
Synaptic Plasticity Is Predicted by Spatiotemporal Firing Rate Patterns and Robust to In Vivo-like Variability


Abstract
:1. Introduction
2. Methods
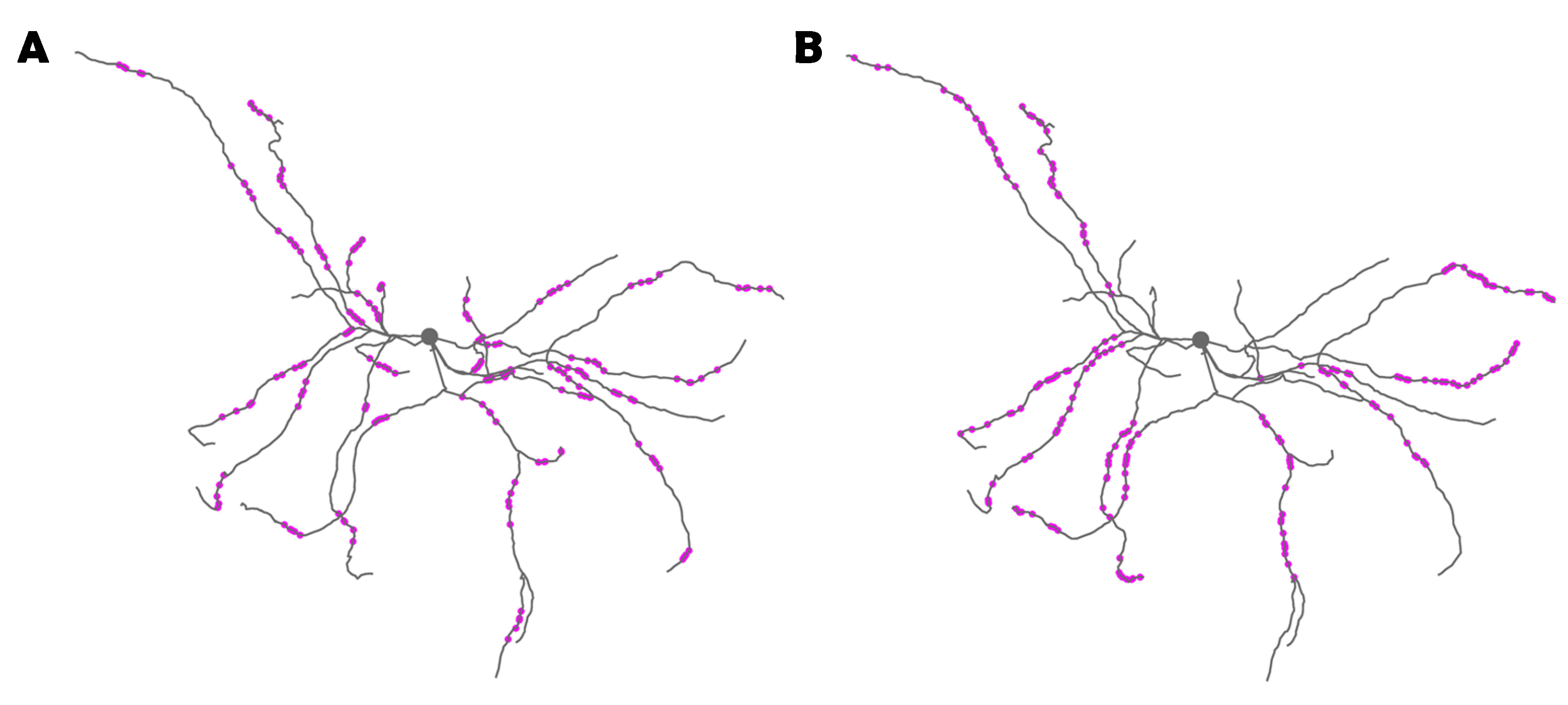
2.1. Spn Model Morphology and Passive Membrane Properties
2.2. Voltage Gated Ion Channels
2.3. Calcium Dynamics
2.4. Synaptic Channels
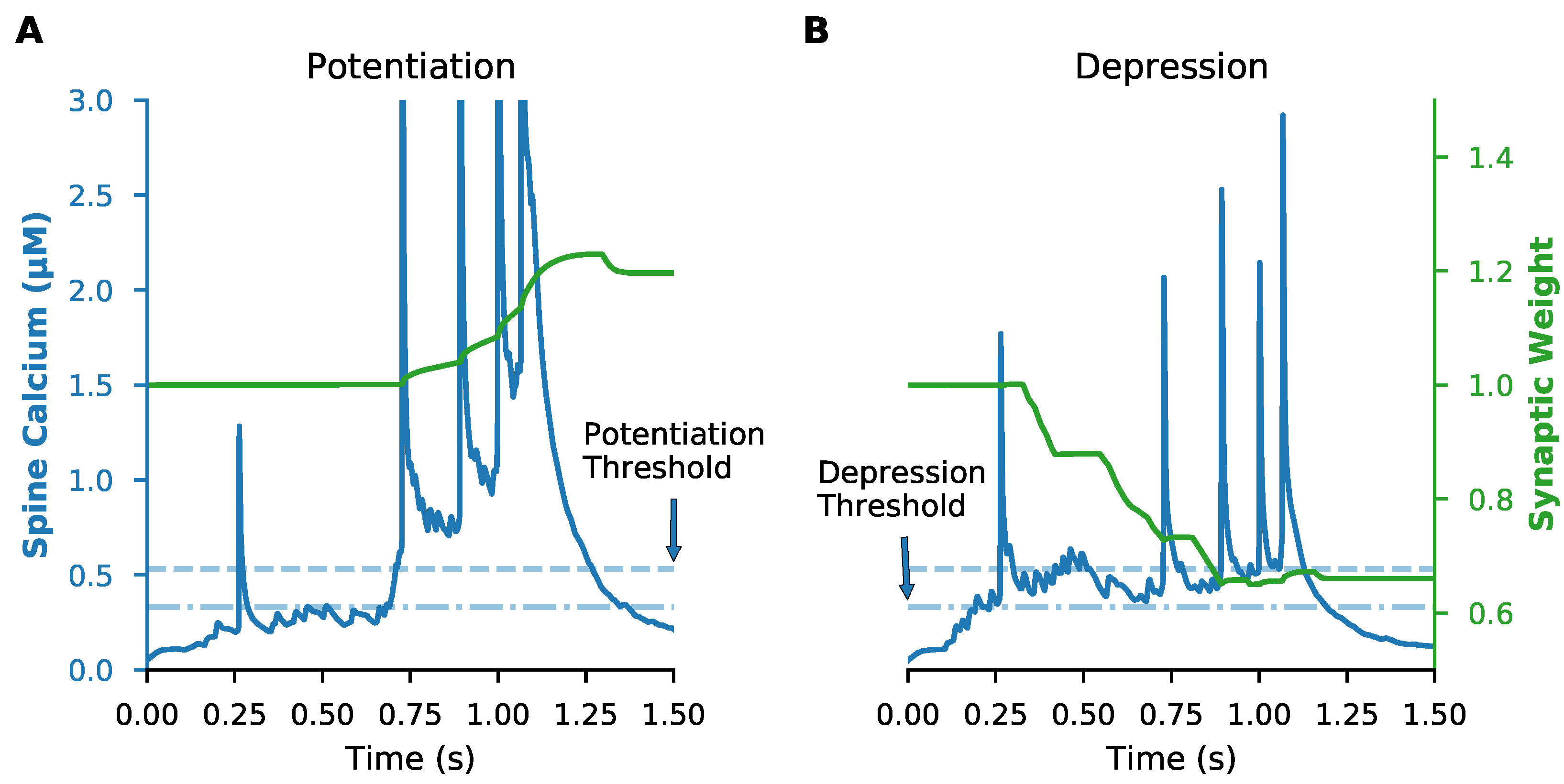
2.5. Plasticity Rule
2.6. Synaptic Inputs
2.7. Analysis
2.8. Code Accessibility
3. Results
3.1. Data-Driven SPN Model Exhibits Calcium-Based Synaptic Plasticity for In Vivo-like Inputs
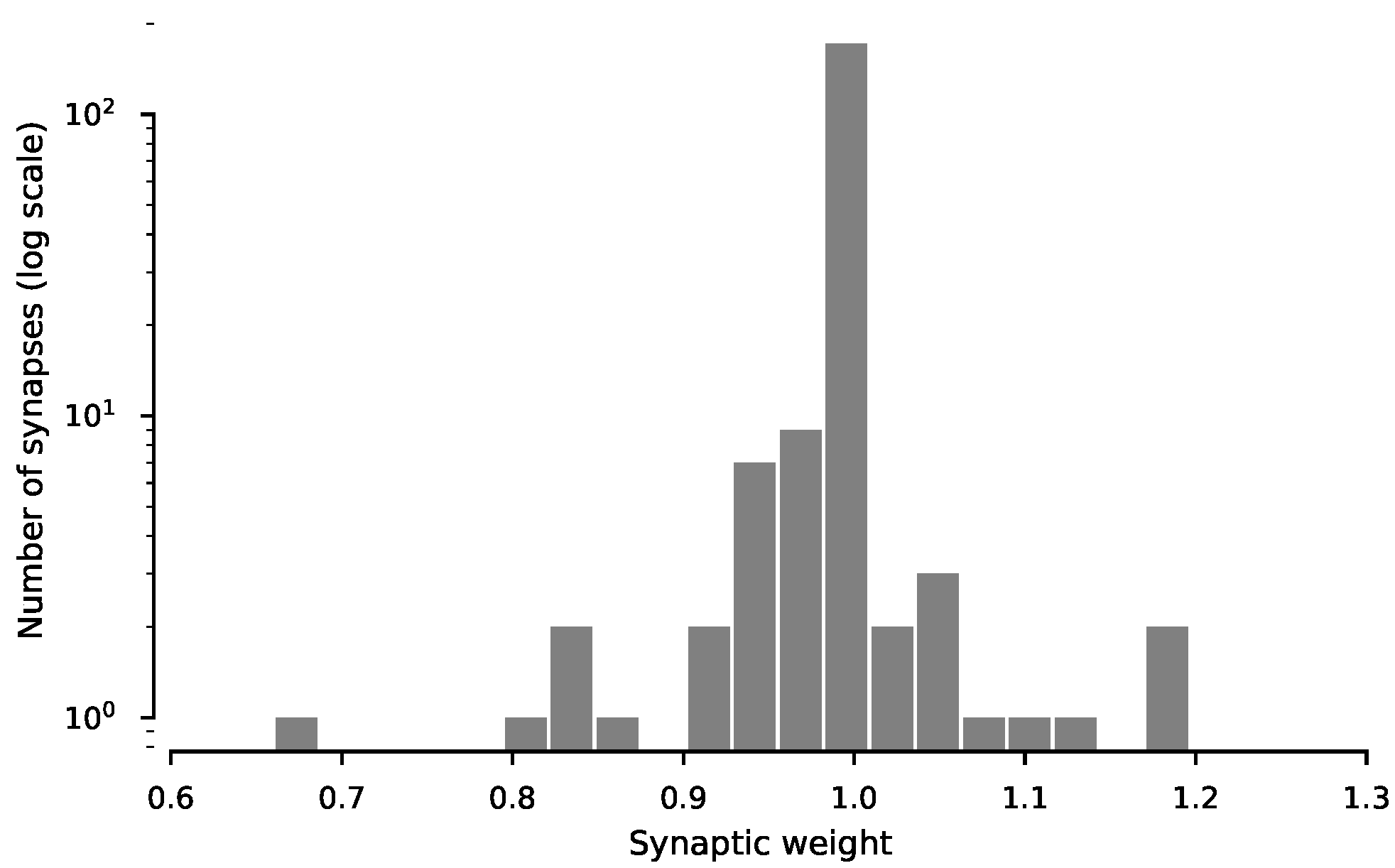
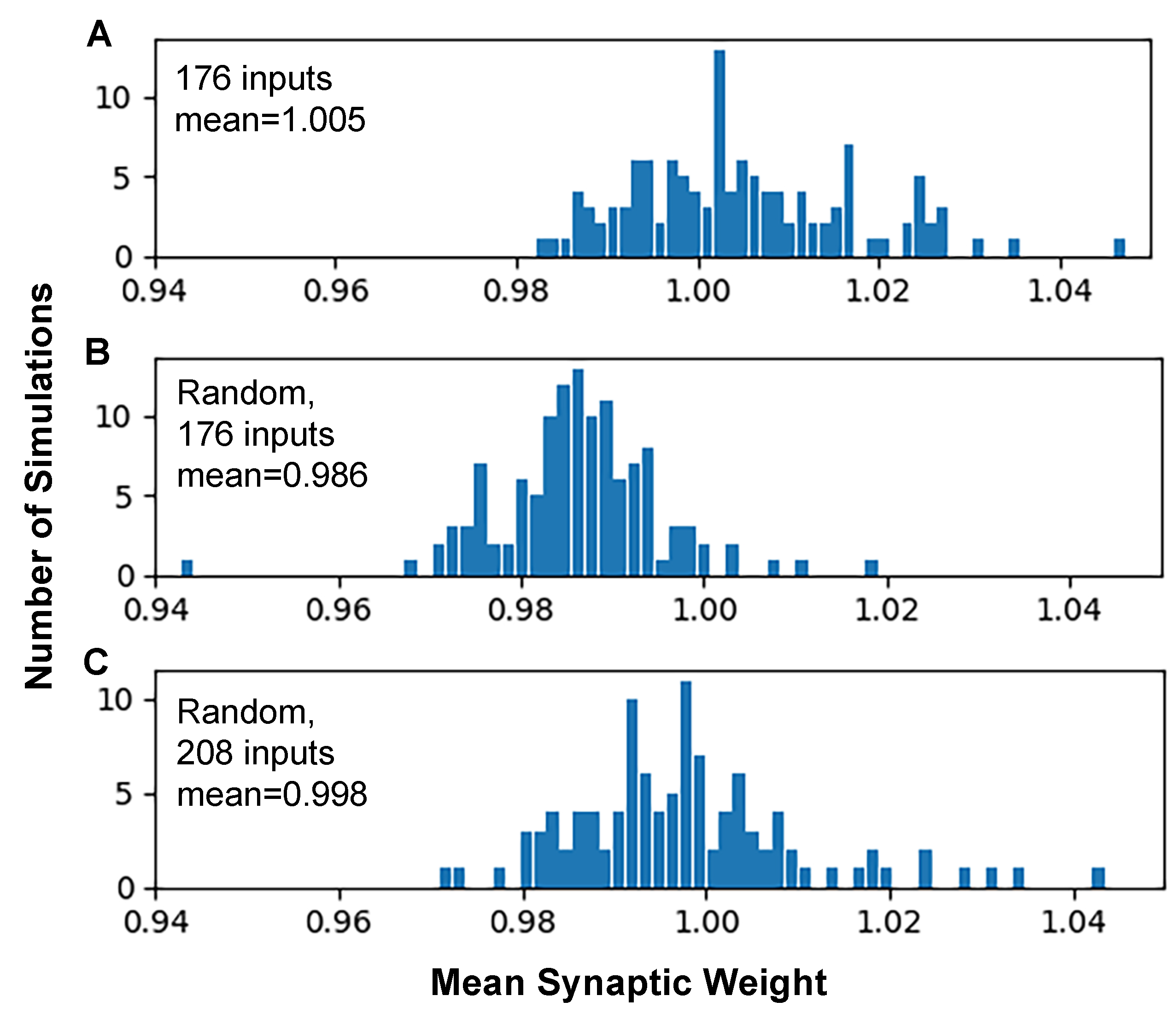
3.2. Synaptic Plasticity Is Highly Robust to Trial-to-Trial Variability
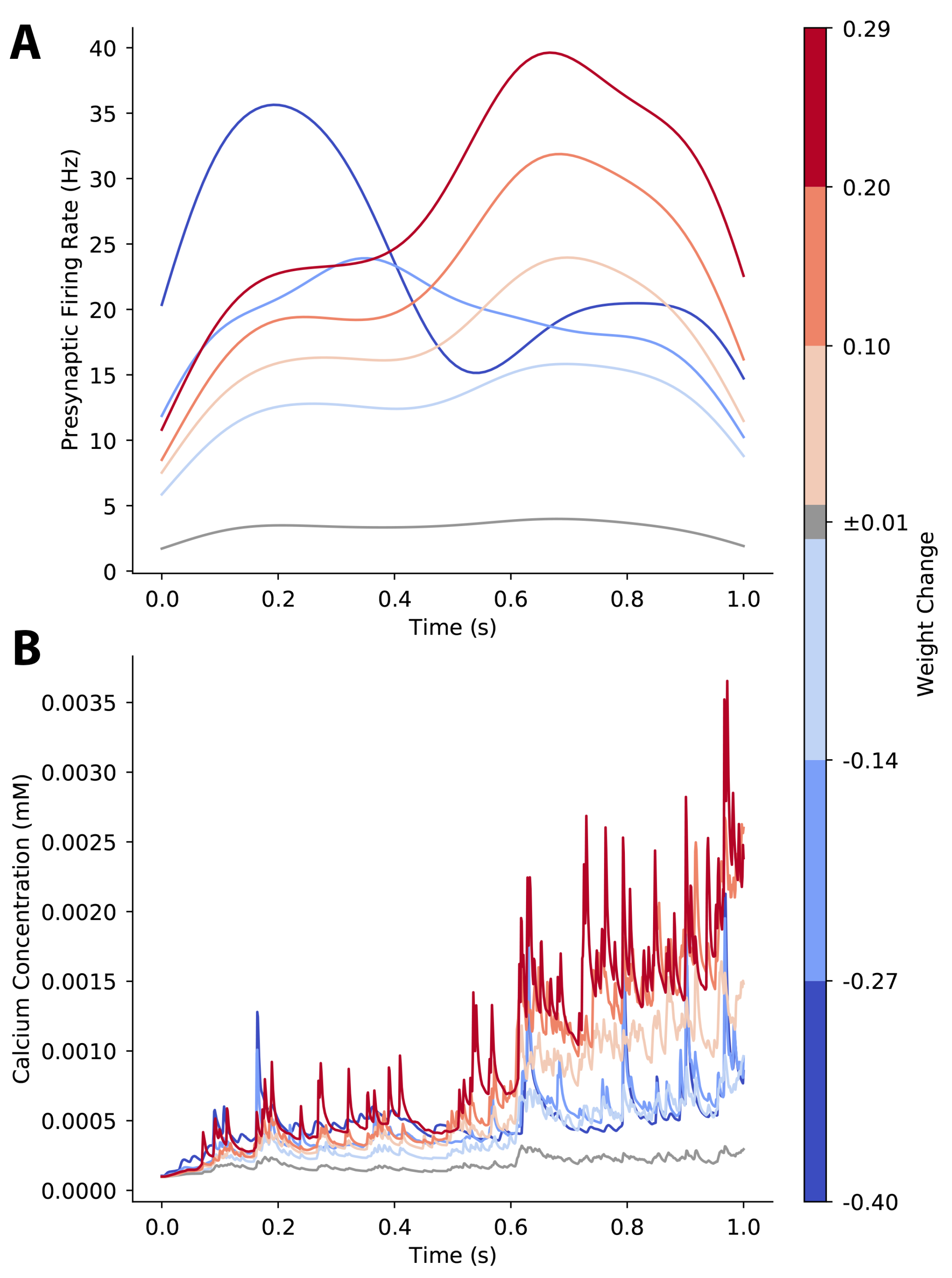
3.3. Plasticity of a Single Synapse Is Only Partially Predicted by Its Pre-Synaptic Activity
3.4. Plasticity of a Single Synapse Is Affected by Temporal Pattern of Pre-Synaptic Firing Rate
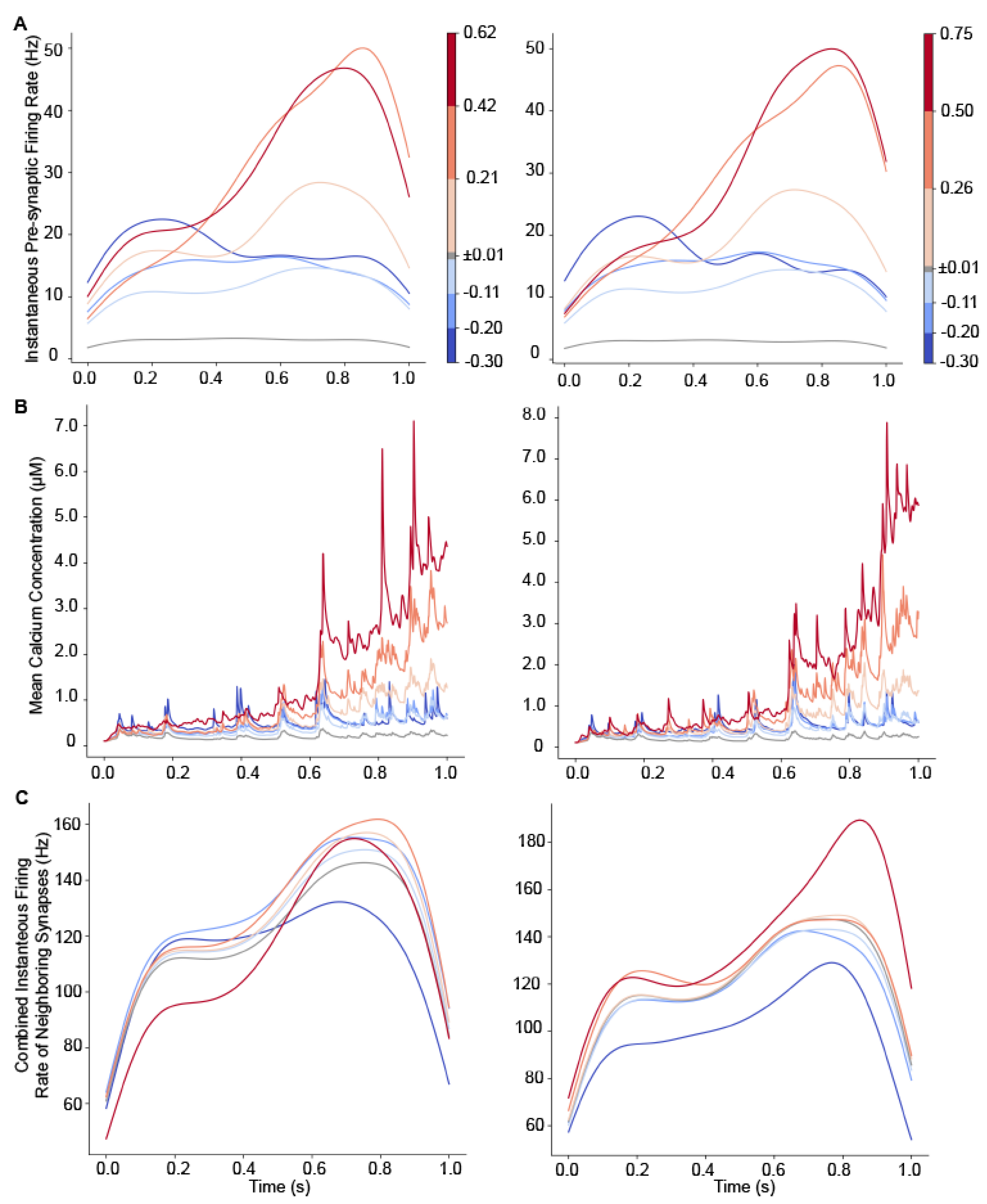
3.5. Plasticity of a Single Synapse Is Affected by Neighboring Synaptic Activity
3.6. Pre-Synaptic Firing Rate and Cluster Length Predict Synaptic Plasticity
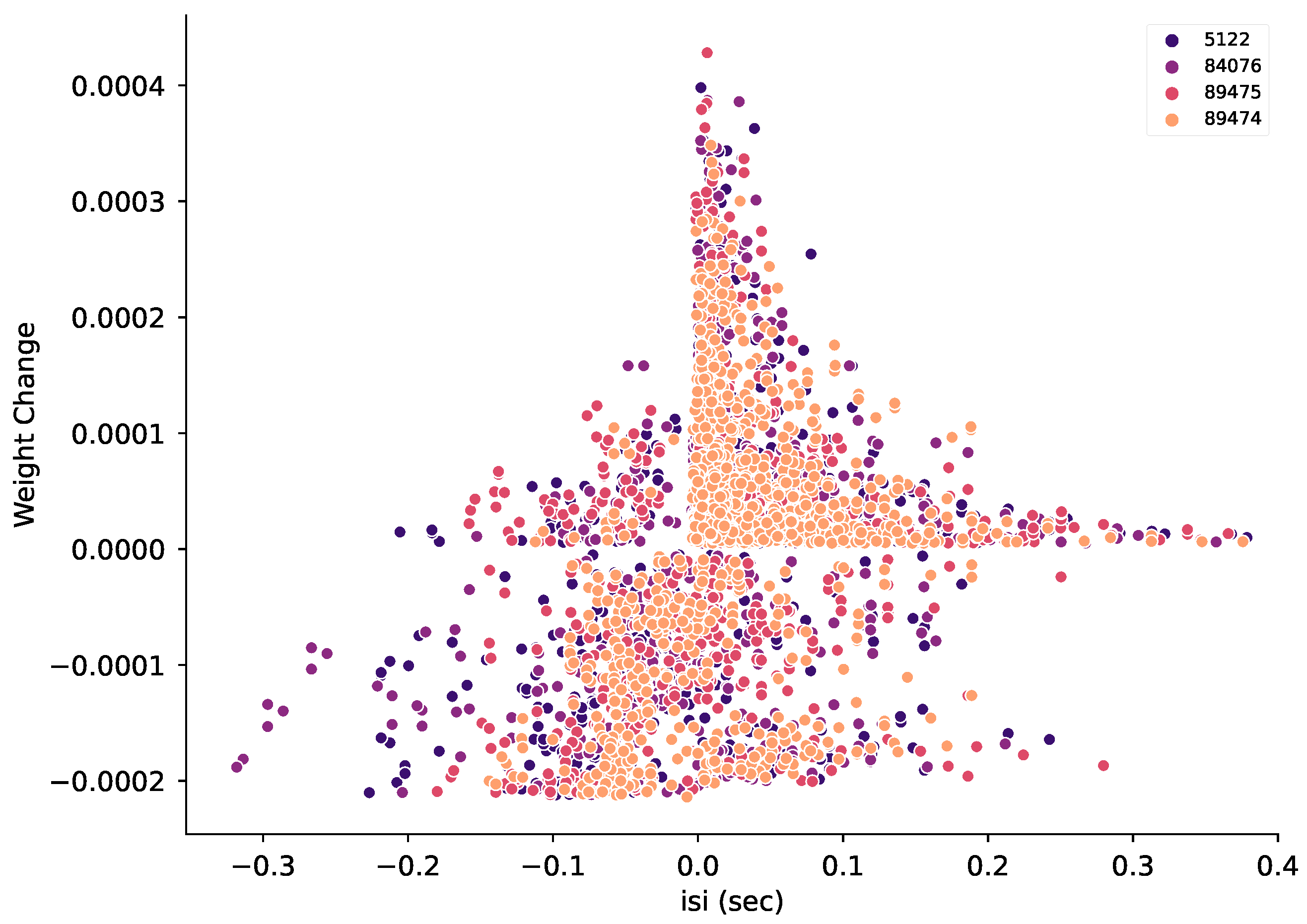
3.7. Spike Timing Together with Pre-Synaptic Firing Rate Can Predict Instantaneous Weight Change
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SPN | Spiny projection neuron |
| LTP | Long term potentiation |
| LTD | Long term depression |
| AHP | Afterhyperpolarization |
| ISI | Inter-spike interval |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Channel | Soma | <50 Microns | >50 Microns |
|---|---|---|---|
| Krp | 27.25 | 27.25 | 27.25 |
| KaF | 592.75 | 198.78 | 70.22 |
| KaS | 918.22 | 121.14 | 1.70 |
| Kir | 3.18 | 28.96 | 12.26 |
| CaL13 | 0.07 | 0.00 | 0.00 |
| CaL12 | 0.00 | 4.09 | 0.02 |
| CaR | 2.54 | 27.28 | 1.16 |
| CaN | 0.00 | 0.00 | 0.00 |
| CaT33 | 0.00 | 0.07 | 0.01 |
| CaT32 | 0.00 | 0.45 | 0.24 |
| NaF | 17,611.46 | 720.28 | 126.05 |
| SKCa | 0.42 | 0.42 | 0.42 |
| BKCa | 46.39 | 46.39 | 46.39 |
| CaCC | 0.79 | 0.79 | 0.79 |





References
- Takeuchi, T.; Duszkiewicz, A.; Morris, R. The Synaptic Plasticity and Memory Hypothesis: Encoding, Storage and Persistence. PHilosophical Trans. R. Soc. Lond. Ser. Biol. Sci. 2014, 369, 20130288. [Google Scholar] [CrossRef] [Green Version]
- Perrin, E.; Venance, L. Bridging the Gap between Striatal Plasticity and Learning. Curr. Opin. Neurobiol. 2019, 54, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Buonomano, D.V.; Merzenich, M.M. Cortical Plasticity: From Synapses to Maps. Annu. Rev. Neurosci. 1998, 21, 149–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.J.; Morris, R.G.M. New Life in an Old Idea: The Synaptic Plasticity and Memory Hypothesis Revisited. Hippocampus 2002, 12, 609–636. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.C.; Robins, A. Memory Retention—The Synaptic Stability versus Plasticity Dilemma. Trends Neurosci. 2005, 28, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Josselyn, S.A.; Tonegawa, S. Memory Engrams: Recalling the Past and Imagining the Future. Science 2020, 367, eaaw4325. [Google Scholar] [CrossRef]
