
The Activity of Chosen Antioxidant Enzymes in Ostrich Meat in Relation to the Type of Packaging and Storage Time in Refrigeration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples and Packaging
2.2. Superoxide Dismutase (SOD) Assay Procedure
2.3. Determination of Glutathione Peroxidase (GPx)
2.4. Determination of Glutathione Reductase (GR)
2.5. Potential to Scavenge the Free DPPH Radical
2.6. Statistical Analysis
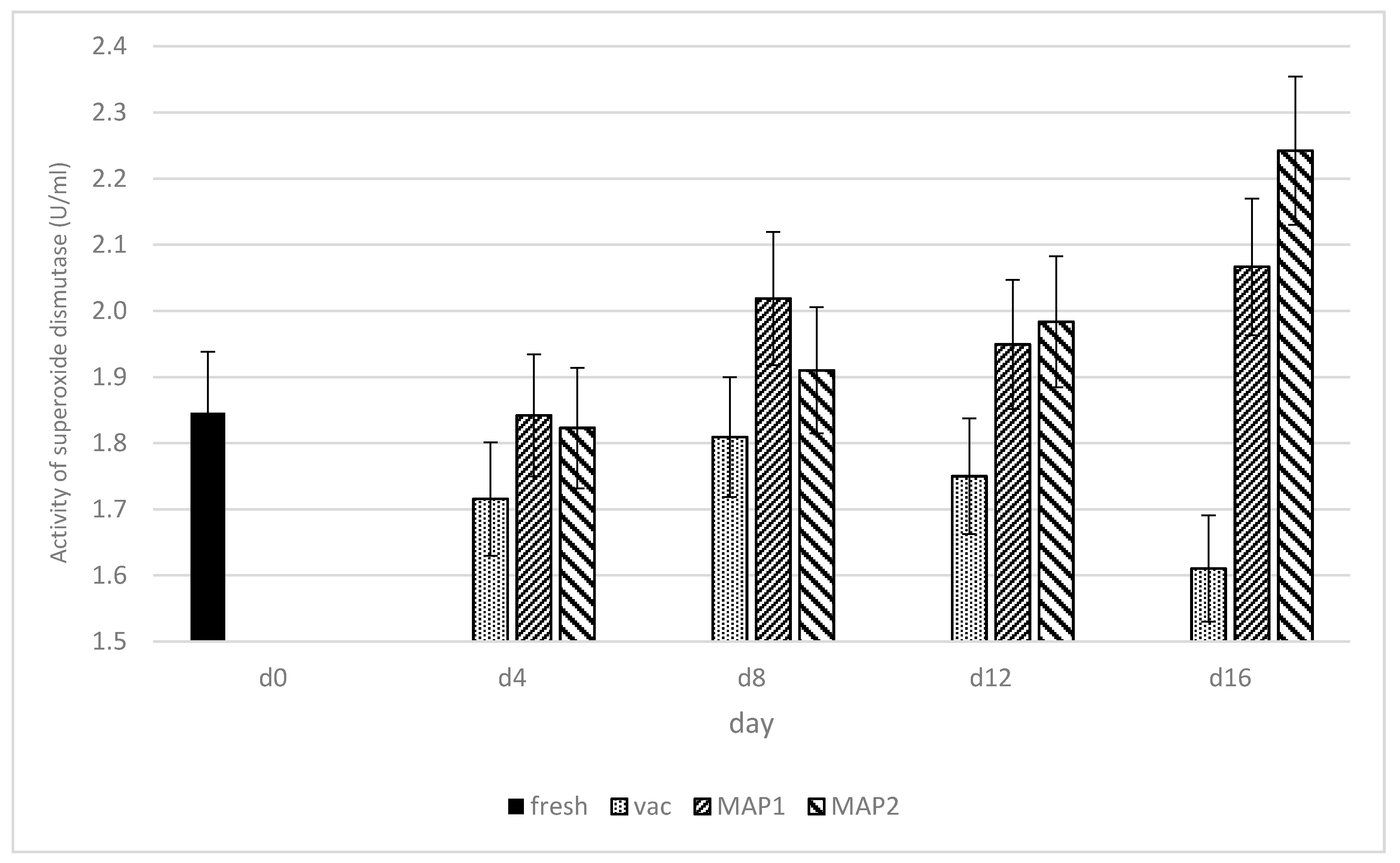
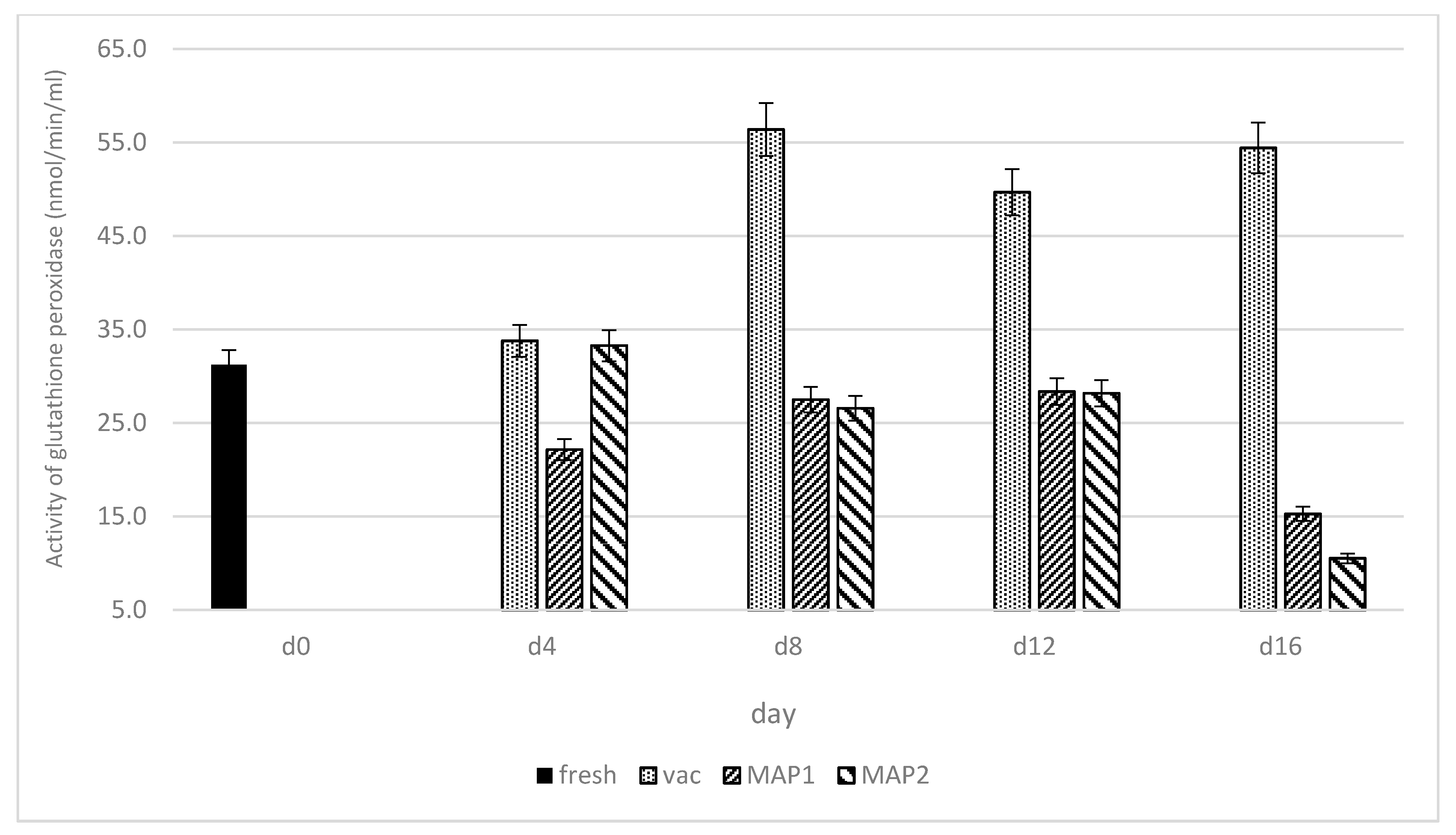
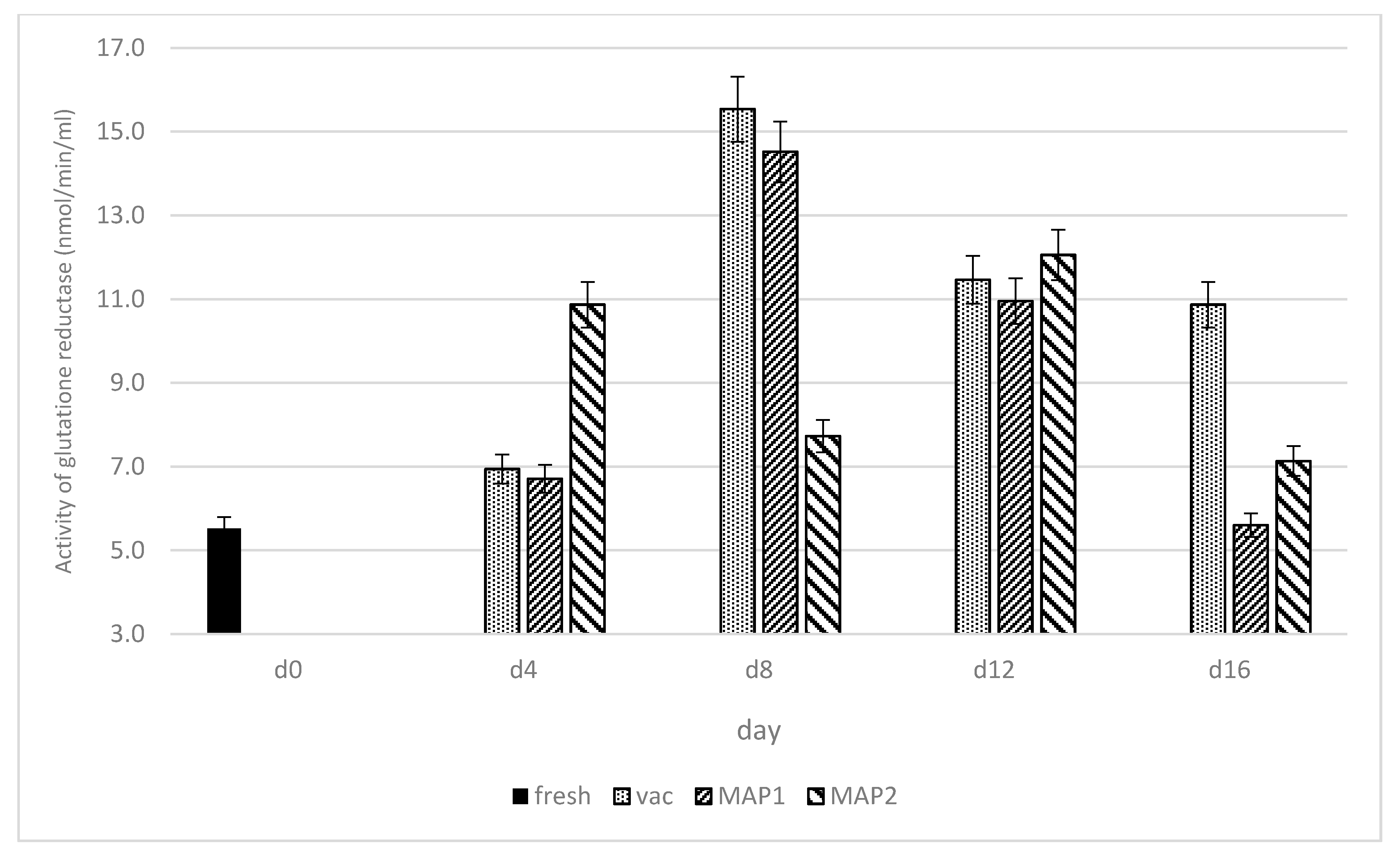
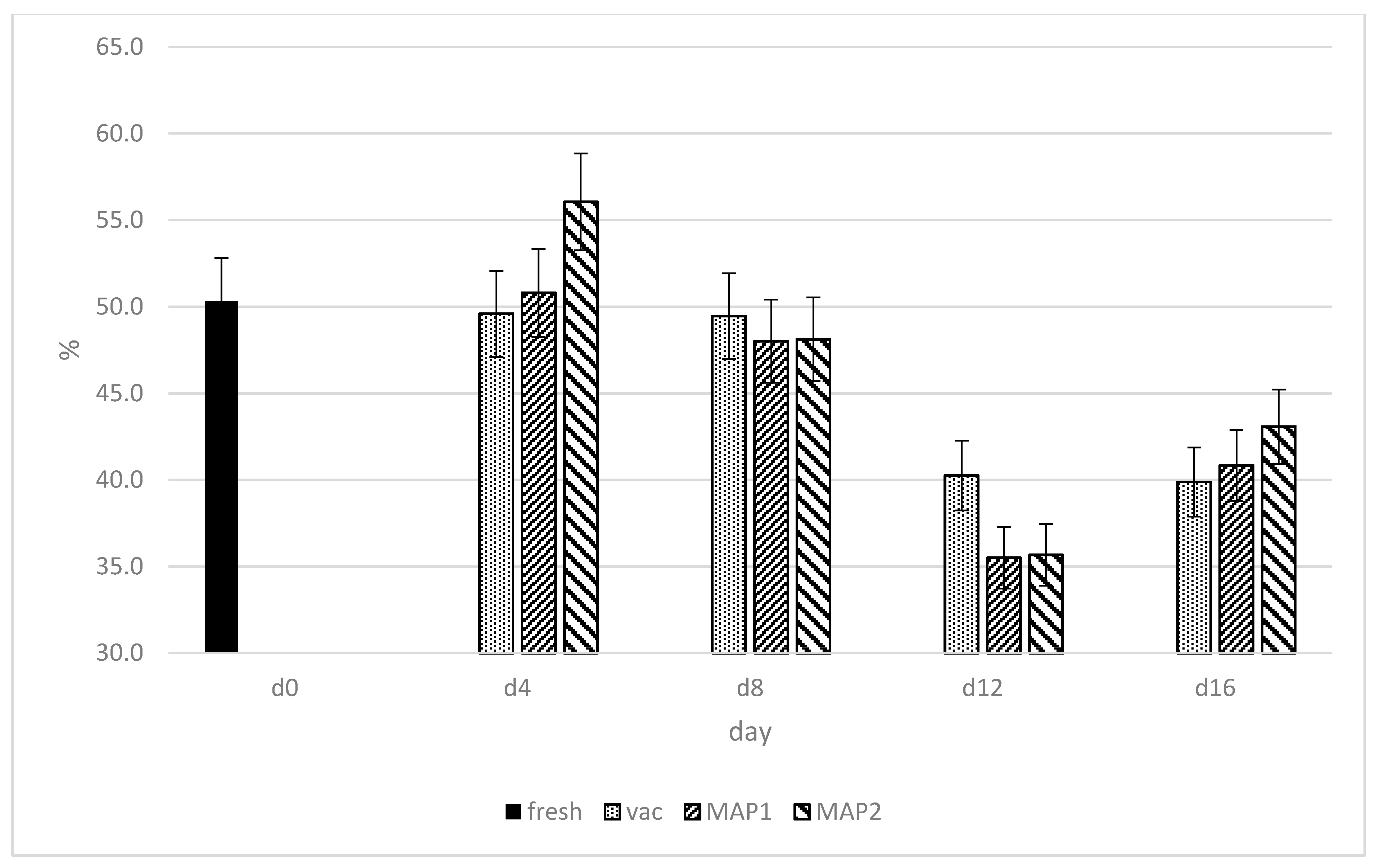
3. Results and Discussion
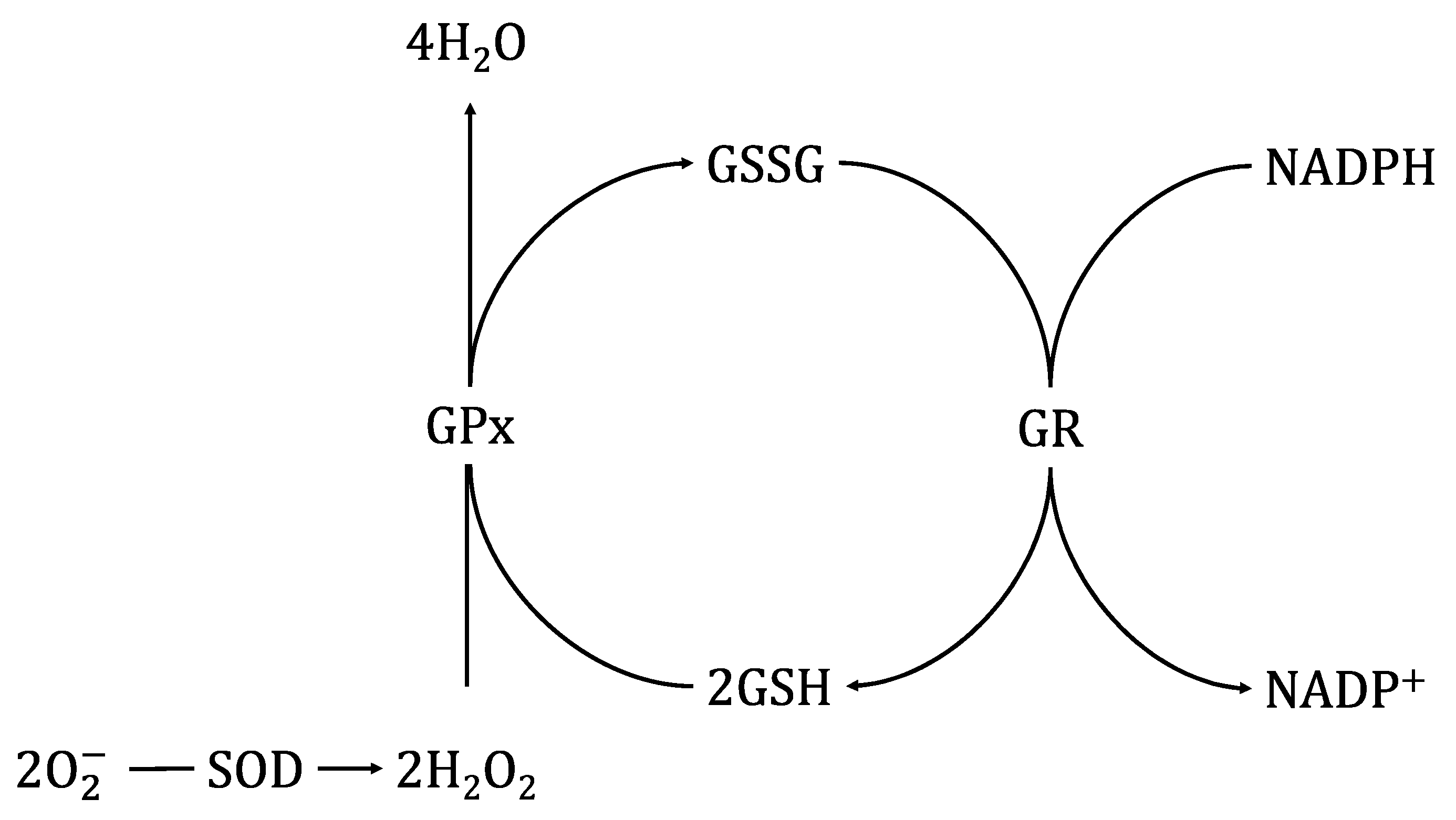
3.1. Glutathione Peroxidase (GPx) and Glutathione Reductase (GR)
3.2. DPPH
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic Metals, Ascorbate and Free Radicals: Combinations to Avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissey, P.A.; Sheehy, P.J.A.; Galvin, K.; Kerry, J.P.; Buckley, D.J. Lipid stability in meat and meat products. Meat Sci. 1998, 49, 73–86. [Google Scholar] [CrossRef]
- King, A.J.; Uijttenboogaart, T.G.; de Vries, A.W. α-Tocopherol, β-Carotene and Ascorbic Acid as Antioxidants in Stored Poultry Muscle in stored poultry muscle. J. Food Sci. 1995, 60, 1009–1012. [Google Scholar] [CrossRef]
