
Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years


Faculty of Pharmaceutical Sciences, Sanyo-Onoda City University, Yamaguchi 756-0884, Japan
Biomolecules 2021, 11(6), 896; https://doi.org/10.3390/biom11060896
Submission received: 20 May 2021
/
Revised: 14 June 2021
/
Accepted: 14 June 2021
/
Published: 16 June 2021
(This article belongs to the Special Issue Hydrogen Sulfide and Polysulfides, Endogenous Mammalian Transmitters——Honorary Special Issue Commemorating the Work of Prof. Hideo Kimura)
{kind=link}
{kind=link}
Abstract
:Since the first description of hydrogen sulfide (H2S) as a toxic gas in 1713 by Bernardino Ramazzini, most studies on H2S have concentrated on its toxicity. In 1989, Warenycia et al. demonstrated the existence of endogenous H2S in the brain, suggesting that H2S may have physiological roles. In 1996, we demonstrated that hydrogen sulfide (H2S) is a potential signaling molecule, which can be produced by cystathionine β-synthase (CBS) to modify neurotransmission in the brain. Subsequently, we showed that H2S relaxes vascular smooth muscle in synergy with nitric oxide (NO) and that cystathionine γ-lyase (CSE) is another producing enzyme. This study also opened up a new research area of a crosstalk between H2S and NO. The cytoprotective effect, anti-inflammatory activity, energy formation, and oxygen sensing by H2S have been subsequently demonstrated. Two additional pathways for the production of H2S with 3-mercaptopyruvate sulfurtransferase (3MST) from l- and d-cysteine have been identified. We also discovered that hydrogen polysulfides (H2Sn, n ≥ 2) are potential signaling molecules produced by 3MST. H2Sn regulate the activity of ion channels and enzymes, as well as even the growth of tumors. S-Sulfuration (S-sulfhydration) proposed by Snyder is the main mechanism for H2S/H2Sn underlying regulation of the activity of target proteins. This mini review focuses on the key findings on H2S/H2Sn signaling during the first 25 years.
1. Identification of H2S as a Signaling Molecule
Patients that recover from H2S poisoning show cognitive decline, and the levels of neurontransmitters in the brains of animals exposed to H2S change, suggesting that the brain is vulnerable to H2S toxicity [1]. Warenycia et al. measured the levels of H2S accumulated in the brain of rats exposed to H2S when they discovered a certain amount of H2S in the brain even without exposure to H2S [2]. Although the concentrations were overestimated, the existence of endogenous H2S was identified in the brain.
Pyridoxal 5′-phosphate-dependent enzymes, cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE), have been suggested to regulate several pathways. CBS catalyzes the first step of the transsulfuration pathway in which cystathionine is produced from serine and homocysteine, and cystathionine is further catalyzed by CSE to cysteine. An alternate pathway exists in which CBS catalyzes the condensation of cysteine with homocysteine to generate cystathionine and H2S [3,4]. CSE catalyzes an elimination reaction which metabolizes cysteine to pyruvate, NH3, and H2S [3,4]. However, rather than being recognized as a physiologically active molecule, in these early studies, H2S was merely thought to be a byproduct of the metabolic pathways.
The observations that H2S is produced by enzymes and exists in the brain prompted us to study a physiological role of this molecule. The activities of CBS and CSE have been intensively studied in the liver and kidney, but little is known about them in the brain. We found CBS in the brain and confirmed the production of H2S, which is augmented by S-adenosyl methionine (SAM) [5].
Other gaseous signaling molecules NO and carbon monoxide (CO) induce hippocampal long-term potentiation (LTP), a synaptic model of memory formation, as retrograde messengers, which are produced at postsynapse and released to presynapse to facilitate a release of a neurotransmitter glutamate from presynapse [6,7,8,9,10]. We examined whether or not H2S has a similar effect. H2S facilitated the induction of LTP by enhancing the activity of N-methyl-d-aspartate (NMDA) receptors but not as a retrograde messenger [5].
NMDA receptors are activated by a reducing substance dithiothreitol (DTT) through the reduction of a cysteine disulfide bond located at the hinge of the ligand-binding domain [11]. Because H2S is a reducing substance, it is likely to be a mechanism for facilitating the induction of LTP. However, H2S with one-tenth of the concentration of DTT exerted a greater effect than that of DTT [5]. This observation suggested that there is an additional mechanism for LTP induction by H2S. The prominent neuroscientist Solomon Snyder commented the following in Science News: “They have very impressive evidence that H2S is a potential neurotransmitter. It is an exciting paper that should stimulate a lot of people’s interest” [12].
The synaptic transmission is regulated not only by events at synapses such as a release of transmitters and the sensitivity of receptors but also by astrocytes, a type of glia, which surround synapses. Astrocytes release gliotransmitters to regulate synaptic activity. We found that H2S induces Ca2+ influx in astrocytes, which was greatly suppressed by La3+, Gd3+, and ruthenium red, broad-spectrum inhibitors known for transient receptor potential (TRP) channels, suggesting that H2S activates TRP channels [13]. H2S was reported to activate TRPA1 channels in urinary bladder and in sensory neurons, but concentrations greater than 1 mM were required for inducing responses [14,15].
2. Identification of H2Sn as Signaling Molecules
During this study, we found that a batch of NaHS, i.e., the sodium salt of H2S, with yellowish color was much more potent than the colorless batch. We successfully reproduced a solution with a similar color by dissolving elemental sulfur into Na2S solution according to a report by Searcy and Lee [16]. The color came from H2Sn, which induces Ca2+ influx in astrocytes much more potently than H2S [17,18,19]. H2Sn are natural inorganic polymeric sulfur–sulfur species or sulfane sulfur, which we later found to be produced by 3-mercaptopyruvate sulfurtransferase (3MST) from 3-mercaptopyruvate [20,21,22] and the partial oxidation of H2S [19], such as via the chemical interaction with NO [23,24]. H2S2 (2.6 µM) exists in the brain almost equivalent to the level of H2S (3 µM) [25]. Ca2+ influx induced in astrocytes by AITC, cinnamaldehyde, selective activators of TRPA1 channels, and Na2S3 was greatly suppressed by HC030031 and AP-18, selective inhibitors of TRPA1 channels. In astrocytes transfected with TRPA1-siRNA, Ca2+ influx was not efficiently induced by Na2S3 [19]. The EC50 value for H2S was 116 µM, while that for H2S3 was 91 nM, suggesting that H2Sn rather than H2S are ligands for TRPA1 channels [13,17,18,19]. The amino terminus of TRPA1 channels has 24 cysteine residues [26], and two cysteine residues Cys422 and Cys634 are sensitive to H2Sn [27].
S-Sulfuration (S-sulfuhydration) was proposed by Snyder and colleagues to regulate the activity of target proteins by H2S [28]. This proposal needs a minor revision to highlight H2Sn but not H2S S-sulfurate cysteine residues. In contrast, H2S S-sulfurates oxidized cysteine residues such as those S-nitrosylated and S-sulfenylated [29]. H2Sn S-sulfurate (S-sulfhydrate) two cysteine residues of TRPA1 channels to induce the conformational changes to activate the channels. As an alternative mechanism, one cysteine residue, which is S-sulfurated, reacts with the remaining cysteine residue to generate a cysteine disulfide bond. Although the conformation has not been examined in detail, the latter mechanism may induce conformational changes more efficiently than the former one.