- Fisher, S.D.; Robertson, P.B.; Black, M.J.; Redgrave, P.; Sagar, M.A.; Abraham, W.C.; Reynolds, J.N. Reinforcement Determines the Timing Dependence of Corticostriatal Synaptic Plasticity in Vivo. Nat. Commun. 2017, 8, 334. [Google Scholar] [CrossRef]
- Trachtenberg, J.T.; Chen, B.E.; Knott, G.W.; Feng, G.; Sanes, J.R.; Welker, E.; Svoboda, K. Long-Term in Vivo Imaging of Experience-Dependent Synaptic Plasticity in Adult Cortex. Nature 2002, 420, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Winnubst, J.; Cheyne, J.E.; Niculescu, D.; Lohmann, C. Spontaneous Activity Drives Local Synaptic Plasticity In Vivo. Neuron 2015, 87, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cudmore, R.H.; Lin, D.T.; Linden, D.J.; Huganir, R.L. Visualization of NMDA Receptor—Dependent AMPA Receptor Synaptic Plasticity in Vivo. Nat. Neurosci. 2015, 18, 402–407. [Google Scholar] [CrossRef]
- Sjöström, P.J.; Turrigiano, G.G.; Nelson, S.B. Rate, Timing, and Cooperativity Jointly Determine Cortical Synaptic Plasticity. Neuron 2001, 32, 1149–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rossum, M.C.W.; Bi, G.Q.; Turrigiano, G.G. Stable Hebbian Learning from Spike Timing-Dependent Plasticity. J. Neurosci. 2000, 20, 8812–8821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporale, N.; Dan, Y. Spike Timing—Dependent Plasticity: A Hebbian Learning Rule. Annu. Rev. Neurosci. 2008, 31, 25–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovinger, D.M.; Tyler, E.C.; Merritt, A. Short- and Long-Term Synaptic Depression in Rat Neostriatum. J. Neurophysiol. 1993, 70, 1937–1949. [Google Scholar] [CrossRef]
- Hawes, S.L.; Gillani, F.; Evans, R.C.; Benkert, E.A.; Blackwell, K.T. Sensitivity to Theta-Burst Timing Permits LTP in Dorsal Striatal Adult Brain Slice. J. Neurophysiol. 2013, 110, 2027–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadlen, M.N.; Newsome, W.T. The Variable Discharge of Cortical Neurons: Implications for Connectivity, Computation, and Information Coding. J. Neurosci. 1998, 18, 3870–3896. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.F.; Zador, A.M. Input Synchrony and the Irregular Firing of Cortical Neurons. Nat. Neurosci. 1998, 1, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Graupner, M.; Wallisch, P.; Ostojic, S. Natural Firing Patterns Imply Low Sensitivity of Synaptic Plasticity to Spike Timing Compared with Firing Rate. J. Neurosci. 2016, 36, 11238–11258. [Google Scholar] [CrossRef] [Green Version]
- Froemke, R.C.; Dan, Y. Spike-timing-dependent synaptic modification induced by natural spike trains. Nature 2002, 416, 433–438. [Google Scholar] [CrossRef]
- Brandalise, F.; Carta, S.; Helmchen, F.; Lisman, J.; Gerber, U. Dendritic NMDA Spikes Are Necessary for Timing-Dependent Associative LTP in CA3 Pyramidal Cells. Nat. Commun. 2016, 7, 13480. [Google Scholar] [CrossRef] [Green Version]
