
Activation of Wnt/β-Catenin Signaling Involves 660 nm Laser Radiation on Epithelium and Modulates Lipid Metabolism


Abstract
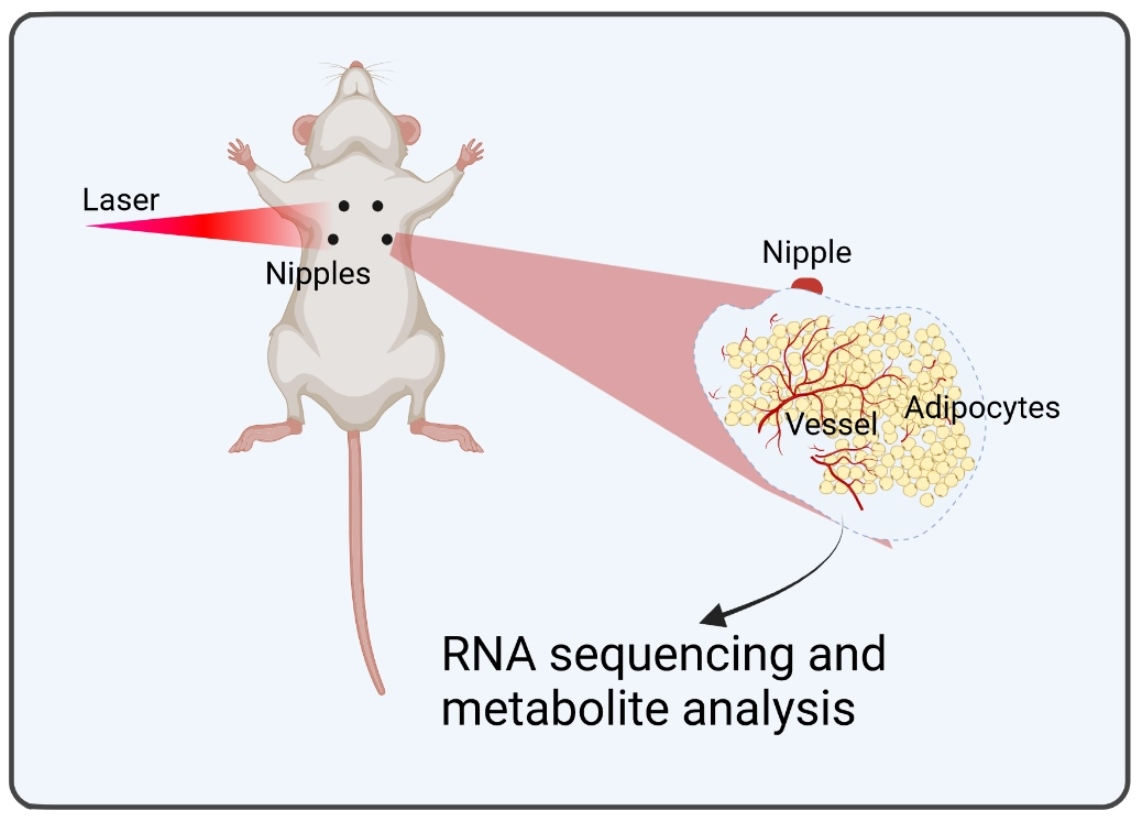
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| qRT-PCR | Real-time quantitative reverse transcription PCR |
| GO | Gene ontology |
| PPI | Protein–protein interaction |
| GSEA | Gene set enrichment analysis |
| DEGs | Differentially expressed genes |
| DAPI | 4′,6′-diamidino-2-phenylindole |
| LED | Light-emitting diode |
| NES | Normalized enrichment score |
| LC–MS | Liquid chromatography–mass spectrometry |
| PPI | Protein–protein interaction |
References
- Wajih, N.; Alipour, E.; Rigal, F.; Zhu, J.; Perlegas, A.; Caudell, D.L.; Kim-Shapiro, D. Effects of nitrite and far-red light on coagulation. Nitric Oxide Biol. Chem. 2021, 107, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, R.; Liu, C.; Liu, H.; Zhu, R.; Guo, S.; Tang, M.; Li, Y.; Niu, J.; Fu, M.; et al. Salvia miltiorrhiza: A Potential Red Light to the Development of Cardiovascular Diseases. Curr. Pharm. Des. 2017, 23, 1077–1097. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Deng, Y.; Huang, P. Application of red light therapy for moderate-to-severe acne vulgaris: A systematic review and meta-analysis. J. Cosmet. Dermatol. 2021, 20, 3498–3508. [Google Scholar] [CrossRef]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic therapy—Mechanisms, photosensitizers and combinations. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Álvarez, C.; Del Olmo-Aguado, S.; Merayo-Lloves, J.; Osborne, N.N. Near infra-red light attenuates corneal endothelial cell dysfunction in situ and in vitro. Exp. Eye Res. 2017, 161, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Cain, N.; Bartel, K.; Micic, G.; Maddock, B.; Gradisar, M. A randomised controlled trial of bright light therapy and morning activity for adolescents and young adults with Delayed Sleep-Wake Phase Disorder. Sleep Med. 2018, 45, 114–123. [Google Scholar] [CrossRef]
- Bashkatov, A.N.; Berezin, K.V.; Dvoretskiy, K.N.; Chernavina, M.L.; Genina, E.A.; Genin, V.D.; Kochubey, V.I.; Lazareva, E.N.; Pravdin, A.B.; Shvachkina, M.E.; et al. Measurement of tissue optical properties in the context of tissue optical clearing. J. Biomed. Opt. 2018, 23, 1–31. [Google Scholar] [CrossRef]
- El Nawam, H.; El Backly, R.; Zaky, A.; Abdallah, A. Low-level laser therapy affects dentinogenesis and angiogenesis of in vitro 3D cultures of dentin-pulp complex. Lasers Med. Sci. 2019, 34, 1689–1698. [Google Scholar] [CrossRef]
- das Neves, L.M.; Leite, G.P.; Marcolino, A.M.; Pinfildi, C.E.; Garcia, S.B.; de Araújo, J.E.; Guirro, E.C. Laser photobiomodulation (830 and 660 nm) in mast cells, VEGF, FGF, and CD34 of the musculocutaneous flap in rats submitted to nicotine. Lasers Med. Sci. 2017, 32, 335–341. [Google Scholar] [CrossRef]
