
Restoration of Vitamin D Levels Improves Endothelial Function and Increases TASK-Like K+ Currents in Pulmonary Arterial Hypertension Associated with Vitamin D Deficiency

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
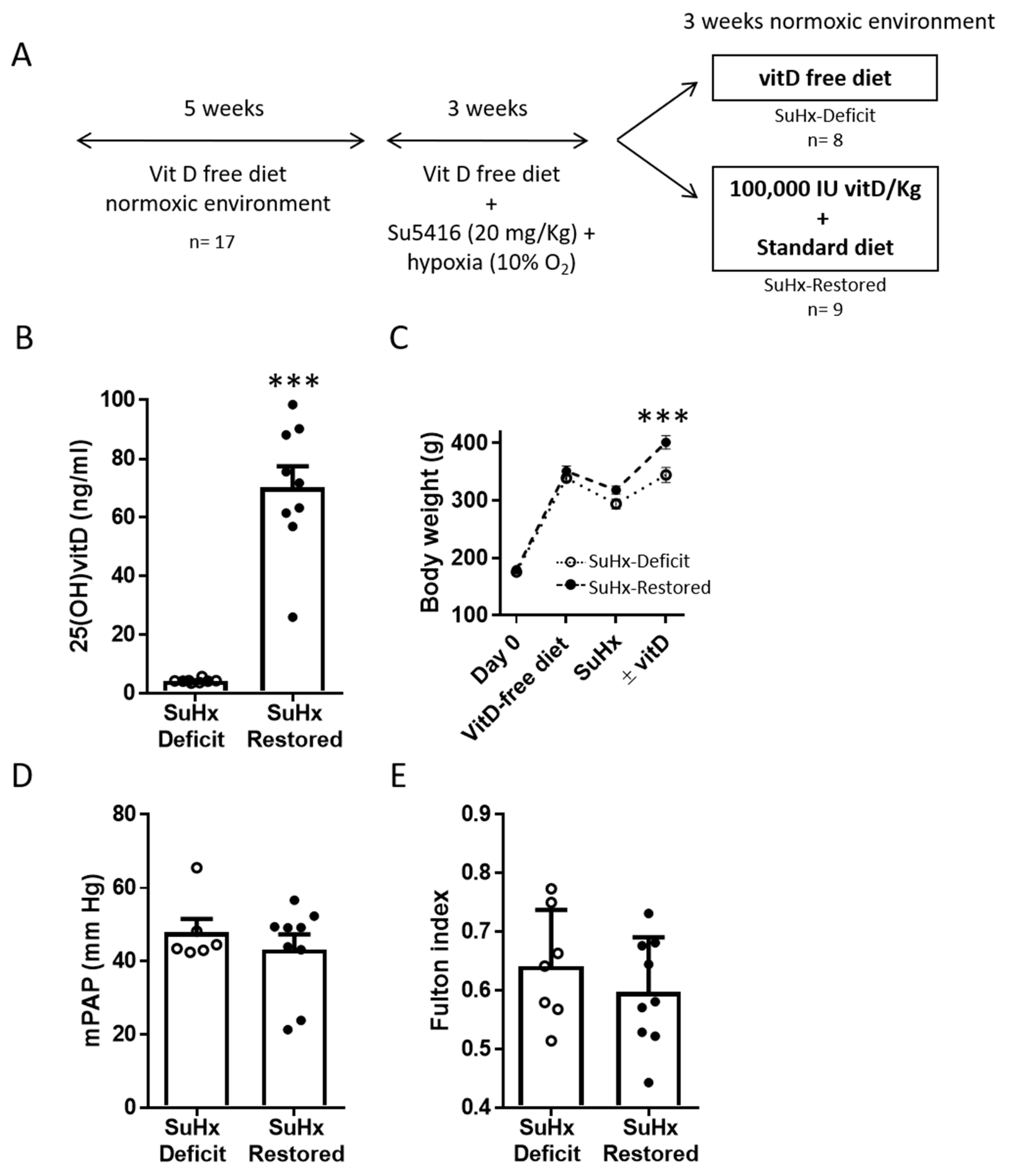
2.1. Vitamin D Replacement in an Animal Model of PAH Associated with Vitamin D Deficiency
2.2. Hemodynamic Measurements
2.3. RV Hypertrophy and Lung Histology
2.4. Electrophysiological Studies
2.5. Arterial Reactivity
2.6. Vitamin D Measurement
2.7. Western Blotting Analysis
2.8. Statistics
3. Results
3.1. Effects of Vitamin D Supplementation on Pulmonary Pressure
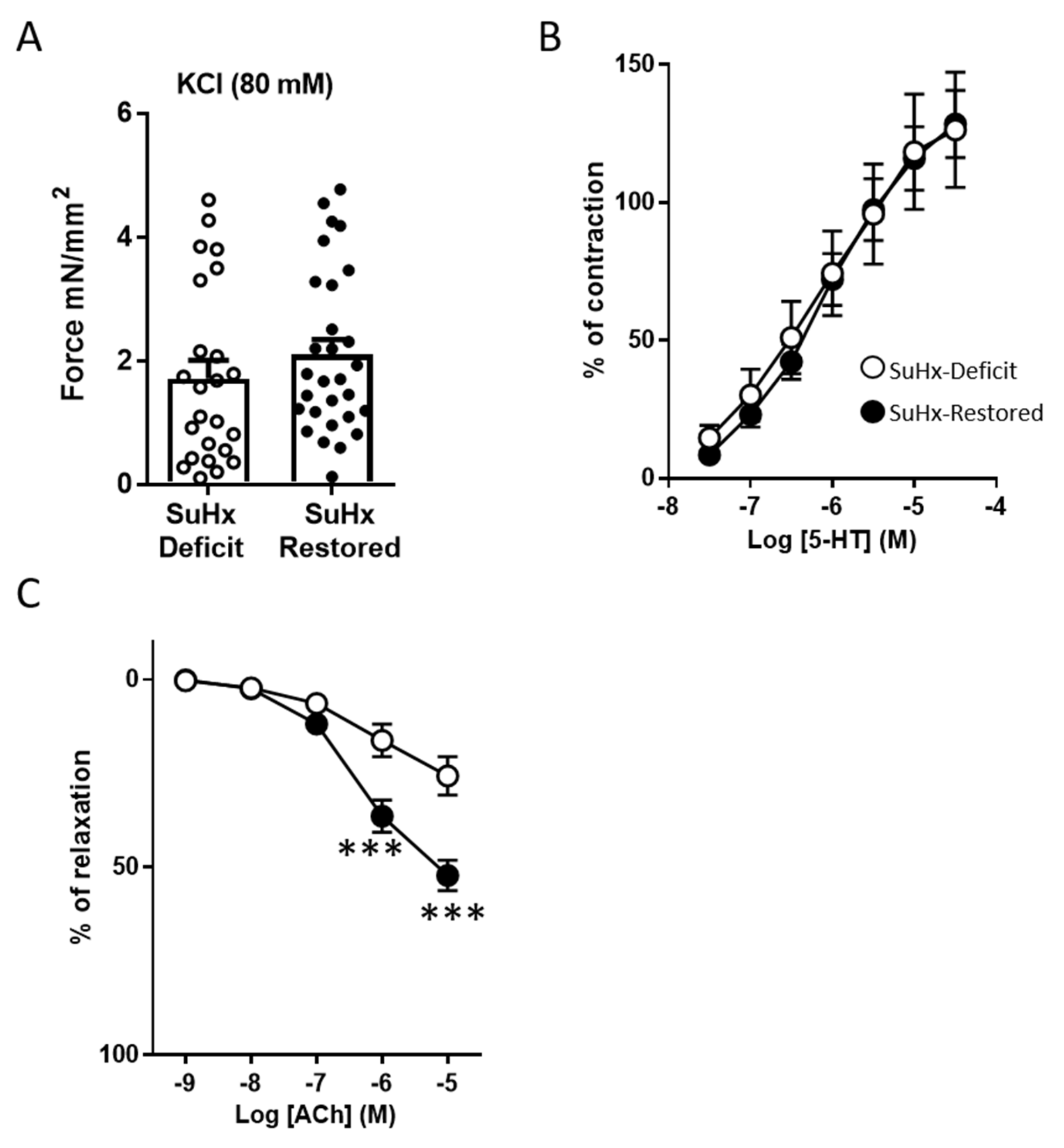
3.2. Effects of Vitamin D Supplementation on Pulmonary Reactivity Endothelial Function
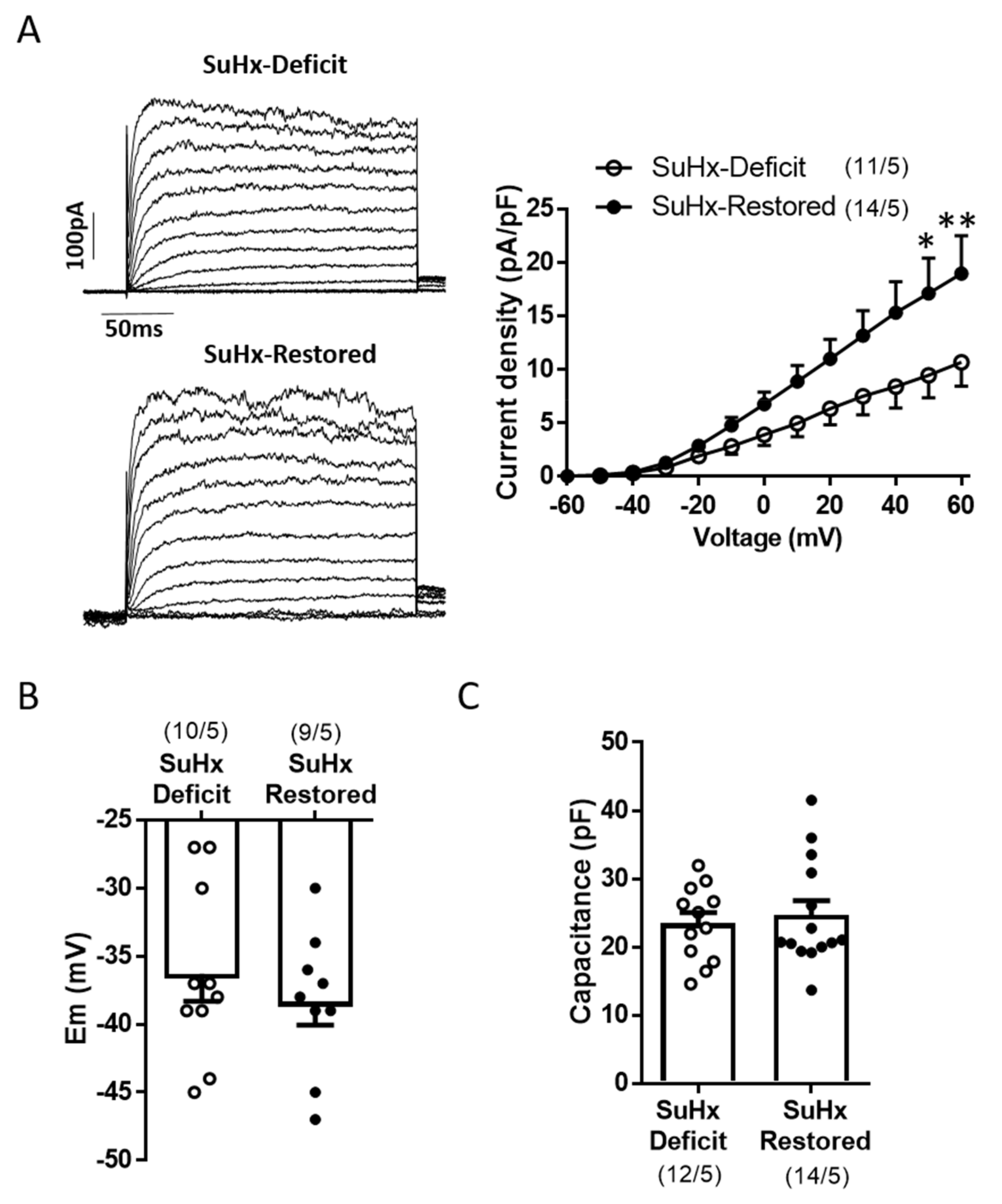
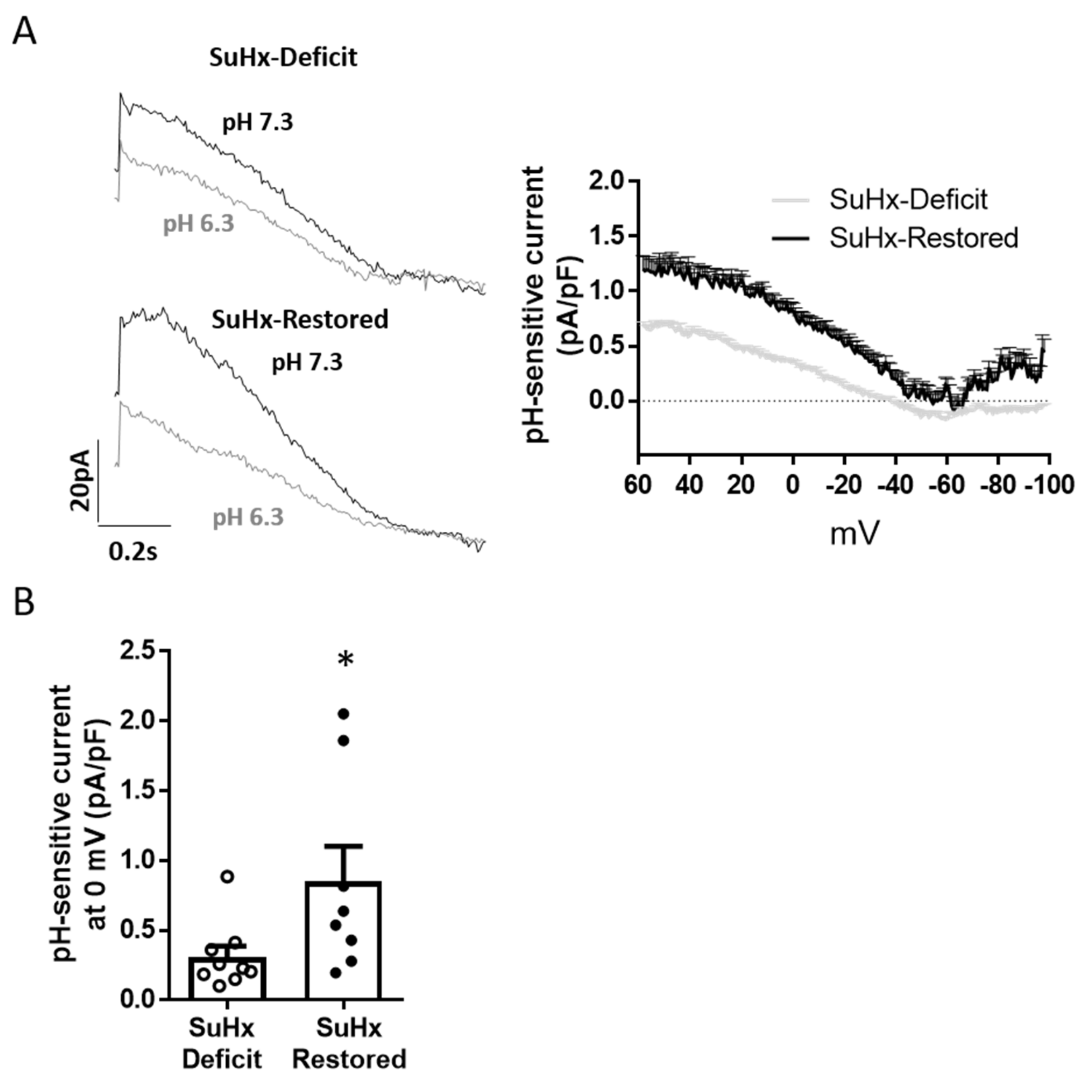
3.3. Restoration of Vitamin D Levels Improved TASK Like-Current
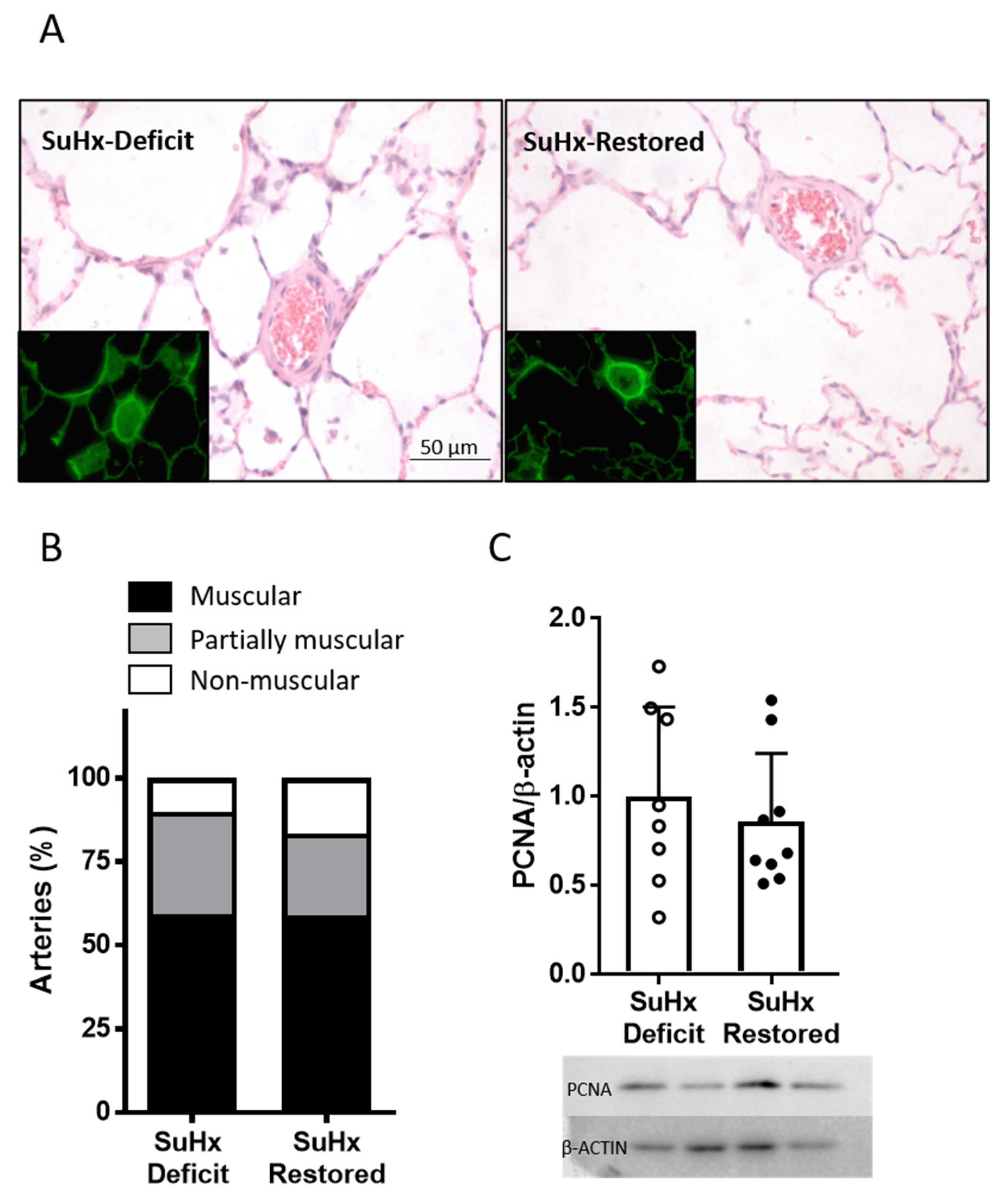
3.4. Effects of Vitamin D Supplementation on Pulmonary Arterial Remodeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M.; et al. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension: The Joint Task Force for the Diagnosis and Treatment of Pulmonary Hypertension of the European Society of Cardiology (ESC) and the European Respiratory Society (ERS): Endorsed by: Association for European Paediatric and Congenital Cardiology (AEPC), International Society for Heart and Lung Transplantation (ISHLT). Eur. Respir. J. 2015, 46, 903–975. [Google Scholar] [CrossRef]
- Thenappan, T.; Ormiston, M.L.; Ryan, J.J.; Archer, S.L. Pulmonary arterial hypertension: Pathogenesis and clinical management. BMJ 2018, 360, j5492. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.; Perros, F.; Tu, L.; Cohen-Kaminsky, S.; Montani, D.; Dorfmuller, P.; Guignabert, C.; Humbert, M. Immune dysregulation and endothelial dysfunction in pulmonary arterial hypertension: A complex interplay. Circulation 2014, 129, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Cogolludo, A.; Moreno, L.; Lodi, F.; Frazziano, G.; Cobeno, L.; Tamargo, J.; Perez-Vizcaino, F. Serotonin inhibits voltage-gated K+ currents in pulmonary artery smooth muscle cells: Role of 5-HT2A receptors, caveolin-1, and KV1.5 channel internalization. Circ. Res. 2006, 98, 931–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondejar-Parreno, G.; Callejo, M.; Barreira, B.; Morales-Cano, D.; Esquivel-Ruiz, S.; Moreno, L.; Cogolludo, A.; Perez-Vizcaino, F. miR-1 is increased in pulmonary hypertension and downregulates Kv1.5 channels in rat pulmonary arteries. J. Physiol. 2019, 597, 1185–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antigny, F.; Hautefort, A.; Meloche, J.; Belacel-Ouari, M.; Manoury, B.; Rucker-Martin, C.; Pechoux, C.; Potus, F.; Nadeau, V.; Tremblay, E.; et al. Potassium Channel Subfamily K Member 3 (KCNK3) Contributes to the Development of Pulmonary Arterial Hypertension. Circulation 2016, 133, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.; Capuano, V.; Olschewski, A.; Sabourin, J.; Nagaraj, C.; Girerd, B.; Weatherald, J.; Humbert, M.; Antigny, F. Ion Channels in Pulmonary Hypertension: A Therapeutic Interest? Int. J. Mol. Sci. 2018, 19, 3162. [Google Scholar] [CrossRef] [Green Version]
- Vinke, P.; Bowen, T.S.; Boekschoten, M.V.; Witkamp, R.F.; Adams, V.; van Norren, K. Anti-inflammatory nutrition with high protein attenuates cardiac and skeletal muscle alterations in a pulmonary arterial hypertension model. Sci. Rep. 2019, 9, 10160. [Google Scholar] [CrossRef] [Green Version]
