stacey, M.J.; Bailey, K.G.H.; Bunn, R.J.; Woods, D.R.; Haworth, K.J.; Brett, S.J.; Folkes, S.E.F. Risk factors for heat illness among British soldiers in the hot collective training environment. J. R. Army Med. Corps 2016, 162, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Druyan, A.; Ketko, I.; Yanovich, R.; Epstein, Y.; Heled, Y. Refining the distinction between heat tolerant and intolerant individuals during a heat tolerance test. J. Therm. Biol. 2013, 38, 539–542. [Google Scholar] [CrossRef]
- Lisman, P.; Kazman, J.B.; O’Connor, F.G.; Heled, Y.; Deuster, P.A. Heat tolerance testing: Association between heat intolerance and anthropometric and fitness measurements. Mil. Med. 2014, 179, 1339–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, Y.; Magazanik, A.; Udassin, R.; Ben-Baruch, G.; Shvartz, E.; Shoenfeld, Y. Heat intolerance in former heatstroke patients. Ann. Intern. Med. 1979, 90, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Alele, F.O.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O.; Crowe, M.J. Individual Anthropometric, Aerobic Capacity and Demographic Characteristics as Predictors of Heat Intolerance in Military Populations. Medicina 2021, 57, 173. [Google Scholar] [CrossRef]
- O’Connor, F.G.; Heled, Y.; Deuster, P.A. Exertional heat stroke, the return to play decision, and the role of heat tolerance testing: A clinician’s dilemma. Curr. Sports Med. Rep. 2018, 17, 244–248. [Google Scholar] [CrossRef]
- Laramie, J.M.; Chung, T.P.; Brownstein, B.; Stormo, G.D.; Cobb, J.P. Transcriptional profiles of human epithelial cells in response to heat: Computational evidence for novel heat shock proteins. Shock 2008, 29, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Ruell, P.A.; Thompson, M.W.; Hoffman, K.M. Heat shock proteins as an aid in the treatment and diagnosis of heat stroke. J. Therm. Biol. 2009, 34, 1–7. [Google Scholar] [CrossRef]
- Sonna, L.A.; Sawka, M.N.; Lilly, C.M. Exertional heat illness and human gene expression. Prog. Brain Res. 2007, 162, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Sonna, L.A.; Gaffin, S.L.; Pratt, R.E.; Cullivan, M.L.; Angel, K.C.; Lilly, C.M. Selected Contribution: Effect of acute heat shock on gene expression by human peripheral blood mononuclear cells. J. Appl. Physiol. 2002, 92, 2208–2220. [Google Scholar] [CrossRef] [PubMed]
- Sonna, L.A.; Wenger, C.B.; Flinn, S.; Sheldon, H.K.; Sawka, M.N.; Lilly, C.M. Exertional heat injury and gene expression changes: A DNA microarray analysis study. J. Appl. Physiol. 2004, 96, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, O.R.; Tuttle, J.A.; Watt, P.W.; Maxwell, N.S.; Taylor, L. Hsp72 and Hsp90α mRNA transcription is characterised by large, sustained changes in core temperature during heat acclimation. Cell Stress Chaperones 2016, 21, 1021–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chand, K.; Iyer, K.; Mitra, D. Comparative analysis of differential gene expression of HSP40 and HSP70 family isoforms during heat stress and HIV-1 infection in T-cells. Cell Stress Chaperones 2021, 26, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graffelman, J.; Jain, D.; Weir, B. A genome-wide study of Hardy–Weinberg equilibrium with next generation sequence data. Hum. Genet. 2017, 136, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Slatkin, M. Linkage disequilibrium—Understanding the evolutionary past and mapping the medical future. Nat. Rev. Genet. 2008, 9, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, S.; Hagiwara, A.; Suzuki, M. Polymorphic analysis of the heat-shock protein 70 gene (HSPA1A) in Ménière’s disease. Acta Otolaryngol. 2008, 128, 1173–1177. [Google Scholar] [CrossRef]