- Wang, Z.X.; Kim, S.H. Effect of Photobiomodulation Therapy (660 nm) on Wound Healing of Rat Skin Infected by Staphylococcus. Photobiomodulation Photomed. Laser Surg. 2020, 38, 419–424. [Google Scholar] [CrossRef]
- Ebrahimpour-Malekshah, R.; Amini, A.; Zare, F.; Mostafavinia, A.; Davoody, S.; Deravi, N.; Rahmanian, M.; Hashemi, S.M.; Habibi, M.; Ghoreishi, S.K.; et al. Combined therapy of photobiomodulation and adipose-derived stem cells synergistically improve healing in an ischemic, infected and delayed healing wound model in rats with type 1 diabetes mellitus. BMJ Open Diabetes Res. Care 2020, 8, e001033. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, C.; Wei, T.; Gayet, O.; Loncle, C.; Borge, L.; Dusetti, N.; Ma, X.; Marson, D.; Laurini, E. Dendrimeric Nanosystem Consistently Circumvents Heterogeneous Drug Response and Resistance in Pancreatic Cancer. In Exploration; Wiley Online Library: Hoboken, NJ, USA, 2021; pp. 21–34. [Google Scholar]
- Bostanciklioglu, M.; Demiryürek, Ş.; Cengiz, B.; Demir, T.; Öztuzcu, S.; Aras, M.H.; Özsevik, S.; Usumez, A.; Ergün, S.; Özbal, H.K.; et al. Assessment of the effect of laser irradiations at different wavelengths (660, 810, 980, and 1064 nm) on autophagy in a rat model of mucositis. Lasers Med. Sci. 2015, 30, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, M.L.; Araújo-Filho, I.; da Silva, E.M.; de Sousa Queiroz, W.S.; Soares, C.D.; de Carvalho, M.G.; Maciel, M.A. Effect of low-level laser therapy on angiogenesis and matrix metalloproteinase-2 immunoexpression in wound repair. Lasers Med. Sci. 2017, 32, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, N.K.; Houreld, N.N.; Abrahamse, H. Photobiomodulation reduces oxidative stress in diabetic wounded fibroblast cells by inhibiting the FOXO1 signaling pathway. J. Cell Commun. Signal. 2021, 15, 195–206. [Google Scholar] [CrossRef]
- Lee, J.H.; Chiang, M.H.; Chen, P.H.; Ho, M.L.; Lee, H.E.; Wang, Y.H. Anti-inflammatory effects of low-level laser therapy on human periodontal ligament cells: In vitro study. Lasers Med. Sci. 2018, 33, 469–477. [Google Scholar] [CrossRef]
- Kaviani, A.; Fateh, M.; Ataie-Fashtami, L.; Yunesian, M.; Najafi, M.; Berry, M.; Rabbani, A. Comparison of carbon dioxide laser and scalpel for breast lumpectomy: A randomized controlled trial. Photomed. Laser Surg. 2008, 26, 257–262. [Google Scholar] [CrossRef]
- Trelles, M.A.; Pardo, L.; Chamorro, J.J.; Bonanad, E.; Allones, I.; Buil, C.; Luna, R. Erbium: YAG laser as a method of deepithelization in corrective and reductive breast surgery. Ann. Plast. Surg. 2005, 55, 122–126. [Google Scholar] [CrossRef]
- Ahcan, U.; Bracun, D.; Zivec, K.; Pavlic, R.; Butala, P. The use of 3D laser imaging and a new breast replica cast as a method to optimize autologous breast reconstruction after mastectomy. Breast 2012, 21, 183–189. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer treatment and survivorship statistics, 2022. CA A Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef]
