
Functional Gait Assessment Using Manual, Semi-Automated and Deep Learning Approaches Following Standardized Models of Peripheral Nerve Injury in Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
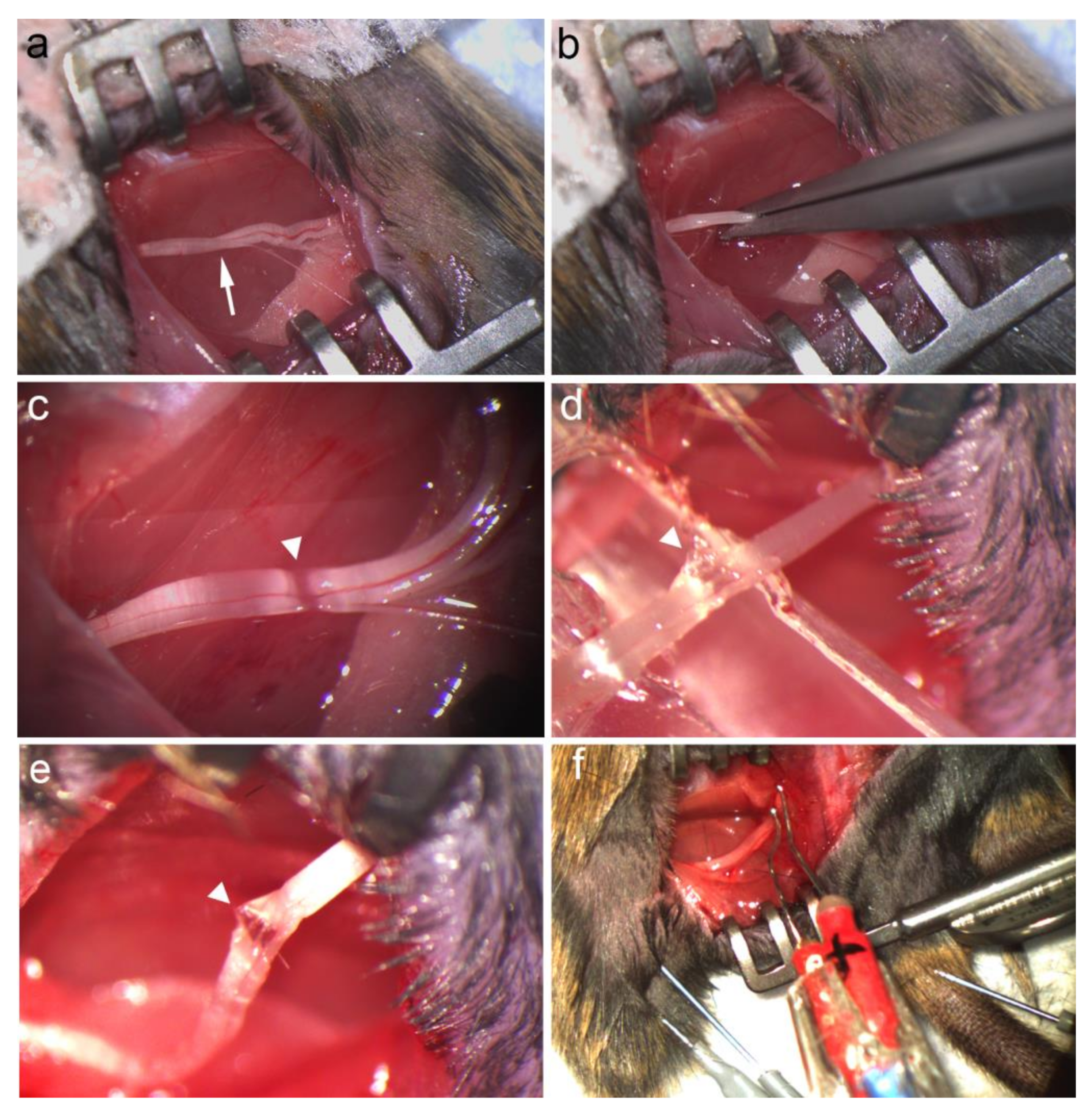
2.2. Nerve Injury Model
2.2.1. Crush Injury
2.2.2. Stretch–Crush Injury
2.3. Behavioral Assessments: Von Frey Monofilament Test, Gait Analysis, and SFI
2.3.1. Von Frey Monofilament Testing
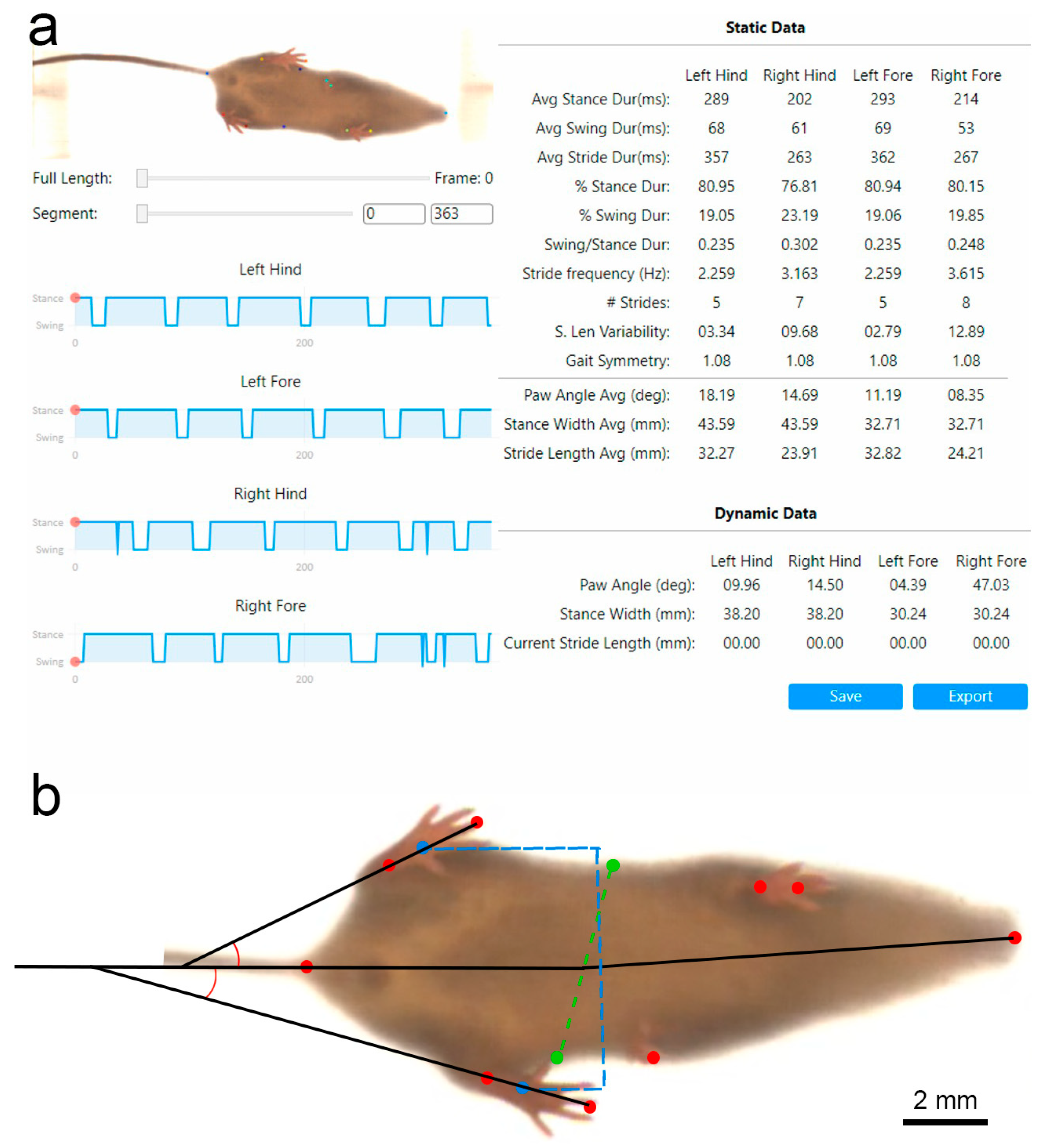
2.3.2. Gait Analysis
2.3.3. Sciatic Functional Index (SFI)
2.4. Electrophysiological Assessment
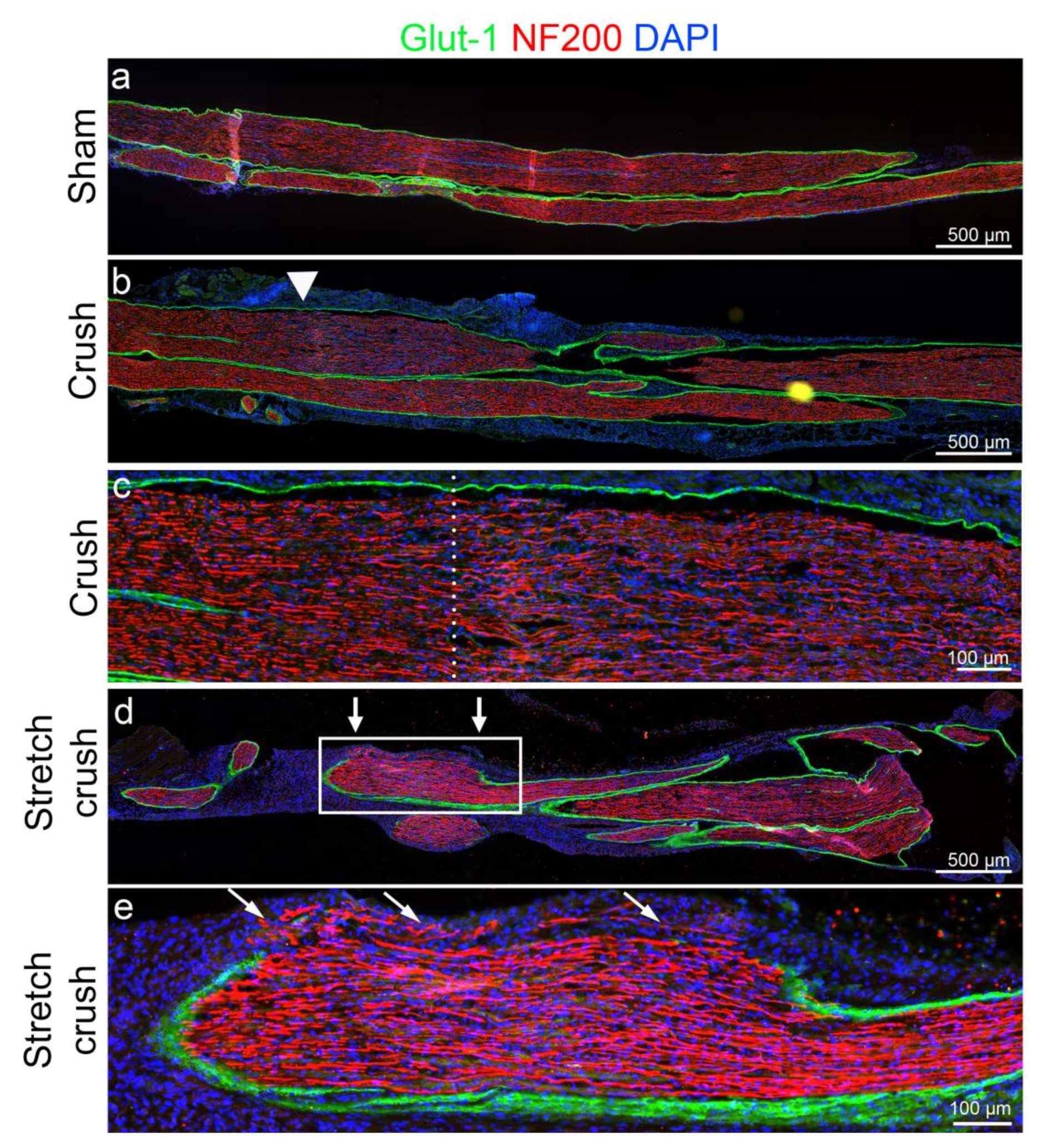
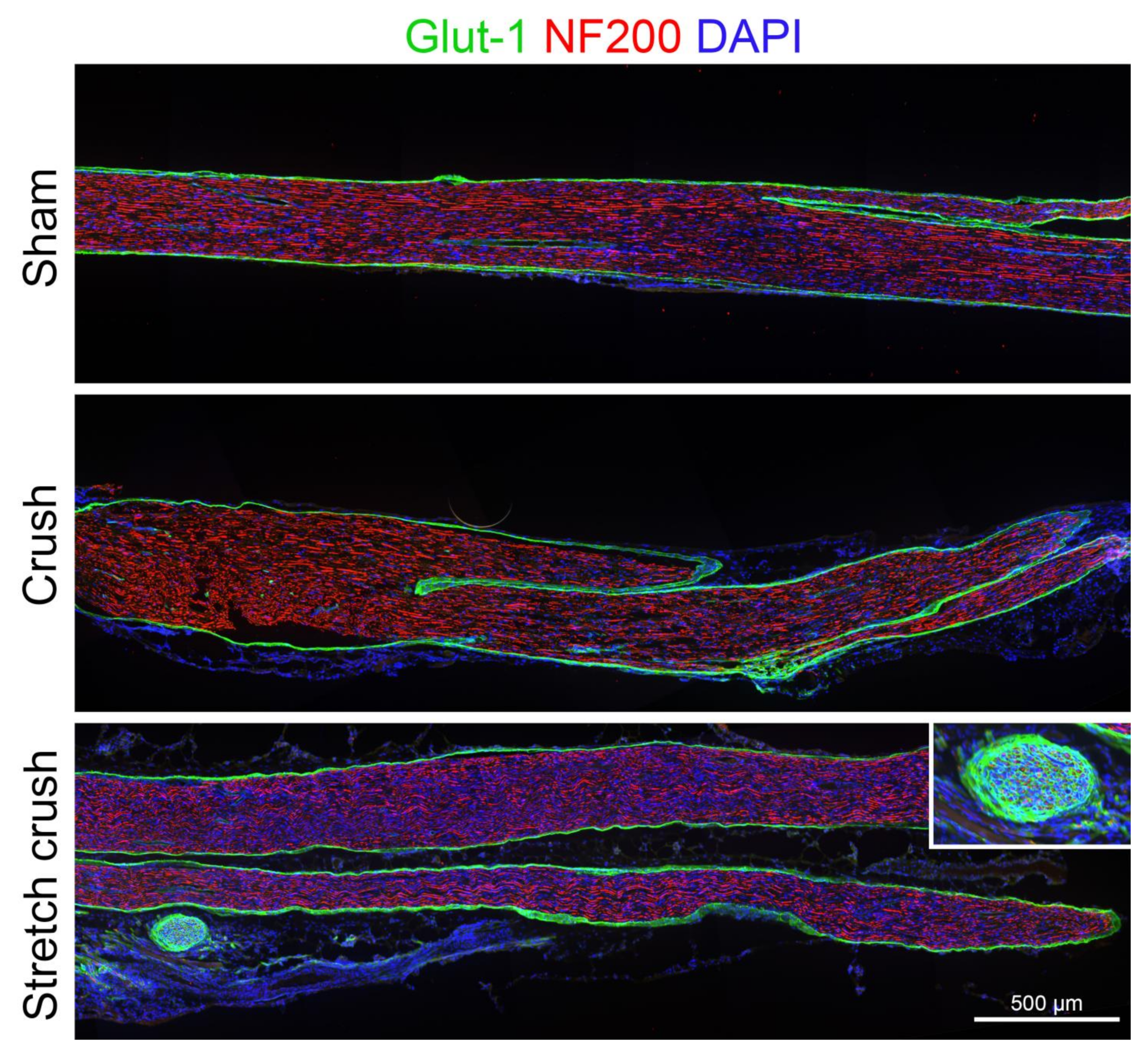
2.5. Histological Assessment
2.6. Muscle Weight Assessment
2.7. Statistics
3. Results
3.1. Histology
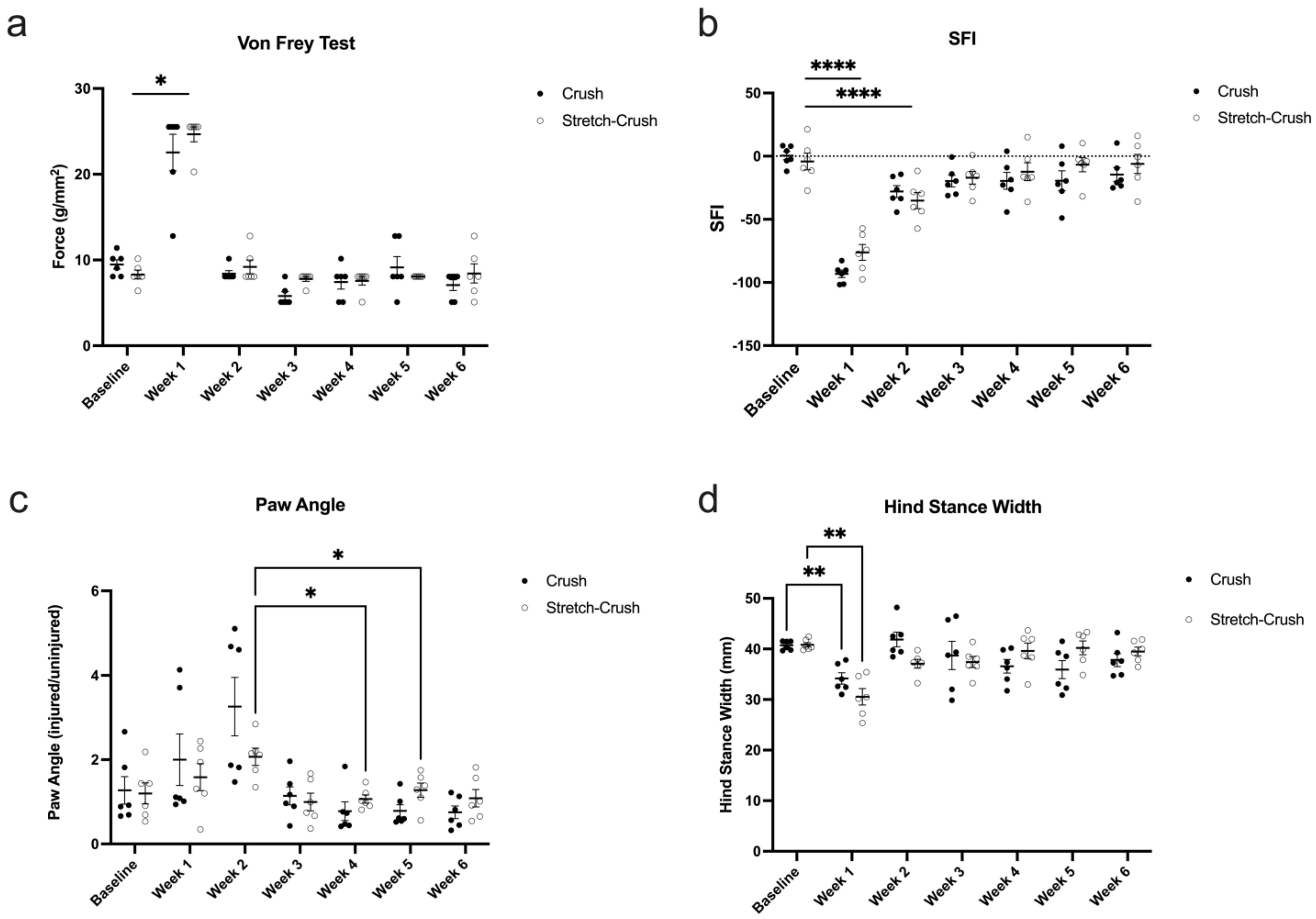
3.2. Behavioral Assessments: Von Frey Monofilament Test, Gait Analysis and SFI
3.2.1. Von Frey Monofilament Testing
3.2.2. SFI
3.2.3. Visual Gait Lab Analysis
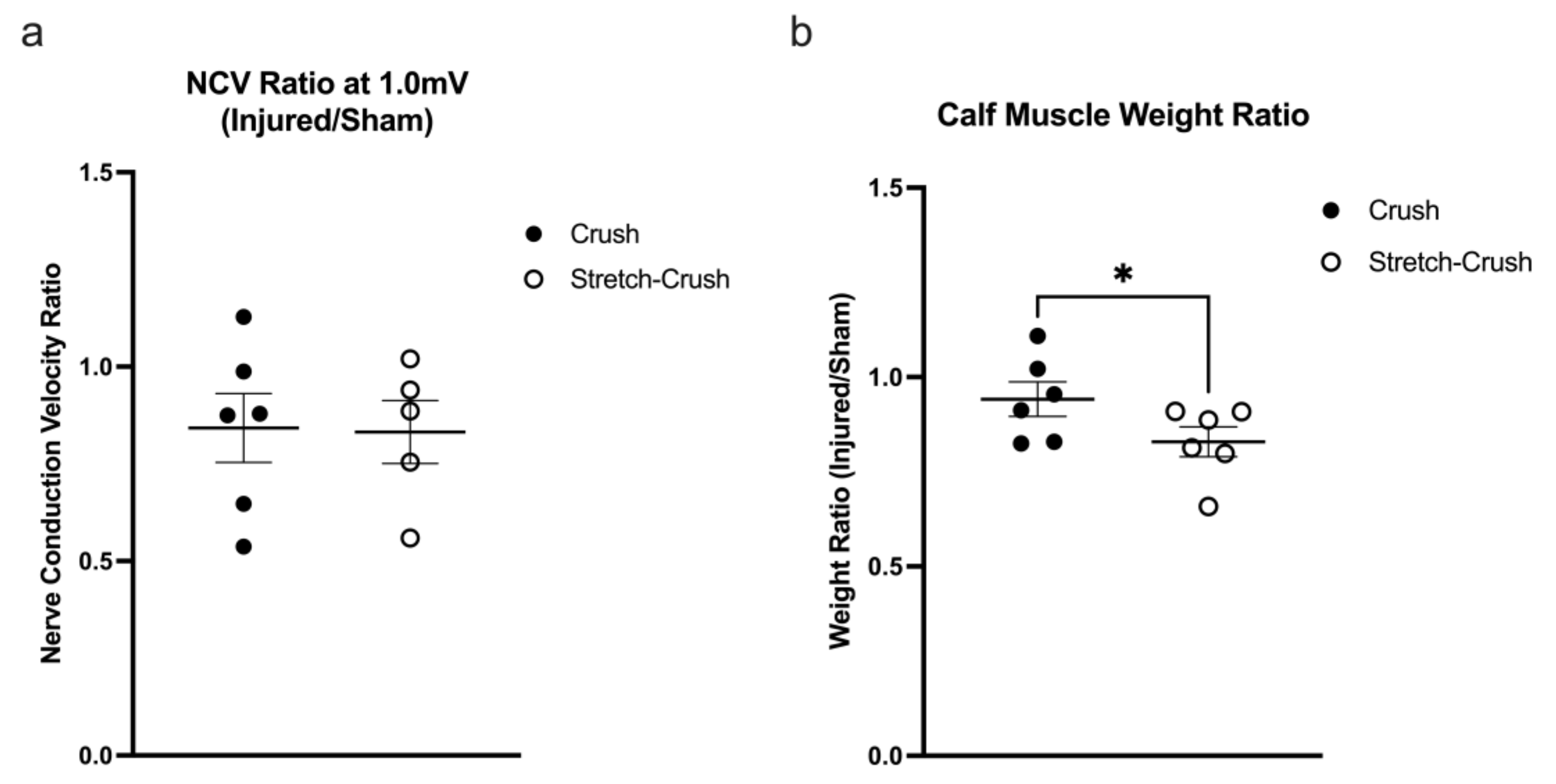
3.3. Electrophysiological and Muscle Weight Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noble, J.; Munro, C.A.; Prasad, V.S.; Midha, R. Analysis of Upper and Lower Extremity Peripheral Nerve Injuries in a Population of Patients with Multiple Injuries. J. Trauma 1998, 45, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, C.A.; Kung, T.A.; Brown, D.L.; Cederna, P.S.; Kemp, S.W.P. State-of-The-Art Techniques in Treating Peripheral Nerve Injury. Plast. Reconstr. Surg. 2018, 141, 702–710. [Google Scholar] [CrossRef]
- Simon, N.G.; Spinner, R.J.; Kline, D.G.; Kliot, M. Advances in the Neurological and Neurosurgical Management of Peripheral Nerve Trauma. J. Neurol. Neurosurg. Psychiatry 2016, 87, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Gonciarz, M.D.; Sundquist, W.I.; Hill, C.P.; Jensen, J. Cryo-EM Structure of Dodecameric Vps4p and Its 2:1 Complex with Vta1p. NIH Public Access. 2009, 377, 364–377. [Google Scholar] [CrossRef]