- Serpen, A.; Gökmen, V.; Fogliano, V. Total antioxidant capacities of raw and cooked meats. Meat Sci. 2012, 90, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.G.; Horbańczuk, J.O. Ostrich nutrition: A review from a Zimbabwean perspective. Rev. Sci. Tech. 2004, 23, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Cooper, R.G.; Naranowicz, H.; Maliszewska, E.; Tennett, A.; Horbańczuk, J.O. Sex-based comparison of limb segmentation in ostriches aged 14 months with and without tibiotarsal rotation. J. S. Afr. Vet. Assoc. 2008, 79, 142–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.G.; Tomasik, C.; Horbańczuk, J.O. Avian influenza in ostriches (Struthio camelus). Avian Poult. Biol. Rev. 2007, 18, 87–92. [Google Scholar] [CrossRef]
- Horbańczuk, J.O.; Sales, J.; Celeda, T.; Konecka, A.; Zięba, G.; Kawka, P. Cholesterol content and fatty acid composition of ostrich meat as influenced by subspecies. Meat Sci. 1998, 50, 385–388. [Google Scholar] [CrossRef]
- Horbańczuk, J.O.; Kawka, M.; Sacharczuk, M.; Cooper, R.G.; Boruszewska, K.; Parada, P.; Jaszczak, K. A search for sequence similarity between chicken (Gallus domesticus) and ostrich (Struthio camelus) microsatellite markers. Anim. Sci. Pap. Rep. 2007, 25, 283–288. [Google Scholar]
- Horbańczuk, J.O.; Tomasik, C.; Cooper, R.G. Ostrich farming in Poland—Its history and current situation after accession to the European Union. Avian Poult. Biol. Rev. 2008, 1, 65–71. [Google Scholar] [CrossRef]