- Kastellakis, G.; Cai, D.J.; Mednick, S.C.; Silva, A.J.; Poirazi, P. Synaptic Clustering within Dendrites: An Emerging Theory of Memory Formation. Prog. Neurobiol. 2015, 126, 19–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöström, P.J.; Rancz, E.A.; Roth, A.; Häusser, M. Dendritic Excitability and Synaptic Plasticity. Physiol. Rev. 2008, 88, 769–840. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.P.; Andrásfalvy, B.K.; Polito, M.; Magó, Á.; Ujfalussy, B.B.; Makara, J.K. Location-Dependent Synaptic Plasticity Rules by Dendritic Spine Cooperativity. Nat. Commun. 2016, 7, 11380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golding, N.L.; Staff, N.P.; Spruston, N. Dendritic Spikes as a Mechanism for Cooperative Long-Term Potentiation. Nature 2002, 418, 326–331. [Google Scholar] [CrossRef]
- Yagishita, S.; Hayashi-Takagi, A.; Ellis-Davies, G.C.; Urakubo, H.; Ishii, S.; Kasai, H. A Critical Time Window for Dopamine Actions on the Structural Plasticity of Dendritic Spines. Science 2014, 345, 1616–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, J.N.; Avvisati, R.; Dodson, P.D.; Fisher, S.D.; Oswald, M.J.; Wickens, J.R.; Zhang, Y.F. Coincidence of cholinergic pauses, dopaminergic activation and depolarisation of spiny projection neurons drives synaptic plasticity in the striatum. Nat. Commun. 2021, 13, 1296. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Huertas, M.; Hong, S.Z.; Tie, X.; Hell, J.W.; Shouval, H.; Kirkwood, A. Distinct Eligibility Traces for LTP and LTD in Cortical Synapses. Neuron 2015, 88, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Kreitzer, A.C.; Malenka, R.C. Striatal Plasticity and Basal Ganglia Circuit Function. Neuron 2008, 60, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.C.; Blackwell, K.T. Calcium: Amplitude, Duration, or Location? Biol. Bull. 2015, 228, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Nevian, T.; Sakmann, B. Spine Ca2+ Signaling in Spike-Timing-Dependent Plasticity. J. Neurosci. 2006, 26, 11001–11013. [Google Scholar] [CrossRef] [Green Version]
- Zucker, R.S. Calcium- and Activity-Dependent Synaptic Plasticity. Curr. Opin. Neurobiol. 1999, 9, 305–313. [Google Scholar] [CrossRef]
- Rubin, J.E.; Gerkin, R.C.; Bi, G.Q.; Chow, C.C. Calcium time course as a signal for spike-timing-dependent plasticity. J. Neurophysiol. 2005, 93, 2600–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graupner, M.; Brunel, N. Calcium-Based Plasticity Model Explains Sensitivity of Synaptic Changes to Spike Pattern, Rate, and Dendritic Location. Proc. Natl. Acad. Sci. USA 2012, 109, 3991–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standage, D.; Trappenberg, T.; Blohm, G. Calcium-Dependent Calcium Decay Explains STDP in a Dynamic Model of Hippocampal Synapses. PLoS ONE 2014, 9, e86248. [Google Scholar] [CrossRef] [PubMed]
- Bush, D.; Jin, Y. Calcium control of triphasic hippocampal STDP. J. Comput. Neurosci. 2012, 33, 495–514. [Google Scholar] [CrossRef] [Green Version]
- Jȩdrzejewska-Szmek, J.; Damodaran, S.; Dorman, D.B.; Blackwell, K.T. Calcium Dynamics Predict Direction of Synaptic Plasticity in Striatal Spiny Projection Neurons. Eur. J. Neurosci. 2017, 45, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Dorman, D.B.; Blackwell, K.T. Ajustador. 2021. Available online: https://github.com/neurord/ajustador/releases/tag/v2.1 (accessed on 16 July 2021).