- Vinke, P.; Jansen, S.M.; Witkamp, R.F.; van Norren, K. Increasing quality of life in pulmonary arterial hypertension: Is there a role for nutrition? Heart Fail Rev. 2018, 23, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Kwant, C.T.; Ruiter, G.; Noordegraaf, A.V. Malnutrition in pulmonary arterial hypertension: A possible role for dietary intervention. Curr. Opin. Pulm. Med. 2019. [Google Scholar] [CrossRef]
- Norman, P.E.; Powell, J.T. Vitamin D and cardiovascular disease. Circ. Res. 2014, 114, 379–393. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D: Newer Concepts of Its Metabolism and Function at the Basic and Clinical Level. J. Endocr. Soc. 2020, 4, bvz038. [Google Scholar] [CrossRef]
- Munns, C.F.; Shaw, N.; Kiely, M.; Specker, B.L.; Thacher, T.D.; Ozono, K.; Michigami, T.; Tiosano, D.; Mughal, M.Z.; Makitie, O.; et al. Global Consensus Recommendations on Prevention and Management of Nutritional Rickets. J. Clin. Endocrinol. Metab. 2016, 101, 394–415. [Google Scholar] [CrossRef]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D’Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Scragg, R. The Vitamin D Assessment (ViDA) study – Design and main findings. J. Steroid Biochem. Mol. Biol. 2020, 198, 105562. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Greenberg, L.; Hooper, R.L.; Griffiths, C.J.; Camargo, C.A., Jr.; Kerley, C.P.; Jensen, M.E.; Mauger, D.; Stelmach, I.; Urashima, M.; et al. Vitamin D supplementation to prevent asthma exacerbations: A systematic review and meta-analysis of individual participant data. Lancet Respir. Med. 2017, 5, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Martineau, A.R.; James, W.Y.; Hooper, R.L.; Barnes, N.C.; Jolliffe, D.A.; Greiller, C.L.; Islam, K.; McLaughlin, D.; Bhowmik, A.; Timms, P.M.; et al. Vitamin D3 supplementation in patients with chronic obstructive pulmonary disease (ViDiCO): A multicentre, double-blind, randomised controlled trial. Lancet Respir. Med. 2015, 3, 120–130. [Google Scholar] [CrossRef]
