
Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”


Abstract
:1. Introduction
2. Adipose-Derived Stem Cells Characteristics
2.1. Nomenclature
2.2. Sources
2.3. Phenotype
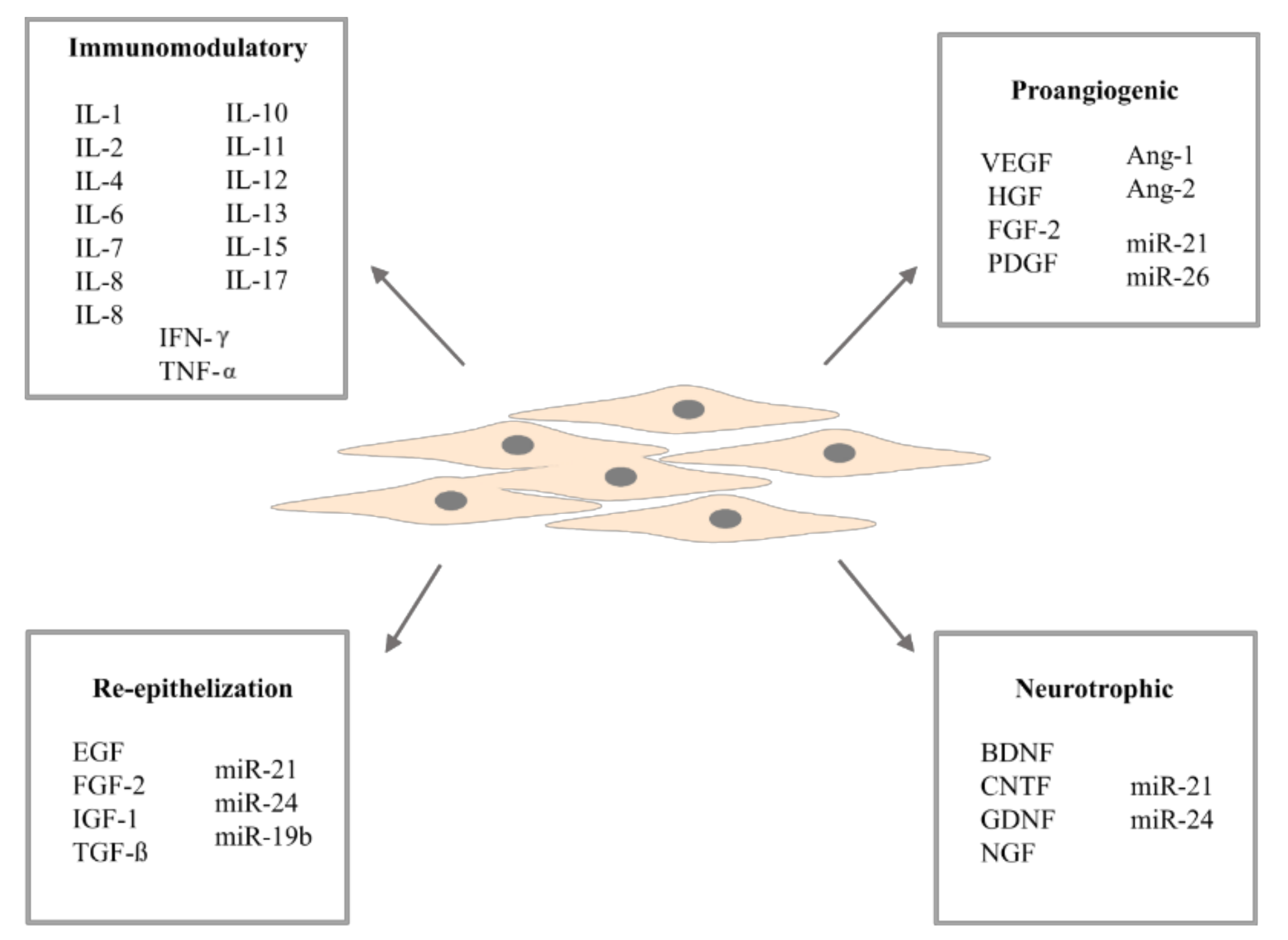
3. Adipose-Derived Stem Cells Secretome
3.1. Cytokines and Growth Factors
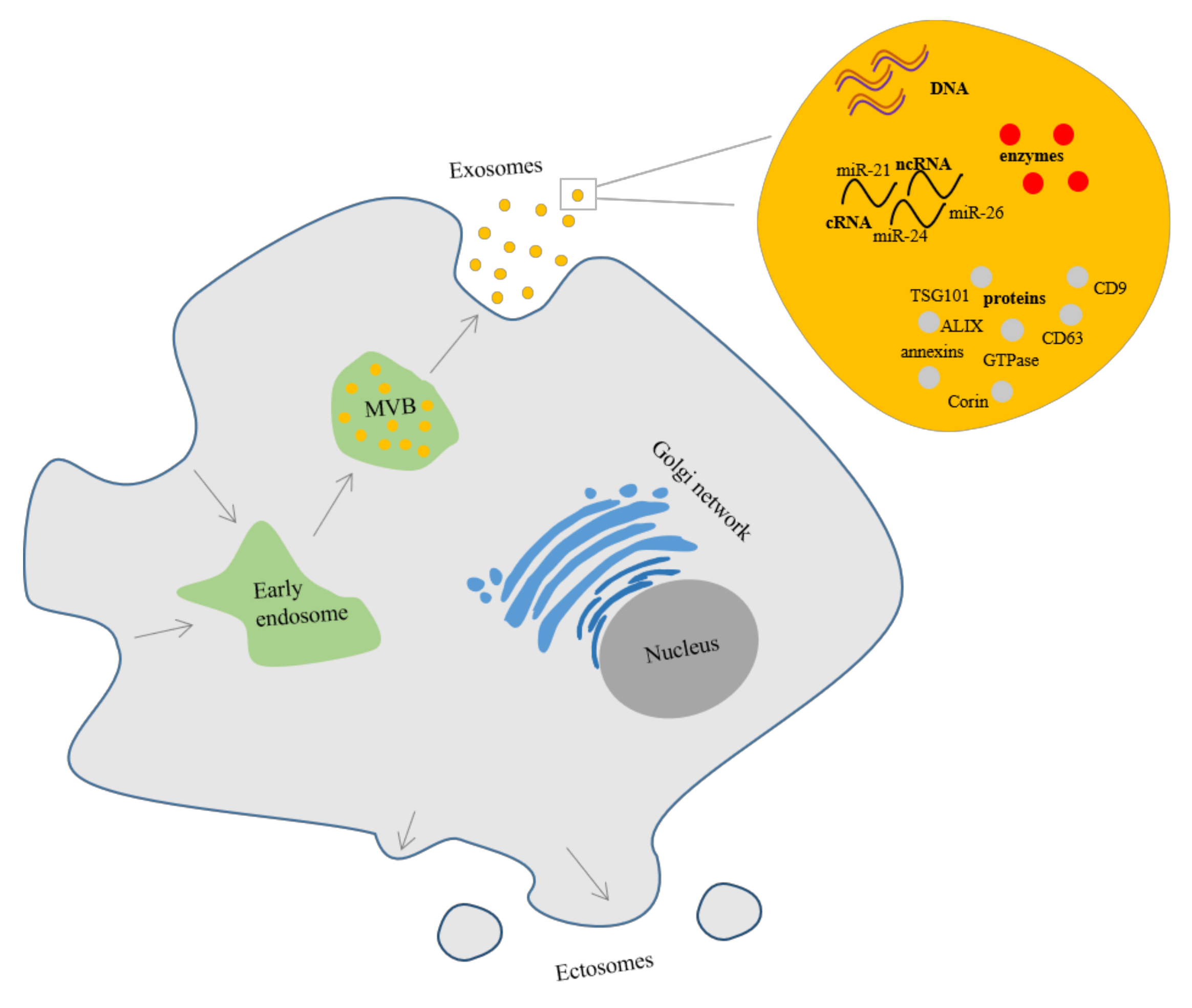
3.2. Extracellular Vesicles
3.2.1. Extracellular Vesicles Composition
3.2.2. Control of the Secretion of Extracellular Vesicles
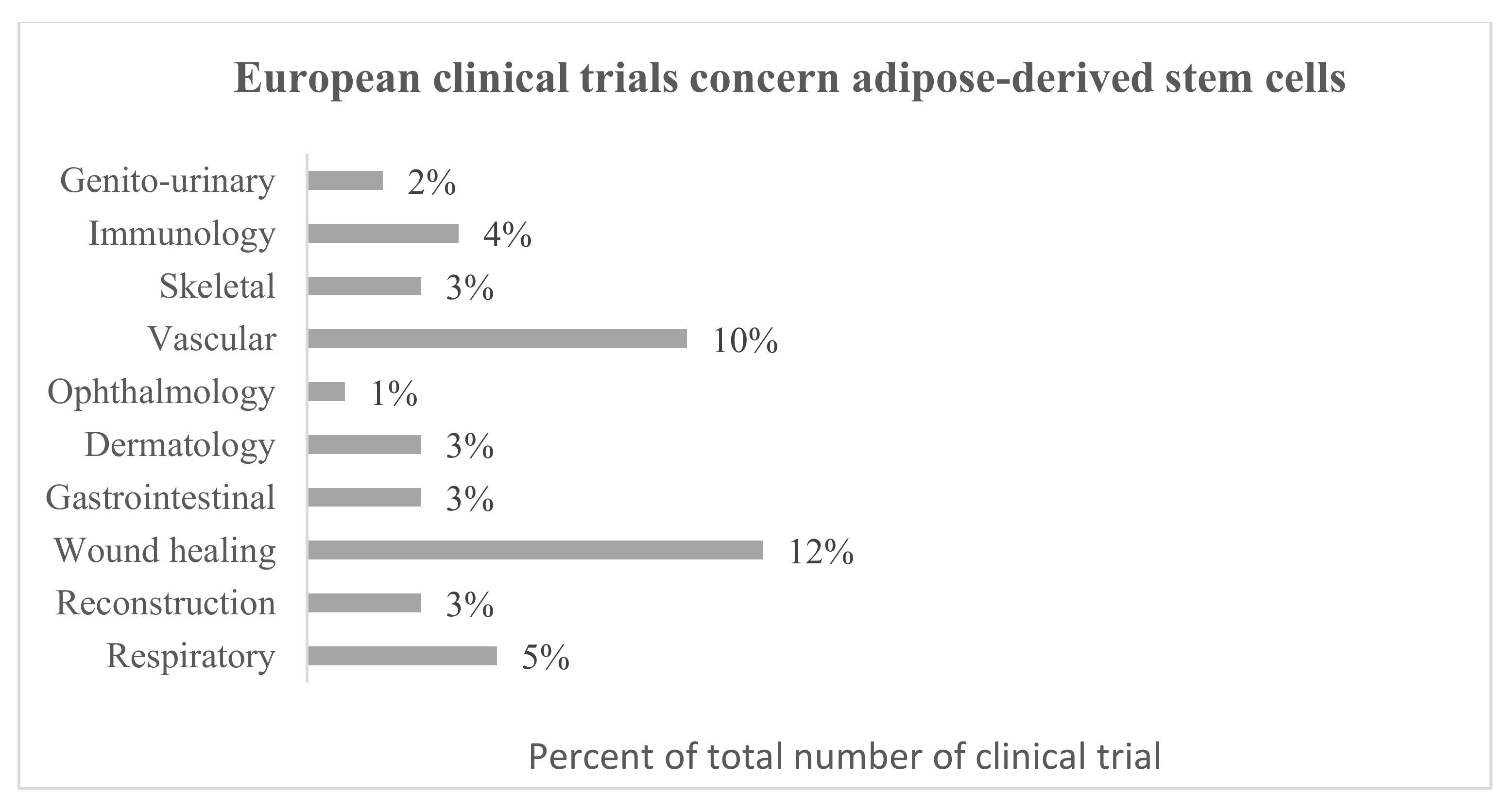
4. Therapeutic Potential of Adipose-Derived Stem Cells Secretome
4.1. Neurodegenerative Diseases
4.2. Cardiovascular Diseases
4.3. Metabolic Diseases
4.4. Respiratory Diseases
4.5. Skeletal Tissue Regeneration
4.6. Wound Healing and Skin Aging
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varderidou-Minasian, S.; Lorenowicz, M.J. Mesenchymal stromal/stem cell-derived extracellular vesicles in tissue repair: Challenges and opportunities. Theranostics 2020, 10, 5979–5997. [Google Scholar] [CrossRef] [PubMed]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; Van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar]
- Prindull, G.; Prindull, B.; Meulen, N. Haematopoietic stem cells (CFUc) in human cord blood. Acta Paediatr. 1978, 67, 413–416. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delo, D.M.; De Coppi, P.; Bartsch, G., Jr.; Atala, A. Amniotic fluid and placental stem cells. Methods Enzymol. 2006, 419, 426–438. [Google Scholar] [CrossRef]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-derived and bone marrow mesenchymal stem cells: A donor-matched comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Dai, R.; Wang, Z.; Samanipour, R.; Koo, K.-I.; Kim, K. Adipose-Derived Stem Cells for Tissue Engineering and Regenerative Medicine Applications. Stem Cells Int. 2016, 2016, 6737345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; He, D.; Kleiner, G.; Kuluz, J.T. Neuron-like Differentiation of Adipose-Derived Stem Cells from Infant Piglets in Vitro. J. Spinal Cord Med. 2007, 30, S35–S40. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.-H.; Tong, Y.-W.; Yeh, W.-L.; Lei, K.F.; Chen, A.C.-Y. Self-Renewal and Differentiation of Adipose-Derived Stem Cells (ADSCs) Stimulated by Multi-Axial Tensile Strain in a Pneumatic Microdevice. Micromachines 2018, 9, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parate, D.; Kadir, N.D.; Celik, C.; Lee, E.H.; Hui, J.H.P.; Franco-Obregón, A.; Yang, Z. Pulsed electromagnetic fields potentiate the paracrine function of mesenchymal stem cells for cartilage regeneration. Stem Cell Res. Ther. 2020, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, O.; Izquierdo, G.; Fernández, V.; Leyva, L.; Reyes, V.; Guerrero, M.; León, A.; Arnaiz, C.; Navarro, G.; Páramo, M.D.; et al. Adipose-derived mesenchymal stem cells (AdMSC) for the treatment of secondary-progressive multiple sclerosis: A triple blinded, placebo controlled, randomized phase I/II safety and feasibility study. PLoS ONE 2018, 13, e0195891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.W.; Shin, I.S.; Song, J.W.; Lee, M.; Yun, T.W.; Yang, J.; Choi, K.-S.; Kim, S.-J. Effects of Transplantation of CTLA4Ig-Overexpressing Adipose Tissue-Derived Mesenchymal Stem Cells in Mice with Sustained Severe Rheumatoid Arthritis. Cell Transplant. 2016, 25, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Ueyama, H.; Okano, T.; Orita, K.; Mamoto, K.; Sobajima, S.; Iwaguro, H.; Nakamura, H. Local transplantation of adipose-derived stem cells has a significant therapeutic effect in a mouse model of rheumatoid arthritis. Sci. Rep. 2020, 10, 3076. [Google Scholar] [CrossRef] [Green Version]
- Sonomoto, K.; Yamaoka, K.; Kaneko, H.; Yamagata, K.; Sakata, K.; Zhang, X.; Kondo, M.; Zenke, Y.; Sabanai, K.; Nakayamada, S.; et al. Spontaneous Differentiation of Human Mesenchymal Stem Cells on Poly-Lactic-Co-Glycolic Acid Nano-Fiber Scaffold. PLoS ONE 2016, 11, e0153231. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, P.D.-A.; Garcia-Arranz, M.; Georgiev-Hristov, T.; Olmo, D.G. A new bronchoscopic treatment of tracheomediastinal fistula using autologous adipose-derived stem cells. Thorax 2008, 63, 374–376. [Google Scholar] [CrossRef] [Green Version]
- Olmo, D.G.; Herreros, D.; De-La-Quintana, P.; Guadalajara, H.; Trébol, J.; Georgiev-Hristov, T.; Garcia-Arranz, M. Adipose-Derived Stem Cells in Crohn’s Rectovaginal Fistula. Case Rep. Med. 2010, 2010, 961758. [Google Scholar] [CrossRef]
- Ikemoto, T.; Tokuda, K.; Wada, Y.; Gao, L.; Miyazaki, K.; Yamada, S.; Saito, Y.; Imura, S.; Morine, Y.; Shimada, M. Adipose Tissue from Type 1 Diabetes Mellitus Patients Can Be Used to Generate Insulin-Producing Cells. Pancreas 2020, 49, 1225–1231. [Google Scholar] [CrossRef]
- Wang, M.; Song, L.; Strange, C.; Dong, X.; Wang, H. Therapeutic Effects of Adipose Stem Cells from Diabetic Mice for the Treatment of Type 2 Diabetes. Mol. Ther. 2018, 26, 1921–1930. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.-Y.; Liu, J.; Wang, Y.-L.; Liu, Y.; Shao, Y.; Han, Y.; Qin, Y.-R.; Xiao, F.-J.; Li, P.-F.; Zhao, L.-J.; et al. Adipose-Derived Mesenchymal Stem Cells Ameliorate Lipid Metabolic Disturbance in Mice. Stem Cells Transl. Med. 2016, 5, 1162–1170. [Google Scholar] [CrossRef]
- Perin, E.C.; Sanz-Ruiz, R.; Sanchez, P.L.; Lasso, J.; Pérez-Cano, R.; Alonso-Farto, J.C.; Pérez-David, E.; Santos, M.E.F.; Serruys, P.W.; Duckers, H.J.; et al. Adipose-derived regenerative cells in patients with ischemic cardiomyopathy: The PRECISE Trial. Am. Heart J. 2014, 168, 88–95.e2. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Scutaru, T.T.; Ghetu, N.; Carasevici, E.; Lupascu, C.D.; Ferariu, D.; Pieptu, D.; Coman, C.-G.; Danciu, M. The Effects of Adipose-Derived Stem Cell-Differentiated Adipocytes on Skin Burn Wound Healing in Rats. J. Burn Care Res. 2017, 38, 1–10. [Google Scholar] [CrossRef]