- Zdanowska-Sąsiadek, Ż.; Marchewka, J.; Horbańczuk, J.O.; Wierzbicka, A.; Lipińska, P.; Jóźwik, A. Nutrients composition in fit snacks made from ostrich, beef and chicken dried meat. Molecules 2018, 23, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales, J.; Horbańczuk, J.O.; Dingle, J.; Coleman, R.; Sensik, S. Carcase characteristics emus (Dromaius novaehollandiae). Brit. Poult. Sci. 1999, 40, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Sales, J.; Horbańczuk, J.O. Ratite Meat. Worlds Poult. Sci. J. 1998, 54, 59–67. [Google Scholar] [CrossRef]
- Horbańczuk, O.K.; Wierzbicka, A. Technological and nutritional properties of ostrich, emu, and rhea meat quality. J. Vet. Res. 2016, 60, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Horbańczuk, O.K.; Wierzbicka, A. Effects of packaging methods on shelf life of ratite meats. J. Vet. Res. 2017, 61, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horbańczuk, O.K.; Jóźwik, A.; Wyrwisz, J.; Marchewka, J.; Wierzbicka, A. Physical Characteristics and Microbial Quality of Ostrich Meat in Relation to the Type of Packaging and Refrigerator Storage Time. Molecules 2021, 26, 3445. [Google Scholar] [CrossRef]