- Callejo, M.; Mondejar-Parreno, G.; Esquivel-Ruiz, S.; Olivencia, M.A.; Moreno, L.; Blanco, I.; Escribano-Subias, P.; Cogolludo, A.; Barbera, J.A.; Perez-Vizcaino, F. Total, Bioavailable, and Free Vitamin D Levels and Their Prognostic Value in Pulmonary Arterial Hypertension. J. Clin. Med. 2020, 9, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demir, M.; Uyan, U.; Keceoclu, S.; Demir, C. The relationship between vitamin D deficiency and pulmonary hypertension. Prague Med. Rep. 2013, 114, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Mirdamadi, A.; Moshkdar, P. Benefits from the correction of vitamin D deficiency in patients with pulmonary hypertension. Caspian J. Intern. Med. 2016, 7, 253–259. [Google Scholar] [PubMed]
- Tanaka, H.; Kataoka, M.; Isobe, S.; Yamamoto, T.; Shirakawa, K.; Endo, J.; Satoh, T.; Hakamata, Y.; Kobayashi, E.; Sano, M.; et al. Therapeutic impact of dietary vitamin D supplementation for preventing right ventricular remodeling and improving survival in pulmonary hypertension. PLoS ONE 2017, 12, e0180615. [Google Scholar] [CrossRef] [Green Version]
- Atamanuk, A.N.; Litewka, D.F.; Baratta, S.J.; Seropian, I.M.; Perez Prados, G.; Payaslian, M.O.; Ortiz Fragola, J.P.; Escribano Subias, P. Vitamin D deficiency among patients with pulmonary hypertension. BMC Pulm. Med. 2019, 19, 258. [Google Scholar] [CrossRef]
- Ulrich, S.; Hersberger, M.; Fischler, M.; Huber, L.C.; Senn, O.; Treder, U.; Speich, R.; Schmid, C. Bone mineral density and secondary hyperparathyroidism in pulmonary hypertension. Open Respir. Med. J. 2009, 3, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Callejo, M.; Mondejar-Parreno, G.; Morales-Cano, D.; Barreira, B.; Esquivel-Ruiz, S.; Olivencia, M.A.; Manaud, G.; Perros, F.; Duarte, J.; Moreno, L.; et al. Vitamin D deficiency downregulates TASK-1 channels and induces pulmonary vascular dysfunction. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L627–L640. [Google Scholar] [CrossRef]