- Shindou, T.; Ochi-Shindou, M.; Wickens, J.R. A Ca2+ Threshold for Induction of Spike-Timing-Dependent Depression in the Mouse Striatum. J. Neurosci. 2011, 31, 13015–13022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotkin, J.L.; Day, M.; Surmeier, D.J. Synaptically Driven State Transitions in Distal Dendrites of Striatal Spiny Neurons. Nat. Neurosci. 2011, 14, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.N.D.; Plenz, D. Dendritic Calcium Encodes Striatal Neuron Output during Up-States. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 1499–1512. [Google Scholar] [CrossRef] [Green Version]
- Corbit, V.L.; Manning, E.E.; Witten, I.B.; Gittis, A.H.; Ahmari, S.E. Strengthened Inputs from Secondary Motor Cortex to Striatum in a Mouse Model of Compulsive Behavior. J. Neurosci. 2019, 39, 2965–2975. [Google Scholar] [CrossRef] [Green Version]
- Corbit, V.L.; Piantadosi, S.C.; Wood, J.; Liu, G.; Choi, C.J.; Witten, I.B.; Gittis, A.H.; Ahmari, S.E. Dissociable roles of central striatum and anterior lateral motor area in initiating and sustaining naturalistic behavior. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dorman, D.B.; Jȩdrzejewska-Szmek, J.; Blackwell, K.T. Inhibition Enhances Spatially-Specific Calcium Encoding of Synaptic Input Patterns in a Biologically Constrained Model. eLife 2018, 7, e38588. [Google Scholar] [CrossRef] [PubMed]
- Goodliffe, J.W.; Song, H.; Rubakovic, A.; Chang, W.; Medalla, M.; Weaver, C.M.; Luebke, J.I. Differential Changes to D1 and D2 Medium Spiny Neurons in the 12-Month-Old Q175+/- Mouse Model of Huntington’s Disease. PLoS ONE 2018, 13, e0200626. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, G.A.; Donohue, D.E.; Halavi, M. NeuroMorpho.Org: A Central Resource for Neuronal Morphologies. J. Neurosci. 2007, 27, 9247–9251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, W.R.; Ambros-Ingerson, J.; Grover, L.M. Fitting Experimental Data to Models That Use Morphological Data from Public Databases. J. Comput. Neurosci. 2006, 20, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.J. Dendritic Morphology, Inward Rectification, and the Functional Properties of Neostriatal Neurons. In Single Neuron Computation; Academic Press: San Diego, CA, USA, 1992; pp. 141–171. [Google Scholar] [CrossRef]
- Ogata, N.; Tatebayashi, H. Sodium Current Kinetics in Freshly Isolated Neostriatal Neurones of the Adult Guinea Pig. Pflugers Arch. Eur. J. Physiol. 1990, 416, 594–603. [Google Scholar] [CrossRef]
- Tkatch, T.; Baranauskas, G.; Surmeier, D.J. Kv4.2 mRNA Abundance and A-Type K(+) Current Amplitude Are Linearly Related in Basal Ganglia and Basal Forebrain Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Hernandez-Lopez, S.; Tkatch, T.; Held, J.E.; Surmeier, D.J. Kv1.2-Containing K+ Channels Regulate Subthreshold Excitability of Striatal Medium Spiny Neurons. J. Neurophysiol. 2004, 91, 1337–1349. [Google Scholar] [CrossRef]
- Steephen, J.E.; Manchanda, R. Differences in Biophysical Properties of Nucleus Accumbens Medium Spiny Neurons Emerging from Inactivation of Inward Rectifying Potassium Currents. J. Comput. Neurosci. 2009, 27, 453–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisenbaum, E.S.; Wilson, C.J. Potassium Currents Responsible for Inward and Outward Rectification in Rat Neostriatal Spiny Projection Neurons. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 4449–4463. [Google Scholar] [CrossRef] [Green Version]
