1. Introduction
Relaxin 2 (RLX2) is a two-chain peptide hormone that has a similar structure to insulin. It is one of the peptides of the so called “insulin-relaxin peptide family” that includes insulin, insulin-like growth factors, relaxins (1, 2, and 3), and relaxin/insulin-like factors (3, 4, 5, and 6) [
1]. RLX1, RXL2, and RLX3 differ in their origins and amino-acid sequence [
1]. RLX2 is currently considered a pleiotropic hormone, and it has important functions in the brain, heart, and kidney, in addition to its role in nitric oxide regulation and neo-angiogenesis [
1,
2]. RLX2′s therapeutic potential has recently been connected with its ability to prevent the tissue remodeling observed in fibrosis and to conserve endogenous tissue structure [
3], while it has been demonstrated to contribute to bone remodeling/expansion [
4]. Furthermore, recombinant human relaxin 2 is currently being clinically evaluated for treating acute heart failure [
5].
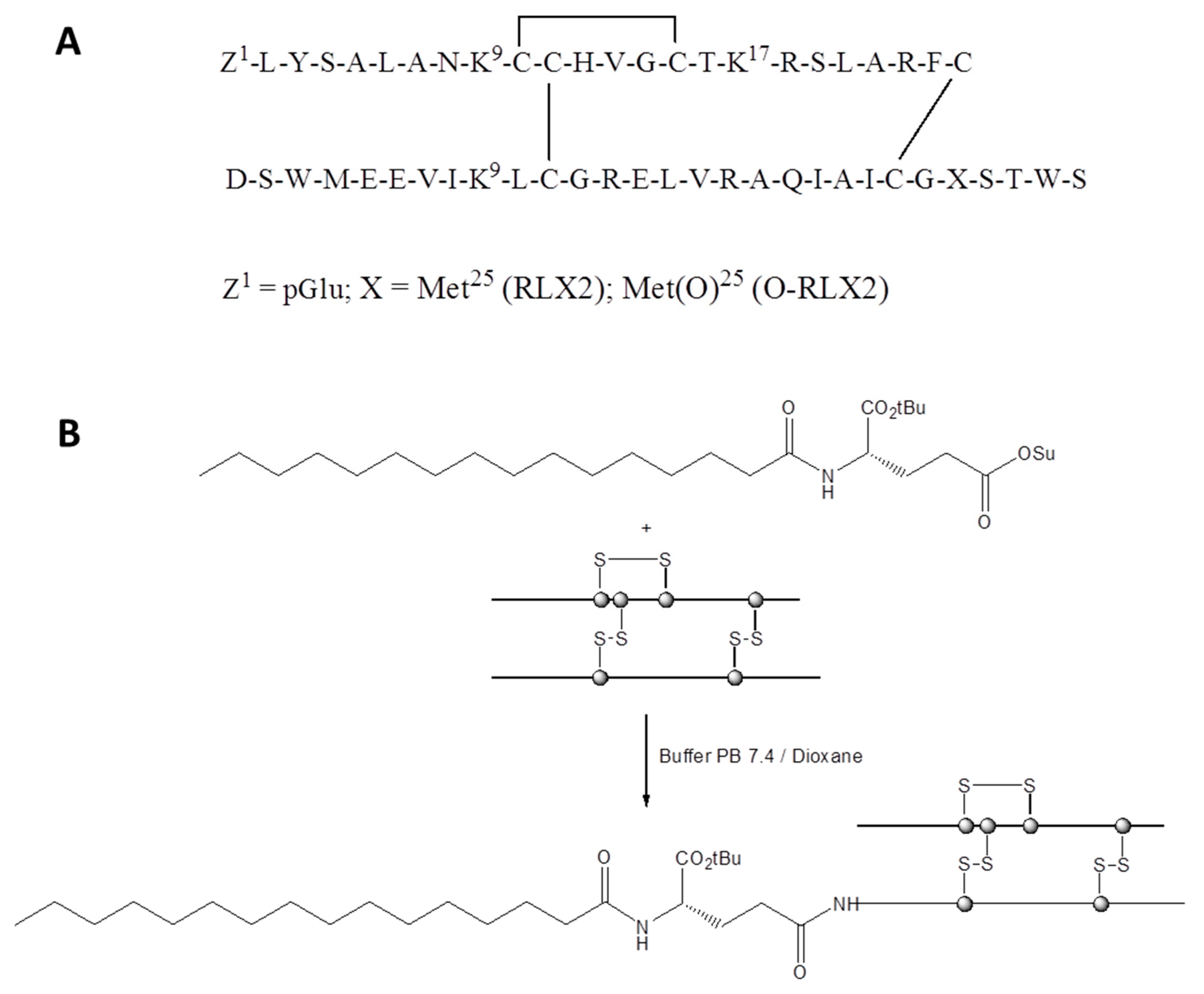
Chemically, RLX2 consists of two peptide chains, the A-chain and the B-chain, that are linked by two cysteine bridges [
6,
7]. The chemical synthesis of RLX2 was not achieved until recently, due to the low solubility of the B-chain, and the low yield achieved when combining the A and B chains. The solubility of RLX2 B-chain was markedly increased by oxidation of its Met
25, and thus effective solid-phase synthesis was recently achieved [
8]. Chemical synthesis of RLX2 facilitates its use as a therapeutic; however, the intrinsic limitations of peptides have impeded the development of peptide drugs, including RLX2. The two major constraints on peptide drug development are their poor oral bioavailability and short half-life in the bloodstream, due to serum enzyme susceptibility and rapid renal clearance [
9]. In particular, RLX2 has a very short in vivo half-life of approximately 10 min in rabbits and monkeys [
10], and also in women [
11]. For this reason, it was necessary to administer RLX2 continuously for 48 h by intravenous infusion into patients, in clinical studies (Phase III trials) that investigated the capability of RLX2 as a therapeutic for treating acute heart failure [
5]. Therefore, there is a clear need for improving the pharmacokinetic properties of RLX2, in order to potentially improve its therapeutic value. Numerous chemical approaches to address this concern have been undertaken in recent years. Conjugating the peptide to an inert chemical moiety is one of the strategies repeatedly proven to be successful in extending the half-life of various peptide therapeutics [
9]. More specifically, a meta-analysis of studies that involved fatty acid conjugates of peptides indicated that acylation contributed to a statistically significant extension of the half-life of the peptides [