- Gaur, M.; Dobke, M.; Lunyak, V.V. Mesenchymal Stem Cells from Adipose Tissue in Clinical Applications for Dermatological Indications and Skin Aging. Int. J. Mol. Sci. 2017, 18, 208. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.S.; Kim, S.; Yang, C.E.; Choi, Y.; Song, S.Y.; Kim, H.O. Human Adipose Mesenchymal Stem Cell-Derived Exosomes: A Key Player in Wound Healing. Tissue Eng. Regen. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Banaś, A.; Teratani, T.; Yamamoto, Y.; Tokuhara, M.; Takeshita, F.; Osaki, M.; Kawamata, M.; Kato, T.; Okochi, H.; Ochiya, T. IFATS Collection: In Vivo Therapeutic Potential of Human Adipose Tissue Mesenchymal Stem Cells after Transplantation into Mice with Liver Injury. Stem Cells 2008, 26, 2705–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzyna, A.; Pikuła, B.; Ludwin, A.; Kocan, B.; Banaś-Ząbczyk, A. The influence of an electromagnetic field on adipose-derived stem/stromal cells’ growth factor secretion: Modulation of FGF-2 production by in vitro exposure. Arch. Biol. Sci. 2020, 72, 339–347. [Google Scholar] [CrossRef]
- Kilroy, G.E.; Foster, S.J.; Wu, X.; Ruiz, J.; Sherwood, S.; Heifetz, A.; Ludlow, J.W.; Stricker, D.M.; Potiny, S.; Green, P.; et al. Cytokine profile of human adipose-derived stem cells: Expression of angiogenic, hematopoietic, and pro-inflammatory factors. J. Cell. Physiol. 2007, 212, 702–709. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Zhao, J.; Nie, F.; Qin, Z.; Xue, H.; Wang, G.; Li, D. Exosomes from Adipose-Derived Stem Cells (ADSCs) Overexpressing miR-21 Promote Vascularization of Endothelial Cells. Sci. Rep. 2019, 9, 12861. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.L.; Chen, H.H.; Chen, K.H.; Chiang, J.Y.; Li, Y.C.; Lin, H.S.; Sung, P.H.; Yip, H.K. Adipose-derived mesenchymal stem cell-derived exosomes markedly protected the brain against sepsis syndrome induced injury in rat. Am. J. Transl. Res. 2019, 11, 3955–3971. [Google Scholar] [PubMed]
- Chen, Z.; Xue, S.; Zhang, S.; Cheng, K.; Ye, Q. Exosomes from donor-derived adipose mesenchymal stem cells prolong the survival of vascularized composite allografts. J. Cell. Physiol. 2021, 236, 5895–5905. [Google Scholar] [CrossRef]
- Dubey, N.K.; Mishra, V.K.; Dubey, R.; Deng, Y.-H.; Tsai, F.-C.; Deng, W.-P. Revisiting the Advances in Isolation, Characterization and Secretome of Adipose-Derived Stromal/Stem Cells. Int. J. Mol. Sci. 2018, 19, 2200. [Google Scholar] [CrossRef] [Green Version]
- Franchi, S.; Castelli, M.; Amodeo, G.; Niada, S.; Ferrari, D.; Vescovi, A.L.; Brini, A.T.; Panerai, A.E.; Sacerdote, P. Adult Stem Cell as New Advanced Therapy for Experimental Neuropathic Pain Treatment. BioMed Res. Int. 2014, 2014, 470983. [Google Scholar] [CrossRef]
- Sacerdote, P.; Niada, S.; Franchi, S.; Arrigoni, E.; Rossi, A.; Yenagi, V.; de Girolamo, L.; Panerai, A.E.; Brini, A.T. Systemic Administration of Human Adipose-Derived Stem Cells Reverts Nociceptive Hypersensitivity in an Experimental Model of Neuropathy. Stem Cells Dev. 2013, 22, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvaro-Gracia, J.M.; Jover, J.A.; García-Vicuña, R.; Carreño, L.; Alonso, A.; Marsal, S.; Blanco, F.; Martínez-Taboada, V.M.; Taylor, P.; Martín-Martín, C.; et al. Intravenous administration of expanded allogeneic adipose-derived mesenchymal stem cells in refractory rheumatoid arthritis (Cx611): Results of a multicentre, dose escalation, randomised, single-blind, placebo-controlled phase Ib/IIa clinical trial. Ann. Rheum. Dis. 2017, 76, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Shin, T.-H.; Kim, H.-S. Current Strategies to Enhance Adipose Stem Cell Function: An Update. Int. J. Mol. Sci. 2019, 20, 3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankrum, J.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zeng, Q.; Han, Q.; Xia, W. Effect of pH, temperature and freezing-thawing on quantity changes and cellular uptake of exosomes. Protein Cell 2019, 10, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Xu, Y.; Yang, E.; Du, Y. Stemness and Regenerative Potential of Corneal Stromal Stem Cells and Their Secretome after Long-Term Storage: Implications for Ocular Regeneration. Investig. Opthalmol. Vis. Sci. 2018, 59, 3728–3738. [Google Scholar] [CrossRef]
- Baer, P.C. Adipose-derived mesenchymal stromal/stem cells: An update on their phenotype in vivo and in vitro. World J. Stem Cells 2014, 6, 256–265. [Google Scholar] [CrossRef]
- Barry, F.P.; Murphy, J. Mesenchymal stem cells: Clinical applications and biological characterization. Int. J. Biochem. Cell Biol. 2004, 36, 568–584. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocan, B.; Maziarz, A.; Tabarkiewicz, J.; Ochiya, T.; Banaś-Ząbczyk, A. Trophic Activity and Phenotype of Adipose Tissue-Derived Mesenchymal Stem Cells as a Background of Their Regenerative Potential. Stem Cells Int. 2017, 2017, 1653254. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wu, H. T Cells in Adipose Tissue: Critical Players in Immunometabolism. Front. Immunol. 2018, 9, 2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, F.J.; Holt, D.J.; Vargas, V.; Yockman, J.; Boudina, S.; Atkinson, D.; Grainger, D.W.; Revelo, M.P.; Sherman, W.; Bull, D.A.; et al. Metabolically Active Human Brown Adipose Tissue Derived Stem Cells. Stem Cells 2014, 32, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Wang, X.; Sun, C.; Kang, Y.; Xu, J.; Wang, X.; Hui, Y. Adipose-derived stem cells: Sources, potency, and implications for regenerative therapies. Biomed. Pharmacother. 2019, 114, 108765. [Google Scholar] [CrossRef] [PubMed]