- Berkefeld, H.; Sailer, C.A.; Bildl, W.; Rohde, V.; Thumfart, J.O.; Eble, S.; Klugbauer, N.; Reisinger, E.; Bischofberger, J.; Oliver, D.; et al. BKCa-Cav Channel Complexes Mediate Rapid and Localized Ca2+-Activated K+ Signaling. Science 2006, 314, 615–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maylie, J.; Bond, C.T.; Herson, P.S.; Lee, W.S.; Adelman, J.P. Small Conductance Ca2+-Activated K+ Channels and Calmodulin. J. Physiol. 2004, 554, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Brevi, S.; de Curtis, M.; Magistretti, J. Pharmacological and Biophysical Characterization of Voltage-Gated Calcium Currents in the Endopiriform Nucleus of the Guinea Pig. J. Neurophysiol. 2001, 85, 2076–2087. [Google Scholar] [CrossRef] [Green Version]
- Foehring, R.C.; Mermelstein, P.G.; Song, W.J.; Ulrich, S.; Surmeier, D.J. Unique Properties of R-Type Calcium Currents in Neocortical and Neostriatal Neurons. J. Neurophysiol. 2000, 84, 2225–2236. [Google Scholar] [CrossRef] [PubMed]
- Bargas, J.; Howe, A.; Eberwine, J.; Cao, Y.; Surmeier, D.J. Cellular and Molecular Characterization of Ca2+ Currents in Acutely Isolated, Adult Rat Neostriatal Neurons. J. Neurosci. 1994, 14, 6667–6686. [Google Scholar] [CrossRef]
- Kasai, H.; Neher, E. Dihydropyridine-Sensitive and Omega-Conotoxin-Sensitive Calcium Channels in a Mammalian Neuroblastoma-Glioma Cell Line. J. Physiol. 1992, 448, 161–188. [Google Scholar] [CrossRef] [Green Version]
- McNaughton, N.C.L.; Randall, A.D. Electrophysiological Properties of the Human N-Type Ca2+ Channel: I. Channel Gating in Ca2+ Ba2+ and Sr2+ Containing Solutions. Neuropharmacology 1997, 36, 895–915. [Google Scholar] [CrossRef]
- Tuckwell, H.C. Quantitative Aspects of L-Type Ca2+ Currents. Prog. Neurobiol. 2012, 96, 1–31. [Google Scholar] [CrossRef]
- McRory, J.E.; Santi, C.M.; Hamming, K.S.C.; Mezeyova, J.; Sutton, K.G.; Baillie, D.L.; Stea, A.; Snutch, T.P. Molecular and Functional Characterization of a Family of Rat Brain T-Type Calcium Channels. J. Biol. Chem. 2001, 276, 3999–4011. [Google Scholar] [CrossRef] [Green Version]
- Song, S.C.; Beatty, J.A.; Wilson, C.J. The Ionic Mechanism of Membrane Potential Oscillations and Membrane Resonance in Striatal LTS Interneurons. J. Neurophysiol. 2016, 116, 1752–1764. [Google Scholar] [CrossRef] [Green Version]
- Pifferi, S.; Dibattista, M.; Menini, A. TMEM16B Induces Chloride Currents Activated by Calcium in Mammalian Cells. PflÜgers Arch. Eur. J. Physiol. 2009, 458, 1023–1038. [Google Scholar] [CrossRef] [PubMed]
- Jȩdrzejewski-Szmek, Z.; Abrahao, K.P.; Jȩdrzejewska-Szmek, J.; Lovinger, D.M.; Blackwell, K.T. Parameter Optimization Using Covariance Matrix Adaptation—Evolutionary Strategy (CMA-ES), an Approach to Investigate Differences in Channel Properties between Neuron Subtypes. Front. Neuroinform. 2018, 12, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, N.; Akimoto, Y.; Baudis, P. CMA-ES/Pycma on Github. Zenodo 2019. [Google Scholar] [CrossRef]
- Sivagnanam, S.; Majumdar, A.; Yoshimoto, K.; Astakhov, V.; Bandrowski, A.E.; Martone, M.E.; Carnevale, N.T. Introducing the Neuroscience Gateway. IWSG 2013, 993, 0. [Google Scholar]