12]. The replacement of disulfide bonds with non-reducible elements has been demonstrated to be effective and to eliminate the relaxin-specific deleterious effect of serum reductases. In particular, substitution with dicarba bonds via ring closure metathesis has been increasingly applied to many bioactive cystine-rich peptides [
13].
An insulin dimer was recently demonstrated to be thermodynamically stable in vitro [
14]. It was thus proposed that perhaps the dimeric form of RLX2 may also be more stable, compared to its monomeric form. However, although dicarba peptide analogues of RLX2 were found to retain strong receptor activity, their stability in serum was drastically reduced [
13]. Nevertheless, other synthetic covalently linked dimeric forms of RLX2 were seen to retain their receptor binding activity and additionally demonstrated improved in vitro serum stability [
15].
Another possibility for the enhancement and prolongation of peptide stability is to encapsulate them into nanometer sized vesicles, as a method to protect them from denaturation (proteolysis and dilution effects). For this reason, several encapsulation methodologies have been optimized to achieve a high encapsulation of chemically unstable molecules, but in the end they could not preserve their functionality [
16,
17]. Elsewhere, it was reported that the liposome preparation method that denatures the least protein is the lipid film hydration technique, but a drawback of this method is that the encapsulation efficiency achieved is low [
18,
19]. Previously, using a modified film hydration method, it was demonstrated that 40% of initial acetylcholinesterase (AChE) could be encapsulated in a functional state in liposomes, but the encapsulated enzyme had reduced activity, due to reduced substrate permeation through the lipid membrane. When the permeability of the liposome membrane was modified by a porin, full function of the enzyme was recovered [
20].
Herein, we evaluated and compared both methods discussed above, for the first time, for their ability to protect RLX2 peptides from degradation and loss of biological activity, in vitro. In particular, we synthesized a novel palmitoyl conjugate of RLX2 peptide (P-RLX2), using the Palmitoyl-
l-Glu-OtBu peptide modifier. This peptide modifier is known to effectively extend the peptide shelf-life of Liraglutide (Victoza), by delaying its absorption and reducing its renal clearance, due to the shielding effect of the fatty acid moiety [
21]. Additionally, we developed novel types of liposomes that encapsulate RLX2, as well as its oxidized form (O-RLX2) and the P-RLX2 conjugate. For liposome formation, the dehydration-rehydration vesicle (DRV) method was applied; a mild method that protects the integrity of peptide/protein drugs and confers high encapsulation for aqueous soluble materials, as reported in [
22,
23].