- Gesta, S.; Tseng, Y.-H.; Kahn, C.R. Developmental Origin of Fat: Tracking Obesity to Its Source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Tsekouras, A.; Mantas, D.; Tsilimigras, I.D.; Moris, D.; Kontos, M.; Zografos, C.G. Comparison of the Viability and Yield of Adipose-Derived Stem Cells (ASCs) from Different Donor Areas. In Vivo 2017, 31, 1229–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feijóo-Bandín, S.; Rodríguez-Penas, D.; García-Rúa, V.; Mosquera-Leal, A.; González-Juanatey, J.R.; Lago, F. Adipokines at the Cardiovascular System: Role in Health and Disease. SM J. Endocrinol. Metab. 2016, 2, 1009. [Google Scholar]
- Prunet-Marcassus, B.; Cousin, B.; Caton, D.; André, M.; Penicaud, L.; Casteilla, L. From heterogeneity to plasticity in adipose tissues: Site-specific differences. Exp. Cell Res. 2006, 312, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Nepali, S.; Park, M.; Lew, H.; Kim, O. Comparative Analysis of Human Adipose-Derived Mesenchymal Stem Cells from Orbital and Abdominal Fat. Stem Cells Int. 2018, 2018, 3932615. [Google Scholar] [CrossRef] [PubMed]
- Schipper, B.M.; Marra, K.G.; Zhang, W.; Donnenberg, A.D.; Rubin, J.P. Regional Anatomic and Age Effects on Cell Function of Human Adipose-Derived Stem Cells. Ann. Plast. Surg. 2008, 60, 538–544. [Google Scholar] [CrossRef]
- Harrison, M.A.A.; Wise, R.M.; Benjamin, B.P.; Hochreiner, E.M.; Mohiuddin, O.A.; Bunnell, B.A. Adipose-Derived Stem Cells from Obese Donors Polarize Macrophages and Microglia toward a Pro-Inflammatory Phenotype. Cells 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Oñate, B.; Vilahur, G.; Camino-López, S.; Díez-Caballero, A.; Ballesta-López, C.; Ybarra, J.; Moscatiello, F.; Herrero, J.; Badimon, L. Stem cells isolated from adipose tissue of obese patients show changes in their transcriptomic profile that indicate loss in stemcellness and increased commitment to an adipocyte-like phenotype. BMC Genom. 2013, 14, 625. [Google Scholar] [CrossRef] [Green Version]
- Badimon, L.; Cubedo, J. Adipose tissue depots and inflammation: Effects on plasticity and resident mesenchymal stem cell function. Cardiovasc. Res. 2017, 113, 1064–1073. [Google Scholar] [CrossRef] [Green Version]
- Badimon, L.; Oñate, B.; Vilahur, G. Adipose-derived Mesenchymal Stem Cells and Their Reparative Potential in Ischemic Heart Disease. Revista Española Cardiología 2015, 68, 599–611. [Google Scholar] [CrossRef]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic review of patient factors affecting adipose stem cell viability and function: Implications for regenerative therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Shang, Q.; Pan, Z.; Bai, Y.; Lining, Z.; Zhang, H.; Zhang, Q.; Guo, C.; Zhang, L.; Wang, Q. Exosomes from Adipose-Derived Stem Cells Attenuate Adipose Inflammation and Obesity through Polarizing M2 Macrophages and Beiging in White Adipose Tissue. Diabetes 2018, 67, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Di Maggio, N.; Martella, E.; Frismantiene, A.; Resink, T.J.; Schreiner, S.; Lucarelli, E.; Jaquiery, C.; Schaefer, D.J.; Martin, I.; Scherberich, A. Extracellular matrix and α5β1 integrin signaling control the maintenance of bone formation capacity by human adipose-derived stromal cells. Sci. Rep. 2017, 7, 44398. [Google Scholar] [CrossRef] [Green Version]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef]
- Gentile, P.; Piccinno, M.S.; Calabrese, C. Characteristics and Potentiality of Human Adipose-Derived Stem Cells (hASCs) Obtained from Enzymatic Digestion of Fat Graft. Cells 2019, 8, 282. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, E.; Kloster, A.; Di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of Human Adipose-Derived Cells: Temporal Changes in Stromal-Associated and Stem Cell-Associated Markers. Stem Cells 2006, 24, 376–385. [Google Scholar] [CrossRef]
- Lin, C.-S.; Ning, H.; Lin, G.; Lue, T.F. Is CD34 truly a negative marker for mesenchymal stromal cells? Cytotherapy 2012, 14, 1159–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Yang, Z.X.; Han, Z.-B.; Ji, Y.R.; Wang, Y.W.; Liang, L.; Chi, Y.; Yang, S.G.; Li, L.N.; Luo, W.F.; Li, J.P.; et al. CD106 Identifies a Subpopulation of Mesenchymal Stem Cells with Unique Immunomodulatory Properties. PLoS ONE 2013, 8, e59354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, H.; Matsumoto, D.; Eto, H.; Inoue, K.; Aoi, N.; Kato, H.; Araki, J.; Yoshimura, K. Functional Implications of CD34 Expression in Human Adipose-Derived Stem/Progenitor Cells. Stem Cells Dev. 2009, 18, 1201–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieczkowska, A.; Schumacher, A.; Filipowicz, N.; Wardowska, A.; Zieliński, M.; Madanecki, P.; Nowicka, E.; Langa, P.; Deptuła, M.; Zieliński, J.; et al. Immunophenotyping and transcriptional profiling of in vitro cultured human adipose tissue derived stem cells. Sci. Rep. 2018, 8, 11339. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, T.; Song, K.; Fan, X.; Ma, X.; Cui, Z. Adipose-derived stem cell: A better stem cell than BMSC. Cell Biochem. Funct. 2008, 26, 664–675. [Google Scholar] [CrossRef]