- Horbańczuk, O.K.; Moczkowska, M.; Marchewka, J.; Atanasov, A.G.; Kurek, M. The Composition of Fatty Acids in Ostrich Meat Influenced by the Type of Packaging and Refrigerated Storage. Molecules 2019, 24, 4128. [Google Scholar] [CrossRef] [Green Version]
- McClements, D.J.; Decker, E.A. Lipids. In Food Chemistry, 4th ed.; Damodarin, S., Parkin, K., Fennema, O.R., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 155–216. [Google Scholar]
- Jensen, C.; Lauridsen, C.; Bertelsen, G. Dietary vitamin E: Quality and storage stability of pork and poultry. Trends Food Sci. Technol. 1998, 9, 62–72. [Google Scholar] [CrossRef]
- Terevinto, A.; Ramos, A.; Castroman, G.; Cabrera, M.C.; Saadoun, A. Oxidative status, in vitro iron-induced lipid oxidation and superoxide dismutase, catalase and glutathione peroxidase activities in rhea meat. Meat Sci. 2010, 84, 706–710. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Yue, H.; Wu, S.; Yang, H.; Wang, Z.; Qi, G. Gas stunning with CO2 affected meat color, lipid peroxidation, oxidative stress, and gene expression of mitogen-activated protein kinases, glutathione S-transferases, and Cu/Zn-superoxide dismutase in the skeletal muscles of broilers. J. Anim. Sci. Biotechnol. 2018, 9, 37. [Google Scholar] [CrossRef]
- Gatellier, P.; Mercier, Y.; Renerre, M. Effect of diet finishing mode (pasture or mixed diet) on antioxidant status of charolais bovine meat. Meat Sci. 2004, 67, 385–394. [Google Scholar] [CrossRef]