Various target proteins of H2Sn have been identified such as a tumor suppressor phosphate and tensin homolog (PTEN), protein kinase G1α, and an enzyme responsible for glycolysis, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) [28,30,31,32]. It has been reported that GAPDH is activated by H2S through S-sulfuration of the active site Cys150 [28], while it is suppressed by H2Sn through S-sulfuration of Cys156, which is not the active site [32]. Cys150 may be an oxidized residue when S-nitrosylated or S-sulfenylated, which can be S-sulfurated by H2S, while Cys156 must be a thiol, which is S-sulfurated by H2Sn.
3. Synergy and Crosstalk between H2S and NO
H2S relaxes vascular smooth muscle in synergy with NO [33]. A similar result was also obtained in the ileum [34]. Whiteman et al. proposed that the chemical interaction of H2S with NO generate nitrosothiol, which releases NO in the presence of Cu2+ [35]. Filipovic et al. reported that H2S and NO produces nitroxyl (HNO) as a major product, as well as H2Sn [36,37], while Cortese-Krott et al. suggested that SSNO− as a major product with H2Sn as a minor one [38]. We proposed that H2Sn are major products [23]. The effect of H2Sn and that of the products obtained from the mixture of Na2S and diethylamine NONOate, an NO donor, were eliminated when they were exposed to cyanide or DTT [23]. In contrast, HNO is resistant to cyanide, and SSNO− is resistant to DTT. Based on these observations, H2Sn are potential chemical entities produced from H2S and NO [23,37,38]. Bogdandi et al. recently suggested that H2Sn transiently activate TRPA1 channels at the early phase of the production from H2S and NO, while the more stable product SSNO− sustainably activates the channels [39].
4. Vascular Tone Regulation by H2S and H2Sn
Since H2S relaxes vascular smooth muscle in synergy with NO [33] and activates ATP-dependent K+ (KATP) channels [40], it has been suggested that H2S is a potential endothelial-derived hyperpolarizing factor (EDHF), which is a component of endothelial-derived relaxing factor (EDRF) [41]. However, previous studies showed that the hyperpolarization induced by EDHF is resistant to glibenclamide, a KATP channel blocker [42,43]. The relaxation of vascular smooth muscle in the mesenteric bed, which is mediated predominantly by EDHF, is rather abolished by apamine, a blocker of Ca2+-activated K+ channels [44].
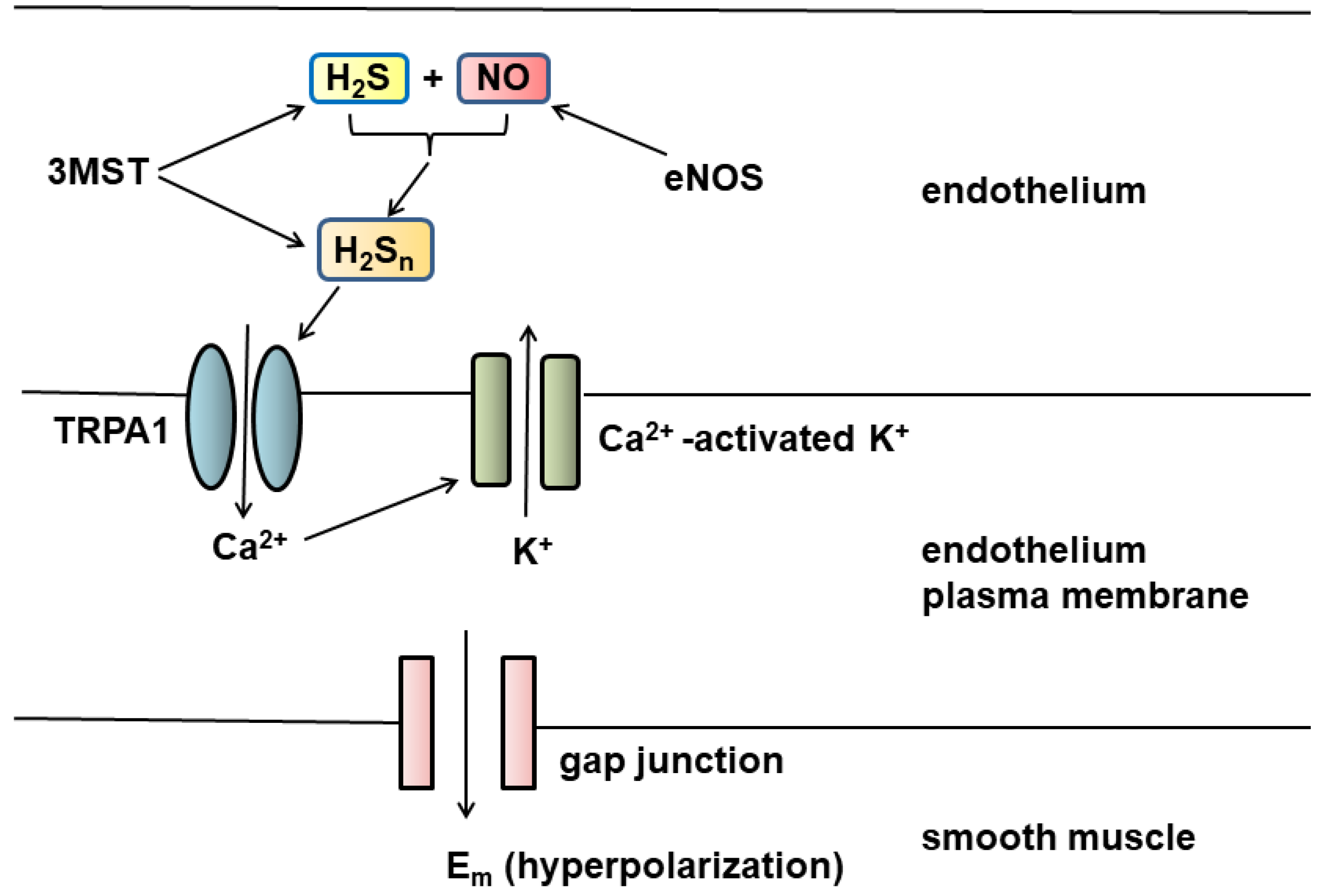
H2Sn are potential EDHFs (Figure 1). H2Sn produced by 3MST together with cysteine aminotransferase (CAT), both of which are localized to the vascular endothelium [20,45,46], or H2Sn generated by the chemical interaction between H2S and NO produced by endothelial NO synthase (eNOS) can activate TRPA1 channels [19,23] localized to myoendothelial junctions. The channels induce Ca2+ influx, which activate Ca2+-activated K+ channels to hyperpolarize the endothelial cell plasma membrane. The change in membrane potential is conducted via myoendothelial gap junctions to hyperpolarize the vascular smooth muscle [47].
H2S has also been demonstrated to relax vascular smooth muscle via the protein kinase G pathway as an endogenous inhibitor of phosphodiesterase and increases the levels of both cyclic GMP and cyclic AMP [48,49], as well as by activating Kv7 potassium channels [50]. Kv7 channels are also involved in CBS-derived H2S induced human malignant hyperthermia syndrome triggered by volatile inhalation anesthetics in skeletal muscle [51].
