
Strontium Binding to α-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Expression and Purification of Human α-PA
2.3. Removal of Metal Ions from PA Samples
2.4. Fluorescence Measurements
2.5. Fluorimetric Evaluation of Ca2+/Sr2+ Affinity of PA
Protein + Me2+ ↔ Protein•Me2+
KMe2
Protein•Me2+ + Me2+ ↔ Me2+•Protein•Me2+,
EDTA + Me2+ ↔ EDTA•Me2+
Protein + Me2+ ↔ Protein•Me2+
2.6. ITC Measurements
2.7. FTIR Spectroscopy Measurements
2.8. Circular Dichroism Measurements
2.9. Scanning Calorimetry Measurements
2.10. Models Used in Theoretical Calculations
2.11. DFT/PCM Calculations (Density Functional Theory/Polarizable Continuum Model)
2.12. Validation of Calculations
3. Results
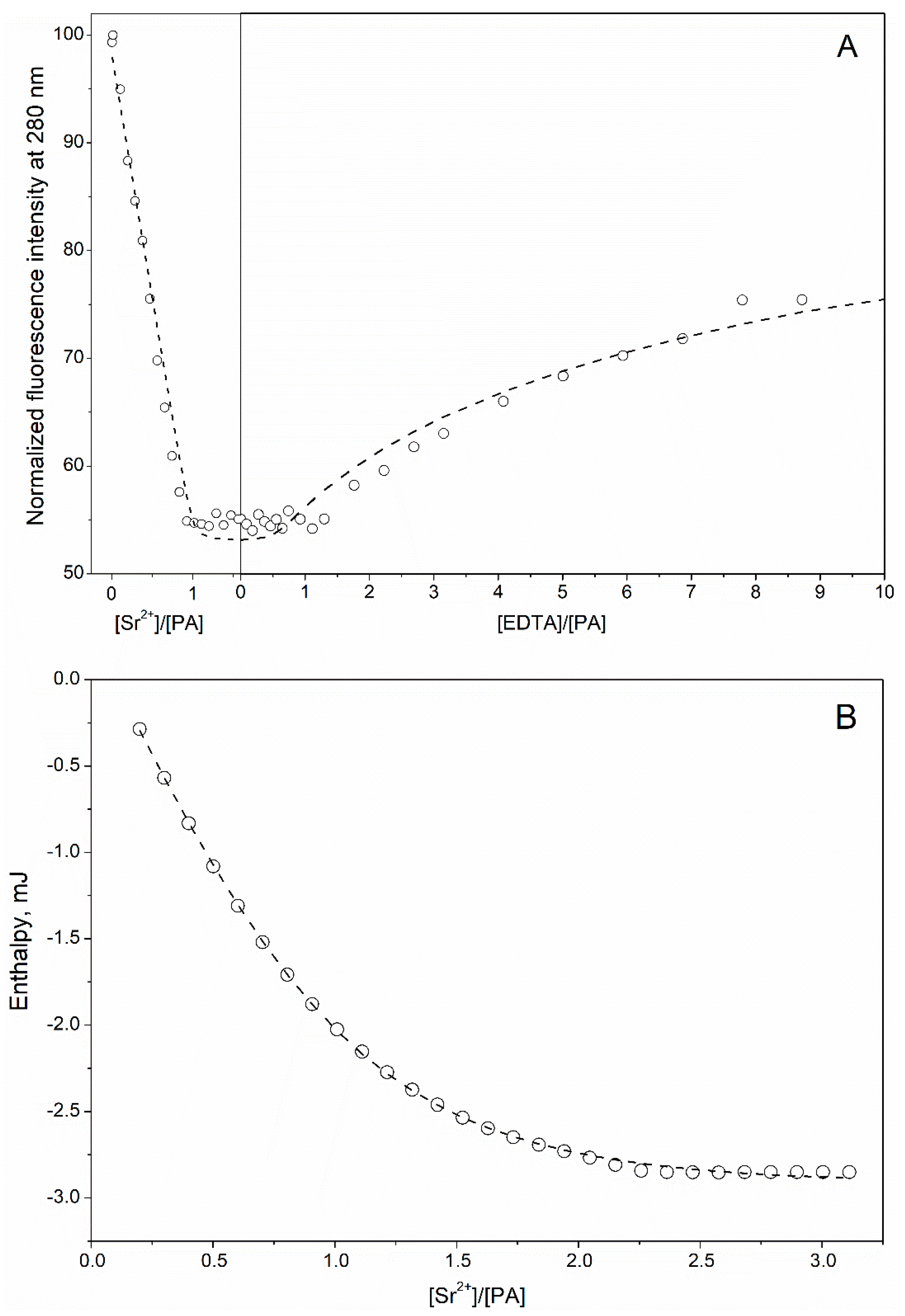
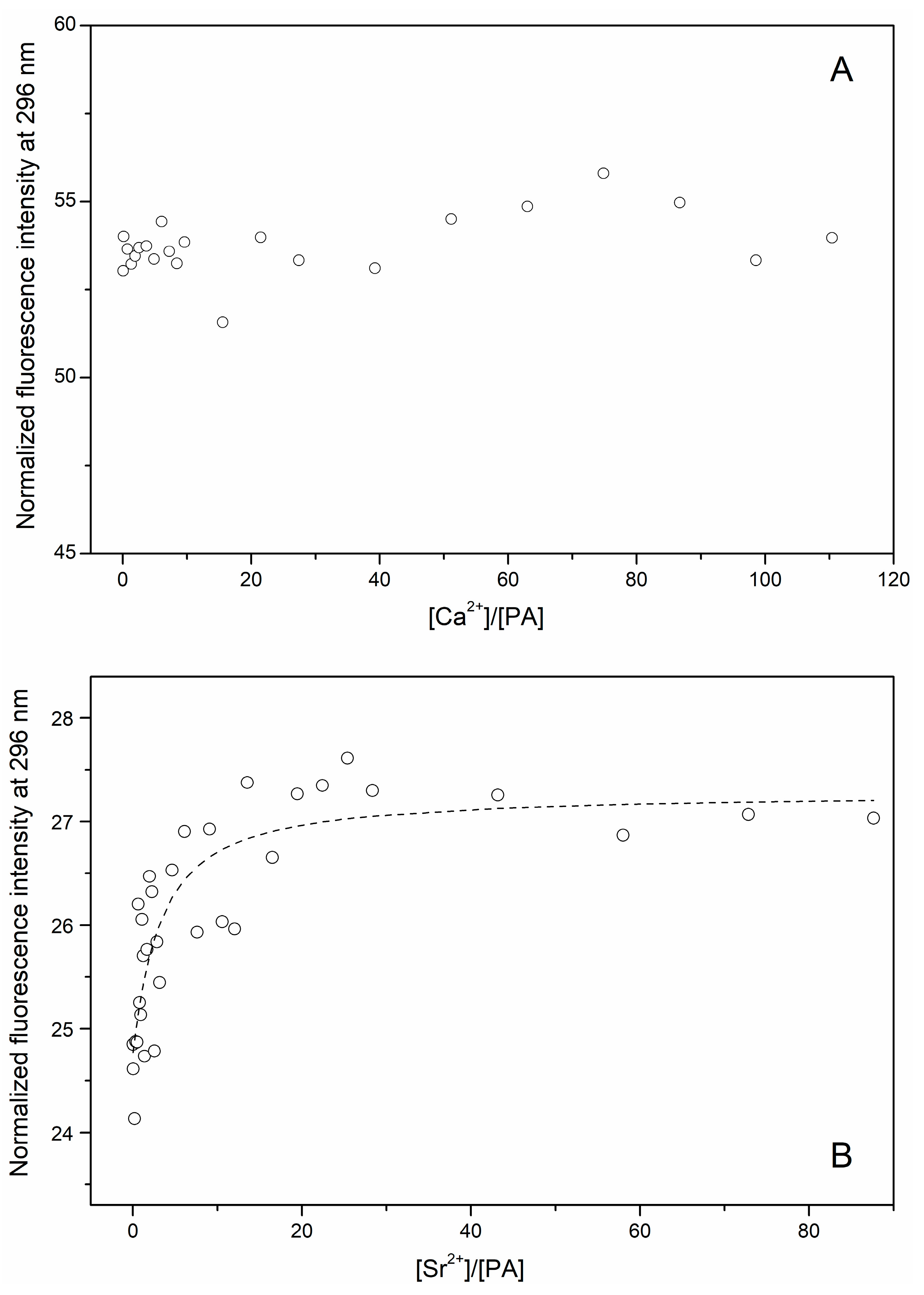
3.1. Ca2+/Sr2+-Binding Properties of Human α-Parvalbumin
3.2. Secondary Sr2+-Binding Site(s) of Human α-Parvalbumin
3.3. Comparison of Structural Peculiarities of Sr2+/Ca2+-Loaded States of α-PA
3.4. Molecular Modeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cabrera, W.E.; Schrooten, I.; De Broe, M.E.; D′Haese, P.C. Strontium and bone. J. Bone Miner. Res. 1999, 14, 661–668. [Google Scholar] [CrossRef]