- Rennere, M.; Dumont, F.; Gatellier, P. Antioxidant enzyme activities in beef in relation to oxidation of lipid and myoglobin. Meat Sci. 1996, 43, 111–121. [Google Scholar] [CrossRef]
- Maraschiello, C.; Sárraga, C.; García Regueiro, J.A. Glutathione Peroxidase Activity, TBARS, and α-Tocopherol in Meat from Chickens Fed Different Diets. J. Agric. Food Chem. 1999, 47, 867–872. [Google Scholar] [CrossRef]
- Petron, M.J.; Raes, K.; Claeys, E.; Lourenco, M.; Fremaut, D.; De Smet, S. Effect of grazing pastures of different botanical composition on antioxidant enzyme activities and oxidative stability of lamb meat. Meat Sci. 2007, 75, 737–745. [Google Scholar] [CrossRef]
- Pradhan, A.A.; Rhee, K.S.; Hernandez, P. Stability of catalase and its potential role in lipid oxidation in meat. Meat Sci. 2000, 54, 385–390. [Google Scholar] [CrossRef]
- Pambuka, S.E.; Adebiyi, A.P.; Muramoto, K.; Naude, R.J. Purification and partial characterisation of a matrix metalloproteinase from ostrich skeletal muscle, and its activity during meat maturation. Meat Sci. 2007, 76, 481–488. [Google Scholar] [CrossRef]
- Hernández, P.; Park, D.K.; Rhee, K.S. Chloride salt type/ionic strength, muscle site and refrigeration effects on antioxidant enzymes and lipid oxidation in pork. Meat Sci. 2002, 61, 405–410. [Google Scholar] [CrossRef]