5. Cytoprotective Effect of H2S, H2Sn, and H2SO3
The impression of H2S as toxic gas led to its cytoprotective effect being overlooked [52]. Expecting that all cells would be killed by H2S, I applied NaHS to cells and incubated for overnight. On the contrary, cells were lively and survived from the toxin. H2S increases the production of glutathione (GSH), a major intracellular antioxidant, by enhancing the activity of cystine/glutamate antiporter, which incorporates cystine into cells, and of glutamate cysteine ligase (GCL), a rate-limiting enzyme for GSH production [52,53]. H2S also facilitates the translocation of GSH into mitochondria [53]. The protective activity of H2S is also exerted through the stabilization of membrane potential by enhancing the activity of KATP channels and cystic fibrosys transmembrane conductance regulator (CFTR) Cl− channels [54]. Lefer and colleagues demonstrated that H2S protects the heart from ischemia/reperfusion injury by preserving mitochondrial function [55].
H2Sn S-sulfurate Keap1 and release Nrf2 from the Keap1/Nrf2 complex to the nucleus, where Nrf2 upregulates antioxidant genes including the GCL gene to increase the production of GSH [56]. H2Sn increase the levels of GSH and protect cells from oxidative stress to a greater level than H2S [57]. Sulfite (H2SO3), a metabolite of H2S and H2Sn, protects neurons by increasing the production of GSH as efficiently as the parental molecules [57].
6. Signaling by H2S, H2Sn through S-Sulfuration and Bound Sulfane Sulfur
In addition to CBS and CSE, 3MST, along with CAT or DAO, was recognized to produce H2S from l- or d-cysteine, respectively [46,58,59]. Subsequently, 3MST was found to produce H2Sn and other S-sulfurated molecules such as cysteine persulfide, GSSH, and S-sulfurated cysteine residues [20,21,60]. Other enzymes such as sulfide-quinone oxidoreductase (SQR), haemoglobin, neuroglobin, catalase, super oxide dismutase (SOD), cysteine tRNA synthetase (CARS), and peroxidases have been identified to produce H2Sn and other S-sulfurated molecules [61,62,63,64,65,66,67,68,69].
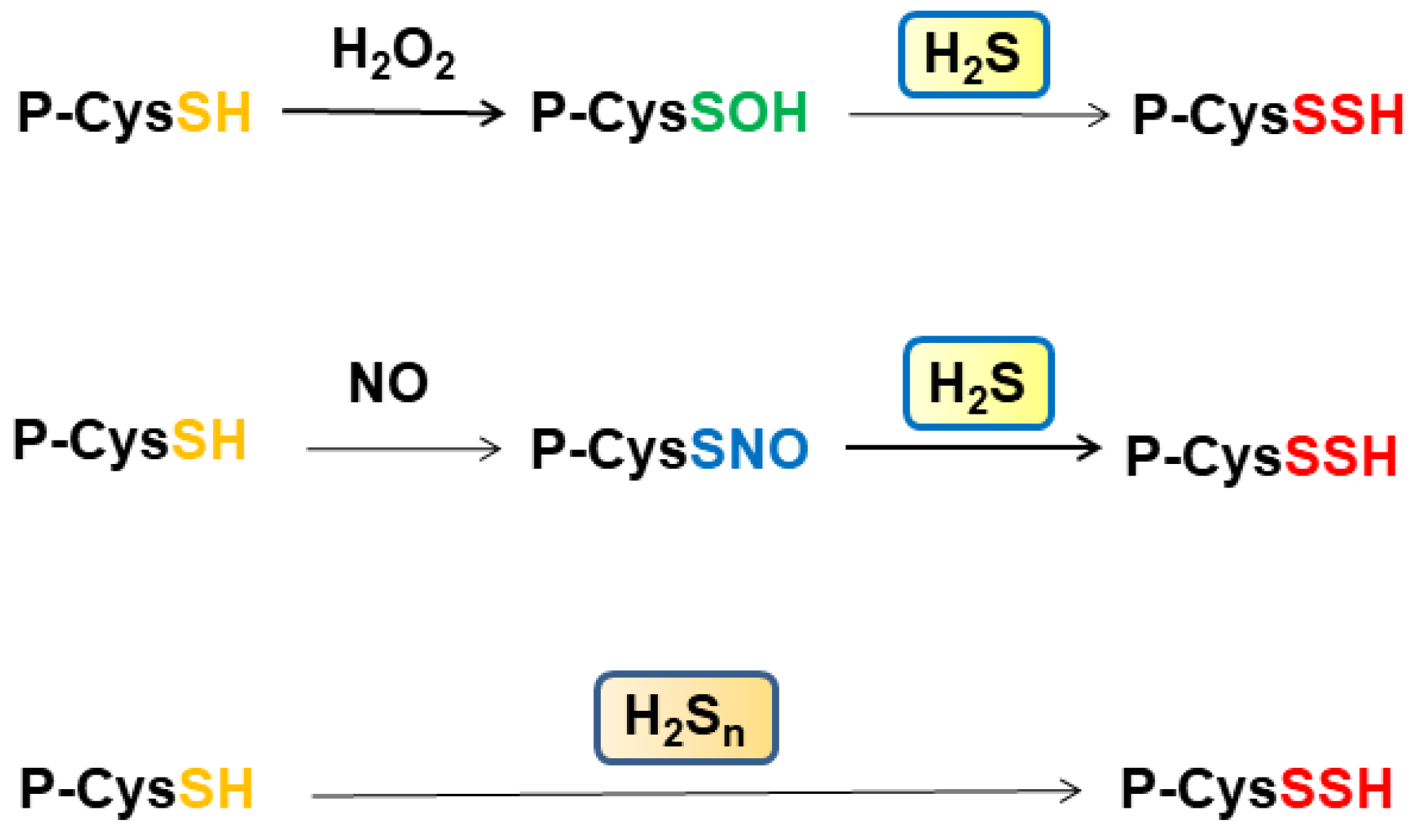
In total, 10–20% of cysteine residues of proteins are S-sulfurated [28], also observed as a part of bound sulfane sulfur, which releases H2S under reducing conditions, including H2Sn, cysteine persulfide, GSSH, and S-sulfurated cysteine residues [70,71,72,73]. In cells and tissues, 5–12% of total protein cysteine residues are oxidized, such as S-nitrosylated (P-CysSNO) and S-sulfenylated (P-CysSOH), and this can be increased to more than 40% under oxidative conditions [74] (Figure 2). The amount of bound sulfane sulfur and its associated species is distinct among tissues. For example, heart homogenates release H2S under reducing conditions much less than those from the liver and the brain, while heart homogenates absorb H2S as fast as liver homogenates [73]. P-CysSNO and P-CysSOH react with H2S to generate P-CysSSH, while they do not release H2S under reducing conditions. These observations suggest that the heart may contain P-CysSNO and P-CysSOH more abundantly than the liver and the brain.
Some cysteine residues are oxidized by H2O2 to generate S-nitrosylated cysteine residues, and some others are S-nitrosylated by NO. These oxidized cysteine residues are S-sulfurated by H2S rather than H2Sn (Figure 2). Cys150 and Cys156 of GAPDH may be in different oxidation states, as described previously [28,32]. Zivanovic et al. demonstrated that the activity of manganese superoxide dismutase is suppressed through S-sulfenylation by H2O2, while its activity is recovered by H2S, which S-sulfurates the S-sulfenylated cysteine residues [75]. The same group showed that epidermal growth factor (EGF) activates its receptor, in which the levels of S-sulfenylated cysteine residues are increased at the early phase, while those of S-sulfurated residues are increased at the late phase when the expression of H2S-producing enzymes is enhanced. H2S S-sulfurates those S-sulfenylated cysteine residues to regulate their activity (Figure 2).
