
Studies on the Catalytic Properties of Crude Freeze-Dried Preparations of Yarrowia lipolytica Extracellular Lipases for Geranyl Ester Derivative Synthesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganisms
2.3. Yeast Cultures
2.4. Freeze-Drying of Supernatant and Biomass of Y. lipolytica KKP 379
2.5. Determination of Water Activity in Lyophilisates
2.6. Determination of the Hydrolytic Activity of Freeze-Dried Supernatants
2.7. Geranyl 4-Hydroxyphenylpropanoate Enzymatic Synthesis
2.8. Evaluation of Antioxidant Activity of Geranyl 4-Hydroxyphenylpropanoate
2.9. Evaluation of Antiobacterial Activity of Geranyl 4-Hydroxyphenylpropanoate
2.10. Calculation of Lipophilicity Properties of Tested Compounds
2.11. Statistical Analysis
3. Results and Discussion
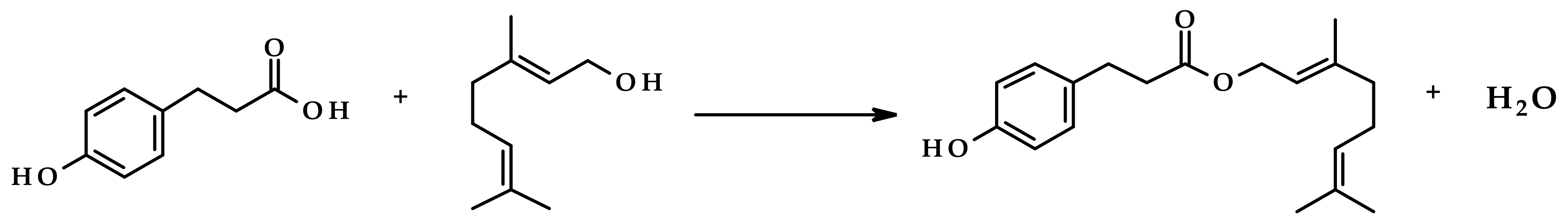
3.1. The Impact of Water Activity in the Crude Freeze-Dried Enzyme Preparations on Their Hydrolytic Activity
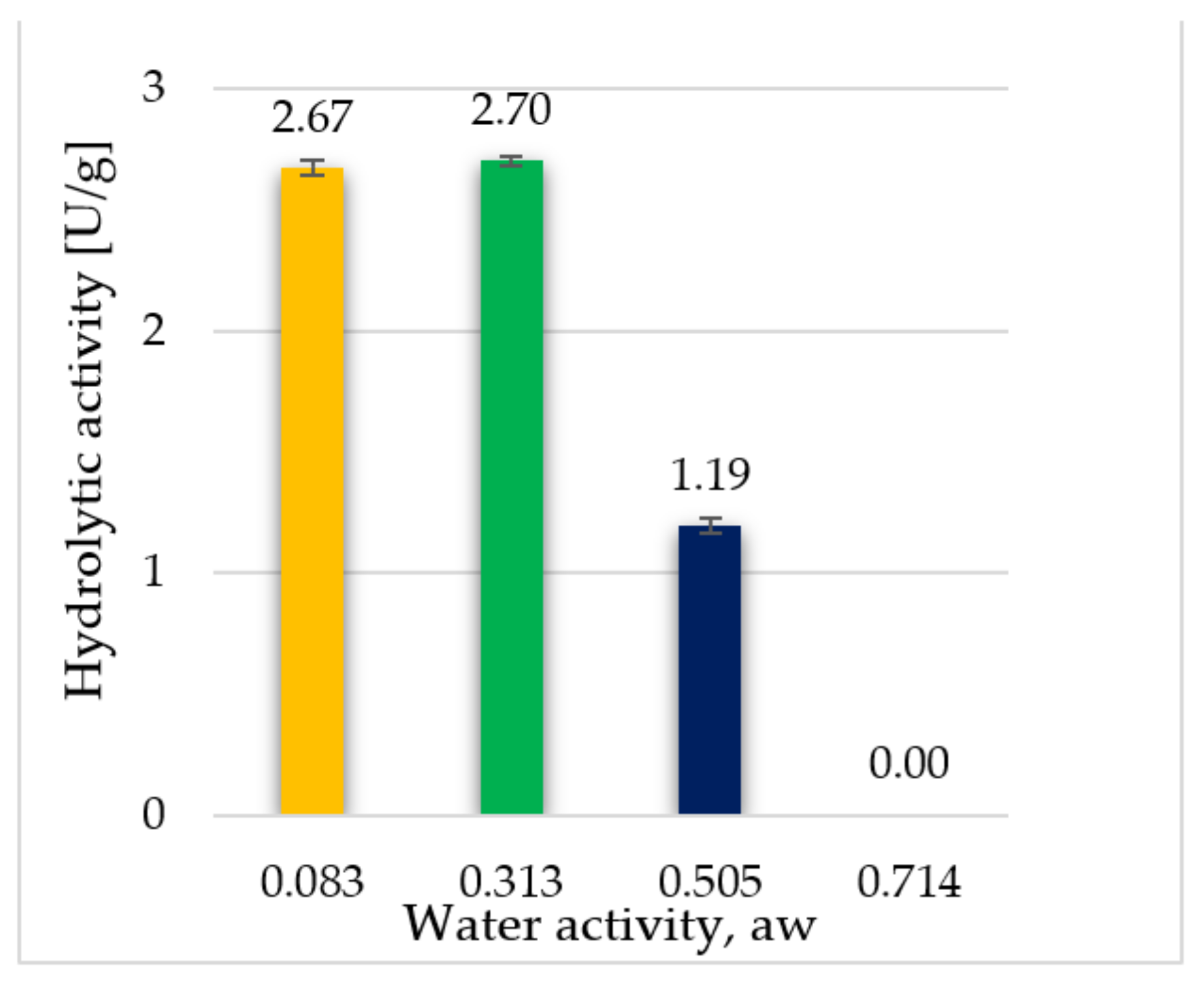
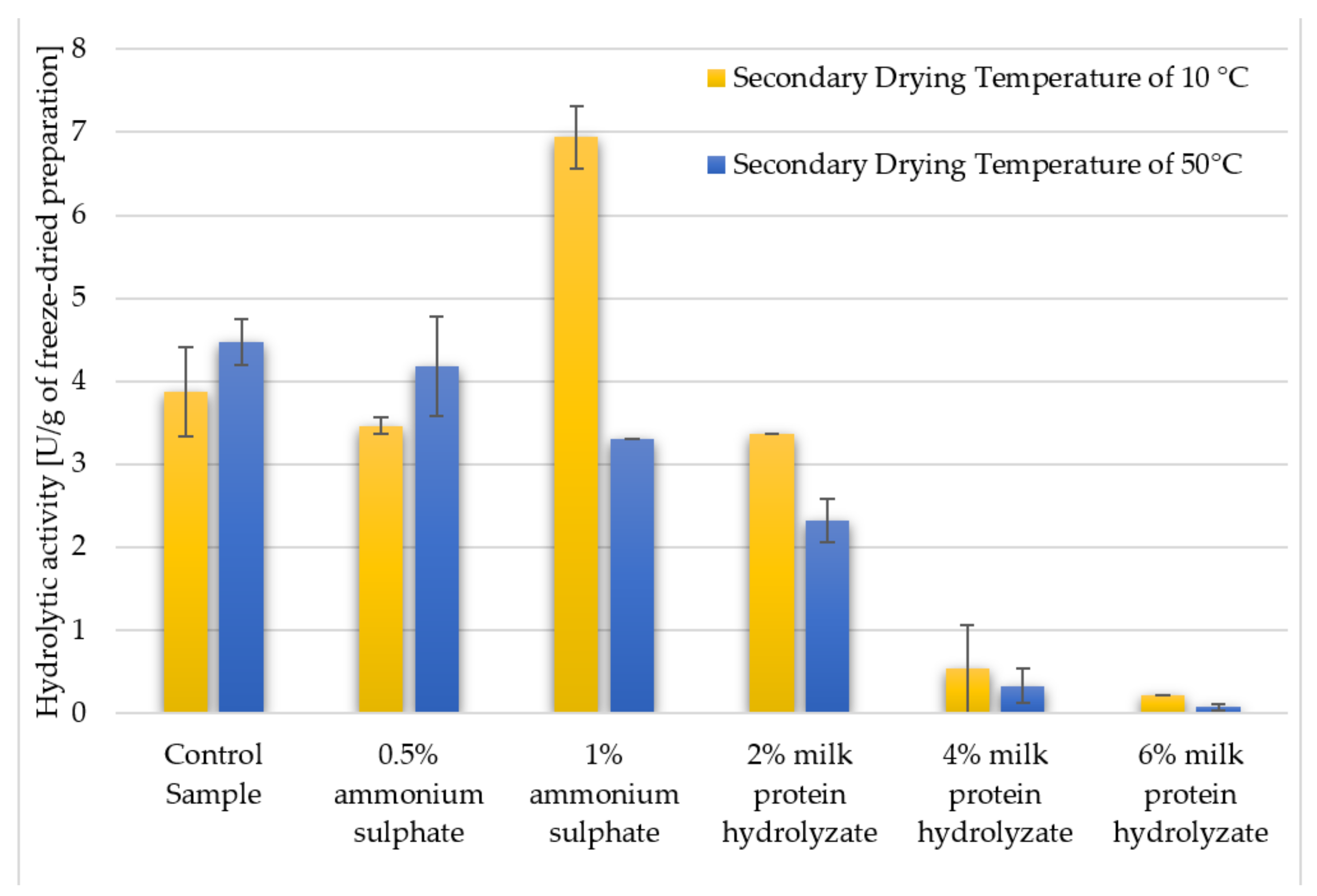
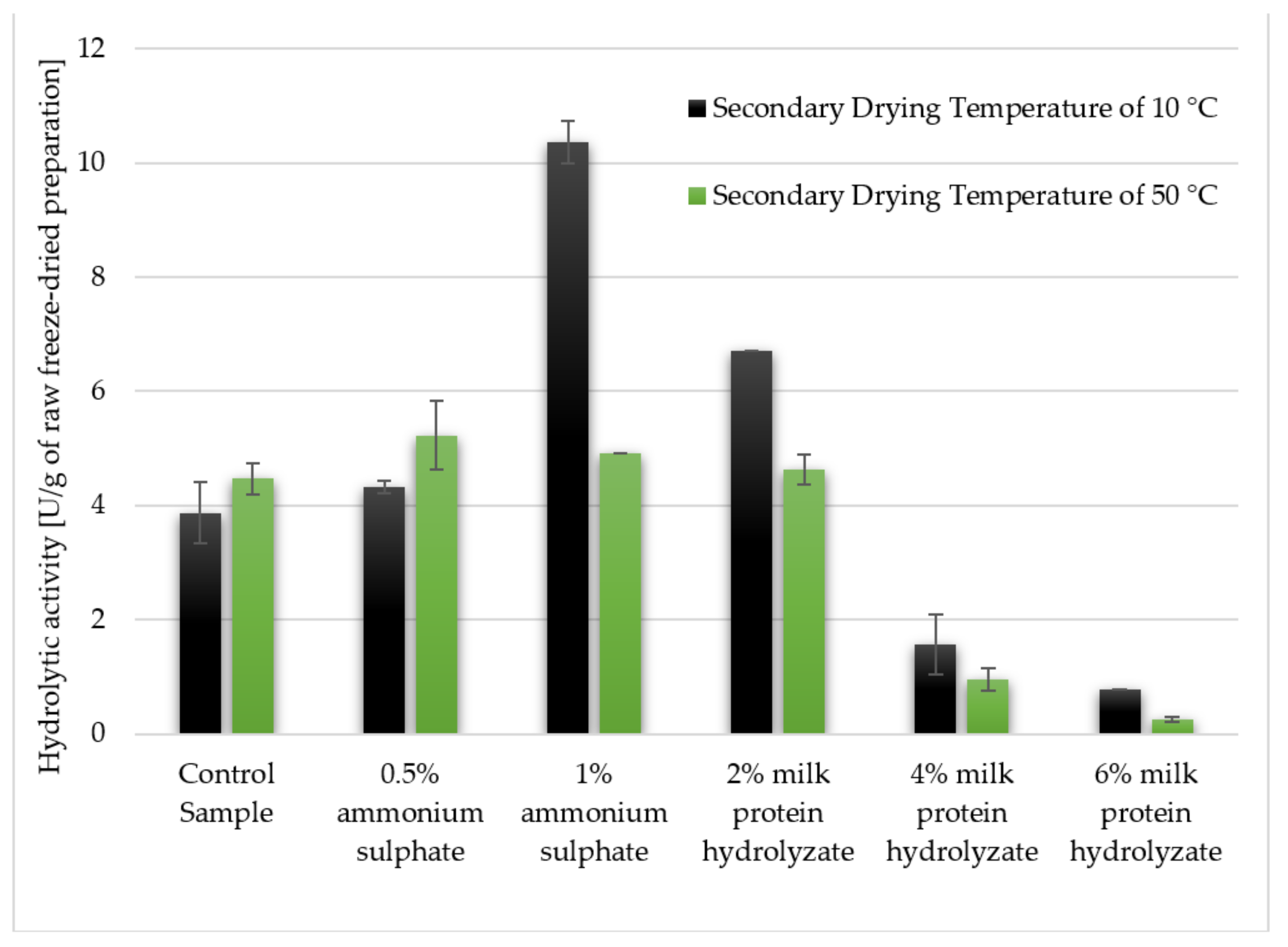
3.2. The Hydrolytic Activity of Crude Freeze-Dried Enzyme Preparations
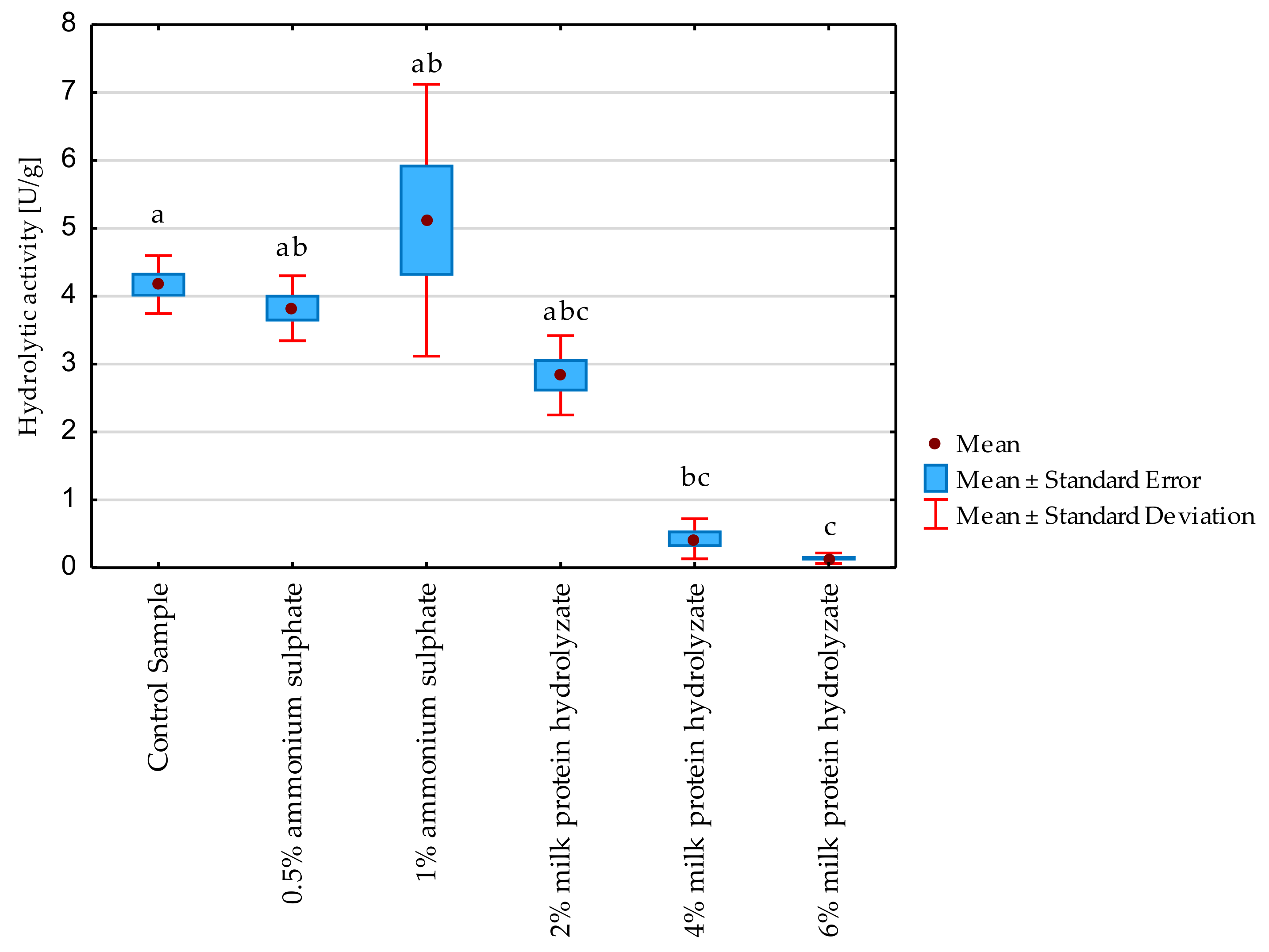
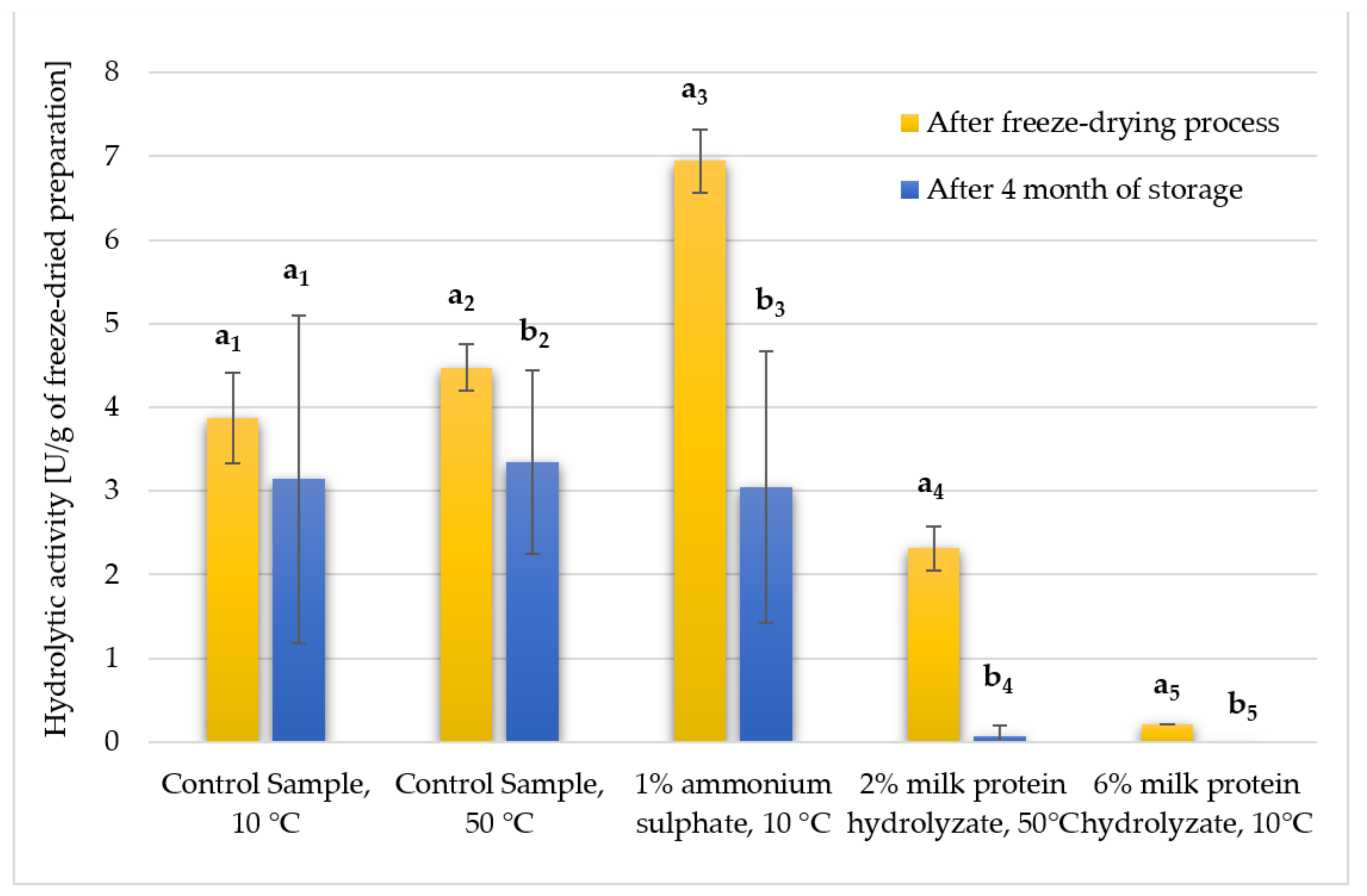
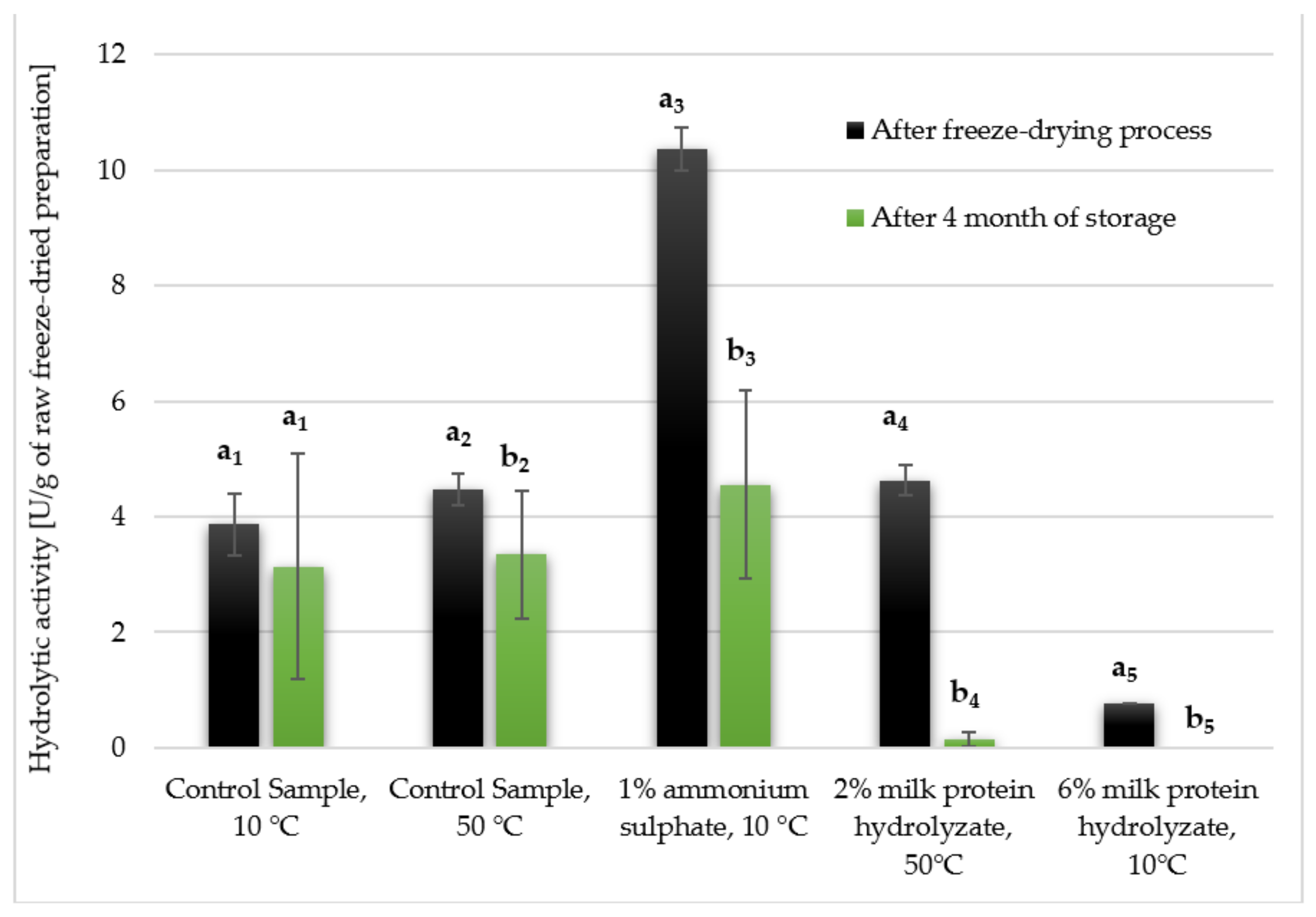
3.3. Hydrolytic Activity of Freeze-Dried Enzyme Preparations after Storage for 4 Months
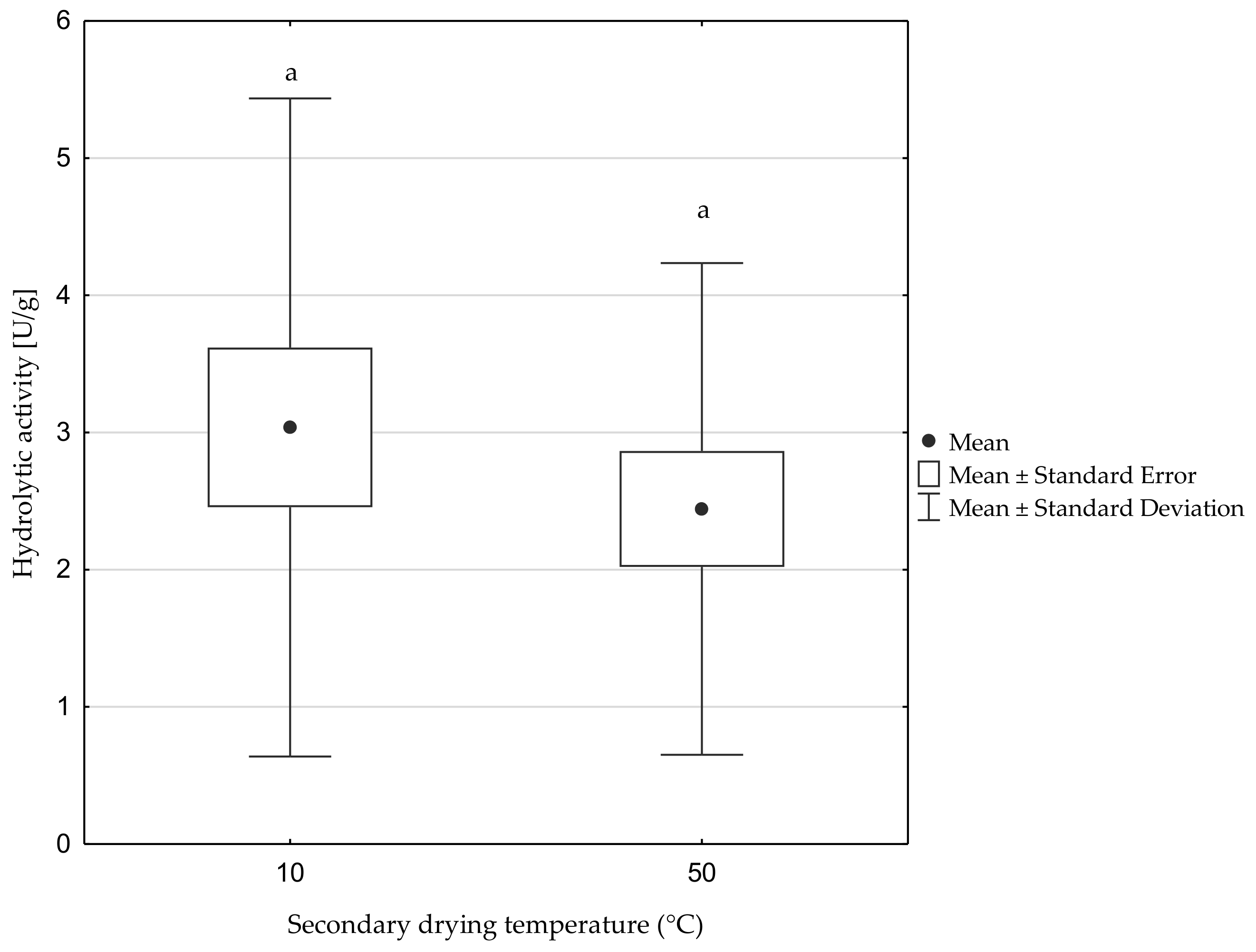
3.4. Esterification of 4-Hydroxyphenylpropanoic Acid with Geraniol
3.5. Evaluation of Ester Biological Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandra, P.; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Hasan, F.; Shah, A.A.; Hameed, A. Industrial applications of microbial lipases. Enzyme Microb. Technol. 2006, 39, 235–251. [Google Scholar] [CrossRef]