- Cianferotti, L.; Gomes, A.R.; Fabbri, S.; Tanini, A.; Brandi, M.L. The calcium-sensing receptor in bone metabolism: From bench to bedside and back. Osteoporos. Int. 2015, 26, 2055–2071. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Ammann, P.; Boivin, G.; Rey, C. Mechanisms of action and therapeutic potential of strontium in bone. Calcif. Tissue Int. 2001, 69, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Pilmane, M.; Salma-Ancane, K.; Loca, D.; Locs, J.; Berzina-Cimdina, L. Strontium and strontium ranelate: Historical review of some of their functions. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 78, 1222–1230. [Google Scholar] [CrossRef]
- Geng, Y.; Mosyak, L.; Kurinov, I.; Zuo, H.; Sturchler, E.; Cheng, T.C.; Subramanyam, P.; Brown, A.P.; Brennan, S.C.; Mun, H.C.; et al. Structural mechanism of ligand activation in human calcium-sensing receptor. Elife 2016, 5, e13662. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.P. The biological role of strontium. Bone 2004, 35, 583–588. [Google Scholar] [CrossRef]
- Kolodziejska, B.; Stepien, N.; Kolmas, J. The Influence of Strontium on Bone Tissue Metabolism and Its Application in Osteoporosis Treatment. Int. J. Mol. Sci. 2021, 22, 6564. [Google Scholar] [CrossRef]
- Diepenhorst, N.A.; Leach, K.; Keller, A.N.; Rueda, P.; Cook, A.E.; Pierce, T.L.; Nowell, C.; Pastoureau, P.; Sabatini, M.; Summers, R.J.; et al. Divergent effects of strontium and calcium-sensing receptor positive allosteric modulators (calcimimetics) on human osteoclast activity. Br. J. Pharmacol. 2018, 175, 4095–4108. [Google Scholar] [CrossRef] [Green Version]