Another role of S-sulfuration is that it enables proteins to recover their functions from over-oxidization. Sulfinic (P-CysSO2H) and sulfonic acids (P-CysSO3H) are not reduced back to P-CysSH by thioredoxin and deteriorate the protein function. In contrast, S-sulfurated proteins P-CysSSO2H and P-CysSSO3H can be reduced by thioredoxin to P-CysSH [75,76].
7. Diseases Caused by the Disturbance of H2S and H2Sn
Both an excess and a deficiency of H2S and H2Sn have been suggested in the pathogenesis of schizophrenia. Thiol homeostasis is shifted to oxidized conditions, reflecting significantly less H2S and more disulfide bond formation in patients than normal individuals [77,78]. In contrast, we suggested that excess H2S and H2Sn are involved in the pathogenesis. Mice with high expression of 3MST impaired prepulse inhibition, an endophenotype for schizophrenia, and 3MST levels were positively correlated with symptom severity scores [79].
CBS and H2S may be involved in regulating proliferation and bioenergetics in breast cancer, ovarian cancer, and colorectal cancer [80,81,82], and high levels of CBS, CSE, and 3MST expression were observed in lung cancer [83].
Gliomas with the highest grades of malignancy contained greater levels of polysulfides than glioma-free brain regions [84], and H2Sn levels were greater in glioblastoma-bearing regions than glioblastoma-free control regions [85]. In contrast, it was reported that CBS is involved in suppressing glioma, whereby glioma with suppressed CBS had high levels of VEGF and HIF-2α and was deeply invaded with dense vascularization and aggressive growth [86].
Parkinson’s disease is a neurodegenerative disorder. Parkin, an E3 ubiquitin ligase responsible for the clearance of misfolded proteins, is suppressed in this disease. Specific cysteine residues of parkin are S-nitrosylated in patients, while they are S-sulfurated in the normal individuals [87]. H2S may be involved in S-sulfurating the S-nitrosylated cysteine residues of parkin.
Down’s syndrome (DS) is characterized by impaired brain growth and maturation that causes mental retardation and is associated with an Alzheimer’s type of dementia in elderly adults. DS involves a trisomy of chromosome 21 where CBS is encoded, and its mRNA level is 12 times greater in myeloblasts of DS children, while CBS protein levels in the brains of patients are approximately three times greater compared to the normal individuals [88,89]. Higher levels of thiosulfate, a metabolite of H2S, were detected in patients [90]. Mice with CBS overexpression showed DS-like neurocognitive deficits [91]. Fibroblasts prepared from DS patients showed profound suppression of mitochondrial electron transport, oxygen consumption, and ATP generation [92].
Ethylmalonyl encephalopathy is an autosomal recessive early-onset disorder, defective in cytochrome c oxidase in the brain and muscle. In this disease, ETHE1, a gene encoding sulfur dioxygenase, which metabolizes H2S in collaboration with SQR, is deficient, and H2S levels are increased to suppress cytochrome c oxidase [93]. High levels of H2S and persulfides also suppress acyl-protein thioesterase in the mouse model of this disease [94,95].
The brain is very sensitive to oxygen deprivation, i.e., hypoxia. During hypoxia, heme oxygenase-2 produces less carbon monoxide, which suppresses the activity of CBS, resulting in the overproduction of H2S that stimulates the carotid body to increase in respiratory rate, heart rate, and blood pressure [96]. On the other hand, the increased levels of H2S, in turn, suppress cytochrome c oxidase to cause hypoxic brain injury [97]. The levels are augmented in SQR-deficient animal models [97].
8. Perspective
Cysteine residues of the target proteins initially found, such as TRPA1 channels, PTEN, and protein kinase G1α, may be thiols whose sulfur has the oxidation state of −2, and that of H2Sn has the oxidation state of −1 or 0 and is able to S-sulfurate thiols to regulate the activity of the targets. On the basis of these observations, S-sulfurated molecules including H2Sn have been recognized as the chemical entities as signaling molecules rather than H2S, which is thought to be a mere precursor of H2Sn or a byproduct of other S-sulfurated molecules. However, depending on the redox conditions of cysteine residues of target proteins, both H2S and S-sulfurated molecules including H2Sn can S-sulfurate the targets. Considering that the endogenous levels of H2S (approximately 3 µM in the brain) and H2Sn (2.6 µM) are well balanced [25] and that the reaction between these signaling molecules and targets is fast [21], both H2S and H2S2 may react with their corresponding targets at a similar frequency. Identifying the redox conditions of target cysteine residues may help understand the functions of H2S or H2Sn as signaling molecules for specific targets.
High concentrations of H2S are toxic, while low concentrations are beneficial. Examples of the former can be observed in Down’s syndrome, ethylmalonyl encephalopathy, and hypoxic brain injury, while examples of the latter can be observed in Parkinson’s disease and Huntington’s disease [87,92,93,96,97,98]. Similarly, H2S exerts opposite effects depending on its concentration as observed in the effect of acetylcholine on the vascular smooth muscle, where low concentrations exert relaxation, while high concentrations exert contraction. Acetylcholine was previously known to contract vasculature, before Furchgott and Zawadzki discovered that lower concentrations of acetylcholine relaxed vasculature where endothelial cells were intact to release EDRF (NO) [99]. Recently, Vellecco et al. identified that vascular contraction by low concentrations of H2S (10 nM to 3 µM) is mediated by cyclic IMP [100]. The effect of nanomolar concentrations of H2S has also been found in T-cell activation [101]. The target, which responds to low concentrations of H2S, should be compared with its response to H2Sn. It will be interesting to know whether or not the targets are oxidized cysteine residues.
Many compounds which release H2S and H2Sn have been developed for clinical use on the basis of their cytoprotective and anti-inflammatory effects. Some of them have successfully completed phase 2 clinical trials and are proceeding further, while many preclinical compounds are awaiting trials [102,103]. It is hoped that various H2S-based compounds will be translated into the field of clinical therapy over the next decade.
Funding
This work was supported by a grant from Strategic Research Program for Brain Sciences from the Japan Agency for Medical Research and Development, AMED, Grant/Award Number JP20dm0107085, and KAKENHI, Grant/Award Number 17K08331 to H.K.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
I thank Csaba Szabo for giving me the opportunity to write a review article. I thank Yuka Kimura for the figures.
Conflicts of Interest
The author declares no conflict of interest.