- Milani, C.; Katayama, M.L.H.; de Lyra, E.C.; Welsh, J.; Campos, L.T.; Brentani, M.M.; Maciel, M.D.; Roela, R.A.; del Valle, P.R.; Goes, J.C.G.S.; et al. Transcriptional effects of 1,25 dihydroxyvitamin D-3 physiological and supra-physiological concentrations in breast cancer organotypic culture. BMC Cancer 2013, 13. [Google Scholar] [CrossRef]
- Shalhoub, V.; Shatzen, E.M.; Ward, S.C.; Young, J.I.; Boedigheimer, M.; Twehues, L.; McNinch, J.; Scully, S.; Twomey, B.; Baker, D.; et al. Chondro/Osteoblastic and Cardiovascular Gene Modulation in Human Artery Smooth Muscle Cells That Calcify in the Presence of Phosphate and Calcitriol or Paricalcitol. J. Cell. Biochem. 2010, 111, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Sempos, C.T.; Vesper, H.W.; Phinney, K.W.; Thienpont, L.M.; Coates, P.M.; Vitamin, D.S.P. Vitamin D status as an international issue: National surveys and the problem of standardization. Scand. J. Clin. Lab. Investig. Suppl. 2012, 243, 32–40. [Google Scholar] [CrossRef]
- Mondejar-Parreno, G.; Barreira, B.; Callejo, M.; Morales-Cano, D.; Barrese, V.; Esquivel-Ruiz, S.; Olivencia, M.A.; Macias, M.; Moreno, L.; Greenwood, I.A.; et al. Uncovered Contribution of Kv7 Channels to Pulmonary Vascular Tone in Pulmonary Arterial Hypertension. Hypertension 2020, 76, 1134–1146. [Google Scholar] [CrossRef]
- Tschopp, O.; Schmid, C.; Speich, R.; Seifert, B.; Russi, E.W.; Boehler, A. Pretransplantation bone disease in patients with primary pulmonary hypertension. Chest 2006, 129, 1002–1008. [Google Scholar] [CrossRef]
- Malik, N.; McCarthy, K.; Minai, O.A. Prevalence and significance of decreased bone density in pulmonary arterial hypertension. South. Med. J. 2012, 105, 344–349. [Google Scholar] [CrossRef]