2. Materials and Methods
2.1. Materials
Egg phosphatidylocholine (PC), 1,2-Distearoyl-sn-glycerol-3-phosphatidyl ethanolamine N-[methoxy (polyethylene-glycol)-2000] (PEG), and Phosphatidyl-glycerine (PG) were purchased from Lipoid, Germany. Cholesterol (Chol) was purchased from Sigma-Aldrich (Darmstadt, Germany). Human Relaxin 2 (RLX2) and Oxidized Human Relaxin 2 (O-RLX2), as well as N-Palmitoyl-l-glutamic acid α-tert-butyl ester γ-succinimidyl ester (Palmitoyl-l-Glu(OSu)-OtBu), were kindly supplied by CBL (Patras, Greece). Agar was from Sigma-Aldrich (Darmstadt, Germany). All solvents used were of analytical or HPLC grade and purchased from Merck (Darmstadt, Germany). All other materials, such as the salts used for buffer preparation, reagents for lipid concentration determination, etc., were of analytical grade and were purchased from Sigma Aldrich (Darmstadt, Germany).
2.2. Characterization (HLPC, ESI-MS)
HPLC analysis was performed on a Shimadzu LC-2010 liquid chromatography system (Canby, OR, USA) using a LiChrospher® 100 RP-18 (5 μm) LiChroCART® 250-4. Mobile phase: THF/H2O; Gradient: 20% THF to 100% THF in 30 min (Conditions A); 50% THF to 100% THF in 30 min (Conditions B); Flow rate: 1 mL/min; Detection at 280 nm. ESI-MS spectra were recorded on a Waters Micromass ZQ 4000 mass detector (positive mode), controlled by MassLynx 4.1 software, by direct infusion, using a syringe pump at a flow rate of 5 μL/min. Cone voltage was set at 30 V and scan time at 1 s, with the interscan delay at 0.1 s.
2.3. Synthesis and Analysis of Nα-(Palmitoyl-l-γ-Glutamyl-α-OtBu)-RLX2 (P-RLX2)
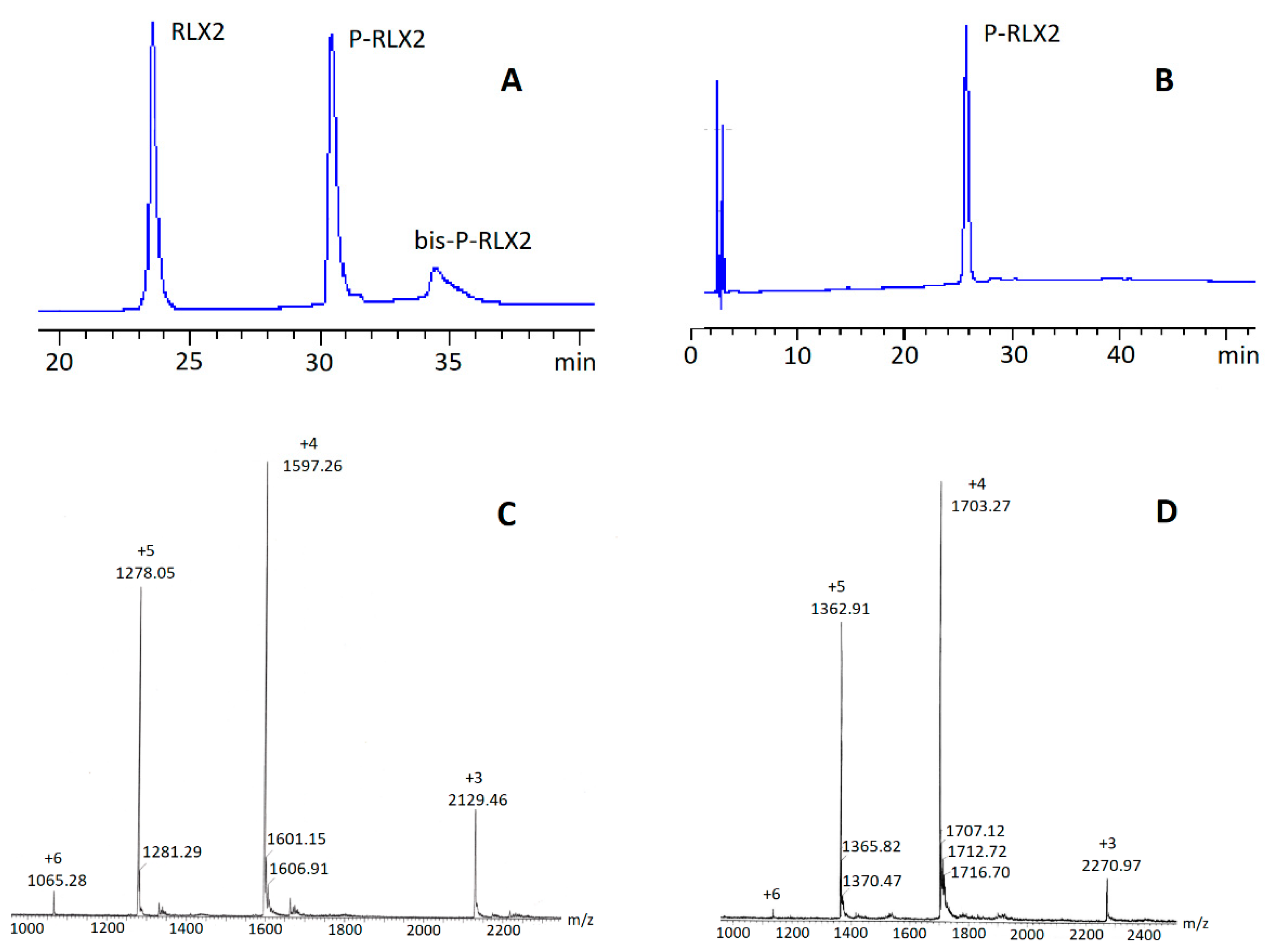
RLX2 (50 mg; 8.39 μmol) was dissolved in buffer phosphate PB at pH 7.40 (2 mL). To the resulting solution, Palmitoyl-l-Glu(OSu)-OtBu (18.08 mg; 4 eq) dissolved in DMSO or Dioxane (2 mL) was added dropwise at 4–5 °C while vigorously stirring, within 2 h, and the reaction mixture was further stirred for 12 h at 20 °C. Next, the reaction mixture was dialyzed over distilled water (3 times) and then lyophilized to afford a white powder that was further purified by semi-preparative HPLC. The collected fractions of P-RLX2 were combined and finally lyophilized to afford P-RLX2 as a white solid (yield 30–35% relative to RLX2). Purity: > 98% (determined by hplc analysis). ESI-MS [M+3H] calcd.: 2128.38; found: 2129.46; [M+4H] calcd.: 1596.54; found: 1597.26; [M+5H] calcd.: 1277.43; found: 1278.05; [M+6H] calcd.: 1064.69; found: 1065.28.
2.4. Preparation of RLX2 Peptide Liposomes
The lipid membrane compositions used for all the liposome formulations prepared were (i) PC/Chol (2:1 mole/mole), (ii) PC/PG/Chol (8:2:5 mole/mole/mole), and (iii) PC/PG/Chol/PEG (8:2:5:1.3 mole/mole/mole/mole). Details about liposome preparation using the two different methods applied are mentioned below.