References
- Reiffenstein, R.J.; Hulbert, W.C.; Roth, S.H. Toxicology of hydrogen sulfide. Annu. Rev. Pharmacol. Toxic 1992, 32, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Warenycia, M.W.; Goodwin, L.R.; Benishin, C.G.; Reiffenstein, R.J.; Grancom, D.M.; Taylor, J.D.; Dieken, F.P. Acute hydrogen sulfide poisoning. Demonstration of selective uptake of sulfide by the brainstem by measurement of brain sulfide levels. Biochem. Pharmacol. 1989, 38, 973–981. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Beck, P.W. Characterization of the enzymic capacity for cysteine desulphhydration in liver and kidney of the rat. Biochem. J. 1982, 206, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, O.W. Mammalian Sulfur Amino Acid Metabolism: An Overview. In Methods in Enzymology; Academic Press: New York, NY, USA, 1987; Volume 143, pp. 366–376. [Google Scholar]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, T.J.; Hawkins, R.D.; Kandel, E.R.; Arancio, O. Tests of the roles of two diffusible substances in long-term potentiation: Evidence for nitric oxide as a possible early retrograde messenger. Proc. Natl. Acad. Sci. USA 1991, 88, 11285–11289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuman, E.M.; Madison, D.V. A requirement for the intercellular messenger nitric oxide in long-term potentiation. Science 1991, 254, 1503–1506. [Google Scholar] [CrossRef]
- Haley, J.E.; Wilcox, G.L.; Chapman, P.F. The role of nitric oxide in hippocampal long-term potentiation. Neuron 1992, 8, 211–216. [Google Scholar] [CrossRef]
- Stevens, C.F.; Wang, Y. Reversal of long-term potentiation by inhibitors of haem oxygenase. Nature 1993, 364, 147–149. [Google Scholar] [CrossRef]
- Zhuo, M.; Small, S.A.; Kandel, E.R.; Hawkins, R.D. Nitric oxide and carbon monoxide produce activity-dependent long-term synaptic enhancement in hippocampus. Science 1993, 260, 1946–1950. [Google Scholar] [CrossRef]
- Aizenman, E.; Lipton, D.A.; Loring, R.H. Selective modulation of NMDA responses by reduction and oxidation. Neuron 1989, 2, 1257–1263. [Google Scholar] [CrossRef]
- Travis, J. The rotten smell of memory: It’s a gas. Sci. News 1996, 149, 116. [Google Scholar] [CrossRef]
- Nagai, Y.; Tsugane, M.; Oka, J.; Kimura, H. Hydrogen sulfide induces calcium waves in astrocytes. FASEB J. 2004, 18, 557–559. [Google Scholar] [CrossRef] [PubMed]
- Streng, T.; Axelsson, H.E.; Hedlund, P.; Andersson, D.A.; Jordt, S.-E.; Bevan, S.; Andersson, K.-E.; Högestätt, E.D.; Zygmunt, P.M. Distribution and Function of the Hydrogen Sulfide-Sensitive TRPA1 Ion Channel in Rat Urinary Bladder. Eur. Urol. 2008, 53, 391–399. [Google Scholar] [CrossRef]
- Ogawa, H.; Takahashi, K.; Miura, S.; Imagawa, T.; Saito, S.; Tominaga, M.; Ohta, T. H2S functions as a nociceptive messenger throughtransient receptor potential ankyrin 1 (TRPA1) activation. Neuroscience 2012, 218, 335–343. [Google Scholar] [CrossRef]
- Searcy, D.G.; Lee, S.H. Sulfur reduction by human erythrocytes. J. Exp. Zool. 1998, 282, 310–322. [Google Scholar] [CrossRef]
- Nagai, Y.; Tsugane, M.; Oka, J.-I.; Kimura, H. Polysulfides induce calcium waves in rat hippocampal astrocytes. J. Pharmacol. Sci. 2006, 100, 200. [Google Scholar]
- Oosumi, K.; Tsugane, M.; Ishigami, M.; Nagai, Y.; Iwai, T.; Oka, J.-I.; Kimura, H. Polysulfide activates TRP channels and increases intracellular Ca2+ in astrocytes. Bull. Jpn. Soc. Neurochem. 2010, 49, 517. [Google Scholar] [CrossRef]
- Kimura, Y.; Mikami, Y.; Osumi, K.; Tsugane, M.; Oka, J.; Kimura, H. Polysulfides are possible H2S-derived signaling molecules in rat brain. FASEB J. 2013, 27, 2451–2457. [Google Scholar] [CrossRef]
- Kimura, Y.; Toyofuku, Y.; Koike, S.; Shibuya, N.; Nagahara, N.; Lefer, D.; Ogasawara, Y.; Kimura, H. Identification of H2S3 and H2S produced by 3-mercaptopyruvate sulfurtransferase in the brain. Sci. Rep. 2015, 5, 14774. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Koike, S.; Shibuya, N.; Lefer, D.; Ogasawara, Y.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces potential redox regularots cysteine- and glutathione-persulfide (Cys-SSH and GSSH) together with signaling molecules H2S2, H2S3 and H2S. Sci. Rep. 2017, 7, 10459. [Google Scholar] [CrossRef]
- Nagahara, N.; Koike, S.; Nirasawa, T.; Kimura, H.; Ogasawara, Y. Alternative pathway of H2S and polysulfides production from sulfurated catalytic-cysteine of reaction intermediates of 3-mercaptopyruvate sulfurtransferase. Biochem. Biophys. Res. Commun. 2018, 496, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, R.; Koike, S.; Takano, Y.; Shibuya, N.; Kimura, Y.; Hanaoka, K.; Urano, Y.; Ogasawara, Y.; Kimura, H. Polysulfides (H2Sn) produced from the interaction of hydrogen sulfide (H2S) and nitric oxide (NO) activate TRPA1 channels. Sci. Rep. 2017, 7, 45995. [Google Scholar] [CrossRef] [Green Version]
- Kharma, A.; Grman, M.; Misak, A.; Domínguez-Álvarez, E.; Nasim, M.J.; Ondrias, K.; Chovanec, M.; Jacob, C. Inorganic Polysulfides and Related Reactive Sulfur–Selenium Species from the Perspective of Chemistry. Molecules 2019, 24, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Kawamura, K.; Kimura, Y.; Shibuya, N.; Kimura, H.; Ogasawara, Y. Analysis of endogenous H2S and H2Sn in mouse brain by high-performance liquid chromatography with fluorescence and tandem mass spectrometric detection. Free Radic. Biol. Med. 2017, 113, 355–362. [Google Scholar] [CrossRef]
- Wang, L.; Cvetkov, T.L.; Chance, M.R.; Moiseenkova-Bell, V.Y. Identification of in Vivo Disulfide Conformation of TRPA1 Ion Channel. J. Biol. Chem. 2012, 287, 6169–6176. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, Y.; Takahashi, K.; Tominaga, M.; Kimura, H.; Ohta, T. Polysulfide Evokes Acute Pain through the Activation of Nociceptive TRPA1 in Mouse Sensory Neurons. Mol. Pain 2015, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S Signals Through Protein S-Sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [Green Version]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, R.; Pálinkás, Z.; Bäsell, K.; Becher, D.; Antelmann, H.; Nagy, P.; Dick, T.P. Polysulfides Link H2S to Protein Thiol Oxidation. Antioxid. Redox Signal. 2013, 19, 1749–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbert, D.; Prysyazhna, O.; Rudyk, O.; Scotcher, J.; Burgoyne, J.; Eaton, P. Protein Kinase G Iα Oxidation Paradoxically Underlies Blood Pressure Lowering by the Reductant Hydrogen Sulfide. Hypertension 2014, 64, 1344–1351. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, A.P.; Wei, W.; Gauld, J.W.; Auld, J.; Özcan, F.; Aslan, M.; Mutus, B. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is inactivated by S-sulfuration in vitro. Free Radic. Biol. Med. 2015, 89, 512–521. [Google Scholar] [CrossRef]