- Katzenellenbogen, J.A.; Johnson, H.J., Jr.; Carlson, K.E.; Myers, H.N. Photoreactivity of some light-sensitive estrogen derivatives. Use of an exchange assay to determine their photointeraction with the rat uterine estrogen binding protein. Biochemistry 1974, 13, 2986–2994. [Google Scholar] [CrossRef]
- Liu, C.; Miyajima, T.; Melangath, G.; Miyai, T.; Vasanth, S.; Deshpande, N.; Kumar, V.; Ong Tone, S.; Gupta, R.; Zhu, S. Ultraviolet A light induces DNA damage and estrogen-DNA adducts in Fuchs endothelial corneal dystrophy causing females to be more affected. Proc. Natl. Acad. Sci. USA 2020, 117, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Rashid, O.M.; Nagahashi, M.; Ramachandran, S.; Dumur, C.; Schaum, J.; Yamada, A.; Terracina, K.P.; Milstien, S.; Spiegel, S.; Takabe, K. An improved syngeneic orthotopic murine model of human breast cancer progression. Breast Cancer Res. Treat. 2014, 147, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Pollack, J.S.; Kim, J.E.; Pulido, J.S.; Burke, J.M. Tissue effects of subclinical diode laser treatment of the retina. Arch. Ophthalmol. 1998, 116, 1633–1639. [Google Scholar] [CrossRef]
- Chuck, R.S.; Behrens, A.; Wellik, S.; Liaw, L.L.; Dolorico, A.M.; Sweet, P.; Chao, L.C.; Osann, K.E.; McDonnell, P.J.; Berns, M.W. Re-epithelialization in cornea organ culture after chemical burns and excimer laser treatment. Arch. Ophthalmol. 2001, 119, 1637–1642. [Google Scholar] [CrossRef]
- Jallow, F.; Brockman, J.L.; Helzer, K.T.; Rugowski, D.E.; Goffin, V.; Alarid, E.T.; Schuler, L.A. 17 β-Estradiol and ICI182, 780 Differentially Regulate STAT5 Isoforms in Female Mammary Epithelium, With Distinct Outcomes. J. Endocr. Soc. 2018, 2, 293–309. [Google Scholar] [CrossRef]
- Ma, R.; Gong, D.; You, H.; Xu, C.; Lu, Y.; Bergers, G.; Werb, Z.; Klein, O.D.; Petritsch, C.K.; Lu, P. LGL1 binds to Integrin β1 and inhibits downstream signaling to promote epithelial branching in the mammary gland. Cell Rep. 2022, 38, 110375. [Google Scholar] [CrossRef]
- Oren, M. Decision making by p53: Life, death and cancer. Cell Death Differ. 2003, 10, 431–442. [Google Scholar] [CrossRef]
- Chen, B.; Lan, J.; Xiao, Y.; Liu, P.; Guo, D.; Gu, Y.; Song, Y.; Zhong, Q.; Ma, D.; Lei, P. Long noncoding RNA TP53TG1 suppresses the growth and metastasis of hepatocellular carcinoma by regulating the PRDX4/β-catenin pathway. Cancer Lett. 2021, 513, 75–89. [Google Scholar] [CrossRef]
- Bautista-García, P.; Reyes, J.L.; Martín, D.; Namorado, M.C.; Chavez-Munguía, B.; Soria-Castro, E.; Huber, O.; González-Mariscal, L. Zona occludens-2 protects against podocyte dysfunction induced by ADR in mice. Am. J. Physiol.-Ren. Physiol. 2013, 304, F77–F87. [Google Scholar] [CrossRef] [Green Version]