2.4.1. Thin-Film Hydration Method
The thin film hydration method was used for P-RLX2 liposome preparation, as previously described in detail [
24]. In brief, the lipids required for lipid composition of PC/PG/Chol/PEG (8:2:5:1.3, mol/mol), together with the required amount of P-RLX2 (to give a P-RLX2/lipid molar ratio of 0.003, and a total lipid concentration of 10 mg/mL), were dissolved in a chloroform/methanol (2:1
v/
v) mixture. The mixture was then placed in a round-bottom flask and connected to a rotor evaporator (BÜCHI Labortechnik AG, Postfach, Switzerland) under vacuum, until complete evaporation of the organic solvents and the formation of a thin lipidic film on the sides of the flask. The film was hydrated with 1 mL of PBS at 40 °C and vortexed until complete removal of the lipids from the sides of the flask, for multilamellar vesicle (MLV) formation.
2.4.2. Dehydration-Rehydration Vesicle Method
For dehydration–rehydration vesicle (DRV) preparation, empty small unilamellar vesicles (SUV) were initially prepared as described in detail before, from empty MLV liposomes [
22,
23,
24]. For this, the lipids were dissolved in a chloroform/methanol (2:1
v/
v) mixture and stored at −20 °C. For each liposome preparation, the appropriate amounts of each solution (depending on the lipid composition required), for a final lipid concentration of 20 mg/mL, were placed in a 100-mL round-bottomed flask. Organic solvents were evaporated by the connection of the flask to a rotary evaporator, until a thin film was formed. For complete removal of organic solvents, the film was flashed-dried with nitrogen for 2–3 min. The lipid film was hydrated with 1 mL of diluted PBS buffer (10%
v/
v) pH 7.40, heated at 40 °C. The resulting dispersions consisting of MLVs were subsequently converted into SUVs by probe sonication with a tapered micro-tip (Vibra cell, Sonics and Materials, Suffolk, UK). In all cases, the initially turbid liposomal suspension was well clarified after sonication for 5–15 min. Following sonication, the liposome suspensions (SUV) were left to stand for at least 1 h at 40 °C, in order to anneal any structural defects. Any Ti-fragment and/lipid aggregate contaminants were removed from SUV suspensions by centrifugation at 15,000×
g for 20 min. Then, 1 mL of the SUV suspension was mixed with 1 mL of a 0.5 mg/mL RLX2 or O-RLX2 solution (in distilled H
2O), and the mixture was freeze-dried. With controlled rehydration of the dried materials, as described previously [
22,
23], multilamellar DRVs were generated.