- Hosoki, R.; Matsuki, N.; Kimura, H. The Possible Role of Hydrogen Sulfide as an Endogenous Smooth Muscle Relaxant in Synergy with Nitric Oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Teague, B.; Asiedu, S.; Moore, P.K. The smooth muscle relaxant effect of hydrogen sulphide in vitro: Evidence for a physiological role to control intestinal contractility. Br. J. Pharmacol. 2002, 137, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, M.; Li, L.; Kostetski, I.; Chu, S.H.; Siau, J.L.; Bhatia, M.; Moore, P.K. Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide. Biochem. Biophys. Res. Commun. 2006, 343, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Miljkovic, J.; Allgauer, A.; Chaurio, R.; Shubina, T.; Herrmann, M.; Ivanovic-Burmazovic, I. Biochemical insight into physiological effects of H2S: Reaction with peroxynitrite and formation of a new nitric oxide donor, sulfinyl nitrite. Biochem. J. 2012, 441, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, M.; Dux, M.; Namer, B.; Miljkovic, J.; Cordasic, N.; Will, C.; Kichko, T.I.; De La Roche, J.; Fischer, M.J.; Suárez, S.A.; et al. H2S and NO cooperatively regulate vascular tone by activating a neuroendocrine HNO–TRPA1–CGRP signaling pathway. Nat. Commun. 2014, 5, 4381. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Kuhnle, G.G.C.; Dyson, A.; Fernandez, B.O.; Grman, M.; Dumond, J.F.; Barrow, M.P.; McLeod, G.; Nakagawa, H.; Ondrias, K.; et al. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc. Natl. Acad. Sci. USA 2015, 112, E4651–E4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdándi, V.; Ditrói, T.; Bátai, I.Z.; Sándor, Z.; Minnion, M.; Vasas, A.; Galambos, K.; Buglyó, P.; Pintér, E.; Feelisch, M.; et al. Nitrosopersulfide (SSNO−) Is a Unique Cysteine Polysulfidating Agent with Reduction-Resistant Bioactivity. Antioxid. Redox Signal. 2020, 33, 1277–1294. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.K.; Sikka, G.; Gazi, S.K.; Steppan, J.; Jung, S.M.; Bhunia, A.K.; Barodka, V.M.; Gazi, F.K.; Barrow, R.K.; Wang, R.; et al. Hydrogen Sulfide as Endothelium-Derived Hyperpolarizing Factor Sulfhydrates Potassium Channels. Circ. Res. 2011, 109, 1259–1268. [Google Scholar] [CrossRef]
- Chen, G.F.; Cheung, D.W. Characterization of acetylcholine-induced membrane hyperpolarization in endothelial cells. Circ. Res. 1992, 70, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Eckman, D.M.; Frankovich, J.D.; Keef, K.D. Comparison of the actions of acetylcholine and BRL 38227 in the guinea-pig coronary artery. Br. J. Pharmacol. 1992, 106, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Garland, C.; Plane, F.; Kemp, B.K.; Cocks, T.M. Endothelium-dependent hyperpolarization: A role in the control of vascular tone. Trends Pharmacol. Sci. 1995, 16, 23–30. [Google Scholar] [CrossRef]
- Shibuya, N.; Mikami, Y.; Kimura, Y.; Nagahara, N.; Kimura, H. Vascular Endothelium Expresses 3-Mercaptopyruvate Sulfurtransferase and Produces Hydrogen Sulfide. J. Biochem. 2009, 146, 623–626. [Google Scholar] [CrossRef]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate Sulfurtransferase Produces Hydrogen Sulfide and Bound Sulfane Sulfur in the Brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Earley, S. TRPA1 channels in the vasculature. Br. J. Pharmacol. 2012, 167, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Bucci, M.; Papapetropoulos, A.; Vellecco, V.; Zhou, Z.; Pyriochou, A.; Roussos, C.; Roviezzo, F.; Brancaleone, V.; Cirino, G. Hydrogen Sulfide Is an Endogenous Inhibitor of Phosphodiesterase Activity. Arter. Thromb. Vasc. Biol. 2010, 30, 1998–2004. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H. Hydrogen Sulfide Induces Cyclic AMP and Modulates the NMDA Receptor. Biochem. Biophys. Res. Commun. 2000, 267, 129–133. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Breschi, M.; Lawson, K.; McKay, N.; Miceli, F.; Taglialatela, M.; Calderone, V. Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol. Res. 2013, 70, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Vellecco, V.; Martelli, A.; Bibli, I.S.; Vallifuoco, M.; Manzo, O.L.; Panza, E.; Citi, V.; Calderone, V.; de Dominicis, G.; Cozzolino, C.; et al. Anomalous Kv7 channel activity in human malignant hyperthermia syndrome unmasks a key role for H2S and persulfidation in skeletal muscle. Br. J. Pharmacol. 2020, 177, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Kimura, H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004, 18, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Goto, Y.-I.; Kimura, H. Hydrogen Sulfide Increases Glutathione Production and Suppresses Oxidative Stress in Mitochondria. Antioxid. Redox Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Dargusch, R.; Schubert, D.; Kimura, H. Hydrogen Sulfide Protects HT22 Neuronal Cells from Oxidative Stress. Antioxid. Redox Signal. 2006, 8, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Elrod, J.W.; Calvert, J.W.; Morrison, J.; Doeller, J.E.; Kraus, D.W.; Tao, L.; Jiao, X.; Scalia, R.; Kiss, L.; Szabo, C.; et al. Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial function. Proc. Natl. Acad. Sci. USA 2007, 104, 15560–15565. [Google Scholar] [CrossRef] [Green Version]
- Koike, S.; Ogasawara, Y.; Shibuya, N.; Kimura, H.; Ishii, K. Polysulfide exerts a protective effect against cytotoxicity caused by t-buthylhydroperoxide through Nrf2 signaling in neuroblastoma cells. FEBS Lett. 2013, 587, 3548–3555. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Shibuya, N.; Kimura, H. Sulfite protects neurons from oxidative stress. Br. J. Pharmacol. 2019, 176, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.J.L. Biochemistry of Sulfur-Containing Amino Acids. Annu. Rev. Biochem. 1983, 52, 187–222. [Google Scholar] [CrossRef]
- Shibuya, N.; Koike, S.; Tanaka, M.; Ishigami-Yuasa, M.; Kimura, Y.; Ogasawara, Y.; Fukui, K.; Nagahara, N.; Kimura, H. A novel pathway for the production of hydrogen sulfide from D-cysteine in mammalian cells. Nat. Commun. 2013, 4, 1366. [Google Scholar] [CrossRef] [Green Version]