- Hasan, F.; Shah, A.A.; Hameed, A. Methods for detection and characterization of lipases: A comprehensive review. Biotechnol. Adv. 2009, 27, 782–798. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Eggert, T. Lipases for biotechnology. Curr. Opin. Biotechnol. 2002, 13, 390–397. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A. Yarrowia lipolytica: A beneficious yeast in biotechnology as a rare opportunistic fungal pathogen: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolarzewicz, I.A.; Zaborniak, P.; Fabiszewska, A.U.; Białecka-Florjańczyk, E. Study on the properties of immobilized biocatalysts with lipase activity produced by Yarrowia lipolytica in batch culture. Chem. Biochem. Eng. Q. 2017, 31, 251–259. [Google Scholar] [CrossRef]
- Brigida, A.I.S.; Amaral, P.F.F.; Coelho, M.A.Z.; Goncalves, L.R.B. Lipase from Yarrowia lipolytica: Production, characterization and application as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2014, 101, 148–158. [Google Scholar] [CrossRef]
- Pignede, G.; Wang, H.; Fudalej, F.; Gaillardin, C.; Seman, M.; Nicaud, J.M. Characterization of an extracellular lipase encoded by LIP2 in Yarrowia lipolytica. J. Bacteriol. 2000, 182, 2802–2810. [Google Scholar] [CrossRef] [Green Version]
- Zieniuk, B.; Fabiszewska, A.; Wołoszynowska, M.; Białecka-Florjańczyk, E. Synthesis of flavor compound ethyl hydrocinnamate by Yarrowia lipolytica lipases. Biocatal. Biotransform. 2020. [Google Scholar] [CrossRef]
- Samborska, K.; Witrowa-Rajchert, D.; Goncalves, A. Spray-Drying of α-Amylase—The Effect of Process Variables on the Enzyme Inactivation. Dry. Technol. 2007, 23, 941–953. [Google Scholar] [CrossRef]