2.4.3. Liposome (MLV and DRV) Size Reduction and Purification
Size reductions of liposomes (both MLVs and DRVs) were carried out by sequential extrusion of the liposomal dispersions at least ten times, through polycarbonate filters with a pore diameter 0.4 µm, followed by a second extrusion cycle through membranes with a 0.1 µm pore diameter, fitted in a syringe-type extruder (Lipo-so-fast, Avestin, Ottawa, ON, Canada). Extrusion was used as a size-reduction method, since sonication was avoided, in order to prevent disruption of the DRV liposomes and leakage of the encapsulated drug and/or destruction of the encapsulated protein. After size reduction, all liposomes were purified from non-entrapped (or incorporated) peptides by size exclusion chromatography on a Sepharose CL-4B column (1 × 40 cm), eluted with PBS, pH 7.40.
2.4.4. Post-Pegylation of Liposomes
For preparation of PEGylated liposomes, a post-pegylation technique [
25] was used, since it was found that the protein encapsulation in DRV liposomes was low when PEG-lipid was added in the lipid phase during DRV preparation. For this, PEG-lipid micellar dispersion was prepared using the thin film hydration method at a final concentration of 11.88 mg/mL of PEG-lipid in PBS, followed by an annealing step at 40 °C for 30 min. Next, a PEG dispersion (11.88 mg/mL) and liposomal dispersion (20 mg/mL), prepared as previously described by the DRV method, were mixed at a volume ratio 1:1,
v/
v and the mixture was incubated at 40 °C for 2 h. After post-pegylation, liposomes were purified from non-incorporated PEG, by size exclusion chromatography, as described above.
2.5. Physicochemical Characterization of RLX2 Peptide Liposomes
2.5.1. Encapsulation Efficiency (EE%)
Liposomes were characterized for peptide encapsulation efficiency (%), calculated according to Equation (1):
where D is drug (peptide) concentration and L is lipid concentration; initial means before purification and final after liposome purification. Liposome lipid concentration was measured using a Stewart assay [
26], a colorimetric method used for the quantification of phospholipids.
RLX2 and O-RLX2 concentration in liposomes was quantified by gradient high-performance liquid chromatography (HPLC) using a Shimadzu 20A5 Gradient HPLC system coupled to a SPD-20A Prominence UV/VIS detector operating at 220 nm. A Luna
® 5 µm C18 (2) 100 Å, LC Column (250 × 4.6 mm) was used; the mobile phase was a mixture of acidified water (0.1%
v/
v trifluoroacetic acid) and acetonitrile at 63:37
v/
v. The column was eluted at a flow rate of 1 mL/min at 25 °C, and RLX2 or O-RLX2 was eluted at 5.2 min and 5.4 min, respectively. The sample injection volume was 100 µL. A calibration curve in the range of 2.5–30 µg/mL was constructed by preparation of standard solutions of RLX2 or O-RLX2 in the presence of 1 mg/mL lipid (lipid composition PC/Chol or PC/PG/Chol) in media with a similar composition to the samples. Liposomes were analyzed after being lysed in isopropanol, for efficient protein extraction, as previously reported [
27,
28]. One volume of sample (150 μL) was mixed with one volume (150 μL) of isopropanol, and the mixture was agitated in a vortex, followed by the addition of 300 μL HEPES buffer (pH 5.0) at a stable final lipid concentration of 1 mg/mL. For the quantification of the peptides encapsulated in the liposomes of lipid composition PC/PG/Chol/PEG, a similar procedure, with a few modifications, was applied. In particular, a calibration curve was constructed in the range of 2.5–15 µg/mL, by preparation of standard solutions of RLX2 or O-RLX2 in the presence of 1 mg/mL lipid (lipid composition PC/PG/Chol/PEG) in media with similar composition as the samples in each case. Liposomes were analyzed after being lysed in isopropanol; for this, 100 μL of liposomal dispersion was mixed with 150 μL of isopropanol, and the mixture was agitated by a vortex, followed by the addition of 300 μL HEPES buffer (pH 5.0) at a stable final lipid concentration of 1 mg/mL.