- Hylin, J.W.; Wood, J.L. Enzymatic Formation of Polysulfides from Mercaptopyruvate. J. Biol. Chem. 1959, 234, 2141–2144. [Google Scholar] [CrossRef]
- Landry, A.P.; Ballou, D.P.; Banerjee, R. H2S oxidation by nanodisc-embedded human sulfide quinone oxidoreductase. J. Biol. Chem. 2017, 292, 11641–11649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitvitsky, V.; Kabil, O.; Banerjee, R. High Turnover Rates for Hydrogen Sulfide Allow for Rapid Regulation of Its Tissue Concentrations. Antioxid. Redox Signal. 2012, 17, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Searcy, D.; Whitehead, J.; Maroney, M. Interaction of Cu, Zn Superoxide Dismutase with Hydrogen Sulfide. Arch. Biochem. Biophys. 1995, 318, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Searcy, D.G. HS−: O2 oxidoreductase activity of Cu, Zn superoxide dismutase. Arch. Biochem. Biophys. 1996, 334, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Gao, Y.; Arif, F.; Arora, K.; Patel, S.; DeLeon, E.R.; Sutton, T.R.; Feelisch, M.; Cortese-Krott, M.; Straub, K.D. Metabolism of hydrogen sulfide (H2S) and Production of Reactive Sulfur Species (RSS) by superoxide dismutase. Redox Biol. 2018, 15, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R.; Gao, Y.; DeLeon, E.R.; Arif, M.; Arif, F.; Arora, N.; Straub, K.D. Catalase as a sulfide-sulfur oxido-reductase: An ancient (and modern?) regulator of reactive sulfur species (RSS). Redox Biol. 2017, 12, 325–339. [Google Scholar] [CrossRef]
- Akaike, T.; Ida, T.; Wei, F.-Y.; Nishida, M.; Kumagai, Y.; Alam, M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Nakamura, M.; Yamazaki, I.; Morrison, M. Reactions of ferryl lactoperoxidase (compound II) with sulfide and sulfhydryl compounds. J. Biol. Chem. 1984, 259, 7080–7085. [Google Scholar] [CrossRef]
- Garai, D.; Ríos-González, B.B.; Furtmüller, P.G.; Fukuto, J.M.; Xian, M.; López-Garriga, J.; Obinger, C.; Nagy, P. Mechanisms of myeloperoxidase catalyzed oxidation of H2S by H2O2 or O2 to produce potent protein Cys–polysulfide-inducing species. Free Radic. Biol. Med. 2017, 113, 551–563. [Google Scholar] [CrossRef]
- Warenycia, M.W.; Goodwin, L.R.; Francom, D.M.; Dieken, F.P.; Kombian, S.B.; Reiffenstein, R.J. Dithiothreitol liberates non-acid labile sulfide from brain tissue of H2S-poisoned animals. Arch. Toxicol. 1990, 64, 650–655. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Ishii, K.; Togawa, T.; Tanabe, S. Determination of Bound Sulfur in Serum by Gas Dialysis/High-Performance Liquid Chromatography. Anal. Biochem. 1993, 215, 73–81. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Isoda, S.; Tanabe, S. Tissue and Subcellular Distribution of Bound and Acid-Labile Sulfur, and the Enzymic Capacity for Sulfide Production in the Rat. Biol. Pharm. Bull. 1994, 17, 1535–1542. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, M.; Hiraki, K.; Umemura, K.; Ogasawara, Y.; Ishii, K.; Kimura, H. A Source of Hydrogen Sulfide and a Mechanism of Its Release in the Brain. Antioxid. Redox Signal. 2009, 11, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, Y.-M.; Jones, D.P. The Redox Proteome. J. Biol. Chem. 2013, 288, 26512–26520. [Google Scholar] [CrossRef] [Green Version]
- Zivanovic, J.; Kouroussis, E.; Kohl, J.B.; Adhikari, B.; Bursac, B.; Schott-Roux, S.; Petrovic, D.; Miljkovic, J.L.; Thomas-Lopez, D.; Jung, Y.; et al. Selective Persulfide Detection Reveals Evolutionarily Conserved Antiaging Effects of S-Sulfhydration. Cell Metab. 2019, 30, 1152–1170.e13. [Google Scholar] [CrossRef]
- Nagahara, N.; Nirasawa, T.; Yoshii, T.; Niimura, Y. Is Novel Signal Transducer Sulfur Oxide Involved in the Redox Cycle of Persulfide at the Catalytic Site Cysteine in a Stable Reaction Intermediate of Mercaptopyruvate Sulfurtransferase? Antioxid. Redox Signal. 2012, 16, 747–753. [Google Scholar] [CrossRef]
- Xiong, J.-W.; Wei, B.; Li, Y.-K.; Zhan, J.-Q.; Jiang, S.-Z.; Chen, H.-B.; Yan, K.; Yu, B.; Yang, Y.-J. Decreased plasma levels of gasotransmitter hydrogen sulfide in patients with schizophrenia: Correlation with psychopathology and cognition. Psychopharmacology 2018, 235, 2267–2274. [Google Scholar] [CrossRef] [PubMed]
- Topcuoglu, C.; Bakirhan, A.; Yilmaz, F.M.; Neselioglu, S.; Erel, O.; Sahiner, S.Y. Thiol/disulfide homeostasis in untreated schizophrenia patients. Psychiatry Res. 2017, 251, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Ide, M.; Ohnishi, T.; Toyoshima, M.; Balan, S.; Maekawa, M.; Shimamoto-Mitsuyama, C.; Iwayama, Y.; Ohba, H.; Watanabe, A.; Ishii, T.; et al. Excess hydrogen sulfide and polysulfides production underlies a schizophrenia pathophysiology. EMBO Mol. Med. 2019, 11, e10695. [Google Scholar] [CrossRef] [PubMed]
- Szabo, C.; Coletta, C.; Chao, C.; Módis, K.; Szczesny, B.; Papapetropoulos, A.; Hellmich, M.R. Tumor-derived hydrogen sulfide, produced by cystathionine-β-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 12474–12479. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, A.L.; et al. Cystathionine beta-synthase (CBS) contributes to advanced ovarian cancer progression and drug resistance. PLoS ONE 2013, 8, e79167. [Google Scholar] [CrossRef]
- Sen, S.; Kawahara, B.; Gupta, D.; Tsai, R.; Khachatryan, M.; Roy-Chowdhuri, S.; Bose, S.; Yoon, A.; Faull, K.; Farias-Eisner, R.; et al. Role of cystathionine β-synthase in human breast cancer. Free Radic. Biol. Med. 2015, 86, 228–238. [Google Scholar] [CrossRef]
- Szczesny, B.; Marcatti, M.; Zatarain, J.R.; Druzhyna, N.; Wiktorowicz, J.E.; Nagy, P.; Hellmich, M.R.; Szabo, C. Inhibition of hydrogen sulfide biosynthesis sensitizes lung adenocarcinoma to chemotherapeutic drugs by inhibiting mitochondrial DNA repair and suppressing cellular bioenergetics. Sci. Rep. 2016, 6, 36125. [Google Scholar] [CrossRef] [Green Version]
- Wróbel, M.; Czubak, J.; Bronowicka-Adamska, P.; Jurkowska, H.; Adamek, D.; Papla, B. Is Development of High-Grade Gliomas Sulfur-Dependent? Molecules 2014, 19, 21350–21362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiota, M.; Naya, M.; Yamamoto, T.; Hishiki, T.; Tani, T.; Takahashi, H.; Kubo, A.; Koike, D.; Itoh, M.; Ohmura, M.; et al. Gold-nanofève surface-enhanced Raman spectroscopy visualizes hypotaurine as a robust anti-oxidant consumed in cancer survival. Nat. Commun. 2018, 9, 1561. [Google Scholar] [CrossRef] [PubMed]