- Ciurzyńska, A.; Lenart, A. Freeze-Drying—Application in Food Processing and Biotechnology—A Review. Pol. J. Food Nutr. Sci. 2011, 61, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Zieniuk, B.; Wołoszynowska, M.; Białecka-Florjańczyk, E.; Fabiszewska, A. Application of freeze-dried Yarrowia lipolytica biomass in the synthesis of lipophilic antioxidants. Biotechnol. Lett. 2021, 43, 601–612. [Google Scholar] [CrossRef]
- Zieniuk, B.; Wołoszynowska, M.; Białecka-Florjańczyk, E.; Fabiszewska, A. Synthesis of Industrially Useful Phenolic Compounds Esters by Means of Biocatalysts Obtained Along with Waste Fish Oil Utilization. Sustainability 2020, 12, 5804. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A.; Białecka-Florjańczyk, E. Screening of solvents for favoring hydrolytic activity of Candida antarctica Lipase B. Bioproc. Biosyst. Eng. 2020, 43, 605–613. [Google Scholar] [CrossRef]
- Nowak, D.; Piechucka, P.; Witrowa-Rajchert, D.; Wiktor, A. Impact of material structure on the course of freezing and freeze-drying and on the properties of dried substance, as exemplified by celery. J. Food Eng. 2016, 180, 22–28. [Google Scholar] [CrossRef]
- Kapturowska, A.; Stolarzewicz, I.; Krzyczkowska, J.; Białecka-Florjańczyk, E. Studies on lipolytic activity of sonicated enzymes from Yarrowia lipolytica. Ultrason. Sonochem. 2012, 19, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Zieniuk, B.; Groborz, K.; Wołoszynowska, M.; Ratusz, K.; Białecka-Florjańczyk, E.; Fabiszewska, A. Enzymatic Synthesis of Lipophilic Esters of Phenolic Compounds, Evaluation of Their Antioxidant Activity and Effect on the Oxidative Stability of Selected Oils. Biomolecules 2021, 11, 314. [Google Scholar] [CrossRef]