- Ashworth, E.T.; Cotter, J.D.; Kilding, A.E. Methods for improving thermal tolerance in military personnel prior to deployment. Mil. Med. Res. 2020, 7, 58. [Google Scholar] [CrossRef]
- Alele, F.O.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O.; Crowe, M.J. Epidemiology of Exertional Heat Illness in the Military: A Systematic Review of Observational Studies. Int. J. Environ. Res. Public Health 2020, 17, 7037. [Google Scholar] [CrossRef] [PubMed]
- Gun, R. Deaths in Australia from Work-Related Heat Stress, 2000–2015. Int. J. Environ. Res. Public Health 2019, 16, 3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Kosoy, R.; Shameer, K.; Kumar, S.; Liu, L.; Readhead, B.; Belbin, G.M.; Lee, H.-C.; Chen, R.; Dudley, J.T. Genome-wide analysis indicates association between heterozygote advantage and healthy aging in humans. BMC Genet. 2019, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.K.; Taff, C.C.; Wheeler, S.S.; Weis, A.M.; Hinton, M.G.; Jones, M.L.; Logsdon, R.M.; Reisen, W.K.; Freund, D.; Sehgal, R.N.M.; et al. Low heterozygosity is associated with vector-borne disease in crows. Ecosphere 2018, 9, e02407. [Google Scholar] [CrossRef] [Green Version]
- Campbell, H.; Carothers, A.D.; Rudan, I.; Hayward, C.; Biloglav, Z.; Barac, L.; Pericic, M.; Janicijevic, B.; Smolej-Narancic, N.; Polasek, O.; et al. Effects of genome-wide heterozygosity on a range of biomedically relevant human quantitative traits. Hum. Mol. Genet. 2007, 16, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Bihlmeyer, N.A.; Brody, J.A.; Smith, A.V.; Lunetta, K.L.; Nalls, M.; Smith, J.A.; Tanaka, T.; Davies, G.; Yu, L.; Mirza, S.S.; et al. Genetic diversity is a predictor of mortality in humans. BMC Genet. 2014, 15, 159. [Google Scholar] [CrossRef] [Green Version]
- Wutich, A.; Lant, T.; White, D.D.; Larson, K.L.; Gartin, M. Comparing Focus Group and Individual Responses on Sensitive Topics: A Study of Water Decision Makers in a Desert City. Field Methods 2009, 22, 88–110. [Google Scholar] [CrossRef]
- Aquino-Gálvez, A.; González-Ávila, G.; Pérez-Rodríguez, M.; Partida-Rodríguez, O.; Nieves-Ramírez, M.; Piña-Ramírez, I.; Ramírez-Martínez, G.; Castillejos-López, M.; Checa, M.; Ruiz, V.; et al. Analysis of heat shock protein 70 gene polymorphisms Mexican patients with idiopathic pulmonary fibrosis. BMC Pulm. Med. 2015, 15, 129. [Google Scholar] [CrossRef] [Green Version]
- Mardan-Nik, M.; Pasdar, A.; Jamialahmadi, K.; Biabangard-Zak, A.; Mirhafez, S.R.; Ghalandari, M.; Tajfard, M.; Mohebati, M.; Esmaily, H.; Ferns, G.A. Association of heat shock protein70-2 (HSP70-2) gene polymorphism with coronary artery disease in an Iranian population. Gene 2014, 550, 180–184. [Google Scholar] [CrossRef]