- Walsh, S.; Midha, R. Use of Stem Cells To Augment Nerve Injury Repair. Neurosurgery 2009, 65, A80–A86. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Begum, R.; Barber, T.; Ibba, V.; Tee, N.C.H.; Hussain, M.; Arastoo, M.; Yang, Q.; Robson, L.G.; Lesage, S.; et al. Regenerative Potential of Silk Conduits in Repair of Peripheral Nerve Injury in Adult Rats. Biomaterials 2012, 33, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Navarro, X.; Vivó, M.; Valero-Cabré, A. Neural Plasticity after Peripheral Nerve Injury and Regeneration. Prog. Neurobiol. 2007, 82, 163–201. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, S.; Warner, W.S.; Eli, I.; Mahan, M.A. Rapid-Stretch Injury to Peripheral Nerves: Comparison of Injury Models. J. Neurosurg. 2020, 135, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Alant, J.D.D.V.; Kemp, S.W.P.; Khu, K.J.O.L.; Kumar, R.; Webb, A.A.; Midha, R. Traumatic Neuroma in Continuity Injury Model in Rodents. J. Neurotrauma 2012, 29, 1691–1703. [Google Scholar] [CrossRef]
- Umansky, D.; Midha, R. Commentary: Rapid-Stretch Injury to Peripheral Nerves: Histologic Results. Neurosurgery 2019, 86, E333–E334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahan, M.A.; Warner, W.S.; Yeoh, S.; Light, A. Rapid-Stretch Injury to Peripheral Nerves: Implications from an Animal Model. J. Neurosurg. 2019, 133, 1–11. [Google Scholar] [CrossRef]
- Varejäo, A.S.P.; Meek, M.F.; Ferreira, A.J.A.; Patrício, J.A.B.; Cabritae, A.M.S. Functional Evaluation of Peripheral Nerve Regeneration in the Rat: Walking Track Analysis. J. Neurosci. Methods 2001, 108, 1–9. [Google Scholar] [CrossRef]
- Varejão, A.S.P.; Cabrita, A.M.; Meek, M.F.; Bulas-Cruz, J.; Melo-Pinto, P.; Raimondo, S.; Geuna, S.; Giacobini-Robecchi, M.G. Functional and Morphological Assessment of a Standardized Rat Sciatic Nerve Crush Injury with a Non-Serrated Clamp. J. Neurotrauma 2004, 21, 1652–1670. [Google Scholar] [CrossRef]
- De Medinaceli, L.; Freed, W.J.; Wyatt, R.J. An Index of the Functional Condition of Rat Sciatic Nerve Based on Measurements Made from Walking Tracks. Exp. Neurol. 1982, 77, 634–643. [Google Scholar] [CrossRef]
- Mahan, M.A.; Yeoh, S.; Monson, K.; Light, A. Rapid Stretch Injury to Peripheral Nerves: Biomechanical Results. Neurosurgery 2018, 85, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Umansky, D.; Midha, R. Editorial. The Quintessential Nerve Injury Model. J. Neurosurg. 2021, 135, 890–892. [Google Scholar] [CrossRef]