- International Organization for Standards. ISO 20776–1. Clinical Laboratory Testing and In Vitro Diagnostic Test Systems–Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices–Part 1: Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; International Organization for Standards: Geneva, Switzerland, 2006. [Google Scholar]
- Páez, B.C.; Medina, A.R.; Rubio, F.C.; Moreno, P.G.; Grima, E.M. Modeling the effect of free water on enzyme activity in immobilized lipase-catalyzed reactions in organic solvents. Enzyme Microb. Technol. 2003, 33, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Bancerz, R.; Ginalska, G. Środowisko niewodne—Miejsce katalizy enzymatycznej. Biotechnologia 2009, 4, 168–182. [Google Scholar]
- Shkidchenko, A.N.; Nikitin, V.A. Correlation between the Cellular Content of Mobile Water and the Viability of Lyophilized Yeast Cells. Microbiology 2004, 73, 431–434. [Google Scholar] [CrossRef]
- Fickers, P.; Fudalej, F.; Dall, M.T.; Casaregola, S.; Gaillardin, C.; Thonart, P.; Nicaud, J.M. Identification and characterisation of LIP7 and LIP8 genes encoding two extracellular triacylglycerol lipases in the yeast Yarrowia lipolytica. Fungal Genet. Biol. 2006, 42, 264–274. [Google Scholar] [CrossRef]
- Abadias, M.; Benabarre, A.; Teixido, N.; Usall, J.; Vinas, I. Effect of freeze drying and protectants on viability of the biocontrol yeast Candida sake. Int. J. Food Microbiol. 2001, 65, 173–182. [Google Scholar] [CrossRef]
- Hubalek, Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003, 46, 205–229. [Google Scholar] [CrossRef]
- Utami, T.S.; Hariyani, I.; Alamsyah, G.; Hermansyah, H. Production of dry extract extracellular lipase from Aspergillus niger by solid state fermentation method to catalyze biodiesel synthesis. Energy Procedia 2017, 136, 41–46. [Google Scholar] [CrossRef]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Klibanov, A.M. Stabilization of Enzymes against Thermal Inactivation. Adv. Appl. Microbiol. 1983, 29, 1–28. [Google Scholar] [PubMed]