- Kraeber-Bodere, F.; Campion, L.; Rousseau, C.; Bourdin, S.; Chatal, J.F.; Resche, I. Treatment of bone metastases of prostate cancer with strontium-89 chloride: Efficacy in relation to the degree of bone involvement. Eur. J. Nucl. Med. 2000, 27, 1487–1493. [Google Scholar] [CrossRef]
- Shannon, R.D. Revised Effective Ionic Radii and Systematic Studies of Interatomic Distances in Halides and Chalcogenides. Acta Crystallogr. 1976, A32, 751–767. [Google Scholar] [CrossRef]
- Dudev, M.; Wang, J.; Dudev, T.; Lim, C. Factors Governing the Metal Coordination Number in Metal Complexes from Cambridge Structural Database Analysis. J. Phys. Chem. 2006, B110, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Hofer, T.S.; Randolf, B.R.; Rode, B.M. Sr(II) in Water: A Labile Hydrate with a Highly Mobile Structure. J. Phys. Chem. 2006, 110, 20409–20417. [Google Scholar] [CrossRef]
- Saidak, Z.; Marie, P.J. Strontium signaling: Molecular mechanisms and therapeutic implications in osteoporosis. Pharmacol. Ther. 2012, 136, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M. Is the calcium receptor a molecular target for the actions of strontium on bone ? Osteoporos. Int. 2003, 14, 25–34. [Google Scholar] [CrossRef]
- Encyclopedia of metalloproteins; Springer: New York, NY, USA, 2013.