- Anwar, H.; Roome, C.J.; Nedelescu, H.; Chen, W.; Kuhn, B.; De Schutter, E. Dendritic Diameters Affect the Spatial Variability of Intracellular Calcium Dynamics in Computer Models. Front. Cell. Neurosci. 2014, 8, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, E.A.; Schoch, S.; Dietrich, D. Tuning Local Calcium Availability: Cell-Type-Specific Immobile Calcium Buffer Capacity in Hippocampal Neurons. J. Neurosci. 2013, 33, 14431–14445. [Google Scholar] [CrossRef] [Green Version]
- Matthews, E.A.; Dietrich, D. Buffer Mobility and the Regulation of Neuronal Calcium Domains. Front. Cell. Neurosci. 2015, 9, 48. [Google Scholar] [CrossRef]
- Carter, A.G.; Sabatini, B.L. State-Dependent Calcium Signaling in Dendritic Spines of Striatal Medium Spiny Neurons. Neuron 2004, 44, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.G.; Soler-Llavina, G.J.; Sabatini, B.L. Timing and Location of Synaptic Inputs Determine Modes of Subthreshold Integration in Striatal Medium Spiny Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 8967–8977. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Chen, T.W.; Guo, Z.V.; Gerfen, C.R.; Svoboda, K. A Motor Cortex Circuit for Motor Planning and Movement. Nature 2015, 519, 51–56. [Google Scholar] [CrossRef]
- Owen, S.F.; Berke, J.D.; Kreitzer, A.C. Fast-spiking interneurons supply feedforward control of bursting, calcium, and plasticity for efficient learning. Cell 2018, 172, 683–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharott, A.; Doig, N.M.; Mallet, N.; Magill, P.J. Relationships between the firing of identified striatal interneurons and spontaneous and driven cortical activities in vivo. J. Neurosci. 2012, 32, 13221–13236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, O.; Pillow, J.W.; Rust, N.C.; Simoncelli, E.P. Spike-Triggered Neural Characterization. J. Vis. 2006, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prager, E.M.; Dorman, D.B.; Hobel, Z.B.; Malgady, J.M.; Blackwell, K.T.; Plotkin, J.L. Dopamine Oppositely Modulates State Transitions in Striosome and Matrix Direct Pathway Striatal Spiny Neurons. Neuron 2020, 108, 1091–1102.e5. [Google Scholar] [CrossRef] [PubMed]
- Shouval, H.Z.; Wang, S.S.H.; Wittenberg, G.M. Spike timing dependent plasticity: A consequence of more fundamental learning rules. Front. Comput. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, A.; Israely, I.; Huang, S.Y.; Tonegawa, S. The Dendritic Branch Is the Preferred Integrative Unit for Protein Synthesis-Dependent LTP. Neuron 2011, 69, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Legenstein, R.; Maass, W. Branch-Specific Plasticity Enables Self-Organization of Nonlinear Computation in Single Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 10787–10802. [Google Scholar] [CrossRef] [Green Version]
- Cichon, J.; Gan, W.B. Branch-Specific Dendritic Ca2+ Spikes Cause Persistent Synaptic Plasticity. Nature 2015, 520, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Magó, Á.; Weber, J.P.; Ujfalussy, B.B.; Makara, J.K. Synaptic Plasticity Depends on the Fine-Scale Input Pattern in Thin Dendrites of CA1 Pyramidal Neurons. J. Neurosci. 2020, 40, 2593–2605. [Google Scholar] [CrossRef] [Green Version]