- El Atat, O.; Antonios, D.; Hilal, G.; Hokayem, N.; Abou-Ghoch, J.; Hashim, H.; Serhal, R.; Hebbo, C.; Moussa, M.; Alaaeddine, N. An Evaluation of the Stemness, Paracrine, and Tumorigenic Characteristics of Highly Expanded, Minimally Passaged Adipose-Derived Stem Cells. PLoS ONE 2016, 11, e0162332. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Ryan, C.M.; Pathan, O.; Abraham, D.; Denton, C.P.; Butler, P.E.M. Characteristics of human adipose derived stem cells in scleroderma in comparison to sex and age matched normal controls: Implications for regenerative medicine. Stem Cell Res. Ther. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Li, Y.-N.; Song, L.; Liu, R.; Li, X.; Shang, Q.; Wang, Y.; Shao, C.; Shi, Y. Macrophages inhibit adipogenic differentiation of adipose tissue derived mesenchymal stem/stromal cells by producing pro-inflammatory cytokines. Cell Biosci. 2020, 10, 88. [Google Scholar] [CrossRef]
- Noverina, R.; Widowati, W.; Ayuningtyas, W.; Kurniawan, D.; Afifah, E.; Laksmitawati, D.R.; Rinendyaputri, R.; Rilianawati, R.; Faried, A.; Bachtiar, I.; et al. Growth factors profile in conditioned medium human adipose tissue-derived mesenchymal stem cells (CM-hATMSCs). Clin. Nutr. Exp. 2019, 24, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Tratwal, J.; Mathiasen, A.B.; Juhl, M.; Brorsen, S.K.; Kastrup, J.; Ekblond, A. Influence of vascular endothelial growth factor stimulation and serum deprivation on gene activation patterns of human adipose tissue-derived stromal cells. Stem Cell Res. Ther. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaber, S.P.; Webster, R.A.; Hill, C.J.; Breen, E.J.; Kuah, D.; Vesey, G.; Herbert, B.R. Analysis of in vitro secretion profiles from adipose-derived cell populations. J. Transl. Med. 2012, 10, 172. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Fu, B.; Yang, X.; Xiao, Y.; Pan, M. Adipose mesenchymal stem cell-derived exosomes promote cell proliferation, migration, and inhibit cell apoptosis via Wnt/β-catenin signaling in cutaneous wound healing. J. Cell. Biochem. 2019, 120, 10847–10854. [Google Scholar] [CrossRef]
- Cao, G.; Chen, B.; Zhang, X.; Chen, H. Human Adipose-Derived Mesenchymal Stem Cells-Derived Exosomal microRNA-19b Promotes the Healing of Skin Wounds through Modulation of the CCL1/TGF-β Signaling Axis. Clin. Cosmet. Investig. Dermatol. 2020, 13, 957–971. [Google Scholar] [CrossRef]
- Mizuta, Y.; Akahoshi, T.; Guo, J.; Zhang, S.; Narahara, S.; Kawano, T.; Murata, M.; Tokuda, K.; Eto, M.; Hashizume, M.; et al. Exosomes from adipose tissue-derived mesenchymal stem cells ameliorate histone-induced acute lung injury by activating the PI3K/Akt pathway in endothelial cells. Stem Cell Res. Ther. 2020, 11, 508. [Google Scholar] [CrossRef]
- Adolfsson, E.; Helenius, G.; Friberg, Ö.; Samano, N.; Frøbert, O.; Johansson, K. Bone marrow- and adipose tissue-derived mesenchymal stem cells from donors with coronary artery disease; growth, yield, gene expression and the effect of oxygen concentration. Scand. J. Clin. Lab. Investig. 2020, 80, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.-C.; Chou, H.-L.; Hung, W.-T.; Liu, H.-W.; Chu, T.-Y. Human adipose-derived stem cells cultured in keratinocyte serum free medium: Donor’s age does not affect the proliferation and differentiation capacities. J. Biomed. Sci. 2013, 20, 59. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Li, J.-Z.; Xie, B.-D.; Tian, H.; Fang, S.-H.; Jiang, S.-L.; Kang, K. Lower Senescence of Adipose-Derived Stem Cells than Donor-Matched Bone Marrow Stem Cells for Surgical Ventricular Restoration. Stem Cells Dev. 2018, 27, 612–623. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Pontecorvi, P.; Anastasiadou, E.; Napoli, C.; Marchese, C. Immunomodulatory Effect of Adipose-Derived Stem Cells: The Cutting Edge of Clinical Application. Front. Cell Dev. Biol. 2020, 8, 236. [Google Scholar] [CrossRef]
- Niu, P.; Smagul, A.; Wang, L.; Sadvakas, A.; Sha, Y.; Pérez, L.M.; Nussupbekova, A.; Amirbekov, A.; Akanov, A.A.; Gálvez, B.G.; et al. Transcriptional profiling of interleukin-2-primed human adipose derived mesenchymal stem cells revealed dramatic changes in stem cells response imposed by replicative senescence. Oncotarget 2015, 6, 17938–17957. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.-E.; Lee, S.Y. IL-6 is produced by adipose-derived stromal cells and promotes osteogenesis. Biochim. Biophys. Acta 2013, 1833, 2608–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastidas-Coral, A.P.; Bakker, A.D.; Zandieh-Doulabi, B.; Kleverlaan, C.J.; Bravenboer, N.; Forouzanfar, T.; Klein-Nulend, J. Cytokines TNF-α, IL-6, IL-17F, and IL-4 Differentially Affect Osteogenic Differentiation of Human Adipose Stem Cells. Stem Cells Int. 2016, 2016, 1318256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Hoogduijn, M.J.; Baan, C.C.; Korevaar, S.S.; De Kuiper, R.; Yan, L.; Wang, L.; Van Besouw, N.M. Adipose Tissue-Derived Mesenchymal Stem Cells Have a Heterogenic Cytokine Secretion Profile. Stem Cells Int. 2017, 2017, 4960831. [Google Scholar] [CrossRef]
- Prichard, H.L.; Reichert, W.; Klitzman, B. IFATS collection: Adipose-derived stromal cells improve the foreign body response. Stem Cells 2008, 26, 2691–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghannam, S.; Bouffi, C.; Djouad, F.; Jorgensen, C.; Noël, D. Immunosuppression by mesenchymal stem cells: Mechanisms and clinical applications. Stem Cell Res. Ther. 2010, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Yin, S.; Liu, W.; Li, N.; Zhang, W.; Cao, Y. Expanded Adipose-Derived Stem Cells Suppress Mixed Lymphocyte Reaction by Secretion of Prostaglandin E2. Tissue Eng. 2007, 13, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.W.; Kang, K.-S.; Koo, H.C.; Park, J.R.; Choi, E.W.; Park, Y.H. Soluble Factors-Mediated Immunomodulatory Effects of Canine Adipose Tissue-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2008, 17, 681–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, C.-H.; Shiau, M.-Y.; Chuang, P.-H.; Chang, Y.-H.; Hwang, J. Interleukin-4 regulates lipid metabolism by inhibiting adipogenesis and promoting lipolysis. J. Lipid Res. 2014, 55, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga, L.A.; Shen, W.-J.; Joyce-Shaikh, B.; Pyatnova, E.A.; Richards, A.G.; Thom, C.; Andrade, S.M.; Cua, D.J.; Kraemer, F.B.; Butcher, E.C. IL-17 Regulates Adipogenesis, Glucose Homeostasis, and Obesity. J. Immunol. 2010, 185, 6947–6959. [Google Scholar] [CrossRef] [Green Version]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of Angiogenic and Antiapoptotic Factors by Human Adipose Stromal Cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef]