- Nowak, D.; Jakubczyk, E. The Freeze-Drying of Foods—The Characteristic of the Process Course and the Effect of Its Parameters on the Physical Properties of Food Materials. Foods 2020, 9, 1488. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto-Shinohara, Y.; Nozawa, F.; Sukenobe, J.; Imaizumi, T. Survival of yeasts stored after freeze-drying or liquid-drying. J. Gen. Appl. Microbiol. 2010, 56, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Paroul, N.; Grzegozeski, L.P.; Chiaradia, V.; Treichel, H.; Cansian, R.L.; Oliveira, J.V.; de Oliveira, D. Solvent-free geranyl oleate production by enzymatic esterification. Bioprocess Biosyst. Eng. 2010, 34, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Kirdi, R.; Ben Akacha, N.; Gargouri, M. Biosynthesis Coupled to the Extraction of Geranyl Acetate in a Liquid-Gas System: Optimization of the Transesterification Reaction and Modeling of the Transfer. Chem. Biochem. Eng. Q. 2016, 30, 117–125. [Google Scholar] [CrossRef]
- Da Silva Correa, L.; Henriques, R.O.; Rios, J.V.; Lerin, L.A.; de Oliveira, D.; Furigo, A., Jr. Lipase-Catalyzed Esterification of Geraniol and Citronellol for the Synthesis of Terpenic Esters. Appl. Biochem. Biotechnol. 2020, 190, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Krzyczkowska, J.; Fabiszewska, A.U. Yarrowia lipolytica—Niekonwencjonalne Drożdże w Biotechnologii. Post. Mikrobiol. 2015, 54, 33–43. [Google Scholar]
- Białecka-Florjańczyk, E.; Krzyczkowska, J.; Stolarzewicz, I.; Kapturowska, A. Synthesis of 2-phenylethyl acetate in the presence of Yarrowia lipolytica KKP 379 biomass. J. Mol. Catal. B Enzym. 2012, 74, 241–245. [Google Scholar] [CrossRef]