- Chan, K.M.; Decker, E.A. Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 1994, 34, 403–426. [Google Scholar] [CrossRef]
- Filgueras, R.S.; Gatellier, P.; Aubry, L.; Thomas, A.; Bauchart, D.; Durand, D.; Zambiazi, R.C.; Santé-Lhoutellier, V. Colour, lipid and protein stability of Rhea americana meat during air- and vacuum-packaged storage: Influence of muscle on oxidative processes. Meat Sci. 2010, 86, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Pastsart, U.; De Boever, M.; Claeys, E.; De Smet, S. Effect of muscle and post-mortem rate of pH and temperature fall on antioxidant enzyme activities in beef. Meat Sci. 2013, 93, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, K.D.; Sabow, A.B.; Shittu, R.M.; Karim, R.; Karsani, S.A.; Sazili, A.Q. Impact of chill storage on antioxidant status, lipid and protein oxidation, color, drip loss and fatty acids of semimembranosus muscle in goats. CYTA—J. Food 2016, 14, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Imazaki, P.H.; Douny, C.; Elansary, M.; Scippo, M.L.; Clinquart, A. Effect of muscle type, aging technique, and aging time on oxidative stability and antioxidant capacity of beef packed in high-oxygen atmosphere. J. Food Process. Preserv. 2018, 42, e13603. [Google Scholar] [CrossRef]