- Bain, J.R.; Mackinnon, S.E.; Hunter, D.A. Functional Evaluation of Complete Sciatic, Peroneal, and Posterior Tibial Nerve Lesions in the Rat. Plast. Reconstr. Surg. 1989, 83, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A.; Szalai, J.P.; Mackinnon, S.E.; Midha, R. Lack of Association between Outcome Measures of Nerve Regeneration. Muscle Nerve 1998, 21, 1095–1097. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Nath, T.; Mathis, A.; Chen, A.C.; Patel, A.; Bethge, M.; Mathis, M.W. Using DeepLabCut for 3D Markerless Pose Estimation across Species and Behaviors. Nat. Protoc. 2019, 14, 2152–2176. [Google Scholar] [CrossRef] [PubMed]
- Fiker, R.; Kim, L.H.; Molina, L.A.; Chomiak, T.; Whelan, P.J. Visual Gait Lab: A User-Friendly Approach to Gait Analysis. J. Neurosci. Methods 2020, 341, 108775. [Google Scholar] [CrossRef] [PubMed]
- Mathis, A.; Mamidanna, P.; Cury, K.M.; Abe, T.; Murthy, V.N.; Mathis, M.W.; Bethge, M. DeepLabCut: Markerless Pose Estimation of User-Defined Body Parts with Deep Learning. Nat. Neurosci. 2018, 21, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Heinzel, J.; Längle, G.; Oberhauser, V.; Hausner, T.; Kolbenschlag, J.; Prahm, C.; Grillari, J.; Hercher, D. Use of the CatWalk Gait Analysis System to Assess Functional Recovery in Rodent Models of Peripheral Nerve Injury – a Systematic Review. J. Neurosci. Methods 2020, 345, 108889. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative Assessment of Tactile Allodynia in the Rat Paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods Used to Evaluate Pain Behaviors in Rodents. Front. Mol. Neurosci. 2017, 10, 1–17. [Google Scholar] [CrossRef]
- Bonin, R.P.; Bories, C.; De Koninck, Y. A Simplified Up-down Method (SUDO) for Measuring Mechanical Nociception in Rodents Using von Frey Filaments. Mol. Pain 2014, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Bradman, M.J.G.; Ferrini, F.; Salio, C.; Merighi, A. Practical Mechanical Threshold Estimation in Rodents Using von Frey Hairs/Semmes–Weinstein Monofilaments: Towards a Rational Method. J. Neurosci. Methods 2015, 255, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.W.; Krug, H.E.; Frizelle, S.P.; Funkenbusch, S.; Mahowald, M.L. A Comparison of DigiGaitTM