- Nockolds, C.E.; Kretsinger, R.H.; Coffee, C.J.; Bradshaw, R.A. Structure of a calcium-binding carp myogen. Proc. Natl. Acad. Sci. USA 1972, 69, 581–584. [Google Scholar] [CrossRef] [Green Version]
- Permyakov, E.A.; Kretsinger, R.H. Calcium Binding Proteins; A John Wiley & Sons, Inc.: New Jersey, NJ, USA, 2011. [Google Scholar]
- Permyakov, E.A.; Uversky, V.N.; Permyakov, S.E. Parvalbumin as a pleomorphic protein. Curr. Protein Pept. Sci. 2016, 18, 780–794. [Google Scholar] [CrossRef] [PubMed]
- Permyakov, E.A. Parvalbumin; Nova Science Publishers, Inc.: New York, NY, USA, 2006. [Google Scholar]
- Vologzhannikova, A.A.; Khorn, P.A.; Kazakov, A.S.; Ismailov, R.G.; Sokolov, A.S.; Uversky, V.N.; Permyakov, E.A. In search for globally disordered apo-parvalbumins: Case of parvalbumin β-1 from coho salmon. Cell Calcium 2017, 67, 53–640. [Google Scholar] [CrossRef] [PubMed]
- Rhyner, J.A.; Durussel, I.; Cox, J.A.; Ilg, E.C.; Schafer, B.W.; Heizmann, C.W. Human recombinant alpha-parvalbumin and nine mutants with individually inactivated calcium- and magnesium-binding sites: Biochemical and immunological properties. Biochim. Biophys. Acta 1996, 1313, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Laible, M.; Boonrod, K. Homemade site directed mutagenesis of whole plasmids. J. Vis. Exp. 2009, 11, 1135. [Google Scholar] [CrossRef] [Green Version]
- Narita, K. Isolation of acetylpeptide from enzymic digests of TMV-protein. Biochim. Biophys. Acta 1958, 28, 184–191. [Google Scholar] [CrossRef]
- Schwaller, B. The use of transgenic mouse models to reveal the functions of Ca(2+) buffer proteins in excitable cells. Biochim. Biophys. Acta 2012, 1820, 1294–1303. [Google Scholar] [CrossRef]
- Blum, H.E.; Lehky, P.; Kohler, L.; Stein, E.A.; Fischer, E.H. Comparative properties of vertebrate parvalbumins. J. Biol. Chem. 1977, 252, 2834–2838. [Google Scholar] [CrossRef]
- Permyakov, S.E.; Bakunts, A.G.; Denesyuk, A.I.; Knyazeva, E.L.; Uversky, V.N.; Permyakov, E.A. Apo-parvalbumin as an intrinsically disordered protein. Proteins 2008, 72, 822–836. [Google Scholar] [CrossRef]
- Permyakov, S.E.; Vologzhannikova, A.A.; Emelyanenko, V.I.; Knyazeva, E.L.; Kazakov, A.S.; Lapteva, Y.S.; Permyakova, M.E.; Zhadan, A.P.; Permyakov, E.A. The impact of alpha-N-acetylation on structural and functional status of parvalbumin. Cell Calcium 2012, 52, 366–376. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Ostrovsky, A.V.; Kalinichenko, L.P. Stopped-flow kinetic studies of Ca(II) and Mg(II) dissociation in cod parvalbumin and bovine alpha-lactalbumin. Biophys. Chem. 1987, 28, 225–233. [Google Scholar] [CrossRef]
- Schwarzenbach, G.; Flaschka, H. Die Komplexometrische Titration; Ferdinand Enke Verlag: Stuttgart, Germany, 1965. [Google Scholar]
- Marquardt, D.W. An algorithm for least-squares estimation of nonlinear parameters. J. Soc. Indust. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Sreerama, N.; Venyaminov, S.Y.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Inclusion of denatured proteins with native proteins in the analysis. Anal. Biochem. 2000, 287, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Makhatadze, G.I.; Privalov, P.L. Heat capacity of proteins. I. Partial molar heat capacity of individual amino acid residues in aqueous solution: Hydration effect. J. Mol. Biol. 1990, 213, 375–384. [Google Scholar] [CrossRef]