- Ieda, Y.; Fujita, J.; Ieda, M.; Yagi, T.; Kawada, H.; Ando, K.; Fukuda, K. G-CSF and HGF: Combination of vasculogenesis and angiogenesis synergistically improves recovery in murine hind limb ischemia. J. Mol. Cell. Cardiol. 2007, 42, 540–548. [Google Scholar] [CrossRef]
- Ball, S.G.; Shuttleworth, C.A.; Kielty, C.M. Mesenchymal stem cells and neovascularization: Role of platelet-derived growth factor receptors. J. Cell. Mol. Med. 2007, 11, 1012–1030. [Google Scholar] [CrossRef] [Green Version]
- Lopatina, T.; Bruno, S.; Tetta, C.; Kalinina, N.; Porta, M.; Camussi, G. Platelet-derived growth factor regulates the secretion of extracellular vesicles by adipose mesenchymal stem cells and enhances their angiogenic potential. Cell Commun. Signal. 2014, 12, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, A.; Aboalola, D.; Han, V.K.M. The Roles of Insulin-Like Growth Factors in Mesenchymal Stem Cell Niche. Stem Cells Int. 2017, 2017, 9453108. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, X.; Dang, H.; Liu, P.; Zhang, B.O.; Xu, F. Insulin-like growth factor 2 regulates the proliferation and differentiation of rat adipose-derived stromal cells via IGF-1R and IR. Cytotherapy 2019, 21, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Chicharro, D.; Carrillo, J.M.; Rubio, M.; Cugat, R.; Cuervo, B.; Guil, S.; Forteza, J.; Moreno, V.; Vilar, J.M.; Sopena, J. Combined plasma rich in growth factors and adipose-derived mesenchymal stem cells promotes the cutaneous wound healing in rabbits. BMC Vet. Res. 2018, 14, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, J.L.; Stapleton, J.T.; Okeoma, C.M. Vehicles of intercellular communication: Exosomes and HIV-1. J. Gen. Virol. 2019, 100, 350–366. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [Green Version]
- Fatima, F.; Ekstrom, K.; Nazarenko, I.; Maugeri, M.; Valadi, H.; Hill, A.F.; Camussi, G.; Nawaz, M. Non-coding RNAs in Mesenchymal Stem Cell-Derived Extracellular Vesicles: Deciphering Regulatory Roles in Stem Cell Potency, Inflammatory Resolve, and Tissue Regeneration. Front. Genet. 2017, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Bai, X.; Zhao, B.; Li, Y.; Zhang, Y.; Li, Z.; Wang, X.; Luo, L.; Han, F.; Zhang, J.; et al. Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway. Exp. Cell Res. 2018, 370, 333–342. [Google Scholar] [CrossRef]
- Cha, H.; Hong, S.; Park, J.H.; Park, H.H. Stem Cell-Derived Exosomes and Nanovesicles: Promotion of Cell Proliferation, Migration, and Anti-Senescence for Treatment of Wound Damage and Skin Ageing. Pharmaceutics 2020, 12, 1135. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Zhang, Q.; Hu, W.; Zhao, C.; Lv, W.; Yi, Y.; Wu, Y.; Wu, M. Exosomes from Adipose-Derived Stem Cells: The Emerging Roles and Applications in Tissue Regeneration of Plastic and Cosmetic Surgery. Front. Cell Dev. Biol. 2020, 8, 574223. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Xing, X.; Han, S.; Cheng, G.; Ni, Y.; Li, Z.; Li, Z. Proteomic Analysis of Exosomes from Adipose-Derived Mesenchymal Stem Cells: A Novel Therapeutic Strategy for Tissue Injury. BioMed Res. Int. 2020, 2020, 6094562. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Li, H.; Zhou, Y.; Gu, B.; Xu, Y.; Fu, Q.; Peng, X.; Cao, N.; Fu, Q.; Jin, M.; et al. Therapeutic Potential of Human Adipose-Derived Stem Cell Exosomes in Stress Urinary Incontinence—An In Vitro and In Vivo Study. Cell. Physiol. Biochem. 2018, 48, 1710–1722. [Google Scholar] [CrossRef] [PubMed]
- González-Cubero, E.; González-Fernández, M.L.; Gutiérrez-Velasco, L.; Navarro-Ramírez, E.; Villar-Suárez, V. Isolation and characterization of exosomes from adipose tissue-derived mesenchymal stem cells. J. Anat. 2021, 238, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.S.; Carter, G.; El Bassit, G.; Patel, A.A.; Cooper, D.R.; Murr, M.; Patel, N.A. Adipose-derived stem cells from lean and obese humans show depot specific differences in their stem cell markers, exosome contents and senescence: Role of protein kinase C delta (PKCδ) in adipose stem cell niche. Stem Cell Investig. 2016, 3, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Bonacquisti, E.E.; Brown, A.D.; Nguyen, J. Boosting the Biogenesis and Secretion of Mesenchymal Stem Cell-Derived Exosomes. Cells 2020, 9, 660. [Google Scholar] [CrossRef] [Green Version]
- Guduric-Fuchs, J.; O’Connor, A.; Camp, B.; O’Neill, C.L.; Medina, R.J.; Simpson, D.A. Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genom. 2012, 13, 357. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Duan, L.; Lu, J.; Xia, J. Engineering exosomes for targeted drug delivery. Theranostics 2021, 11, 3183–3195. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: www.clinicaltrials.gov (accessed on 8 April 2021).
- European Union Clinical Trials Register. Available online: www.clinicaltrialsregister.eu (accessed on 8 April 2021).