- Fantin, G.; Fogagnolo, M.; Guerrini, A.; Medici, A.; Pedrini, P.; Fontana, S. Enantioselective hydrolyses with Yarrowia lipolytica: A versatile strain for esters, enol esters, epoxides, and lactones. Tetrahedron Asymmetry 2001, 12, 2709–2713. [Google Scholar] [CrossRef]
- Reis, B.; Martins, M.; Barreto, B.; Milhazes, N.; Garrido, E.M.; Silva, P.; Garrido, J.; Borges, F. Structure-property-activity relationship of phenolic acids and derivatives. Protocatechuic acid alkyl esters. J. Agric. Food Chem. 2010, 58, 6986–6993. [Google Scholar] [CrossRef] [PubMed]
- Konate, K.; Hilou, A.; Mavoungou, J.F.; Lepengue, A.N.; Souza, A.; Barro, N.; Datte, J.Y.; M’Bachti, B.; Nacoulma, O.G. Antimicrobial activity of polyphenol-rich fractions from Sida alba L. (Malvaceae) against co-trimoxazol-resistant bacteria strains. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieniuk, B.; Białecka-Florjańczyk, E.; Fabiszewska, A. Anti-Listerial Effect of 4-Hydroxyphenylpropanoic Acid Esters Synthesized by Lipase-Catalyzed Esterification. Proceedings 2021, 70, 17. [Google Scholar] [CrossRef]
- Merkl, R.; Hradkova, I.; Filip, V.; Smidrkal, J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J. Food Sci. 2010, 28, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Shi, Y.; Gu, Q.; Fang, M.; Chen, Y.; Fang, S.; Dang, Y.; Chen, J. Antimicrobial effect and mechanism of non-antibiotic alkyl gallates against Pseudomonas fluorescens on the surface of Russian sturgeon (Acipenser gueldenstaedti). Int. J. Food Microbiol. 2021, 342, 109093. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-W.; Liao, C.-C.; Hung, K.-C.; Wang, Z.-Y.; Tung, Y.-T.; Wu, J.