- Lindmark-Månsson, H.; Chen, J.; Paulsson, M.; Aldén, G.; Ren, B.; Ladenstein, R. The effect of storage and heat treatment on glutathione peroxidase in bovine milk and whey. Int. Dairy J. 2001, 11, 71–81. [Google Scholar] [CrossRef]
- Hoac, T.; Daun, C.; Trafikowska, U.; Zackrisson, J.; Åkesson, B. Influence of heat treatment on lipid oxidation and glutathione peroxidase activity in chicken and duck meat, 2006. Innov. Food Sci. Emerg. Technol. 2006, 7, 88–93. [Google Scholar] [CrossRef]
- Yang, M.S.; Chan, H.W.; Yu, L.C. Glutathione peroxidase and glutathione reductase activities are partially responsible for determining the susceptibility of cells to oxidative stress. Toxicology 2006, 226, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Goto, M.; Abe, K.; Nakano, Y. Glutathione peroxidase activity during storage of fish muscle. J. Food Sci. 1996, 61, 734–735. [Google Scholar] [CrossRef]
- Daun, C.; Johansson, M.; Önning, G.; Akesson, B. Glutathione peroxidase activity, tissue and soluble selenium content in beef and pork in relation to meat ageing and pig RN phenotype. Food Chem. 2001, 73, 313–319. [Google Scholar] [CrossRef]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid–protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef]