and TreadScanTM Imaging Systems: Assessment of Pain Using Gait Analysis in Murine Monoarthritis. J. Pain Res. 2014, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Guillot, T.S.; Asress, S.A.; Richardson, J.R.; Glass, J.D.; Miller, G.W. Treadmill Gait Analysis Does Not Detect Motor Deficits in Animal Models of Parkinson’s Disease or Amyotrophic Lateral Sclerosis. J. Mot. Behav. 2008, 40, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Inserra, M.M.; Bloch, D.A.; Terris, D.J. Functional Indices for Sciatic, Peroneal, and Posterior Tibial Nerve Lesions in the Mouse. Microsurgery 1998, 18, 119–124. [Google Scholar] [CrossRef]
- Stratton, J.A.; Holmes, A.; Rosin, N.L.; Sinha, S.; Vohra, M.; Burma, N.E.; Trang, T.; Midha, R.; Biernaskie, J. Macrophages Regulate Schwann Cell Maturation after Nerve Injury. Cell Rep. 2018, 24, 2561–2572.e6. [Google Scholar] [CrossRef] [PubMed]
- Lele, P.P. Effects of Focused Ultrasonic Radiation on Peripheral Nerve, with Observations on Local Heating. Exp. Neurol. 1963, 8, 47–83. [Google Scholar] [CrossRef]
- Takata, K.; Hirano, H.; Kasahara, M. Transport of Glucose across the Blood-Tissue Barriers. Int. Rev. Cytol. 1997, 172, 1–53. [Google Scholar] [CrossRef]
- Tserentsoodol, N.; Shin, B.C.; Suzuki, T.; Takata, K. Colocalization of Tight Junction Proteins, Occludin and ZO-1, and Glucose Transporter GLUT1 in Cells of the Blood-Ocular Barrier in the Mouse Eye. Histochem. Cell Biol. 1998, 110, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Shakhbazau, A.; Kawasoe, J.; Hoyng, S.A.; Kumar, R.; van Minnen, J.; Verhaagen, J.; Midha, R. Early Regenerative Effects of NGF-Transduced Schwann Cells in Peripheral Nerve Repair. Mol. Cell. Neurosci. 2012, 50, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Martini, R.; Fischer, S.; Lopez-Vales, R.; David, S. Interactions between Schwann Cells and Macrophages in Injury and Inherited Demyelinating Disease. Glia 2008, 56, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Midha, R. Mechanisms and Pathology of Injury. In Kline and Hudson’s Nerve Injuries: Operative Results for Major Nerve Injuries, Entrapments and Tumors; Kim, D.H., Kline, D.G., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2008; pp. 23–42. [Google Scholar]