- Kazanskaya, O.; Glinka, A.; del Barco Barrantes, I.; Stannek, P.; Niehrs, C.; Wu, W. R-Spondin2 is a secreted activator of Wnt/β-catenin signaling and is required for Xenopus myogenesis. Dev. Cell 2004, 7, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Sano, M.; Songyang, Z.; Schneider, M.D. A Wnt-and β-catenin-dependent pathway for mammalian cardiac myogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 5834–5839. [Google Scholar] [CrossRef] [PubMed]
- Hardie, R.C.; Minke, B. The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron 1992, 8, 643–651. [Google Scholar] [CrossRef]
- Golovynska, I.; Golovynskyi, S.; Stepanov, Y.V.; Stepanova, L.I.; Qu, J.; Ohulchanskyy, T.Y. Red and near-infrared light evokes Ca2+ influx, endoplasmic reticulum release and membrane depolarization in neurons and cancer cells. J. Photochem. Photobiol. B Biol. 2021, 214, 112088. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kou, S.; Gu, L.; Zheng, Q.; Li, M.; Qi, F.; Zhao, H.; Wang, L. Effects of Zuogui Pill () and Yougui Pill () on the expression of brain-derived neurotrophic factor and cyclic adenosine monophosphate/protein kinase A signaling transduction pathways of axonal regeneration in model rats with experimental autoimmune encephalomyelitis. Chin. J. Integr. Med. 2012, 20, 24–30. [Google Scholar]
- Gong, B.; Shen, W.; Xiao, W.; Meng, Y.; Meng, A.; Jia, S. The Sec14-like phosphatidylinositol transfer proteins Sec14l3/SEC14L2 act as GTPase proteins to mediate Wnt/Ca2+ signaling. eLife 2017, 6, e26362. [Google Scholar] [CrossRef]
- Sinha, S.; Yang, W. Cellular signaling for activation of Rho GTPase Cdc42. Cell. Signal. 2008, 20, 1927–1934. [Google Scholar] [CrossRef]
- Agarwal, R.; Jurisica, I.; Mills, G.B.; Cheng, K.W. The emerging role of the RAB25 small GTPase in cancer. Traffic 2009, 10, 1561–1568. [Google Scholar] [CrossRef]
- Jiang, C.; Li, H.T.; Zhou, Y.M.; Wang, X.; Wang, L.; Liu, Z.Q. Cardiac optogenetics: A novel approach to cardiovascular disease therapy. Ep Eur. 2018, 20, 1741–1749. [Google Scholar] [CrossRef]
- Mongardini, C.; Di Tanna, G.L.; Pilloni, A. Light-activated disinfection using a light-emitting diode lamp in the red spectrum: Clinical and microbiological short-term findings on periodontitis patients in maintenance. A randomized controlled split-mouth clinical trial. Lasers Med. Sci. 2014, 29, 1–8. [Google Scholar] [CrossRef]
- Grant, W.; MacRobert, A.; Bown, S.; Hopper, C.; Speight, P. Photodynamic therapy of oral cancer: Photosensitisation with systemic aminolaevulinic acid. Lancet 1993, 342, 147–148. [Google Scholar] [CrossRef]
- Banci, H.; Strazzi-Sahyon, H.; Duarte, M.; Cintra, L.; Gomes-Filho, J.; Chalub, L.; Berton, S.; de Oliveira, V.; Dos Santos, P.; Sivieri-Araujo, G. Influence of photodynamic therapy on bond strength and adhesive interface morphology of MTA based root canal sealer to different thirds of intraradicular dentin. Photodiagnosis Photodyn. Ther. 2020, 32, 102031. [Google Scholar] [CrossRef] [PubMed]