- Gheisari, H.R.; Motamedi, H. Chloride salt type/ionic strength and refrigeration effects on antioxidant enzymes and lipid oxidation in cattle, camel and chicken meat. Meat Sci. 2010, 86, 377–383. [Google Scholar] [CrossRef]
- Fasseas, M.K.; Mountzouris, K.C.; Tarantilis, P.A.; Polissiou, M.; Zervas, G. Antioxidant activity in meat treated with oregano and sage essential oils. Food Chem. 2008, 106, 1188–1194. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horbańczuk, O.K.; Jóźwik, A.; Wyrwisz, J.; Marchewka, J.; Atanasov, A.G.; Wierzbicka, A. The Activity of Chosen Antioxidant Enzymes in Ostrich Meat in Relation to the Type of Packaging and Storage Time in Refrigeration. Biomolecules 2021, 11, 1338. https://doi.org/10.3390/biom11091338
Horbańczuk OK, Jóźwik A, Wyrwisz J, Marchewka J, Atanasov AG, Wierzbicka A. The Activity of Chosen Antioxidant Enzymes in Ostrich Meat in Relation to the Type of Packaging and Storage Time in Refrigeration. Biomolecules. 2021; 11(9):1338. https://doi.org/10.3390/biom11091338
Chicago/Turabian StyleHorbańczuk, Olaf K., Artur Jóźwik, Jarosław Wyrwisz, Joanna Marchewka, Atanas G. Atanasov, and Agnieszka Wierzbicka. 2021. "The Activity of Chosen Antioxidant Enzymes in Ostrich Meat in Relation to the Type of Packaging and Storage Time in Refrigeration" Biomolecules 11, no. 9: 1338. https://doi.org/10.3390/biom11091338