- Lundborg, G. Nerve Injury and Repair—A Challenge to the Plastic Brain. J. Peripher. Nerv. Syst. 2003, 8, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.Y.; Gordon, T. The Cellular and Molecular Basis of Peripheral Nerve Regeneration. Mol. Neurobiol. 1997, 14, 67–116. [Google Scholar] [CrossRef]
- Kaplan, H.M.; Mishra, P.; Kohn, J. The Overwhelming Use of Rat Models in Nerve Regeneration Research May Compromise Designs of Nerve Guidance Conduits for Humans. J. Mater. Sci. Mater. Med. 2015, 26, 1–5. [Google Scholar] [CrossRef]
- Fadia, N.B.; Bliley, J.M.; Di Bernardo, G.A.; Crammond, D.J.; Schilling, B.K.; Sivak, W.N.; Spiess, A.M.; Washington, K.M.; Waldner, M.; Liao, H.-T.; et al. Long-Gap Peripheral Nerve Repair through Sustained Release of a Neurotrophic Factor in Nonhuman Primates. Sci. Transl. Med. 2020, 12, eaav7753. [Google Scholar] [CrossRef] [PubMed]
- Grochmal, J.; Teo, W.; Gambhir, H.; Kumar, R.; Stratton, J.A.; Dhaliwal, R.; Brideau, C.; Biernaskie, J.; Stys, P.K.; Midha, R. A Novel Approach to 32-Channel Peripheral Nervous System Myelin Imaging in vivo, with Single Axon Resolution. J. Neurosurg. 2018, 130, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McLean, N.A.; Verge, V.M.K. Dynamic Impact of Brief Electrical Nerve Stimulation on the Neural Immune Axis—Polarization of Macrophages toward a pro-Repair Phenotype in Demyelinated Peripheral Nerve. Glia 2016, 64, 1546–1561. [Google Scholar] [CrossRef]
- Warner, W.S.; Yeoh, S.; Light, A.; Zhang, J.; Mahan, M.A. Rapid-Stretch Injury to Peripheral Nerves: Histologic Results. Neurosurgery 2019, 86, 437–445. [Google Scholar] [CrossRef]
- De Alant, J.D.V.; Senjaya, F.; Ivanovic, A.; Forden, J.; Shakhbazau, A.; Midha, R. The Impact of Motor Axon Misdirection and Attrition on Behavioral Deficit Following Experimental Nerve Injuries. PLoS ONE 2013, 8, e82546. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Inserra, M.M.; Duh, M.J.; Terris, D.J. A Longitudinal, Functional Study of Peripheral Nerve Recovery in the Mouse. Laryngoscope 1998, 108, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Inserra, M.M.; Yao, M.; Murray, R.; Terris, D.J. Peripheral Nerve Regeneration in Interleukin 6-Deficient Mice. Arch. Otolaryngol. Head Neck Surg. 2000, 126, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Siqueira Mietto, B.; Kroner, A.; Girolami, E.I.; Santos-Nogueira, E.; Zhang, J.; David, S. Role of IL-10 in Resolution of Inflammation and Functional Recovery after Peripheral Nerve Injury. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 16431–16442. [Google Scholar] [CrossRef]
- Mietto, B.S.; Jurgensen, S.; Alves, L.; Pecli, C.; Narciso, M.S.; Assunção-Miranda, I.; Villa-Verde, D.M.S.; de Souza Lima, F.R.; de Menezes, J.R.L.; Benjamim, C.F.; et al. Lack of Galectin-3 Speeds Wallerian Degeneration by Altering TLR and pro-Inflammatory Cytokine Expressions in Injured Sciatic Nerve. Eur. J. Neurosci. 2013, 37, 1682–1690. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.; Sanes, J.R.; Lichtman, J.W. Pre-Existing Pathways Promote Precise Projection Patterns. Nat. Neurosci. 2002, 5, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, K.R.; Fricke, S.T.; Faden, A.I. Neuropathological Differences between Rats and Mice after Spinal Cord Injury. J. Magn. Reson. Imaging 2010, 32, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Sroga, J.M.; Jones, T.B.; Kigerl, K.A.; McGaughy, V.M.; Popovich, P.G. Rats and Mice Exhibit Distinct Inflammatory Reactions after Spinal Cord Injury. J. Comp. Neurol. 2003, 462, 223–240. [Google Scholar] [CrossRef]
- Irintchev, A. Potentials and Limitations of Peripheral Nerve Injury Models in Rodents with Particular Reference to the Femoral Nerve. Ann. Anat. Anat. Anz. 2011, 193, 276–285. [Google Scholar] [CrossRef]
- Pavić, R.; Pavić, M.L.; Tot, O.K.; Bensić, M.; Heffer-Lauc, M. Side Distinct Sciatic Nerve Recovery Differences between Rats and Mice. Somatosens. Mot. Res. 2008, 25, 163–170. [Google Scholar] [CrossRef]
- Lakes, E.H.; Allen, K.D. Gait Analysis Methods for Rodent Models of Arthritic Disorders: Reviews and Recommendations. Osteoarthr. Cartil. 2016, 24, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.Y.; Kloefkorn, H.E.; Allen, K.D. Gait Analysis Methods for Rodent Models of Osteoarthritis. Curr. Pain Headache Rep. 2014, 18, 456. [Google Scholar] [CrossRef] [PubMed]