- Ruetze, M.; Richter, W. Adipose-derived stromal cells for osteoarticular repair: Trophic function versus stem cell activity. Expert Rev. Mol. Med. 2014, 16, e9. [Google Scholar] [CrossRef]
- Phelps, J.; Sanati-Nezhad, A.; Ungrin, M.; Duncan, N.A.; Sen, A. Bioprocessing of Mesenchymal Stem Cells and Their Derivatives: Toward Cell-Free Therapeutics. Stem Cells Int. 2018, 2018, 9415367. [Google Scholar] [CrossRef] [Green Version]
- Araldi, R.P.; D’Amelio, F.; Vigerelli, H.; De Melo, T.C.; Kerkis, I. Stem Cell-Derived Exosomes as Therapeutic Approach for Neurodegenerative Disorders: From Biology to Biotechnology. Cells 2020, 9, 2663. [Google Scholar] [CrossRef]
- Sadat, S.; Gehmert, S.; Song, Y.-H.; Yen, Y.; Bai, X.; Gaiser, S.; Klein, H.; Alt, E. The cardioprotective effect of mesenchymal stem cells is mediated by IGF-I and VEGF. Biochem. Biophys. Res. Commun. 2007, 363, 674–679. [Google Scholar] [CrossRef]
- Nakagami, H.; Maeda, K.; Morishita, R.; Iguchi, S.; Nishikawa, T.; Takami, Y.; Kikuchi, Y.; Saito, Y.; Tamai, K.; Ogihara, T.; et al. Novel Autologous Cell Therapy in Ischemic Limb Disease through Growth Factor Secretion by Cultured Adipose Tissue-Derived Stromal Cells. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2542–2547. [Google Scholar] [CrossRef]
- Kandasamy, M.; Lehner, B.; Kraus, S.; Sander, P.R.; Marschallinger, J.; Rivera, F.J.; Trümbach, D.; Ueberham, U.; Reitsamer, H.A.; Strauss, O.; et al. TGF-β signalling in the adult neurogenic niche promotes stem cell quiescence as well as generation of new neurons. J. Cell. Mol. Med. 2014, 18, 1444–1459. [Google Scholar] [CrossRef] [Green Version]
- Razavi, S.; Razavi, M.R.; Esfahani, H.Z.; Kazemi, M.; Mostafavi, F.S. Comparing brain-derived neurotrophic factor and ciliary neurotrophic factor secretion of induced neurotrophic factor secreting cells from human adipose and bone marrow-derived stem cells. Dev. Growth Differ. 2013, 55, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Zhao, L.; Zhong, J.; Gu, H.; Feng, D.; Johnstone, B.; March, K.; Farlow, M.; Du, Y. Adipose stromal cells-secreted neuroprotective media against neuronal apoptosis. Neurosci. Lett. 2009, 462, 76–79. [Google Scholar] [CrossRef]
- Park, H.; Chang, K.-A. Therapeutic Potential of Repeated Intravenous Transplantation of Human Adipose-Derived Stem Cells in Subchronic MPTP-Induced Parkinson’s Disease Mouse Model. Int. J. Mol. Sci. 2020, 21, 8129. [Google Scholar] [CrossRef]
- Kalbermatten, D.F.; Schaakxs, D.; Kingham, P.J.; Wiberg, M. Neurotrophic activity of human adipose stem cells isolated from deep and superficial layers of abdominal fat. Cell Tissue Res. 2011, 344, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Kim, H.J.; Oh, J.-H.; Park, H.-G.; Ra, J.C.; Chang, K.-A.; Suh, Y.-H. Therapeutic potentials of human adipose-derived stem cells on the mouse model of Parkinson’s disease. Neurobiol. Aging 2015, 36, 2885–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-S.; Hong, Z.-X.; Lin, S.-Z.; Harn, H.-J. Identifying Therapeutic Targets for Spinocerebellar Ataxia Type 3/Machado–Joseph Disease through Integration of Pathological Biomarkers and Therapeutic Strategies. Int. J. Mol. Sci. 2020, 21, 3063. [Google Scholar] [CrossRef] [PubMed]
- Riordan, N.H.; Ichim, T.E.; Min, W.-P.; Wang, H.; Solano, F.; Lara, F.; Alfaro, M.; Rodriguez, J.P.; Harman, R.J.; Patel, A.N.; et al. Non-expanded adipose stromal vascular fraction cell therapy for multiple sclerosis. J. Transl. Med. 2009, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Fei, K.; Tian, F.; Gao, C.; Song, Y. Adipose-derived mesenchymal stem cells attenuate ischemic brain injuries in rats by modulating miR-21-3p/MAT2B signaling transduction. Croat. Med. J. 2019, 60, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Ban, J.-J.; Yang, S.; Im, W.; Kim, M. The exosome of adipose-derived stem cells reduces β-amyloid pathology and apoptosis of neuronal cells derived from the transgenic mouse model of Alzheimer’s disease. Brain Res. 2018, 1691, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, T.; Tsuchiya, R.; Kosaka, N.; Yoshioka, Y.; Takagaki, K.; Oki, K.; Takeshita, F.; Sakai, Y.; Kuroda, M.; Ochiya, T. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci. Rep. 2013, 3, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Liu, T.; Im, W.; Kim, M. Exosomes from adipose-derived stem cells ameliorate phenotype of Huntington’s disease in vitro model. Eur. J. Neurosci. 2016, 44, 2114–2119. [Google Scholar] [CrossRef]
- Lee, M.; Ban, J.-J.; Kim, K.Y.; Jeon, G.S.; Im, W.; Sung, J.-J.; Kim, M. Adipose-derived stem cell exosomes alleviate pathology of amyotrophic lateral sclerosis in vitro. Biochem. Biophys. Res. Commun. 2016, 479, 434–439. [Google Scholar] [CrossRef]
- Laso-García, F.; Ramos-Cejudo, J.; Carrillo-Salinas, F.J.; Ortega, L.O.; Feliú, A.; Frutos, M.G.-D.; Mecha, M.; Díez-Tejedor, E.; Guaza, C.; Gutiérrez-Fernández, M. Therapeutic potential of extracellular vesicles derived from human mesenchymal stem cells in a model of progressive multiple sclerosis. PLoS ONE 2018, 13, e0202590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.A.; Moss, L.D.; Lee, J.-Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. J. Neuroinflamm. 2018, 15, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Du, Z.; Zhao, L.; Feng, D.; Wei, G.; He, Y.; Tan, J.; Lee, W.-H.; Hampel, H.; Dodel, R.; et al. IFATS Collection: The Conditioned Media of Adipose Stromal Cells Protect against Hypoxia-Ischemia-Induced Brain Damage in Neonatal Rats. Stem Cells 2009, 27, 478–488. [Google Scholar] [CrossRef]
- Kranz, A.; Rau, C.; Kochs, M.; Waltenberger, J. Elevation of Vascular Endothelial Growth Factor-A Serum Levels Following Acute Myocardial Infarction. Evidence for its Origin and Functional Significance. J. Mol. Cell. Cardiol. 2000, 32, 65–72. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, J.; Hu, X.; Wang, Z.; Wu, S.; Yi, Y. Extracellular vesicles derived from human adipose-derived stem cells promote the exogenous angiogenesis of fat grafts via the let-7/AGO1/VEGF signalling pathway. Sci. Rep. 2020, 10, 5313. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Witman, N.; Yan, D.; Zhang, S.; Zhou, M.; Yan, Y.; Yao, Q.; Ding, F.; Yan, B.; Wang, H.; et al. Human adipose-derived stem cells enriched with VEGF-modified mRNA promote angiogenesis and long-term graft survival in a fat graft transplantation model. Stem Cell Res. Ther. 2020, 11, 490. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-H.; Chen, C.-H.; Wallace, C.G.; Yuen, C.-M.; Kao, G.-S.; Chen, Y.-L.; Shao, P.-L.; Chen, Y.-L.; Chai, H.-T.; Lin, K.-C.; et al. Intravenous administration of xenogenic adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes markedly reduced brain infarct volume and preserved neurological function in rat after acute ischemic stroke. Oncotarget 2016, 7, 74537–74556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopatina, T.; Favaro, E.; Grange, C.; Cedrino, M.; Ranghino, A.; Occhipinti, S.; Fallo, S.; Buffolo, F.; Gaykalova, D.A.; Zanone, M.M.; et al. PDGF enhances the protective effect of adipose stem cell-derived extracellular vesicles in a model of acute hindlimb ischemia. Sci. Rep. 2018, 8, 17458. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.-Y.; Dass, K.T.P.; Lin, S.-Z.; Harn, H.-J.; Liu, S.-P. The application of stem cell therapy and brown adipose tissue transplantation in metabolic disorders. Cytotherapy 2020, 22, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.R.; Baptista, L.S. Adipose-derived stromal/stem cells from different adipose depots in obesity development. World J. Stem Cells 2019, 11, 147–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.P.; Anderson, A.E.; McCartney, A.; Ory, X.; Ma, G.; Pappalardo, E.; Bader, J.; Elisseeff, J.H. Metabolically Active Three-Dimensional Brown Adipose Tissue Engineered from White Adipose-Derived Stem Cells. Tissue Eng. 2017, 23, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisani, D.F.; Djedaini, M.; Beranger, G.E.; Elabd, C.; Scheideler, M.; Ailhaud, G.P.; Amri, E.-Z. Differentiation of human adipose-derived stem cells into “brite” (brown-in-white) adipocytes. Front. Endocrinol. 2011, 2, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Pan, Q.; Dong, H.; Yuan, X.; Li, Y.; Sun, Z.; Dong, X.; Wang, H. Adipose-derived mesenchymal stem cells improve glucose homeostasis in high-fat diet-induced obese mice. Stem Cell Res. Ther. 2015, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Capilla-González, V.; López-Beas, J.; Escacena, N.; Aguilera, Y.; de la Cuesta, A.; Ruiz-Salmerón, R.; Martín, F.; Hmadcha, A.; Soria, B. PDGF Restores the Defective Phenotype of Adipose-Derived Mesenchymal Stromal Cells from Diabetic Patients. Mol. Ther. 2018, 26, 2696–2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brini, A.T.; Amodeo, G.; Ferreira, L.M.; Milani, A.; Niada, S.; Moschetti, G.; Franchi, S.; Borsani, E.; Rodella, L.F.; Panerai, A.E.; et al. Therapeutic effect of human adipose-derived stem cells and their secretome in experimental diabetic pain. Sci. Rep. 2017, 7, 9904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodeo, G.; Niada, S.; Moschetti, G.; Franchi, S.; Savadori, P.; Brini, A.T.; Sacerdote, P. Secretome of human adipose-derived mesenchymal stem cell relieves pain and neuroinflammation independently of the route of administration in experimental osteoarthritis. Brain Behav. Immun. 