Due to the low absorptivity of the conjugate P-RLX2, its concentration in the liposomes could not be measured by the HPLC method (as used for calculation of the yield of the synthetic procedure), and its complete incorporation in the liposomes was verified by HPLC studies carried out in the combined liposome and free molecule fractions collected, following size exclusion chromatography on a Sepharose CL-4B column (1 × 40 cm), eluted with PBS, pH 7.40.
2.5.2. Vesicle Physicochemical Properties
The particle size distribution (mean hydrodynamic diameter and polydispersity index) of RLX2, O-RLX2, and P-RLX2 loaded liposomes dispersed at 0.4 mg/mL lipid, in phosphate-buffered saline (10 mM) with pH 7.40, was measured by dynamic light scattering (DLS) (Malvern Nano-Zs, Malvern Instruments, Malvern, Worcestershire, UK) at 25 °C and a 173° angle [
24]. Each sample was measured 11 times in three independent measurements. The polydispersity index (PDI) was used as a measure of homogeneity of liposomal dispersions. Dispersions having a PDI of less than 0.200 or 0.250 are generally considered to have a narrow size distribution. Zeta potential was measured in the same dispersions, at 25 °C, utilizing the Doppler electrophoresis technique, as recently reported [
24].
2.5.3. RLX2 Peptide Stability Studies in Buffer and Serum (In Vitro)
To ensure peptide stability during incubation in media containing, or not containing, serum proteins, solutions of RLX2 peptides, as well as liposomal dispersions with lipid membrane compositions of PC/PG/Chol (8:2:5 mol/mol/mol) and PC/PG/Chol/PEG (8:2:5:1.3 mol/mol/mol/mol), were incubated at a final peptide concentration of 35 ± 8.1 μg/mL (when PBS was used as media), and 58 ± 5.0 μg/mL (when FBS (80% v/v) was used as media). Samples were placed separately in hermetically sealed screw-tubes (to avoid evaporation) and then in an orbital incubator (Stuart S1500, UK) set at 50 rpm and 37 °C, for a period of up to 30 d. At specific time intervals, samples were analyzed by HPLC, for quantification of intact RLX2 or O-RLX2, with the method mentioned above, in the case of samples in PBS. For quantification of free or liposomal peptides in the presence of 80% FBS, serum proteins precipitation was accomplished by mixing one volume of sample (100 μL) with three volumes (300 μL) of acidified isopropanol (with 0.1% TFA), followed by vortex agitation and centrifugation at 15000 rpm for 15 min. Next, the supernatant was transferred to a new eppendorf tube and 400 μL of HEPES (pH 5.0) were added, followed by vigorous vortex for 1 min, in order to retrieve the peptide. Finally, the samples were analyzed by HPLC, based on a calibration curve constructed at the exact same conditions as the samples to be analyzed.
For direct comparison of the stability of the different peptides, the results were normalized and expressed as the percentage of the corresponding initial peptide concentration measured at time point 0, for each peptide and each formulation (solution or liposome). Experiments were carried out in triplicate, and the results are expressed as the mean value ± standard deviation.
2.6. RLX2 Peptide Biological Activity Evaluation (In Vitro)
2.6.1. Cell culture, Transfection, and Stimulation for cAMP Production
Human HEK-293 embryonic kidney cells (HEK) (American Type Culture Collection, Manassas, VA, USA) were provided by Prof. G.T. Stathopoulos (Medical School, University of Patras, Patras, Greece). Cells were grown in Dulbecco’s modified Eagle’s medium (D-MEM; Gibco BRL, Gaithersburg, MD, USA) supplemented with 10% fetal bovine serum (FBS; Biosera, Nuaille, France), 100 IU penicillin/mL, and 100 μg streptomycin/mL (Gibco, Gran Iskand, NY, USA), and incubated at 37 °C and 5% CO
2. A total of 6 × 10
5 cells were seeded in 6-well plates and allowed to attach overnight. The following day, cells were transected with RXFP1 (LGR7, OriGene, Rockville, MD, USA) with Xfect transfection reagent (Takara, San Jose, CA, USA). After 48 h, RXFP1-HEK293 cells with transient expression were prepared for stimulation assay, with various receptor agonists. In more detail, cells were washed twice with Hanks’ balanced salt solution and were incubated in Hanks’ balanced salt solution in the presence of 0.1 mM 3-isobutyl-1-methylxanthine (IBMX, stimulation buffer, Sigma-Merck, Darmstadt, Germany) for 10 min. Cells were then stimulated for 30 min with vehicle or RLX2 peptide types at concentrations between 0.1–100 nM, media were aspirated, and 200 μL of HCL 0.1 M was added into each well, to extract cAMP. After 10 min, the extracts were collected and centrifuged at 600 g for 5 min and subsequently samples were used for Cyclic adenosine monophosphate (cAMP) assay [
29]. HEK293 cells without drug stimulation were employed as the control group.