-H. Proteomics Reveals Octyl Gallate as an Environmentally Friendly Wood Preservative Leading to Reactive Oxygen Species-Driven Metabolic Inflexibility and Growth Inhibition in White-Rot Fungi (Lenzites betulina and Trametes versicolor). J. Fungi 2021, 7, 145. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of the Freeze-Dried Enzyme Preparation | Temperature of Secondary Drying (°C) | Water Activity aw |
|---|---|---|
| control sample—preparation without additives | 10 | 0.160 |
| control sample—preparation without additives | 50 | 0.097 |
| freeze-dried supernatant with the addition of 0.5% ammonium sulphate | 10 | 0.141 |
| freeze-dried supernatant with the addition of 0.5% ammonium sulphate | 50 | 0.117 |
| freeze-dried supernatant with the addition of 1% ammonium sulphate | 10 | 0.156 |
| freeze-dried supernatant with the addition of 1% ammonium sulphate | 50 | 0.122 |
| freeze-dried supernatant with the addition of 2% milk protein hydrolyzate | 10 | 0.099 |
| freeze-dried supernatant with the addition of 2% milk protein hydrolyzate | 50 | 0.091 |
| freeze-dried supernatant with the addition of 4% milk protein hydrolyzate | 10 | 0.050 |
| freeze-dried supernatant with the addition of 4% milk protein hydrolyzate | 50 | 0.055 |
| freeze-dried supernatant with the addition of 6% milk protein hydrolyzate | 10 | 0.052 |
| freeze-dried supernatant with the addition of 6% milk protein hydrolyzate | 50 | 0.095 |
| Water Activity in the Desiccator | Water Activity of the Freeze-Dried Enzyme Preparations |
|---|---|
| 0.000 | 0.083 |
| 0.329 | 0.313 |
| 0.529 | 0.505 |
| 0.750 | 0.714 |
| Biocatalyst | Esterification Yield (%) |
|---|---|
| Lipase B from C. antarctica | 73.4 ± 3.7 A |
| Freeze-dried biomass of Y. lipolytica KKP 379 | 47.9 ± 1.9 B |
| Freeze-dried supernatant of Y. lipolytica KKP 379 | 5.3 ± 0.3 C |