- Ziqi, W.; Kai, C.; Costabel, U.; Xiaoju, Z. Nanotechnology-Facilitated Vaccine Development During the Coronavirus Disease 2019 (COVID-19) Pandemic. In Exploration; Wiley Online Library: Hoboken, NJ, USA, 2022; p. 20210082. [Google Scholar]
- Niu, T.; Tian, Y.; Cai, Q.; Ren, Q.; Wei, L. Red light combined with blue light irradiation regulates proliferation and apoptosis in skin keratinocytes in combination with low concentrations of curcumin. PLoS ONE 2015, 10, e0138754. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.; Tian, Y.; Wang, G.; Guo, G.; Tong, Y.; Shi, Y. Inhibition of ROS-NF-κB-dependent autophagy enhances Hypocrellin A united LED red light-induced apoptosis in squamous carcinoma A431 cells. Cell. Signal. 2020, 69, 109550. [Google Scholar] [CrossRef]
- Song, S.; Zhang, Y.; Fong, C.-C.; Tsang, C.-H.; Yang, Z.; Yang, M. cDNA microarray analysis of gene expression profiles in human fibroblast cells irradiated with red light. J. Investig. Dermatol. 2003, 120, 849–857. [Google Scholar] [CrossRef]
- Zhang, J.; Yue, X.; Luo, H.; Jiang, W.; Mei, Y.; Ai, L.; Gao, G.; Wu, Y.; Yang, H.; An, J. Illumination with 630 nm red light reduces oxidative stress and restores memory by photo-activating catalase and formaldehyde dehydrogenase in SAMP8 mice. Antioxid. Redox Signal. 2019, 30, 1432–1449. [Google Scholar] [CrossRef]
- Ma, C.; Dai, S. Advances in photoreceptor-mediated signaling transduction in flowering time regulation. Chin. Bull. Bot. 2019, 54, 9. [Google Scholar]
- Jakobs, K.H.; Gierschik, P.; Grandt, R.; Marquetant, R.; Strasser, R.H. Signal Transduction by the Adenylate Cyclase System. In Signal Transduction and Protein Phosphorylation; Springer: Berlin/Heidelberg, Germany, 1987; pp. 55–63. [Google Scholar]
- Gemel, J.; Randall, D.D. Light regulation of leaf mitochondrial pyruvate dehydrogenase complex: Role of photorespiratory carbon metabolism. Plant Physiol. 1992, 100, 908–914. [Google Scholar] [CrossRef]
- Ferraresi, C.; Kaippert, B.; Avci, P.; Huang, Y.Y.; de Sousa, M.V.; Bagnato, V.S.; Parizotto, N.A.; Hamblin, M.R. Low-level laser (light) therapy increases mitochondrial membrane potential and ATP synthesis in C2C12 myotubes with a peak response at 3–6 h. Photochem. Photobiol. 2015, 91, 411–416. [Google Scholar] [CrossRef]
- Yamauchi, N.; Minagawa, E.; Imai, K.; Kobuchi, K.; Li, R.; Taguchi, Y.; Umeda, M. High-Intensity Red Light-Emitting Diode Irradiation Suppresses the Inflammatory Response of Human Periodontal Ligament Stem Cells by Promoting Intracellular ATP Synthesis. Life 2022, 12, 736. [Google Scholar] [CrossRef]
- Pastore, D.D.; Martino, C.D.; Bosco, G.; Passarella, S. Stimulation of ATP synthesis via oxidative phosphorylation in wheat mitochondria irradiated with helium-neon laser. IUBMB Life 1996, 39, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Li, K.; Hu, B.; Li, C.; Zhang, M.; Hussain, A.; Wang, X.; Cheng, Q.; Yang, F.; Ge, K. Membrane-Destabilizing Ionizable Lipid Empowered Imaging—Guided Sirna Delivery and Cancer Treatment. In Exploration; Wiley Online Library: Hoboken, NJ, USA, 2021; pp. 35–49. [Google Scholar]