- Permyakov, S.E.; Bakunts, A.G.; Permyakova, M.E.; Denesyuk, A.I.; Uversky, V.N.; Permyakov, E.A. Metal-controlled interdomain cooperativity in parvalbumins. Cell Calcium 2009, 46, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Declercq, J.P.; Tinant, B.; Parello, J.; Rambaud, J. Ionic interactions with parvalbumins. Crystal structure determination of pike 4.10 parvalbumin in four different ionic environments. J. Mol. Biol. 1991, 220, 1017–1039. [Google Scholar] [CrossRef]
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-class functionals and 12 other functionals. Theor. Chem. Acc. 2006, 120, 215–241. [Google Scholar]
- Dudev, T.; Cheshmedzhieva, D.; Doudeva, L. Competition between abiogenic Al3+ and native Mg2+, Fe2+ and Zn2+ ions in protein binding sites: Implications for aluminum toxicity. J. Mol. Modeling 2018, 24, 55. [Google Scholar] [CrossRef]
- Dudev, T.; Nikolova, V. Determinants of Fe2+ over M2+ (M = Mg, Mn, Zn) Selectivity in Non-Heme Iron Proteins. Inorg. Chem. 2016, 55, 12644–12650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaussian 09; Revision A.02; Gaussian, Inc.: Wallingford, CT, USA, 2016.
- Zheng, J.; Truhlar, D.G. Including Torsional Anharmonicity in Canonical and Microcanonical Reaction Path Calculations. J. Chem. Theory Comput. 2013, 9, 2875–2881. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Determinants of K+ vs Na+ Selectivity in potassium channels. J. Am. Chem. Soc. 2009, 131, 8092–8101. [Google Scholar] [CrossRef]
- Ambjørn, M.; Ejlerskov, P.; Liu, Y.; Lees, M.; Jäättelä, M.; Issazadeh-Navikas, S. IFNB1/interferon-β-induced autophagy in MCF-7 breast cancer cells counteracts its proapoptotic function. Autophagy 2013, 9, 287–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertz, E.L.; Krishtalik, L.I. Low dielectric response in enzyme active site. Proc. Natl. Acad. Sci. USA 2000, 97, 2081–2086. [Google Scholar] [CrossRef] [Green Version]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal solvation model based on solute electron density and on a continuum model of the solvent defined by the bulk dielectric constant and atomic surface tensions. J. Phys. Chem. 2009, 113, 6378–6396. [Google Scholar] [CrossRef]
- Marcus, Y. Thermodynamics of solvation of ions. 5. Gibbs free energy of hydration at 298.15 K. J. Chem. Soc. Faraday Trans. 1991, 87, 2995–2999. [Google Scholar] [CrossRef]
- Smith, R.M.; Martell, A.E. Critical stability constants, enthalpies and entropies for the formation of metal complexes of aminopolycarboxylic acids and carboxylic acids. Sci. Total Environ. 1987, 64, 125–147. [Google Scholar] [CrossRef]
- Barth, A. The infrared absorption of amino acid side chains. Prog. Biophys. Mol. Biol. 2000, 74, 141–173. [Google Scholar] [CrossRef]
- Permyakov, S.E. Differential scanning microcalorimetry of intrinsically disordered proteins. Methods Mol. Biol. 2012, 896, 283–296. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Calcium Ion Selectivity in Biological Systems. In Encyclopedia of Metalloproteins; Uversky, V.N., Kretsinger, R.H., Permyakov, E.A., Eds.; Springer Science: New York, NY, USA, 2013; pp. 478–484. [Google Scholar]
- Permyakov, E.A.; Kalinichenko, L.P.; Medvedkin, V.N.; Burstein, E.A.; Gerday, C. Sodium and potassium binding to parvalbumins measured by means of intrinsic protein fluorescence. Biochim. Biophys. Acta 1983, 749, 185–191. [Google Scholar] [CrossRef]