2.6.2. Cyclic Adenosine Monophosphate (cAMP) Assay
The biological activity of RLX2 peptides was measured by their ability to stimulate cAMP production in RXFP1-HEK293 cells [
29]. The dose–response bioactivity of the three peptide types evaluated was measured at peptide concentrations 0.1, 1, 10, and 100 nM. cAMP concentration was measured according to the instructions of the cAMP Direct Immunoassay Kit (ab65355, Abcam, Cambridge, UK). For this, after carrying out the stimulation procedure as described above, and prior to quantification, cAMP standards and samples were neutralized and acetylated using the neutralizing buffer and acetylating reagent supplied in the kit, respectively. For quantification, 50 μL of standard cAMP solutions or 50 µL of samples were added to the Protein G coated 96-well plate provided. After blending with 10 μL cAMP antibody, the suspension was incubated for 1 h at room temperature with gentle agitation and for another 1 h after adding 10 mL of cAMP-HRP. Then the plate was washed 5 times, and the wells were incubated with 100 μL of HRP substrate for 1 h. The reaction was stopped using 100 μL of 1Μ HCl, and absorbance at 450 nm was measured using a microtiter plate reader (FLUOstar Omega, BMG LABTECH). The molar concentration of cAMP was determined from standard curves generated using standard solutions, normalized to the corresponding protein concentrations and measured using the Bradford Protein Assay (Thermo Scientific) and finally expressed as pmol cAMP/μg protein. Each experiment was performed in triplicate, and the results are expressed as mean ± S.D.
2.6.3. Preservation of Bioactivity of Liposomal and Free RLX2 Peptides
For these experiments, RLX2 peptides (400 nM) in the form of solutions (free) or liposomes (with lipid membrane composition: PC/PG/Chol/PEG, 8.0:2.0:5.0:1.3 mol/mol) were incubated in FBS (80% v/v) at 37 °C, for 30 d. At specific time points (0, 1, 3, 4, 10, and 30 d), 250 μL of each sample was added to 750 μL of culture medium (4× dilution), in order to realize a peptide concentration of 100 nM for all tested samples. HEK-293 cells were transfected and stimulated with each compound, and the cAMP concentration was measured, following the procedures described above. Experiments were carried out in triplicate, and the results are expressed as mean value ± standard deviation.
2.7. Statistical Methods
The IBM SPSS statistics pack was used for the statistical analysis of the results. All experiments were performed in triplicate. All data are presented as the mean ± standard deviation of the mean of independent experiments. Statistical significance was evaluated using one-way ANOVA or two-way ANOVA and LSD’s post hoc test with a significance level of p < 0.05.
4. Discussion
Liposome encapsulation and fatty acid conjugation were investigated as methods to retain the stability and prolong the biological activity of RLX2 peptides. To the best of our knowledge this is the first report that has compared the effect of chemical modification and liposome encapsulation of any peptide on the peptide activity. A palmitoyl conjugate of RLX2 peptide (P-RLX2) was synthesized, using
N-Palmitoyl-
l-glutamic acid α-tert-butyl ester γ-succinimidyl ester (Palmitoyl-
l-Glu(OSu)-OtBu) as the fatty acid modifier, after identifying an efficient synthetic pathway and the appropriate conditions that conferred a high production yield of the monopalmitate RLX2 peptide conjugate, in the final stage of its synthesis. This peptide modifier is known to delay peptide absorption and extend the half-life of Liraglutide, while renal clearance is also reduced, due to the shielding effect of the fatty acid moiety [
21].
RLX2 peptide encapsulating nanosized liposomes were additionally formulated (with mean diameters between 114 nm–160 nm) for all the peptide types, by appropriate methods that confer a high encapsulation. In more detail, the DRV method [
22,
23], a mild method, which protects the integrity of peptide/protein drugs and confers high encapsulation for aqueous soluble materials, was used for encapsulation of RLX2 and O-RLX2 peptides, and it was found that inclusion of a negatively charged lipid (PG) conferred the maximum encapsulation of these peptides (EE up to 22%), due to the fact that they were oppositely charged at pH 7.40. Protein encapsulation in liposomes has been reported to be dependent on the extent of interactions that occur between the encapsulated proteins and the phospholipid bilayer [
34]. For the P-RLX2 lipid conjugate of RLX2 incorporation in the liposome membrane, the thin film method was applied, and it was fully incorporated in the liposomes.