- Takano, N.; Sarfraz, Y.; Gilkes, D.M.; Chaturvedi, P.; Xiang, L.; Suematsu, M.; Zagzag, D.; Semenza, G.L. Decreased expression of cystathionine β-synthase promotes glioma tumorigenesis. Mol. Cancer Res. 2014, 12, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Vandiver, M.S.; Paul, B.D.; Xu, R.; Karuppagounder, S.; Rao, F.; Snowman, A.M.; Ko, H.S.; Lee, Y., II; Dawson, V.L.; Dawson, T.M.; et al. Sulfhydration mediates neuroprotective actions of parkin. Nat. Commun. 2013, 4, 1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taub, J.W.; Huang, X.; Matherly, L.H.; Stout, M.L.; Buck, S.A.; Massey, G.V.; Becton, D.L.; Chang, M.N.; Weinstein, H.J.; Ravindranath, Y. Expression of chromosome 21-localized genes in acute myeloid leukemia: Differences between Down syndrome and non-Down syndrome blast cells and relationship to in vitro sensitivity to cytosine arabinoside and daunorubicin. Blood 1999, 94, 1393–1400. [Google Scholar]
- Ichinohe, A.; Kanaumi, T.; Takashima, S.; Enokido, Y.; Nagai, Y.; Kimura, H. Cystathionine β-synthase is enriched in the brains of Down’s patients. Biochem. Biophys. Res. Commun. 2005, 338, 1547–1550. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, P.; Belardinelli, M.-C.; Chabli, A.; Lallouchi, K.; Chadefaux-Vekemans, B. Endogenous hydrogen sulfide overproduction in Down syndrome. Am. J. Med Genet. Part A 2002, 116A, 310–311. [Google Scholar] [CrossRef]
- Marechal, D.; Brault, V.; Leon, A.; Martin, D.; Pereira, P.L.; Loaëc, N.; Birling, M.-C.; Friocourt, G.; Blondel, M.; Herault, Y. Cbs overdosage is necessary and sufficient to induce cognitive phenotypes in mouse models of Down syndrome and interacts genetically with Dyrk1a. Hum. Mol. Genet. 2019, 28, 1561–1577. [Google Scholar] [CrossRef] [Green Version]
- Panagaki, T.; Randi, E.B.; Augsburger, F.; Szabo, C. Overproduction of H2S, generated by CBS, inhibits mitochondrial Complex IV and suppresses oxidative phosphorylation in Down syndrome. Proc. Natl. Acad. Sci. USA 2019, 116, 18769–18771. [Google Scholar] [CrossRef] [Green Version]
- Tiranti, V.; Viscomi, C.; Hildebrandt, T.; Di Meo, I.; Mineri, R.; Tiveron, C.; Levitt, M.D.; Prelle, A.; Fagiolari, G.; Rimoldi, M.; et al. Loss of ETHE1, a mitochondrial dioxygenase, causes fatal sulfide toxicity in ethylmalonic encephalopathy. Nat. Med. 2009, 15, 200–205. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Meo, I.D.; Zeviani, M.; Viscomi, C.; Braun, H.P. Proteome adaptations in Ethe1-deficient mice indicate a role in lipid catabolism and cytoskeleton organization via post-translational protein modifications. Biosci. Rep. 2013, 33, e00052. [Google Scholar] [CrossRef]
- Cardoso, G.M.F.; Pletsch, J.T.; Parmeggiani, B.; Grings, M.; Glanzel, N.M.; Bobermin, L.D.; Amaral, A.U.; Wajner, M.; Leipnitz, G. Bioenergetics dysfunction, mitochondrial permeability transition pore opening and lipid peroxidation induced by hydrogen sulfide as relevant pathomechanisms underlying the neurological dysfunction characteristic of ethylmalonic encephalopathy. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 2192–2201. [Google Scholar] [CrossRef]
- Peng, Y.-J.; Zhang, X.; Gridina, A.; Chupikova, I.; McCormick, D.L.; Thomas, R.J.; Scammell, T.E.; Kim, G.; Vasavda, C.; Nanduri, J.; et al. Complementary roles of gasotransmitters CO and H2S in sleep apnea. Proc. Natl. Acad. Sci. USA 2017, 114, 1413–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marutani, E.; Morita, M.; Hirai, S.; Kai, S.; Grange, R.M.H.; Miyazaki, Y.; Nagashima, F.; Traeger, L.; Magliocca, A.; Ida, T.; et al. Sulfide catabolism ameliorates hypoxic brain injury. Nat. Commun. 2021, 12, 3108. [Google Scholar] [CrossRef]
- Paul, B.D.; Sbodio, J.I.; Xu, R.; Vandiver, M.S.; Cha, J.Y.; Snowman, A.M.; Snyder, S.H. Cystathionine γ-lyase deficiency mediates neurodegeneration in Huntington’s disease. Nature 2014, 509, 96–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Mitidieri, E.; Vellecco, V.; Brancaleone, V.; Vanacore, D.; Manzo, O.L.; Martin, E.; Sharina, I.; Krutsenko, Y.; Monti, M.C.; Morretta, E.; et al. Involvement of 3′,5′ cyclic inosine monophosphate in cystathionine γ-lyase -dependent regulation of the vascular tone. Br. J. Pharmacol. 2021. [Google Scholar] [CrossRef]
- Miller, T.W.; Wang, E.A.; Gould, S.; Stein, E.V.; Kaur, S.; Lim, L.; Amarnath, S.; Fowler, D.H.; Roberts, D.D. Hydrogen sulfide is an endogenous potentiator of T cell activation. J. Biol. Chem. 2012, 287, 4211–4221. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Gojon, G.; Morales, G.A. SG1002 and Catenated Divalent Organic Sulfur Compounds as Promising Hydrogen Sulfide Prodrugs. Antioxid. Redox Signal. 2020, 33, 1010–1045. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
H2Sn are potential EDHFs. Both 3MST and eNOS are localized to endothelium. H2Sn produced by 3MST or by the chemical interaction between H2S and NO activate TRPA1 channels present in myoendothelial junctions to induce Ca2+ influx, which activates Ca2+-dependent K+ channels. The change in membrane potential is conducted via gap junction to hyperpolarize the smooth muscle plasma membrane.
Figure 1.
H2Sn are potential EDHFs. Both 3MST and eNOS are localized to endothelium. H2Sn produced by 3MST or by the chemical interaction between H2S and NO activate TRPA1 channels present in myoendothelial junctions to induce Ca2+ influx, which activates Ca2+-dependent K+ channels. The change in membrane potential is conducted via gap junction to hyperpolarize the smooth muscle plasma membrane.

Figure 2.
S-Sulfuration of cysteine residues by H2S and H2Sn. Cysteine residues are S-sulfenylated by H2O2 and S-nitrosylated by NO. These oxidized cysteine residues are S-sulfurated by H2S. In contrast, cysteine residues are S-sulfurated by H2Sn.
Figure 2.
S-Sulfuration of cysteine residues by H2S and H2Sn. Cysteine residues are S-sulfenylated by H2O2 and S-nitrosylated by NO. These oxidized cysteine residues are S-sulfurated by H2S. In contrast, cysteine residues are S-sulfurated by H2Sn.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kimura, H. Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years. Biomolecules 2021, 11, 896. https://doi.org/10.3390/biom11060896
AMA Style
Kimura H. Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years. Biomolecules. 2021; 11(6):896. https://doi.org/10.3390/biom11060896
Chicago/Turabian StyleKimura, Hideo. 2021. "Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years" Biomolecules 11, no. 6: 896. https://doi.org/10.3390/biom11060896
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.