- Permyakov, S.E.; Cherskaya, A.M.; Senin, I.I.; Zargarov, A.A.; Shulga-Morskoy, S.V.; Alekseev, A.M.; Zinchenko, D.V.; Lipkin, V.M.; Philippov, P.P.; Uversky, V.N.; et al. Effects of mutations in the calcium-binding sites of recoverin on its calcium affinity: Evidence for successive filling of the calcium binding sites. Protein Eng. 2000, 13, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Chandra, K.; Ramakrishnan, V.; Sharma, Y.; Chary, K.V. N-terminal myristoylation alters the calcium binding pathways in neuronal calcium sensor-1. J. Biol. Inorg. Chem. 2011, 16, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Yamniuk, A.P.; Nguyen, L.T.; Hoang, T.T.; Vogel, H.J. Metal ion binding properties and conformational states of calcium- and integrin-binding protein. Biochemistry 2004, 43, 2558–2568. [Google Scholar] [CrossRef] [PubMed]
- Kiran, U.; Regur, P.; Kreutz, M.R.; Sharma, Y.; Chakraborty, A. Intermotif Communication Induces Hierarchical Ca(2+) Filling of Caldendrin. Biochemistry 2017, 56, 2467–2476. [Google Scholar] [CrossRef]
- Shirran, S.L.; Barran, P.E. The use of ESI-MS to probe the binding of divalent cations to calmodulin. J. Am. Soc. Mass. Spectrom. 2009, 20, 1159–1171. [Google Scholar] [CrossRef]
- Schaer, J.J.; Milos, M.; Cox, J.A. Thermodynamics of the binding of calcium and strontium to bovine alpha-lactalbumin. FEBS Lett. 1985, 190, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Blancuzzi, Y.; Padilla, A.; Parello, J.; Cave, A. Symmetrical rearrangement of the cation-binding sites of parvalbumin upon Ca2+/Mg2+ exchange. A study by 1H 2D NMR. Biochemistry 1993, 32, 1302–1309. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ca2+ | ||
|---|---|---|
| Protein | Fluorimetric titrations | |
| K1, M−1 | K2, M−1 | |
| rWT | 4 × 109 | 4 × 109 |
| PA-CD | 5 × 104 | |
| PA-EF | 4 × 108 | |
| Sr2+ | ||
| Protein | Fluorimetric titrations/ITC | |
| K1, M−1 | K2, M−1 | |
| rWT | 3 × 107/(2 × 107 and 2 × 106) | |
| PA-CD | 1 × 106 | |
| PA-EF | 2 × 106 | |
| Protein State | α-Helices, % | β-Sheets, % | Turns, % | Random Coil, % |
|---|---|---|---|---|
| apo | 30.6 ± 0.2 | 17.6 ± 0.3 | 19.6 ± 0.3 | 31.50 ± 0.10 |
| Sr2+ | 50.1 ± 0.2 | 7.4 ± 0.3 | 15.5 ± 0.3 | 26.80 ± 0.10 |
| Ca2+ | 48.2 ± 0.2 | 8.6 ± 0.3 | 15.7 ± 0.3 | 27.30 ± 0.10 |
| T0, °C | ΔH0, J/g | ΔCp, J/(g·K) | n |
|---|---|---|---|
| 86.0 | 37.5 | 0.40 | 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vologzhannikova, A.A.; Shevelyova, M.P.; Kazakov, A.S.; Sokolov, A.S.; Borisova, N.I.; Permyakov, E.A.; Kircheva, N.; Nikolova, V.; Dudev, T.; Permyakov, S.E. Strontium Binding to α-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family. Biomolecules 2021, 11, 1158. https://doi.org/10.3390/biom11081158
Vologzhannikova AA, Shevelyova MP, Kazakov AS, Sokolov AS, Borisova NI, Permyakov EA, Kircheva N, Nikolova V, Dudev T, Permyakov SE. Strontium Binding to α-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family. Biomolecules. 2021; 11(8):1158. https://doi.org/10.3390/biom11081158
Chicago/Turabian StyleVologzhannikova, Alisa A., Marina P. Shevelyova, Alexey S. Kazakov, Andrey S. Sokolov, Nadezhda I. Borisova, Eugene A. Permyakov, Nikoleta Kircheva, Valya Nikolova, Todor Dudev, and Sergei E. Permyakov. 2021. "Strontium Binding to α-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family" Biomolecules 11, no. 8: 1158. https://doi.org/10.3390/biom11081158