- Losonczy, A.; Makara, J.K.; Magee, J.C. Compartmentalized Dendritic Plasticity and Input Feature Storage in Neurons. Nature 2008, 452, 436–441. [Google Scholar] [CrossRef]
- Du, K.; Wu, Y.W.; Lindroos, R.; Liu, Y.; Rózsa, B.; Katona, G.; Ding, J.B.; Kotaleski, J.H. Cell-Type–Specific Inhibition of the Dendritic Plateau Potential in Striatal Spiny Projection Neurons. Proc. Natl. Acad. Sci. USA 2017, 114, E7612–E7621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, U.; Polsky, A.; Schiller, J. Plasticity Compartments in Basal Dendrites of Neocortical Pyramidal Neurons. J. Neurosci. 2006, 26, 12717–12726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkum, M.E.; Nevian, T.; Sandler, M.; Polsky, A.; Schiller, J. Synaptic Integration in Tuft Dendrites of Layer 5 Pyramidal Neurons: A New Unifying Principle. Science 2009, 325, 756–760. [Google Scholar] [CrossRef] [Green Version]
- Makara, J.K.; Magee, J.C. Variable Dendritic Integration in Hippocampal CA3 Pyramidal Neurons. Neuron 2013, 80, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, J.; Major, G.; Koester, H.J.; Schiller, Y. NMDA Spikes in Basal Dendrites of Cortical Pyramidal Neurons. Nature 2000, 404, 285–289. [Google Scholar] [CrossRef]
- Tazerart, S.; Mitchell, D.E.; Miranda-Rottmann, S.; Araya, R. A spike-timing-dependent plasticity rule for dendritic spines. Nat. Commun. 2020, 11, 4276. [Google Scholar] [CrossRef]
- Takahashi, N.; Kitamura, K.; Matsuo, N.; Mayford, M.; Kano, M.; Matsuki, N.; Ikegaya, Y. Locally Synchronized Synaptic Inputs. Science 2012, 335, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.E.; Whitney, D.E.; Scholl, B.; Fitzpatrick, D. Orientation Selectivity and the Functional Clustering of Synaptic Inputs in Primary Visual Cortex. Nat. Neurosci. 2016, 19, 1003–1009. [Google Scholar] [CrossRef]
- Kerlin, A.; Mohar, B.; Flickinger, D.; MacLennan, B.J.; Davis, C.; Spruston, N.; Svoboda, K. Functional Clustering of Dendritic Activity during Decision-Making. bioRxiv 2018, 440396. [Google Scholar] [CrossRef]
- Hwang, F.J.; Roth, R.H.; Wu, Y.W.; Sun, Y.; Kwon, D.K.; Liu, Y.; Ding, J.B. Motor learning selectively strengthens cortical and striatal synapses of motor engram neurons. Neuron 2022, 110, 1–12. [Google Scholar] [CrossRef]
- Ujfalussy, B.B.; Makara, J.K. Impact of Functional Synapse Clusters on Neuronal Response Selectivity. Nat. Commun. 2020, 11, 1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huertas, M.A.; Newton, A.J.; McDougal, R.A.; Sacktor, T.C.; Shouval, H.Z. Conditions for synaptic specificity during the maintenance phase of synaptic plasticity. Eneuro 2022, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.C.; Parajuli, L.K.; Zito, K. Heterosynaptic Structural Plasticity on Local Dendritic Segments of Hippocampal CA1 Neurons. Cell Rep. 2015, 10, 162–169. [Google Scholar] [CrossRef] [Green Version]