For the reduction of the size of both types of peptide-loaded liposomes, DRVs and MLVs, extrusion was applied, since it was previously reported that extrusion resulted in a much higher retention of heparin in nanosized DRV liposomes compared to sonication [
35]. We anticipate that extrusion may additionally be less harmful to such bioactive peptides than sonication; however, this remains to be proven.
It is well known that PEGylated liposomes have a higher integrity compared to non-PEGylated ones (in presence of serum proteins, FBS) [
24,
30,
31], and thereby it was essential to study PEGylated liposomes. In order to coat the RLX2-peptide entrapping liposomes with PEG, a post-PEGylation method was required, since when PEG was added initially (during lipid film formation (conventional method)), a very low encapsulation was realized (2.85%). In good agreement with previous findings, when the stability of RLX2 peptides was evaluated in buffer, the entrapment of the peptide in PEGylated liposomes was demonstrated to confer maximum peptide integrity. However, the peptide integrity in the presence of serum was not affected by liposome PEGylation, at least under the conditions applied herein. In fact the integrity of RLX2 peptide was significantly higher in serum, compared to that measured in buffer (without proteins), probably due to the increased molecular shielding of the peptides by the proteins.
The results of the peptide stability studies indicated that both peptide types, RLX2 (
Figure 3) and O-RLX2 (
Figure 4) were highly protected against degradation when they were entrapped in liposomes.
The current results about the liposomal forms of RLX2 peptide types cannot be compared with others, since the liposomal encapsulation of RLX2 had not been previously studied as a method to prolong this peptide’s activity; the beneficial effect of a special type of targeted RLX2 liposomes has been reported [
4]; however, the properties and/or stability of RLX2 in those formulations have not been reported.
On the other hand, several chemical modifications were previously reported for RLX2. Dicarba RLX2 analogues were found to retain their activity but lose their stability in serum [
13], while other synthetic covalently linked dimeric forms of human RLX2 were seen to retain their (receptor binding) activity and also demonstrated improved in vitro serum stability [
15]. Nevertheless, a direct comparison of the reported results of RLX2 stability in serum with the current results could not be made, since much lower stabilities in serum were reported by others for the monomeric amidated form of H2 relaxin, and also for native H2 relaxin [
13,
36], probably due to the fact that different peptide concentrations and a different type of serum (human serum) were used in those studies. In any case, it should be pointed out that, according to previous studies, the in vivo half-life of peptides is always much lower compared to in vitro results, since renal clearance is a major factor that determines the in vivo half-life of peptides [
15].
Comparing the retention of the bioactivity of the three peptide types studied herein, from the dose response bioactivity experiments carried out in the absence of proteins, it was indicated that peptide oxidation on Met25 has an activity reducing effect, and palmitoylation of RLX2 with a Palmitoyl-
l-Glu-OtBu modifier at the a-amino group of the
N-terminal amino acid of the Β-chain, had a similar effect on the bioactivity of the peptide as that caused by oxidation (
Figure 5). However, when the RLX2 bioactivity was measured in presence of proteins (
Figure 6), the results suggested that interactions of the various peptide types with serum proteins affected their bioactivity in different ways, since P-RLX2 had a significantly lower bioactivity compared to O-RLX2, and both latter peptide types had a lower bioactivity compared to RLX2.
Interestingly, although the bioactivity measured for P-RLX2 conjugate was significantly lower than that of RLX2 (and O-RLX2), it was highly preserved. Indeed, after 30 d incubation in FBS at 37 °C, the bioactivity of the conjugate was much higher (approx. 30% of its initial value) compared to the preservation of the bioactivity of the liposomal form of RLX2 peptide (the only other peptide type that retained a significant bioactivity value after 30 d, which was approx. 13% of its initial value). However, due to the higher initial bioactivity of liposomal RLX2, its bioactivity was practically the same as that of P-RLX2 at the 30 d time point. Nevertheless, liposomal RLX2 demonstrated a much higher bioactivity at all other time points evaluated; therefore, according to the current results, entrapment of RLX2 peptide in PEGylated liposomes was the most successful method, among those evaluated herein, for attaining a high and prolonged biological activity of RLX2. Another very interesting finding from the current study is that the stability study resulted for for the free and liposomal forms of RLX2 and O-RLX2 peptides are in good agreement with the results of the bioactivity preservation studies, indicating that such studies may provide good predictions for the preservation of their biological activity.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}