| Compound | DPPH• Method | CUPRAC Method | |
|---|---|---|---|
| Antioxidant Activity (%) | IC50 (mM) | TEAC | |
| 4-Hydroxyphenylpropanoic acid | 32.22 ± 2.70 A | 81.20 ± 5.69 A | 0.746 ± 0.039 A |
| Geranyl 4-hydroxyphenylpropanoate | 28.71 ± 1.82 A | 90.25 ± 3.64 A | 0.513 ± 0.035 B |
| Geraniol | 2.43 ± 0.73 B | n.a. | 0.045 ± 0.003 C |
| 4HPPA | Geraniol | G4HPP | ||
|---|---|---|---|---|
| L. monocytogenes PCM 2191 | MIC (mM) | 16 | 16 | 0.125 |
| MBC (mM) | 32 | 16 | 0.5 | |
| MBC/MIC | 2 | 1 | 4 | |
| B. subtilis PCM 486 | MIC (mM) | 8 | 8 | 2 |
| MBC (mM) | >64 | >64 | >64 | |
| MBC/MIC | >16 | >16 | >16 | |
| K. pneumoniae PCM 1 | MIC (mM) | 8 | 16 | 4 |
| MBC (mM) | 32 | 32 | 32 | |
| MBC/MIC | 4 | 2 | 8 | |
| E. coli PCM 2057 | MIC (mM) | 16 | 16 | 8 |
| MBC (mM) | 64 | 32 | 32 | |
| MBC/MIC | 4 | 2 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasińska, K.; Zieniuk, B.; Nowak, D.; Fabiszewska, A. Studies on the Catalytic Properties of Crude Freeze-Dried Preparations of Yarrowia lipolytica Extracellular Lipases for Geranyl Ester Derivative Synthesis. Biomolecules 2021, 11, 839. https://doi.org/10.3390/biom11060839
Jasińska K, Zieniuk B, Nowak D, Fabiszewska A. Studies on the Catalytic Properties of Crude Freeze-Dried Preparations of Yarrowia lipolytica Extracellular Lipases for Geranyl Ester Derivative Synthesis. Biomolecules. 2021; 11(6):839. https://doi.org/10.3390/biom11060839
Chicago/Turabian StyleJasińska, Karina, Bartłomiej Zieniuk, Dorota Nowak, and Agata Fabiszewska. 2021. "Studies on the Catalytic Properties of Crude Freeze-Dried Preparations of Yarrowia lipolytica Extracellular Lipases for Geranyl Ester Derivative Synthesis" Biomolecules 11, no. 6: 839. https://doi.org/10.3390/biom11060839