- Plana-Bonamaisó, A.; López-Begines, S.; Fernández-Justel, D.; Junza, A.; Soler-Tapia, A.; Andilla, J.; Loza-Alvarez, P.; Rosa, J.L.; Miralles, E.; Casals, I. Post-translational regulation of retinal IMPDH1 in vivo to adjust GTP synthesis to illumination conditions. eLife 2020, 9, e56418. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Álvarez, C.; Suárez-Barrio, C.; Del Olmo Aguado, S.; Osborne, N.N. Blue light negatively affects the survival of ARPE19 cells through an action on their mitochondria and blunted by red light. Acta Ophthalmol. 2019, 97, e103–e115. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.N.; Núñez-Álvarez, C.; Del Olmo-Aguado, S.; Merrayo-Lloves, J. Visual light effects on mitochondria: The potential implications in relation to glaucoma. Mitochondrion 2017, 36, 29–35. [Google Scholar] [CrossRef] [PubMed]
- del Olmo-Aguado, S.; Manso, A.G.; Osborne, N.N. Light might directly affect retinal ganglion cell mitochondria to potentially influence function. Photochem. Photobiol. 2012, 88, 1346–1355. [Google Scholar] [CrossRef]
- Ming, M.; Wang, S.; Wu, W.; Senyuk, V.; Le Beau, M.M.; Nucifora, G.; Qian, Z. Activation of Wnt/β-catenin protein signaling induces mitochondria-mediated apoptosis in hematopoietic progenitor cells. J. Biol. Chem. 2012, 287, 22683–22690. [Google Scholar] [CrossRef]
- Shares, B.H.; Busch, M.; White, N.; Shum, L.; Eliseev, R.A. Active mitochondria support osteogenic differentiation by stimulating β-catenin acetylation. J. Biol. Chem. 2018, 293, 16019–16027. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front. Immunol. 2018, 9, 745. [Google Scholar] [CrossRef]
- Perugorria, M.J.; Olaizola, P.; Labiano, I.; Esparza-Baquer, A.; Marzioni, M.; Marin, J.J.G.; Bujanda, L.; Banales, J.M. Wnt-β-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 121–136. [Google Scholar] [CrossRef]
- Alonso, L.; Fuchs, E. Stem cells in the skin: Waste not, Wnt not. Genes Dev. 2003, 17, 1189–1200. [Google Scholar] [CrossRef]
- Alberici, P.; Fodde, R. The role of the APC tumor suppressor in chromosomal instability. Genome Dyn. 2006, 1, 149–170. [Google Scholar]
- Okerlund, N.D.; Cheyette, B.N. Synaptic Wnt signaling-a contributor to major psychiatric disorders? J. Neurodev. Disord. 2011, 3, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Schinner, S. Wnt-signalling and the metabolic syndrome. Horm. Metab. Res. Horm. Und Stoffwechs. Horm. Et Metab. 2009, 41, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Chang, Z.F.; Chau, Y.P.; Chen, A.; Yang, W.C.; Yang, A.H.; Lee, O.K. Circulating Wnt/β-catenin signalling inhibitors and uraemic vascular calcifications. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc. Eur. Ren. Assoc. 