- Boiocchi, C.; Osera, C.; Monti, M.C.; Ferraro, O.E.; Govoni, S.; Cuccia, M.; Montomoli, C.; Pascale, A.; Bergamaschi, R. Are Hsp70 protein expression and genetic polymorphism implicated in multiple sclerosis inflammation? J. Neuroimmunol. 2014, 268, 84–88. [Google Scholar] [CrossRef]
- Buraczynska, M.; Swatowski, A.; Buraczynska, K.; Dragan, M.; Ksiazek, A. Heat-shock protein gene polymorphisms and the risk of nephropathy in patients with Type 2 diabetes. Clin. Sci. 2008, 116, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Li, S.S.; Xie, Q.M.; Xu, J.H.; Sun, X.X.; Pan, F.M.; Xu, S.Q.; Liu, S.X.; Tao, J.H.; Liu, S.; et al. Associations of HSP90AA2 gene polymorphisms with disease susceptibility, glucocorticoids efficacy and health-related quality of life in Chinese systemic lupus erythematosus patients. Genes Genom. 2018, 40, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Stallings, J.D.; Ippolito, D.L.; Rakesh, V.; Baer, C.E.; Dennis, W.E.; Helwig, B.G.; Jackson, D.A.; Leon, L.R.; Lewis, J.A.; Reifman, J. Patterns of gene expression associated with recovery and injury in heat-stressed rats. BMC Genom. 2014, 15, 1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehbi, M.; Baturcam, E.; Eldali, A.; Ahmed, M.; Kwaasi, A.; Chishti, M.A.; Bouchama, A. Hsp-72, a candidate prognostic indicator of heatstroke. Cell Stress Chaperones 2010, 15, 593–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deuster, P.A.; Contreras-Sesvold, C.L.; O’Connor, F.G.; Campbell, W.W.; Kenney, K.; Capacchione, J.F.; Landau, M.E.; Muldoon, S.M.; Rushing, E.J.; Heled, Y. Genetic polymorphisms associated with exertional rhabdomyolysis. Eur. J. Appl. Physiol. 2013, 113, 1997–2004. [Google Scholar] [CrossRef]
- Landau, M.E.; Kenney, K.; Deuster, P.; Gonzalez, R.S.; Contreras-Sesvold, C.; Sambuughin, N.; O’Connor, F.G.; Campbell, W.W. Investigation of the relationship between serum creatine kinase and genetic polymorphisms in military recruits. Mil. Med. 2012, 177, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Schermann, H.; Hazut-Krauthammer, S.; Weksler, Y.; Spitzer, S.; Epstein, Y.; Kalmanovich, G.; Yanovich, R. When Should a Heat-Tolerance Test Be Scheduled After Clinical Recovery From an Exertional Heat Illness? J. Athl. Train. 2020, 55, 289–294. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Heat Tolerant | Heat Intolerant | p-Value |
|---|---|---|---|
| Age | 31 (15) | 27 (16) | 0.252 |
| Gender | |||
| Male | 17 (63.0%) | 17 (81.0%) | 0.174 |
| Female | 10 (37.0%) | 4 (19.0%) | |
| EHS history | |||
| Previous history of EHS | 6 (22.2%) | 8 (38.1%) | 0.230 |
| No history of EHS | 21 (77.8%) | 13 (61.9%) |
| Gene | SNP Locus | Genotype | Heat Tolerant | Heat Intolerant | p-Value |
|---|---|---|---|---|---|
| HSPA1Ba | g.31829044G>A | AA | 8 (30.8%) | 9 (45.0%) | 0.489 |
| AG | 15 (57.7%) | 8 (40.0%) | |||
| GG | 3 (11.5%) | 3 (15.0%) | |||
| g.31829851G>C | GG | 7 (26.9%) | 5 (25.0%) | 0.690 | |
| CG | 14 (56.8%) | 9 (45.0%) | |||
| CC | 5 (19.2%) | 6 (30.0%) | |||
| HSP90AA2b | g.27889377T>A | AA | 19 (70.4%) | 14 (70.0%) | 0.978 |
| AT | 8 (29.6%) | 6 (30.0%) | |||
| g.27890100C>T | CC | 19 (70.4%) | 14 (70.0%) | 0.978 | |
| CT | 8 (29.6%) | 6 (30.0%) | |||
| g.27890332T>C | TT | 19 (70.4%) | 14 (70.0%) | 0.978 | |
| CT | 8 (29.6%) | 6 (30.0%) |