2021, 94, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mi, Y.; Wu, S.; You, X.; Huang, Y.; Zhu, J.; Zhu, L. Exosomes from adipose-derived stem cells protect against high glucose-induced erectile dysfunction by delivery of corin in a streptozotocin-induced diabetic rat model. Regen. Ther. 2020, 14, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Wartchow, K.M.; Rodrigues, L.; Lissner, L.J.; Federhen, B.C.; Selistre, N.G.; Moreira, A.; Gonçalves, C.-A.; Sesterheim, P. Insulin-producing cells from mesenchymal stromal cells: Protection against cognitive impairment in diabetic rats depends upon implant site. Life Sci. 2020, 251, 117587. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Shi, Y.; Gong, J.; Zhao, L.; Li, Y.; He, Q.; Huang, H. Exosome secreted from adipose-derived stem cells attenuates diabetic nephropathy by promoting autophagy flux and inhibiting apoptosis in podocyte. Stem Cell Res. Ther. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Luo, Q.; Wang, Y.; Ma, Y.; Chen, F.; Zhu, X.; Shi, J. Adipose mesenchymal stem cell-derived extracellular vesicles containing microRNA-26a-5p target TLR4 and protect against diabetic nephropathy. J. Biol. Chem. 2020, 295, 12868–12884. [Google Scholar] [CrossRef]
- Gao, F.; Zuo, B.; Wang, Y.; Li, S.; Yang, J.; Sun, D. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci. 2020, 255, 117719. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 18. [Google Scholar] [CrossRef]
- Brave, H.; MacLoughlin, R. State of the Art Review of Cell Therapy in the Treatment of Lung Disease, and the Potential for Aerosol Delivery. Int. J. Mol. Sci. 2020, 21, 6435. [Google Scholar] [CrossRef]
- Shigemura, N.; Okumura, M.; Mizuno, S.; Imanishi, Y.; Nakamura, T.; Sawa, Y. Autologous transplantation of adipose tissue-derived stromal cells ameliorates pulmonary emphysema. Am. J. Transplant. 2006, 6, 2592–2600. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Kim, J.-Y.; Cho, R.; Shin, D.-M.; Lee, S.W.; Oh, Y.-M. Adipose stem cell-derived nanovesicles inhibit emphysema primarily via an FGF2-dependent pathway. Exp. Mol. Med. 2017, 49, e284. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Danchuk, S.D.; Bonvillain, R.W.; Xu, B.; Scruggs, B.A.; Strong, A.L.; Semon, J.A.; Gimble, J.M.; Betancourt, A.M.; Sullivan, D.E.; et al. Interleukin 6 Mediates the Therapeutic Effects of Adipose-Derived Stromal/Stem Cells in Lipopolysaccharide-Induced Acute Lung Injury. Stem Cells 2014, 32, 1616–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Guijo, F.; García-Arranz, M.; López-Parra, M.; Monedero, P.; Mata-Martínez, C.; Santos, A.; Sagredo, V.; Álvarez-Avello, J.-M.; Guerrero, J.E.; Pérez-Calvo, C.; et al. Adipose-derived mesenchymal stromal cells for the treatment of patients with severe SARS-CoV-2 pneumonia requiring mechanical ventilation. A proof of concept study. EClinicalMedicine 2020, 25, 100454. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.-T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Y.; Qin, H.; Tong, S.; Sun, Q.; Wang, T.; Zhang, H.; Cui, M.; Guo, S. Osteogenically-induced exosomes stimulate osteogenesis of human adipose-derived stem cells. Cell Tissue Bank. 2021, 22, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, J.; Peng, W.; Yuan, B.; Bi, Q.; Xu, Y. Exosomes from adipose-derived stem cells promote chondrogenesis and suppress inflammation by upregulating miR-145 and miR-221. Mol. Med. Rep. 2020, 21, 1881–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofiño-Vian, M.; Guillén, M.I.; Del Caz, M.D.P.; Silvestre, A.; Alcaraz, M.J. Microvesicles from Human Adipose Tissue-Derived Mesenchymal Stem Cells as a New Protective Strategy in Osteoarthritic Chondrocytes. Cell. Physiol. Biochem. 2018, 47, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.H.; Kim, H.K.; Jung, G.Y.; Jung, Y.J.; Lee, K.S.; Yun, Y.E.; Han, J.; Lee, J.; Kim, W.S.; Choi, J.S.; et al. Small extracellular vesicles from human adipose-derived stem cells attenuate cartilage degeneration. J. Extracell. Vesicles 2020, 9, 1735249. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Song, W.; Chen, B.; Liu, X.; He, Y. Exosomes Isolated from Adipose-Derived Stem Cells: A New Cell-Free Approach to Prevent the Muscle Degeneration Associated with Torn Rotator Cuffs. Am. J. Sports Med. 2019, 47, 3247–3255. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, J.A.; Harmsen, M.C.; Van Der Lei, B.; Stevens, H.P. Augmentation of Dermal Wound Healing by Adipose Tissue-Derived Stromal Cells (ASC). Bioengineering 2018, 5, 91. [Google Scholar] [CrossRef] [Green Version]
- Stessuk, T.; Puzzi, M.B.; Chaim, E.A.; Alves, P.C.M.; De Paula, E.V.; Forte, A.; Izumizawa, J.M.; Oliveira, C.C.; Frei, F.; Ribeiro-Paes, J.T. Platelet-rich plasma (PRP) and adipose-derived mesenchymal stem cells: Stimulatory effects on proliferation and migration of fibroblasts and keratinocytes in vitro. Arch. Dermatol. Res. 2016, 308, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Seo, G.; Oh, E.; Yun, M.; Lee, J.-Y.; Bae, J.S.; Joo, K.; Chae, G.T.; Lee, S.-B. Adipose-derived stem cell conditioned medium accelerates keratinocyte differentiation via the upregulation of miR-24. Exp. Dermatol. 2015, 24, 792–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, C.-M.; Chen, Y.-C.; Chen, Y.-C.; Lee, T.-L.; Peng, Y.-S.; Chen, S.-H.; Yen, Y.-H.; Chien, C.-L.; Hsieh, J.-H.; Chen, Y.-L. Interleukin-6 from Adipose-Derived Stem Cells Promotes Tissue Repair by the Increase of Cell Proliferation and Hair Follicles in Ischemia/Reperfusion-Treated Skin Flaps. Mediat. Inflamm. 2019, 2019, 2343867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Han, Y.-D.; Yan, X.-L.; Ren, J.; Zeng, Q.; Li, X.-D.; Pei, X.-T.; Han, Y. Adipose mesenchymal stem cell-derived exosomes stimulated by hydrogen peroxide enhanced skin flap recovery in ischemia-reperfusion injury. Biochem. Biophys. Res. Commun. 2018, 500, 310–317. [Google Scholar] [CrossRef]
- Yang, C.; Luo, L.; Bai, X.; Shen, K.; Liu, K.; Wang, J.; Hu, D. Highly-expressed micoRNA-21 in adipose derived stem cell exosomes can enhance the migration and proliferation of the HaCaT cells by increasing the MMP-9 expression through the PI3K/AKT pathway. Arch. Biochem. Biophys. 2020, 681, 108259. [Google Scholar] [CrossRef]
- Mazini, L.; Rochette, L.; Malka, G. Adipose-Derived Stem Cells (ADSCs) and Growth Differentiation Factor 11 (GDF11): Regenerative and Antiaging Capacity for the Skin. Regen. Med. 2020. [Google Scholar] [CrossRef]
- Lv, Q.; Deng, J.; Chen, Y.; Wang, Y.; Liu, B.; Liu, J. Engineered Human Adipose Stem-Cell-Derived Exosomes Loaded with miR-21-5p to Promote Diabetic Cutaneous Wound Healing. Mol. Pharm. 2020, 17, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.S.; Kim, J.O.; Ha, D.H.; Yi, Y.W. Exosomes derived from human adipose tissue-derived mesenchymal stem cells alleviate atopic dermatitis. Stem Cell Res. Ther. 2018, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.-O.; Ha, D.H.; Kim, J.O.; Crumrine, D.A.; Meyer, J.M.; Wakefield, J.S.; Lee, Y.; Kim, B.; Kim, S.; Kim, H.-K.; et al. Exosomes from Human Adipose Tissue-Derived Mesenchymal Stem Cells Promote Epidermal Barrier Repair by Inducing De Novo Synthesis of Ceramides in Atopic Dermatitis. Cells 2020, 9, 680. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Fan, Y.; Lin, G.; Park, Y.K.; Pak, C.S.; Jeong, J.H.; Kim, S. Synergistic Effect of Adipose-Derived Stem Cells and Fat Graft on Wrinkles in Aged Mice. Plast. Reconstr. Surg. 2019, 143, 1637–1646. [Google Scholar] [CrossRef]
- Guo, S.; Wang, T.; Zhang, S.; Chen, P.; Cao, Z.; Lian, W.; Guo, J.; Kang, Y. Adipose-derived stem cell-conditioned medium protects fibroblasts at different senescent degrees from UVB irradiation damages. Mol. Cell. Biochem. 2020, 463, 67–78. [Google Scholar] [CrossRef]
- Li, L.; Ngo, H.T.; Hwang, E.; Wei, X.; Liu, Y.; Liu, J.; Yi, T.-H. Conditioned Medium from Human Adipose-Derived Mesenchymal Stem Cell Culture Prevents UVB-Induced Skin Aging in Human Keratinocytes and Dermal Fibroblasts. Int. J. Mol. Sci. 2019, 21, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NCT no. | Title | Status | Condition/ Disease | Administration | Intervention/ Treatment | Results |
|---|---|---|---|---|---|---|
| NCT04313647 | A tolerance clinical study on aerosol inhalation of mesenchymal stem cells exosomes in healthy volunteers | Recruiting | Healthy volunteers | Aerosol inhalation | 2 × 108, 4 × 108, 8 × 108, 16 × 108, 20 × 108 nano vesicles/3 mL to be administrated at once to different participants sets | N/A |