2015, 30, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Benz, F.; Wichitnaowarat, V.; Lehmann, M.; Germano, R.F.; Mihova, D.; Macas, J.; Adams, R.H.; Taketo, M.M.; Plate, K.H.; Guérit, S.; et al. Low wnt/β-catenin signaling determines leaky vessels in the subfornical organ and affects water homeostasis in mice. eLife 2019, 8, e43818. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.P.; Pan, J.; Li, Y.; Geng, J.; Liu, C.; Zhang, L.Y.; Zhou, P.; Tang, Y.H.; Wang, Y.; Zhang, Z.; et al. Oligodendrocyte precursor cell transplantation promotes angiogenesis and remyelination via Wnt/β-catenin pathway in a mouse model of middle cerebral artery occlusion. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2021, 42, 757–770. [Google Scholar] [CrossRef]
- Qiao, N.; Du, G.; Zhong, X.; Sun, X. Recombinant Lactic Acid Bacteria as Promising Vectors for Mucosal Vaccination. In Exploration; Wiley Online Library: Hoboken, NJ, USA, 2021; p. 20210026. [Google Scholar]
- Cha, B.; Geng, X.; Mahamud, M.R.; Zhang, J.Y.; Chen, L.; Kim, W.; Jho, E.H.; Kim, Y.; Choi, D.; Dixon, J.B.; et al. Complementary Wnt Sources Regulate Lymphatic Vascular Development via PROX1-Dependent Wnt/β-Catenin Signaling. Cell Rep. 2018, 25, 571–584.e5. [Google Scholar] [CrossRef]
- Fang, Q.; Kaeli, D.R. Accelerating mesh-based Monte Carlo method on modern CPU architectures. Biomed. Opt. Express 2012, 3, 3223–3230. [Google Scholar] [CrossRef]
- Dogdas, B.; Stout, D.; Chatziioannou, A.F.; Leahy, R.M. Digimouse: A 3D whole body mouse atlas from CT and cryosection data. Phys. Med. Biol. 2007, 52, 577. [Google Scholar] [CrossRef]
- Cheong, W.-F.; Prahl, S.A.; Welch, A.J. A review of the optical properties of biological tissues. IEEE J. Quantum Electron. 1990, 26, 2166–2185. [Google Scholar] [CrossRef]
- Strangman, G.; Franceschini, M.A.; Boas, D.A. Factors affecting the accuracy of near-infrared spectroscopy concentration calculations for focal changes in oxygenation parameters. Neuroimage 2003, 18, 865–879. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Q.; Wang, L.; Zhang, J.; Zhong, X.; Guo, Z.; Yu, J.; Ma, Y.; Wu, H. Activation of Wnt/β-Catenin Signaling Involves 660 nm Laser Radiation on Epithelium and Modulates Lipid Metabolism. Biomolecules 2022, 12, 1389. https://doi.org/10.3390/biom12101389
Xiao Q, Wang L, Zhang J, Zhong X, Guo Z, Yu J, Ma Y, Wu H. Activation of Wnt/β-Catenin Signaling Involves 660 nm Laser Radiation on Epithelium and Modulates Lipid Metabolism. Biomolecules. 2022; 12(10):1389. https://doi.org/10.3390/biom12101389
Chicago/Turabian StyleXiao, Qiyang, Lijing Wang, Juling Zhang, Xinyu Zhong, Zhou Guo, Jiahao Yu, Yuanyuan Ma, and Haigang Wu. 2022. "Activation of Wnt/β-Catenin Signaling Involves 660 nm Laser Radiation on Epithelium and Modulates Lipid Metabolism" Biomolecules 12, no. 10: 1389. https://doi.org/10.3390/biom12101389