- El-Boustani, S.; Ip, J.P.K.; Breton-Provencher, V.; Knott, G.W.; Okuno, H.; Bito, H.; Sur, M. Locally Coordinated Synaptic Plasticity of Visual Cortex Neurons in Vivo. Science 2018, 360, 1349–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; Prokin, I.; Mendes, A.; Berry, H.; Venance, L. Robustness of STDP to Spike Timing Jitter. Sci. Rep. 2018, 8, 8139. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.H.; Poole, B.; Maheswaranathan, N.; Dhawale, A.K.; Fisher, T.; Wilson, C.D.; Brann, D.H.; Trautmann, E.M.; Ryu, S.; Shusterman, R.; et al. Discovering Precise Temporal Patterns in Large-Scale Neural Recordings through Robust and Interpretable Time Warping. Neuron 2019, 105, 246–259. [Google Scholar] [CrossRef]
- Lappalainen, J.; Herpich, J.; MTetzlaff, C. A Theoretical Framework to Derive Simple, Firing-Rate-Dependent Mathematical Models of Synaptic Plasticity. Front. Comput. Neurosci. 2019, 13, 26. [Google Scholar] [CrossRef]
- Shindou, T.; Shindou, M.; Watanabe, S.; Wickens, J. A Silent Eligibility Trace Enables Dopamine-Dependent Synaptic Plasticity for Reinforcement Learning in the Mouse Striatum. Eur. J. Neurosci. 2019, 49, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Perez De La Mora, M.; Manger, P.; Narváez, M.; Beggiato, S.; Crespo-Ramírez, M.; Navarro, G.; Wydra, K.; Díaz-Cabiale, Z.; Rivera, A.; et al. Brain Dopamine Transmission in Health and Parkinson’s Disease: Modulation of Synaptic Transmission and Plasticity Through Volume Transmission and Dopamine Heteroreceptors. Front. Synaptic Neurosci. 2018, 10, 20. [Google Scholar] [CrossRef]
- Zoli, M.; Torri, C.; Ferrari, R.; Jansson, A.; Zini, I.; Fuxe, K.; Agnati, L.F. The Emergence of the Volume Transmission Concept. Brain Res. Rev. 1998, 26, 136–147. [Google Scholar] [CrossRef]
- Santos, F.J.; Oliveira, R.F.; Jin, X.; Costa, R.M. Corticostriatal Dynamics Encode the Refinement of Specific Behavioral Variability during Skill Learning. eLife 2015, 4, e09423. [Google Scholar] [CrossRef] [PubMed]
- Legenstein, R.; Naeger, C.; Maass, W. What Can a Neuron Learn with Spike-Timing-Dependent Plasticity? Neural Comput. 2005, 17, 2337–2382. [Google Scholar] [CrossRef] [PubMed]
- Berthet, P.; Lindahl, M.; Tully, P.J.; Hellgren-Kotaleski, J.; Lansner, A. Functional Relevance of Different Basal Ganglia Pathways Investigated in a Spiking Model with Reward Dependent Plasticity. Front. Neural Circuits 2016, 10, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunovan, K.; Vich, C.; Clapp, M.; Verstynen, T.; Rubin, J. Reward-driven changes in striatal pathway competition shape evidence evaluation in decision-making. PLoS Comput. Biol. 2019, 15, e1006998. [Google Scholar] [CrossRef] [PubMed] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorman, D.B.; Blackwell, K.T. Synaptic Plasticity Is Predicted by Spatiotemporal Firing Rate Patterns and Robust to In Vivo-like Variability. Biomolecules 2022, 12, 1402. https://doi.org/10.3390/biom12101402
Dorman DB, Blackwell KT. Synaptic Plasticity Is Predicted by Spatiotemporal Firing Rate Patterns and Robust to In Vivo-like Variability. Biomolecules. 2022; 12(10):1402. https://doi.org/10.3390/biom12101402
Chicago/Turabian StyleDorman, Daniel B., and Kim T. Blackwell. 2022. "Synaptic Plasticity Is Predicted by Spatiotemporal Firing Rate Patterns and Robust to In Vivo-like Variability" Biomolecules 12, no. 10: 1402. https://doi.org/10.3390/biom12101402