| NCT04388982 | Open-label, single-center, phase I/Ⅱ clinical trial to evaluate the safety and the efficacy of exosomes derived from allogenic adipose mesenchymal stem cells in patients with mild to moderate dementia due to Alzheimer’s disease | Recruiting | Dementia due to Alzheimer’s disease | Nasal drip | 5 µg, 10 µg, 20 µg exosomes/1 mL exosomes, twice a week for 12 weeks | N/A |
| NCT04544215 | A clinical study of allogeneic human adipose-derived mesenchymal progenitor cell exosomes (haMPC-Exos) nebulizer for the treatment of carbapenem-resistant gram-negative bacilli-induced pulmonary infection | Recruiting | Drug resistant pulmonary infection | Aerosol inhalation | 8 × 108, 16 × 108 nano vesicles/3 mL per day for 7 days | N/A |
| NCT04276987 | A pilot clinical study on aerosol inhalation of the exosomes derived from allogenic adipose mesenchymal stem cells in the treatment of severe patients with novel coronavirus pneumonia | Completed | Coronavirus pneumonia | Aerosol inhalation | 2 × 108 nano vesicles/3 mL per day for 5 days | N/A |
| Paracrine Factors | Model | Therapeutic Effect | References |
|---|---|---|---|
| Neurodegenerative Diseases | |||
| BDNF, GDNF | Mouse Parkinson disease model | Protection of dopaminergic neurons. | [128] |
| Exosomes | Mice Alzheimer disease Model | Reduction of amyloid beta (Aβ) levels and the Aβ42/40. Increasing cell survival and decrease of neuronal death in the hippocampus and the cerebral cortex. Decrease deposition of Aβ plaques in Alzheimer Disease by exosomal neprilysin. | [134,135] |
| Exosomes | Mice-derived neural cells from Huntington’s disease mouse model | Reduction of accumulation of mHtt aggregates, mitochondrial dysfunction and cell apoptosis by p-CREB-PGC1α pathway activation in Huntington’s disease. | [136] |
| Exosomes | Mice-derived neuronal cells from Amyotrophic lateral sclerosis disease mouse model | Reduction of cytosolic superoxide dismutase 1 and restore of the abnormal reduction of mitochondrial proteins. | [137] |
| Exosomes | Mouse progressive Multiple Sclerosis model | Brain atrophy reduction and remyelination promotion. Reduction of Th1 and Th17 levels. | [138] |
| Exosomes | Brain injured male Sprague-Dawley rats | Reduction of inflammatory indicator (Ly6G+/CD11b/c+) and immune (CD3+/CD4+/CD3+/CD8+) cells. Reduction of early and late apoptotic cells. | [31] |
| Exosomes (containing or depleted of MALAT1) | Rats following a mild controlled cortical impact | Modulation of inflammation-related after traumatic brain injury by MALAT1. | [139] |
| Cardiovascular diseases | |||
| Exosomes | Endothelial cells | Promotion of vascularization by overexpressing miR-21. | [30] |
| VEGF | Human microvascular endothelial cells and mouse Hindlimb Ischemia model | Reduction of endothelial cells apoptosis. Perfusion improvement in ischemic hindlimbs. | [95] |
| IGF-1, BDNF | Cerebellar granule neurons | Blocking postischemic p38 activation. Protection of neural cells. | [140] |
| Exosomes | Rat Acute Ischemic Stroke model | Increase of neural regeneration. Reduction of brain infarct zone, brain swelling, and shrinkage. | [144] |
| Exosomes released by PDGF-stimulated adipose-derived stem cells | Acute Hindlimb Ischemia mouse model | Muscle protection from Acute Ischemia. | [145] |
| Engineered modified exosomes encoding VEGF | Nude mouse fat transplantation model | Improvement of neo-angiogenesis and vascularization by promoting cell proliferation and vascular maturity. | [143] |
| Exosomes | Vascular endothelial cells and nude mice transplantation model | Promoting exogenous angiogenesis. Increase the proliferation, migration, tube formation, and VEGF secretion. Improving the survival of fat grafts. | [142] |
| Metabolic diseases | |||
| PDGF | Diabetic patients adipose-derived stem cells and SCID Wound mice model | Increased proliferation, migration, and homing to sites of inflammation. | [151] |
| Exosomes | Diabetic rat model | Restored erectile function due to corin content in exosomes. | [154] |
| Exosomes | Mouse podocyte MPC5 cells and spontaneous diabetes mice | Reducing high glucose-induced increase of cell death. Reduction of urine protein, serum creatinine, blood urea nitrogen, and podocyte apoptosis. | [156] |
| Exosomes (miR-26a-5p) | Mouse podocyte MPC5 cells and spontaneous diabetes mice | Protection of cells from injury. Protection against diabetic nephropathy. | [157] |
| Exosomes | Peritoneal macrophages and obesity mouse model | Improving obesity-related inflammation and metabolism. | [60] |
| Exosomes | Sepsis-induced acute kidney injury mouse model | Renal Protective effect in acute kidney injury. | [158] |
| Respiratory diseases | |||
| HGF | Pulmonary emphysema rat model | Increased alveolar and vascular repair. | [161] |
| Artifical Nanovesicles | MLE-12 epithelial cells and elastase-induced emphysema mice | Proliferation increase. Inhibition of emphysema primarily by an FGF2-dependent pathway. | [162] |
| Exosomes | Histone-induced acute lung injury mice | Improvement of survival. Inhibition histone-mediated lung hemorrhage edema, Reduction of vascular hyper-permeability. | [80] |
| Skeletal tissue regeneration | |||
| Exosomes | Adipose-derived stem cells | Induction of osteogenic differentiation. | [166] |
| Exosomes | Synovial fibroblasts and periosteal cells | Chondrogenesis promotion. Increased Collagen type II and β-catenin expression. Increased proliferation (miR-145) and chondrogenic potential (miR-221). | [167] |
| Conditioned Media | Osteoarthritic mice model | Rapid and lasting pain relief. No effect on tissue regeneration. | [153] |
| Exosomes | Chondrocytes from osteoarthritic patients | Reduction of MMP-1, MMP-3, MMP-13 and ADAMTS-5. Increased collagen II expression. | [169] |
| Exosomes | Chondrocytes from osteoarthritic patients | Reduction of MMP-3 expression. Increased collagen II expression. | [168] |
| Whole secretome/isolated exosomes | Mouse myoblast cell line C2C12 and C57BL/6 mice | Increased cell proliferation, skeletal muscle differentiation and migration. Enhancing of regeneration of skeletal muscle in acute damage. | [165] |
| Exosomes | Massive rotator cuff tear rat model | Prevention of atrophy, fatty infiltration, inflammation, and vascularization of muscles. | [170] |
| Wound healing and skin aging | |||
| Platelet-rich plasma and conditioned Media | Fibroblasts and keratinocytes isolated from skin sample | Increased cell proliferation and migration. | [172] |
| Conditioned Media | Human epidermal keratinocyte neonatal cells | Acceleration of keratinocyte differentiation via miR-24 upregulation. | [173] |
| Exosomes | Human keratinocyte cell line HaCaT | Increased migration and proliferation by exosomal miR-21. Enhancing MMP-9 expression. Promotion of wound healing. | [176] |
| Exosomes | Human keratinocyte cell line HaCaT | TGF-β pathway regulation by miR-19b. Promotion of wound healing. | [79] |
| Exosomes | Human keratinocyte cell line HaCaT | Promotion of wound healing via Wnt/β-catenin pathway. Increased proliferation and migration. Apoptosis inhibition. | [78] |
| Conditioned media | Human keratinocyte cell line HaCaTs and normal human dermal fibroblasts | Anti-photoaging activity. Reduced IL-6 secretion. | [183] |
| Engineered modified exosomes miR-21-5p | Human keratinocyte cell line HaCaT and fullthickness skin defects diabetic rat model | Promotion of wound healing via Wnt/β-catenin pathway. Improved wound healing by re-epithelialization, collagen remodeling, angiogenesis, and vessel maturation. | [178] |
| Conditioned Media | Ischemia/reperfusion flap mice model | Increased cell proliferation and the number of hair follicles. Prevention from flap necrosis after skin flap transplantation. | [174] |
| Exosomes | Human umbilical vein endothelial cell | Reduction of inflammation and apoptosis. Enhancing skin flap recovery. | [175] |
| Exosomes | Human embryonic kidney 293 cells | Prolonged the survival of vascularized composite allografts after transplantation. Downregulation of CD4 + T and Th1 cells. Upregulation Tr1 and Treg cell. | [32] |
| Conditioned media | Human dermal fibroblasts | Reduced cellular senescence of skin cells. Improved collagen I, collagen III, elastin, and TIMP-1 expression. | [182] |
| Exosomes | Atopic dermatitis mouse model | Decreased level of IgE and eosinophiles in blood and CD86+ and CD206+ cells in skin lesion. Reduction of (IL)-4, IL-23, IL-31. | [179] |
| Exosomes | Chronic allergic dermatitis mouse model | Promotion of epidermal barrier repair. Reduction of IL-5, IL-13, TNF-α, IFN-γ, IL-17, and TSLP. | [180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trzyna, A.; Banaś-Ząbczyk, A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules 2021, 11, 878. https://doi.org/10.3390/biom11060878
Trzyna A, Banaś-Ząbczyk A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules. 2021; 11(6):878. https://doi.org/10.3390/biom11060878
Chicago/Turabian StyleTrzyna, Anna, and Agnieszka Banaś-Ząbczyk. 2021. "Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”" Biomolecules 11, no. 6: 878. https://doi.org/10.3390/biom11060878