- Hamers, F.P.; Lankhorst, A.J.; van Laar, T.J.; Veldhuis, W.B.; Gispen, W.H. Automated Quantitative Gait Analysis during Overground Locomotion in the Rat: Its Application to Spinal Cord Contusion and Transection Injuries. J. Neurotrauma 2001, 18, 187–201. [Google Scholar] [CrossRef]
- Beare, J.E.; Morehouse, J.R.; DeVries, W.H.; Enzmann, G.U.; Burke, D.A.; Magnuson, D.S.K.; Whittemore, S.R. Gait Analysis in Normal and Spinal Contused Mice Using the TreadScan System. J. Neurotrauma 2009, 26, 2045–2056. [Google Scholar] [CrossRef]
- Akula, S.K.; McCullough, K.B.; Weichselbaum, C.; Dougherty, J.D.; Maloney, S.E. The Trajectory of Gait Development in Mice. Brain Behav. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Vinsant, S.; Mansfield, C.; Jimenez-Moreno, R.; Del Gaizo Moore, V.; Yoshikawa, M.; Hampton, T.G.; Prevette, D.; Caress, J.; Oppenheim, R.W.; Milligan, C. Characterization of Early Pathogenesis in the SOD1(G93A) Mouse Model of ALS: Part I, Background and Methods. Brain Behav. 2013, 3, 335–350. [Google Scholar] [CrossRef]
- Hampton, T.G.; Amende, I. Treadmill Gait Analysis Characterizes Gait Alterations in Parkinson’s Disease and Amyotrophic Lateral Sclerosis Mouse Models. J. Mot. Behav. 2009, 42, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Mathis, M.W.; Mathis, A. Deep Learning Tools for the Measurement of Animal Behavior in Neuroscience. Curr. Opin. Neurobiol. 2020, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, S. Deep Learning: The Bad, the Ugly, and the Good. VAHPERD J. 2019, 28, 8. [Google Scholar]
- Xu, Y.; Tian, N.X.; Bai, Q.Y.; Chen, Q.; Sun, X.H.; Wang, Y. Gait Assessment of Pain and Analgesics: Comparison of the DigiGaitTM and CatWalkTM Gait Imaging Systems. Neurosci. Bull. 2019, 35, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Dellon, E.S.; Dellon, A.L. Functional Assessment of Neurologic Impairment: Track Analysis in Diabetic and Compression Neuropathies. Plast. Reconstr. Surg. 1991, 88, 686–694. [Google Scholar] [CrossRef]