| Gene | SNP Locus | Final Core Temp | Final HR |
|---|---|---|---|
| HSPA1B | g.31829044G>A | −0.05 | −0.106 |
| g.31829851 G>C | 0.072 | 0.118 | |
| HSP90AA2 | g.27889377T>C | −0.041 | 0.063 |
| g.27890100C>T | −0.041 | 0.063 | |
| g.27890332T>C | −0.041 | 0.063 |
| Gene | SNP Locus | Genotype | EHS History | No EHS History | p-Value |
|---|---|---|---|---|---|
| HSPA1Ba | g.31829044G>A | AA | 4 (28.6%) | 13 (40.6%) | 0.736 |
| AG | 8 (57.1%) | 15 (46.9%) | |||
| GG | 2 (14.3%) | 4 (12.5%) | |||
| g.31829851G>C | GG | 4 (28.6%) | 8 (25.0%) | 0.360 | |
| CG | 5 (35.7%) | 18 (56.3%) | |||
| CC | 5 (35.7%) | 6 (18.8%) | |||
| HSP90AA2b | g.27889377T>A | AA | 9 (64.3%) | 24 (72.7%) | 0.563 |
| AT | 5 (35.7%) | 9 (27.3%) | |||
| g.27890100C>T | CC | 9 (64.3%) | 24 (72.7%) | 0.563 | |
| CT | 5 (35.7%) | 9 (27.3%) | |||
| g.27890332T>C | TT | 9 (64.3%) | 24 (72.7%) | 0.563 | |
| CT | 5 (35.7%) | 9 (27.3%) |
| EHS History | No History of EHS | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SNP | Genotype | Heat Tolerant n (%) | Heat Intolerant n (%) | p-Value | FDR p-Value | Heat Tolerant n (%) | Heat Intolerant n (%) | p-Value | FDR p-Value |
| HSPA1B | g.31829044G>A | AA | 0 (0%) | 4 (50%) | 0.024 | 0.048 | 8 (40.0%) | 5 (41.7%) | 1 | 1 |
| AG | 6 (100%) | 2 (25%) | 9 (45.5%) | 6 (50.0%) | ||||||
| GG | 0 (0%) | 2 (25%) | 3 (15.0%) | 1 (8.3%) | ||||||
| g.31829851G>C | GG | 1 (16.7%) | 3 (37.5%) | 0.201 | 0.402 | 6 (30.0%) | 2 (16.7%) | 0.703 | 0.703 | |
| CG | 4 (66.7%) | 1 (12.5%) | 10 (50.0%) | 8 (66.7%) | ||||||
| CC | 1 (16.7%) | 4 (50.0%) | 4 (20.0%) | 2 (16.7%) | ||||||
| HSP90AA2 | g.27889377T>A | AA | 4 (66.7%) | 5 (62.5%) | 0.872 | 0.872 | 15 (71.4%) | 9 (75.0%) | 0.825 | 0.872 |
| AT | 2 (33.3%) | 3 (37.5%) | 6 (28.6%) | 3 (25.0%) | ||||||
| g.27890100C>T | TT | 4 (66.7%) | 5 (62.5%) | 0.872 | 0.872 | 15 (71.4%) | 9 (75.0%) | 0.825 | 0.872 | |
| CT | 2 (33.3%) | 3 (37.5%) | 6 (28.6%) | 3 (25.0%) | ||||||
| g.27890332T>C | CC | 4 (66.7%) | 5 (62.5%) | 0.872 | 0.872 | 15 (71.4%) | 9 (75.0%) | 0.825 | 0.872 | |
| CT | 2 (33.3%) | 3 (37.5%) | 6 (28.6%) | 3 (25.0%) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alele, F.O.; Otto, J.R.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O. Next Generation Sequencing of Genotype Variants and Genetic Association between Heat Shock Proteins HSPA1B Single Nucleotide Polymorphism at the g.31829044 Locus and Heat Tolerance: A Pilot Quasi-Experimental Study. Biomolecules 2022, 12, 1465. https://doi.org/10.3390/biom12101465
Alele FO, Otto JR, Malau-Aduli BS, Malau-Aduli AEO. Next Generation Sequencing of Genotype Variants and Genetic Association between Heat Shock Proteins HSPA1B Single Nucleotide Polymorphism at the g.31829044 Locus and Heat Tolerance: A Pilot Quasi-Experimental Study. Biomolecules. 2022; 12(10):1465. https://doi.org/10.3390/biom12101465
Chicago/Turabian StyleAlele, Faith O., John R. Otto, Bunmi S. Malau-Aduli, and Aduli E. O. Malau-Aduli. 2022. "Next Generation Sequencing of Genotype Variants and Genetic Association between Heat Shock Proteins HSPA1B Single Nucleotide Polymorphism at the g.31829044 Locus and Heat Tolerance: A Pilot Quasi-Experimental Study" Biomolecules 12, no. 10: 1465. https://doi.org/10.3390/biom12101465