- Yu, H.; Xu, M.; Dong, Y.; Liu, J.; Li, Y.; Mao, W.; Wang, J.; Wang, L. 1,25(OH)2D3 attenuates pulmonary arterial hypertension via microRNA-204 mediated Tgfbr2/Smad signaling. Exp. Cell Res. 2018, 362, 311–323. [Google Scholar] [CrossRef]
- Binkley, N.; Ramamurthy, R.; Krueger, D. Low vitamin D status: Definition, prevalence, consequences, and correction. Endocrinol. Metab. Clin. N. Am. 2010, 39, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Pilz, S.; Verheyen, N.; Grubler, M.R.; Tomaschitz, A.; Marz, W. Vitamin D and cardiovascular disease prevention. Nat. Rev. Cardiol. 2016, 13, 404–417. [Google Scholar] [CrossRef]
- Ni, W.; Watts, S.W.; Ng, M.; Chen, S.; Glenn, D.J.; Gardner, D.G. Elimination of vitamin D receptor in vascular endothelial cells alters vascular function. Hypertension 2014, 64, 1290–1298. [Google Scholar] [CrossRef] [Green Version]
- Borgi, L.; McMullan, C.; Wohlhueter, A.; Curhan, G.C.; Fisher, N.D.; Forman, J.P. Effect of Vitamin D on Endothelial Function: A Randomized, Double-Blind, Placebo-Controlled Trial. Am. J. Hypertens. 2017, 30, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Beveridge, L.A.; Khan, F.; Struthers, A.D.; Armitage, J.; Barchetta, I.; Bressendorff, I.; Cavallo, M.G.; Clarke, R.; Dalan, R.; Dreyer, G.; et al. Effect of Vitamin D Supplementation on Markers of Vascular Function: A Systematic Review and Individual Participant Meta-Analysis. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Szczurek, M.; Hassan, C.; Masrur, M.; Gangemi, A.; Phillips, S.A. Vitamin D Improves Nitric Oxide-Dependent Vasodilation in Adipose Tissue Arterioles from Bariatric Surgery Patients. Nutrients 2019, 11, 2521. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, F.; Yamamoto, K.; Matsuda, K.; Kawata, K.; Negishi, M.; Shinomiya, K.; Shimamur, K.; Sunano, S. Endothelium-dependent relaxation in pulmonary arteries of L-NAME-treated Wistar and stroke-prone spontaneously hypertensive rats. J. Smooth Muscle Res. 2002, 38, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Mondejar-Parreno, G.; Cogolludo, A.; Perez-Vizcaino, F. Potassium (K(+)) channels in the pulmonary vasculature: Implications in pulmonary hypertension Physiological, pathophysiological and pharmacological regulation. Pharmacol. Ther. 2021, 225, 107835. [Google Scholar] [CrossRef]
- Boucherat, O.; Chabot, S.; Antigny, F.; Perros, F.; Provencher, S.; Bonnet, S. Potassium channels in pulmonary arterial hypertension. Eur. Respir. J. 2015, 46, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Le Ribeuz, H.; Dumont, F.; Ruellou, G.; Lambert, M.; Balliau, T.; Quatredeniers, M.; Girerd, B.; Cohen-Kaminsky, S.; Mercier, O.; Yen-Nicolay, S.; et al. Proteomic Analysis of KCNK3 Loss of Expression Identified Dysregulated Pathways in Pulmonary Vascular Cells. Int. J. Mol. Sci. 2020, 21, 7400. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.; Mendes-Ferreira, P.; Ghigna, M.R.; LeRibeuz, H.; Adao, R.; Boet, A.; Capuano, V.; Rucker-Martin, C.; Bras-Silva, C.; Quarck, R.; et al. Kcnk3 Dysfunction Exaggerates The Development Of Pulmonary Hypertension Induced By Left Ventricular Pressure Overload. Cardiovasc. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, K.P.; Holden, R.G.; Escribano-Subias, P.M.; Cogolludo, A.; Veale, E.L.; Mathie, A. Characterization and regulation of wild-type and mutant TASK-1 two pore domain potassium channels indicated in pulmonary arterial hypertension. J. Physiol. 2019, 597, 1087–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olschewski, A.; Li, Y.; Tang, B.; Hanze, J.; Eul, B.; Bohle, R.M.; Wilhelm, J.; Morty, R.E.; Brau, M.E.; Weir, E.K.; et al. Impact of TASK-1 in human pulmonary artery smooth muscle cells. Circ. Res. 2006, 98, 1072–1080. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.; Capuano, V.; Boet, A.; Tesson, L.; Bertero, T.; Nakhleh, M.K.; Remy, S.; Anegon, I.; Pechoux, C.; Hautefort, A.; et al. Characterization of Kcnk3-Mutated Rat, a Novel Model of Pulmonary Hypertension. Circ. Res. 2019, 125, 678–695. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SuHx-Deficit | SuHx-Restored | Student’s’ t Test | |
|---|---|---|---|
| n | 6 * | 9 | |
| sPAP (mmHg) | 81.19 ± 7.75 | 72.38 ± 9.54 | 0.48 |
| dPAP (mmHg) | 29.83 ± 1.29 | 25.45 ± 2.67 | 0.18 |
| mPAP (mmHg) | 47.86 ± 3.63 | 43.21 ± 4.67 | 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callejo, M.; Morales-Cano, D.; Mondejar-Parreño, G.; Barreira, B.; Esquivel-Ruiz, S.; Olivencia, M.A.; Moreno, L.; Cogolludo, A.; Perez-Vizcaino, F. Restoration of Vitamin D Levels Improves Endothelial Function and Increases TASK-Like K+ Currents in Pulmonary Arterial Hypertension Associated with Vitamin D Deficiency. Biomolecules 2021, 11, 795. https://doi.org/10.3390/biom11060795
Callejo M, Morales-Cano D, Mondejar-Parreño G, Barreira B, Esquivel-Ruiz S, Olivencia MA, Moreno L, Cogolludo A, Perez-Vizcaino F. Restoration of Vitamin D Levels Improves Endothelial Function and Increases TASK-Like K+ Currents in Pulmonary Arterial Hypertension Associated with Vitamin D Deficiency. Biomolecules. 2021; 11(6):795. https://doi.org/10.3390/biom11060795
Chicago/Turabian StyleCallejo, Maria, Daniel Morales-Cano, Gema Mondejar-Parreño, Bianca Barreira, Sergio Esquivel-Ruiz, Miguel Angel Olivencia, Laura Moreno, Angel Cogolludo, and Francisco Perez-Vizcaino. 2021. "Restoration of Vitamin D Levels Improves Endothelial Function and Increases TASK-Like K+ Currents in Pulmonary Arterial Hypertension Associated with Vitamin D Deficiency" Biomolecules 11, no. 6: 795. https://doi.org/10.3390/biom11060795