- Sakuma, T.; Kamoda, H.; Miyagi, M.; Ishikawa, T.; Arai, G.; Eguchi, Y.; Suzuki, M.; Oikawa, Y.; Sakuma, Y.; Kubota, G.; et al. Comparison of Catwalk Analysis and von Frey Testing for Pain Assessment in a Rat Model of Nerve Crush plus Inflammation. Spine 2013, 38, 919–924. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Sheu, M.L.; Cheng, F.C.; Chen, C.J.; Su, H.L.; Sheehan, J.; Pan, H.C. Comprehensive Analysis of Neurobehavior Associated with Histomorphological Alterations in a Chronic Constrictive Nerve Injury Model through Use of the CatWalk XT System: Laboratory Investigation. J. Neurosurg. 2014, 120, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Lemcke, S.; Müller, S.; Möller, S.; Schillert, A.; Ziegler, A.; Cepok-Kauffeld, S.; Comabella, M.; Montalban, X.; Rülicke, T.; Nandakumar, K.S.; et al. Nerve Conduction Velocity Is Regulated by the Inositol Polyphosphate-4-Phosphatase II Gene. Am. J. Pathol. 2014, 184, 2420–2429. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Oka, Y. The Relationship between Nerve Conduction Velocity and Fiber Morphology during Peripheral Nerve Regeneration. Brain Behav. 2012, 2, 382–390. [Google Scholar] [CrossRef]
- Gordon, T. Peripheral Nerve Regeneration and Muscle Reinnervation. Int. J. Mol. Sci. 2020, 21, 8652. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Stage | Number | Strain |
|---|---|---|
| Exploratory | 19 | Varying |
| Acute | 4 | C57/Bl6 |
| Behavioral | 12 | 129S6/SvEvTac |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umansky, D.; Hagen, K.M.; Chu, T.H.; Pathiyil, R.K.; Alzahrani, S.; Ousman, S.S.; Midha, R. Functional Gait Assessment Using Manual, Semi-Automated and Deep Learning Approaches Following Standardized Models of Peripheral Nerve Injury in Mice. Biomolecules 2022, 12, 1355. https://doi.org/10.3390/biom12101355
Umansky D, Hagen KM, Chu TH, Pathiyil RK, Alzahrani S, Ousman SS, Midha R. Functional Gait Assessment Using Manual, Semi-Automated and Deep Learning Approaches Following Standardized Models of Peripheral Nerve Injury in Mice. Biomolecules. 2022; 12(10):1355. https://doi.org/10.3390/biom12101355
Chicago/Turabian StyleUmansky, Daniel, Kathleen M. Hagen, Tak Ho Chu, Rajesh K. Pathiyil, Saud Alzahrani, Shalina S. Ousman, and Rajiv Midha. 2022. "Functional Gait Assessment Using Manual, Semi-Automated and Deep Learning Approaches Following Standardized Models of Peripheral Nerve Injury in Mice" Biomolecules 12, no. 10: 1355. https://doi.org/10.3390/biom12101355